
Identification of Signatures of Positive Selection That Have Shaped the Genomic Landscape of South African Pig Populations

 , ,
, ,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animal Samples, Genotyping, and Quality Control
2.2. Detection of Signatures Using iHS
2.3. Detection of Signatures Using XP-EHH
2.4. Detection of Signatures Using HapFLK
2.5. Annotation and Function Analyses of Identified Genomic Regions
3. Results
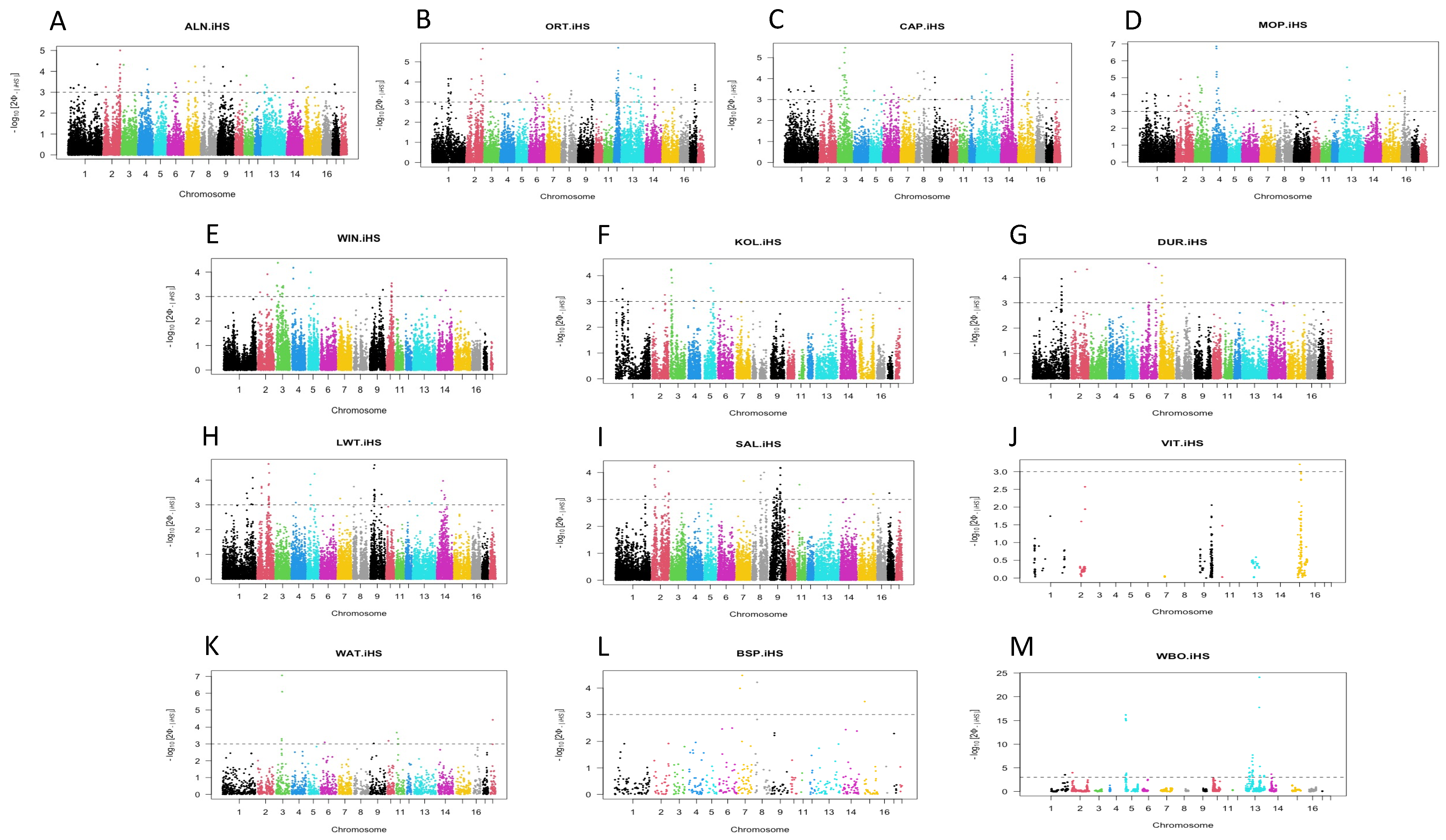
3.1. Detection of Signatures within a Population Using iHS
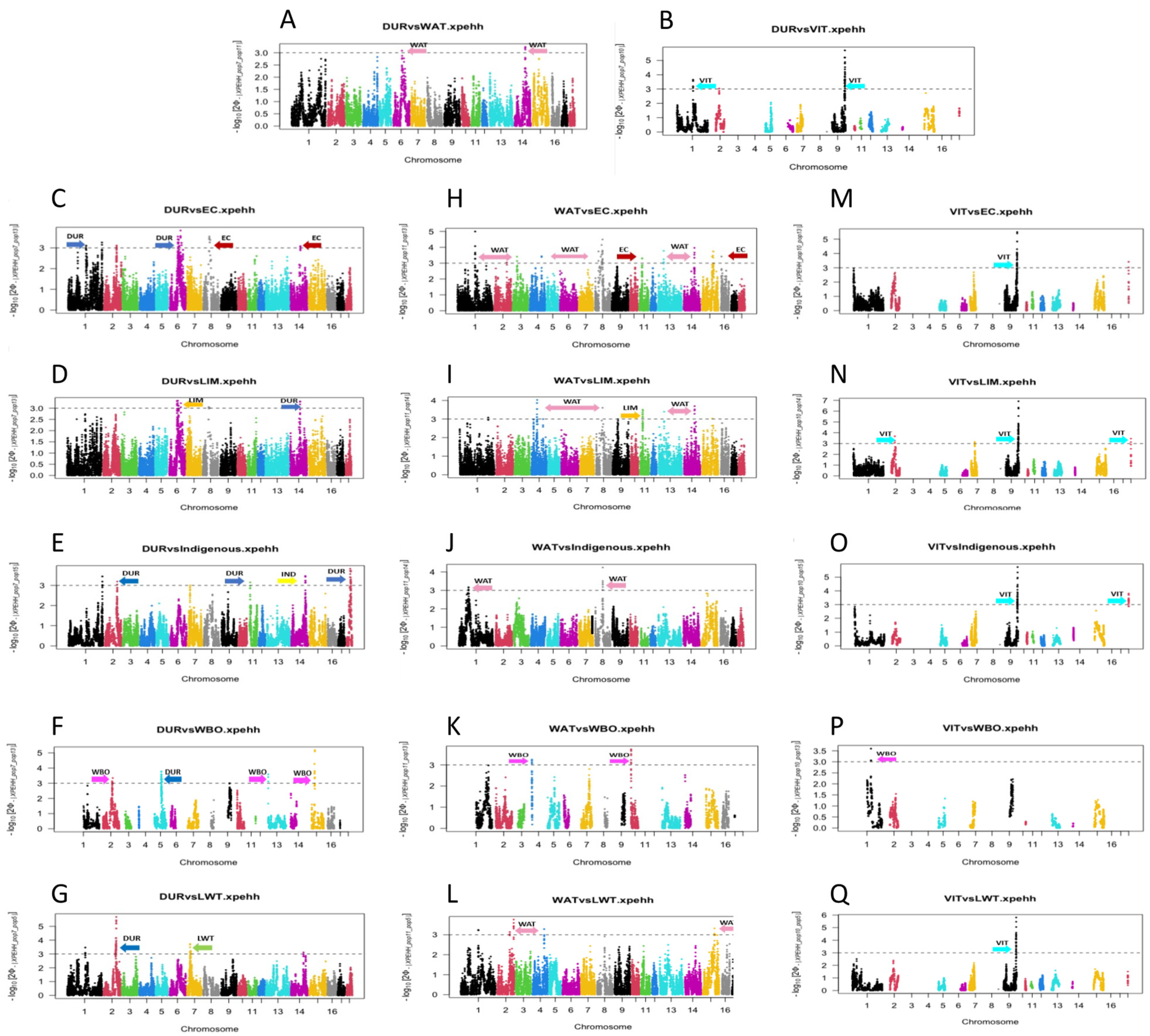
3.2. Detection of Selection of Signatures between Populations Using XP-EHH
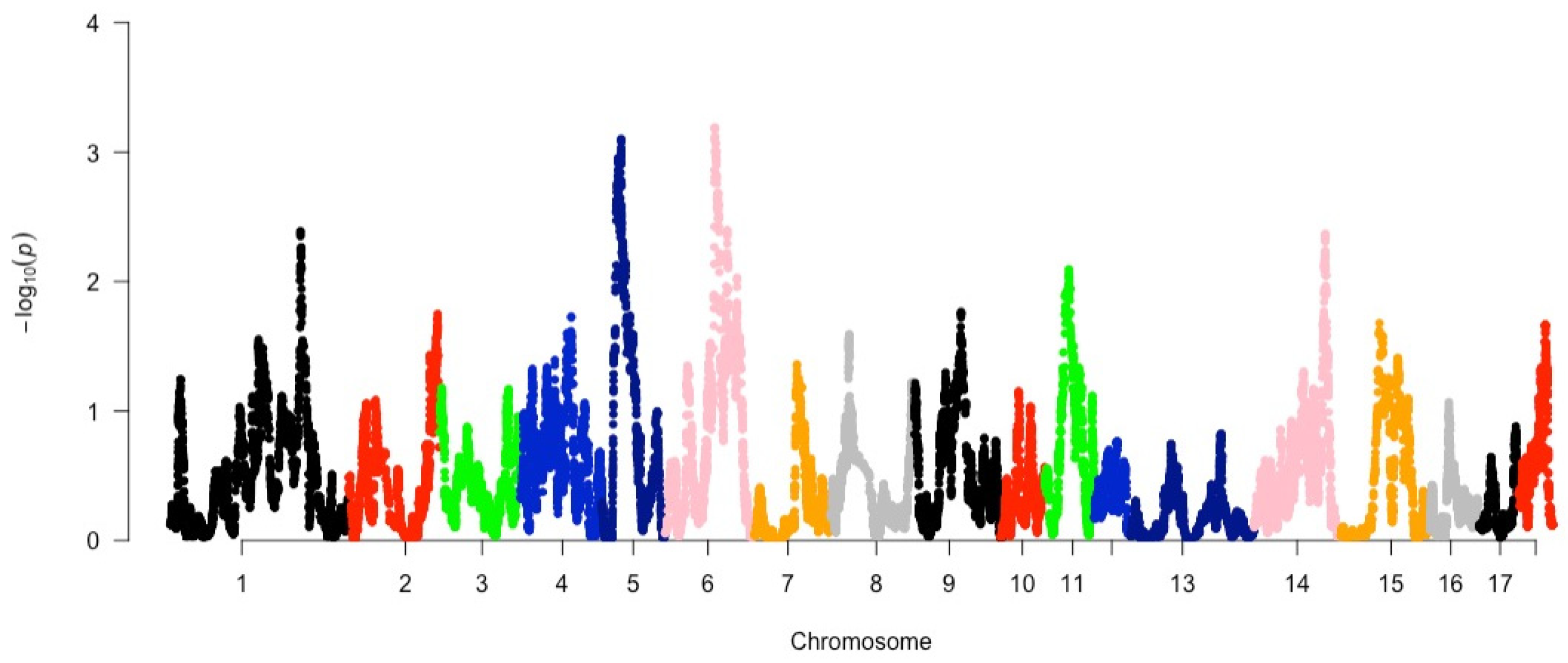
3.3. Detection of Selection of Signatures between Populations Using HapFLK
3.4. Genes Identified Using Different Signatures of Selection Methods
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Statista. Global Number of Pigs 2012–2021, by Country. Available online: https://www.statista.com/statistics/263963/number-of-pigs-worldwide-since-1990/ (accessed on 18 August 2021).
- Lunney, J.K. Advances in swine biomedical model genomics. Int. J. Biol. Sci. 2007, 3, 179–184. [Google Scholar] [CrossRef]
- Giuffra, E.; Kijas, J.M.; Amarger, V.; Carlborg, O.; Jeon, J.T.; Andersson, L. The origin of the domestic pig: Independent domestication and subsequent introgression. Genetics 2000, 154, 1785–1791. [Google Scholar] [CrossRef]
- Li, Y.; Mei, S.; Zhang, X.; Peng, X.; Liu, G.; Tao, H.; Wu, H.; Jiang, S.; Xiong, Y.; Li, F. Identification of genome-wide copy number variations among diverse pig breeds by array CGH. BMC Genom. 2012, 13, 725. [Google Scholar] [CrossRef]
- Gurgul, A.; Jasielczuk, I.; Ropka-Molik, K.; Semik-Gurgul, E.; Pawlina-Tyszko, K.; Szmatoła, T.; Szyndler-Nędza, M.; Bugno-Poniewierska, M.; Blicharski, T.; Szulc, K.; et al. A genome-wide detection of selection signatures in conserved and commercial pig breeds maintained in Poland. BMC Genet. 2018, 19, 95. [Google Scholar] [CrossRef] [PubMed]
- Larson, G.; Dobney, K.; Albarella, U.; Fang, M.; Matisoo-Smith, E.; Robins, J.; Lowden, S.; Finlayson, H.; Brand, T.; Willerslev, E.; et al. Worldwide phylogeography of wild boar reveals multiple centers of pig domestication. Science 2005, 307, 1618–1621. [Google Scholar] [CrossRef] [PubMed]
- Wu, G.S.; Yao, Y.G.; Qu, K.X.; Ding, Z.L.; Li, H.; Palanichamy, M.G.; Duan, Z.Y.; Li, N.; Chen, Y.S.; Zhang, Y.P. Population phylogenomic analysis of mitochondrial DNA in wild boars and domestic pigs revealed multiple domestication events in East Asia. Genome Biol. 2007, 8, R245. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Rodríguez, F.; Navas, M.J.; Mar, C.H.; Almagro-Delgado, V.; López, E.; Cuadrado, R.; Bosch-Camós, L.; Accensi Alemany, F.; Pina-Pedrero, S.; et al. Fecal microbiota transplantation from warthog to pig confirms the influence of the gut microbiota on African Swine Fever susceptibility. Sci. Rep. 2020, 10, 17605. [Google Scholar] [CrossRef]
- van Laere, A.S.; Nguyen, M.; Braunschweig, M.; Nezer, C.; Collette, C.; Moreau, L.; Archibald, A.L.; Haley, C.S.; Buys, N.; Tally, M.; et al. A regulatory mutation in IGF2 causes a major QTL effect on muscle growth in the pig. Nature 2003, 425, 832–836. [Google Scholar] [CrossRef]
- Rubin, C.J.; Megens, H.J.; Barrio, A.M.; Maqbool, K.; Sayyab, S.; Schwochow, D.; Wang, C.; Carlborg, O.; Jern, P.; Jorgensen, C.B.; et al. Strong signatures of selection in the domestic pig genome. Proc. Natl. Acad. Sci. USA 2012, 109, 19529–19536. [Google Scholar] [CrossRef]
- Zhang, Z.G. Pig Breeds in China; Shanghai Scientific and Technical Publishers: Shanghai, China, 1986; pp. 73–277. [Google Scholar]
- Guo, Y.; Mao, H.; Ren, J.; Yan, X.; Duan, Y.; Yang, G.; Ren, D.; Zhang, Z.; Yang, B.; Ouyang, J.; et al. A linkage map of the porcine genome from a large-scale White Duroc X Erhualian resource population and evaluation of factors affecting recombination rates. Anim. Genet. 2009, 40, 47–52. [Google Scholar] [CrossRef]
- Li, K.; Ren, J.; Xing, Y.; Zhang, Z.; Ma, J.; Guo, Y.; Huang, L. Quantitative trait loci for litter size and prenatal loss in a White Duroc X Chinese Erhualian resource population. Anim. Genet. 2009, 40, 963–966. [Google Scholar] [CrossRef]
- Watanabe, T.; Hayashi, Y.; Kimura, J.; Yasuda, Y.; Saitou, N.; Tomita, T.; Ogasawara, N. Pig mitochondrial DNA: Polymorphism, restriction map orientation, and sequence data. Biochem. Genet. 1986, 24, 385–396. [Google Scholar] [CrossRef]
- Scandura, M.; Iacolina, L.; Apollonio, M. Genetic diversity in the European wild boar Sus scrofa: Phylogeography, population structure and wild domestic hybridization. Mammal Rev. 2011, 41, 125–137. [Google Scholar] [CrossRef]
- Nyafu, K. Warthogs as an Introduced Species in the Eastern Cape. Master’s Thesis, Nelson Mandela Metropolitan University, Port Elizabeth, South Africa, 2009. [Google Scholar]
- Amar, S.; De Boni, L.; de Voux, A.; Heath, L.; Geertsma, P. An outbreak of African Swine Fever in small-scale pigs, Gauteng, South Africa, July 2020. Int. J. Infect. Dis. 2021, 110, S44–S49. [Google Scholar] [CrossRef]
- Molotsi, A.H.; Cupido, M.; Hoffman, L.C.H. Characterization of smallholder pig production systems in the Western Cape, South Africa. Trop. Anim. Health Prod. 2021, 53, 325. [Google Scholar] [CrossRef]
- NAFU. NAFU Farmer Headlines; June Issue Newsletter, 18–20 November 2007, Issue 9; NAFU: Pretoria, South Africa, 2007. [Google Scholar]
- Penrith, M.L.; Vosloo, W.; Mather, C. Classical Swine Fever (Hog Cholera): Review of aspects relevant to control. Transbound. Emerg. Dis. 2011, 58, 187–196. [Google Scholar] [CrossRef]
- Krige, J.E. The Social Systems of the Zulus; Shuter and Shooter: Pietermaritzburg, South Africa, 1988. [Google Scholar]
- Blench, R.M.; MacDonald, K.C. The Origins and Development of African Livestock: Archaeology, Genetics, Linguistics and Ethnography; UCL Press: London, UK, 2000. [Google Scholar]
- Swart, H.; Kotze, A.; Olivier, P.A.S.; Grobler, J.P. Microsatellite-based characterization of Southern African domestic pigs (Sus scrofa Domestica). S. Afr. J. Anim. Sci. 2010, 40, 121–132. [Google Scholar] [CrossRef]
- Visser, D.P. Structuring of Breeding Objectives in the Pork Supply Chain in South Africa. Ph.D. Thesis, University of Pretoria, Pretoria, South Africa, 2004. [Google Scholar]
- Visser, D. Modern Pig Production. Available online: https://books.google.co.za/books?id=mpXorQEACAAJ (accessed on 14 October 2016).
- Munzhelele, P. Evaluation of the Production Systems and Constraints of Smallholder Pig Farming in Three Agro-Ecological Zones of Mpumalanga Province, South Africa. Master’s Thesis, University of South Africa, Pretoria, South Africa, 2015. [Google Scholar]
- Madzimure, J.; Chimonyo, M.; Zander, K.K.; Dzama, K. Diurnal heat-related physiological and behavioural responses in South African indigenous gilts. J. Arid Environ. 2012, 87, 29–34. [Google Scholar] [CrossRef]
- Moyo, D.; Gomes, M.; Erlwanger, K.H. Comparison of the histology of the skin of the Windsnyer, Kolbroek and Large White pigs. J. S. Afr. Vet. Assoc. 2018, 89, 1–10. [Google Scholar] [CrossRef]
- Meissner, H.H.; Scholtz, M.M.; Palmer, A.R. Sustainability of the South African livestock sector towards 2050 Part 1: Worth and impact of the sector. S. Afr. J. Anim. Sci. 2013, 43, 282–297. [Google Scholar] [CrossRef]
- Madzimure, J.; Chimonyo, M.; Zander, K.K.; Dzama, K. Potential for using indigenous pigs in subsistence-oriented and market-oriented small-scale farming systems of Africa. Trop. Anim. Health Prod. 2012, 45, 135–142. [Google Scholar] [CrossRef] [PubMed]
- Halimani, T.E.; Muchadeyi, F.C.; Chimonyo, M.; Dzama, K. Pig genetic resource conservation: The Southern African perspective. Ecol. Econ. 2010, 69, 944–951. [Google Scholar] [CrossRef]
- Esfandyari, H.; Sørensen, A.C.; Bijma, P. Maximizing crossbred performance through purebred genomic selection. Genet. Sel. Evol. 2015, 47, 16. [Google Scholar] [CrossRef]
- Mohlatlole, R.P.; Madoroba, E.; Muchadeyi, F.C.; Chimonyo, M.; Kanengoni, A.T.; Dzomba, E.F. Virulence profiles of enterotoxigenic, shiga toxin and enteroaggregative Escherichia coli in South African pigs. Trop. Anim. Health Prod. 2013, 45, 399–405. [Google Scholar] [CrossRef]
- Raudsepp, T.; Finno, C.J.; Bellone, R.R.; Petersen, J.L. Ten years of the Horse Reference Genome: Insights into equine biology, domestication and population dynamics in the post genome era. Anim. Genet. 2019, 50, 569–597. [Google Scholar] [CrossRef] [PubMed]
- Ai, H.; Huang, L.; Ren, J. Genetic diversity, linkage disequilibrium and selection signatures in Chinese and Western pigs revealed by genome-wide SNP markers. PLoS ONE 2013, 8, e56001. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Chen, Q.; Yang, Y.; Yang, H.; He, P.; Zhang, Z.; Chen, Z.; Liao, R.; Tu, Y.; Zhang, X.; et al. A genome-wide scan for selection signatures in Yorkshire and Landrace pigs based on sequencing data. Anim. Genet. 2014, 45, 808–816. [Google Scholar] [CrossRef]
- D’Alessandro, E.; Arfuso, F.; Floridia, V.; Tardiolo, G.; Fazio, F.; Giannetto, C.; Piccione, G.; Zumbo, A. Different genotype and a liquid whey-supplemented diet influence the resilience of pigs through immune-modulation and anti-inflammatory response. Front. Vet. Sci. 2022, 9, 1046101. [Google Scholar] [CrossRef]
- Sutera, A.M.; Arfuso, F.; Tardiolo, G.; Riggio, V.; Fazio, F.; Aiese Cigliano, R.; Paytuví, A.; Piccione, G.; Zumbo, A. Effect of a co-feed liquid whey-integrated diet on crossbred pigs’ fecal microbiota. Animals 2023, 13, 1750. [Google Scholar] [CrossRef]
- Floridia, V.; Giuffrè, L.; Giosa, D.; Arfuso, F.; Aragona, F.; Fazio, F.; Chen, C.; Song, C.; Romeo, O.; D’Alessandro, E. Comparison of the faecal microbiota composition following a dairy by-product supplemented diet in nero siciliano and large white × landrace pig breeds. Animals 2023, 13, 2323. [Google Scholar] [CrossRef]
- Hlongwane, N.L.; Hadebe, K.; Soma, P.; Dzomba, E.F.; Muchadeyi, F.C. Genome wide assessment of genetic variation and population distinctiveness of the pig family in South Africa. Front. Genet. 2020, 11, 344. [Google Scholar] [CrossRef] [PubMed]
- Voight, B.F.; Kudaravalli, S.; Wen, X.; Pritchard, J.K. A map of recent positive selection in the human genome. PLoS Biol. 2006, 4, e72. [Google Scholar]
- Sabeti, P.C.; Varilly, P.; Fry, B.; Lohmueller, J.; Hostetter, E.; Cotsapas, C.; Xie, X.; Byrne, E.H.; McCarroll, S.A.; Gaudet, R.; et al. Genome-wide detection and characterization of positive selection in human populations. Nature 2007, 449, 913–918. [Google Scholar] [CrossRef] [PubMed]
- Sabeti, P.C.; Reich, D.E.; Higgins, J.M.; Levine, H.Z.; Richter, D.J.; Schaffner, S.F.; Gabriel, S.B.; Platko, J.V.; Patterson, N.J.; McDonald, G.J.; et al. Detecting recent positive selection in the human genome from haplotype structure. Nature 2002, 419, 832–837. [Google Scholar] [CrossRef] [PubMed]
- Eydivandi, S.; Roudbar, M.A.; Ardestani, S.S.; Momen, M.; Sahana, G. A selection signatures study among Middle Eastern and European sheep breeds. J. Anim. Breed. Genet. 2021, 138, 574–588. [Google Scholar] [CrossRef]
- Chen, Z.; Zhu, M.; Wu, Q.; Lu, H.; Lei, C.; Ahmed, Z.; Sun, J. Analysis of genetic diversity and selection characteristics using the whole genome sequencing data of five buffaloes, including Xilin buffalo, in Guangxi, China. Front. Genet. 2023, 13, 1084824. [Google Scholar] [CrossRef] [PubMed]
- Fariello, M.I.; Boitard, S.; Naya, H.; SanCristobal, M.; Servin, B. Detecting signatures of selection through haplotype differentiation among hierarchically structured populations. Genetics 2013, 193, 929–941. [Google Scholar] [CrossRef]
- Zhao, F.; Deng, T.; Shi, L.; Wang, W.; Zhang, Q.; Du, L.; Wang, L. Genomic scan for selection signature reveals fat deposition in Chinese indigenous sheep with extreme tail types. Animals 2020, 10, 773. [Google Scholar] [CrossRef]
- Storey, J.D.; Tibshirani, R. Statistical Significance for Genomewide Studies. Proc. Natl. Acad. Sci. USA 2003, 100, 9440–9445. [Google Scholar] [CrossRef]
- Willing, E.; Dreyer, C.; Oosterhout, C. Estimates of genetic differentiation measured by fst do not necessarily require large sample sizes when using many SNP markers. PLoS ONE 2012, 7, e42649. [Google Scholar] [CrossRef]
- Devani, K.; Crowley, J.J.; Plastow, G.; Orsel, K.; Valente, T.S. Genetic parameter estimations and genomic insights for teat and udder structure in young and mature Canadian Angus cows. J. Anim. Sci. 2021, 99, skab087. [Google Scholar] [CrossRef] [PubMed]
- Abraham, G. The importance of muscarinic receptors in domestic animal diseases and therapy: Current and future perspectives. Vet. J. 2016, 208, 13–21. [Google Scholar] [CrossRef] [PubMed]
- Tyra, M.; Ropka-Molik, K.; Piórkowska, K.; Oczkowicz, M.; Szyndler-Nędza, M.; Małopolska, M. Association of ghrelin gene polymorphisms with fattening traits and feed intake in pig: A preliminary study. Animals 2019, 9, 410. [Google Scholar] [CrossRef] [PubMed]
- El-Magd, M.A.; Saleh, A.A.; Abdel-Hamid, T.M.; Saleh, R.M.; Afifi, M.A. Is really endogenous ghrelin a hunger signal in chickens? Association of GHSR SNPs with increase appetite, growth traits, expression and serum level of GHRL, and GH. Gen. Comp. Endocrinol. 2016, 237, 131–139. [Google Scholar] [CrossRef] [PubMed]
- Duarte, D.A.S.; Fortes, M.R.S.; de Souza Duarte, M.; Guimarães, S.E.; Verardo, L.L.; Veroneze, R.; Ribeiro, A.M.F.; Lopes, P.S.; de Resende, M.D.V.; e Silva, F.F. Genome-wide association studies, meta-analyses and derived gene network for meat quality and carcass traits in pigs. Anim. Prod. Sci. 2017, 58, 1100–1108. [Google Scholar] [CrossRef]
- Qin, M.; Li, C.; Li, Z.; Chen, W.; Zeng, Y. Genetic diversities and differentially selected regions between Shandong indigenous pig breeds and Western pig breeds. Front. Genet. 2020, 10, 1351. [Google Scholar] [CrossRef] [PubMed]
- Hoffman, L.C.; Styger, E.; Muller, M.; Brand, T.S. Sensory, Physical and chemical quality characteristics of bacon derived from South African indigenous and commercial pig breeds. S. Afr. Soc. Anim. Sci. 2005, 6, 38–48. [Google Scholar]
- Kogelman, L.J.; Kadarmideen, H.N.; Mark, T.; Karlskov-Mortensen, P.; Bruun, C.S.; Cirera, S.; Jacobsen, M.J.; Jørgensen, C.B.; Fredholm, M. An F2 pig resource population as a model for genetic studies of obesity and obesity-related diseases in humans: Design and genetic parameters. Front. Genet. 2013, 4, 29. [Google Scholar] [CrossRef]
- Reardon, W.; Mullen, A.M.; Sweeney, T.; Hamill, R.M. Association of polymorphisms in candidate genes with colour, water-holding capacity, and composition traits in bovine M. longissimus and M. semimembranosus. Meat Sci. 2010, 86, 270–275. [Google Scholar] [CrossRef]
- Jung, J.H.; Shim, K.S.; Na, C.S.; Choe, H.S. Studies on intramuscular fat percentage in live swine using real-time ultrasound to determine pork quality. Asian Australas. J. Anim. Sci. 2015, 28, 318–322. [Google Scholar] [CrossRef]
- Ren, Z.; Wang, Y.; Ren, Y.; Zhang, Z.; Gu, W.; Wu, Z.; Chen, L.; Mou, L.; Li, R.; Yang, H.; et al. Enhancement of porcine intramuscular fat content by overexpression of the cytosolic form of phosphoenolpyruvate carboxykinase in skeletal muscle. Sci. Rep. 2017, 7, 43746. [Google Scholar] [CrossRef] [PubMed]
- Switonski, M.; Stachowiak, M.; Cieslak, J.; Bartz, M.; Grzes, M. Genetics of fat tissue accumulation in pigs: A comparative approach. J. Appl. Genet. 2010, 51, 153–168. [Google Scholar] [CrossRef]
- Stachowiak, M.; Szczerbal, I.; Switonski, M. Genetics of adiposity in large animal models for human obesity-studies on pigs and dogs. Prog. Mol. Biol. Transl. Sci. 2016, 140, 233–270. [Google Scholar]
- Lopes-Bote, C.J. Sustained utilization of the Iberian pig breed. Meat Sci. 1998, 49, S17–S27. [Google Scholar] [CrossRef]
- Egerszegi, I.; Rátky, J.; Solti, L.; Brüssow, K.P. Mangalica—An indigenous swine breed from Hungary. Arch. Anim. Breed. 2003, 46, 245–256. [Google Scholar] [CrossRef]
- Hoffman, L.C.; Styger, W.F.; Brand, T.S.; Muller, M. The growth, carcass yield, physical and chemical characteristic of two South African indigenous pig breeds. S. Afr. J. Anim. Sci. 2005, 6, 25–35. [Google Scholar]
- Ludwiczak, A.; Składanowska-Baryza, J.; Stanisz, M. Effect of age and sex on the quality of offal and meat of the wild boar (Sus scrofa). Animals 2020, 10, 660. [Google Scholar] [CrossRef] [PubMed]
- Niewiadomska, K.; Kosicka-Gębska, M.; Gębski, J.; Gutkowska, K.; Jeżewska-Zychowicz, M.; Sułek, M. Game meat consumption—Conscious choice or just a game? Foods 2020, 9, 1357. [Google Scholar] [CrossRef]
- Ciobanu, M.M.; Postolache, A.N.; Lipşa, F.D.; Munteanu, M.; Rațu, R.N.; Murariu, O.C.; Boișteanu, P.C. Meat fatty acid composition of wild boars hunted in Romania in relationship to gender and age-class. Animals 2022, 12, 810. [Google Scholar] [CrossRef]
- Li, D.; Huang, M.; Zhuang, Z.; Ding, R.; Gu, T.; Hong, L.; Zheng, E.; Li, Z.; Cai, G.; Wu, Z.; et al. Genomic analyses revealed the genetic difference and potential selection genes of growth traits in two Duroc lines. Front. Vet. Sci. 2021, 8, 725367. [Google Scholar] [CrossRef]
- Chen, Z.; Li, Y.; Zhang, Z.; Zhao, W.; Zhang, Z.; Xiang, Y.; Wang, Q.; Pan, Y.; Guo, X.; Wang, Z. Genome-wide epistatic interactions of litter size at birth in Chinese indigenous pigs. Anim. Genet. 2021, 52, 739–743. [Google Scholar] [CrossRef]
- Miller, L.C.; Fleming, D.S.; Lager, K.M. Comparison of the transcriptome response within the swine tracheobronchial lymphnode following infection with PRRSV, PCV-2 or IAV-S. Pathogens 2020, 9, 99. [Google Scholar] [CrossRef]
- Mészárosová, M.; Mészáros, G.; Moravčíková, N.; Pavlík, I.; Margetín, M.; Kasarda, R. Within-and between-breed selection signatures in the original and improved Valachian sheep. Animals 2022, 12, 1346. [Google Scholar] [CrossRef]
- Krupa, E.; Wolfová, M.; Krupová, Z.; Žáková, E. Estimation of economic weights for number of teats and sperm quality traits in pigs. J. Anim. Breed Genet. 2020, 137, 189–199. [Google Scholar] [CrossRef]
- Halimani, T.E.; Muchadeyi, F.C.; Chimonyo, M.; Dzama, K. Some Insights into the phenotypic and genetic diversity of indigenous pigs in Southern Africa. S. Afr. J. Anim. Sci. 2012, 42, 507–510. [Google Scholar] [CrossRef]
- Oliviero, C.; Junnikkala, S.; Peltoniemi, O. The challenge of large litters on the immune system of the sow and the piglets. Reprod. Domest. Anim. 2019, 54, 12–21. [Google Scholar] [CrossRef]
- Lopes, M.S.; Bastiaansen, J.W.; Harlizius, B.; Knol, E.F.; Bovenhuis, H. A Genome-wide association study reveals dominance effects on number of teats in pigs. PLoS ONE 2014, 9, e105867. [Google Scholar] [CrossRef]
- Duijvesteijn, N.; Veltmaat, J.M.; Knol, E.F.; and Harlizius, B. High-resolution association mapping of number of teats in pigs reveals regions controlling vertebral development. BMC Genom. 2014, 15, 542. [Google Scholar] [CrossRef]
- Makhanya, L.G. Phenotypic and Reproductive Characterisation of Kolbroek Pigs. Master’s Thesis, Central University of Technology, Bloemfontein, South Africa, 2018. [Google Scholar]
- Verardo, L.L.; Silva, F.F.; Varona, L.; Resende, M.D.V.; Bastiaansen, J.W.M.; Lopes, P.S.; Guimarães, S.E.F. Bayesian GWAS and network analysis revealed new candidate genes for number of teats in pigs. J. Appl. Genet. 2015, 56, 123–132. [Google Scholar] [CrossRef]
- Van Hossou, S.F.U.; Yin, T.; Scheper, C.; Fries, R.; Dossa, L.H.; König, S. Unravelling admixture, inbreeding, and recent selection signatures in West African indigenous cattle populations in Benin. Front. Genet. 2021, 12, 657282. [Google Scholar] [CrossRef]
- VonHoldt, B.M.; Hinton, J.W.; Shutt, A.C.; Murphy, S.M.; Karlin, M.L.; Adams, J.R.; Waits, L.P.; Brzeski, K.E. Reviving ghost alleles: Genetically admixed coyotes along the American Gulf Coast are critical for saving the endangered red wolf. Sci. Adv. 2022, 8, eabn7731. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Category | Population | Code | N |
|---|---|---|---|
| Village | Mopani | MOP | 27 |
| Village | Capricorn | CAP | 25 |
| Village | Oliver Reginald Tambo | ORT | 22 |
| Village | Alfred Nzo | ALN | 17 |
| Commercial | Large White | LWT | 20 |
| Commercial | SA Landrace | SAL | 20 |
| Commercial | Duroc | DUR | 20 |
| Indigenous | Kolbroek | KOL | 20 |
| Indigenous | Windsnyer Type | WIN | 20 |
| Asian | Vietnamese Potbelly | VIT | 5 |
| Wild | Wild boar | WBO | 4 |
| Wild | Wild boar | WAT | 31 |
| Wild | Wild boar | BSP | 3 |
| Populations | Chr | Start | End | Gene | QTLs |
|---|---|---|---|---|---|
| ALN | 2 | 25247505 | 25378565 | TRIM44 | Spinal Curvature |
| 7 | 95982739 | 96254303 | DPF3 | Teat number | |
| 8 | 31921587 | 32301975 | APBB2 | Stearic acid content | |
| 13 | 29753308 | 29775371 | PTH1R | Front leg conformation, Hind leg conformation, Hip structure | |
| ORT | 1 | 162364055 | 162737492 | NEDD4L | Intramuscular fat content |
| 2 | 87678811 | 87866856 | DMGDH | Litter weight piglets born alive | |
| 8 | 80290715 | 80669404 | NR3C2 | Loin muscle, Teat number | |
| 12 | 53979215 | 54055377 | PIK3R5 | Intramuscular fat content | |
| 12 | 27957732 | 28617585 | CA10 | Hemaglobin | |
| 12 | 44617499 | 44660271 | VTN | Body depth, Drip loss, Hind leg conformation, Hip structure, pH 24 h post-mortem (loin), ph 45 min post-mortem | |
| MOP | 4 | 71045333 | 71318030 | NKAIN3 | Body weight (birth) |
| 8 | 31921587 | 32301975 | APBB2 | Stearic acid content | |
| 16 | 27537885 | 27631130 | SELENOP | Meat colour a | |
| 16 | 32142034 | 32148335 | PELO | Obesity index, Teat number | |
| 16 | 32336292 | 32437103 | ITGA2 | Body weight (5 weeks) | |
| CAP | 1 | 183915339 | 184140122 | SAMD4A | Intramuscular fat content |
| 12 | 27416337 | 27436160 | NME1 | Conductivity 24 h post-mortem (loin), Cooking loss, Loin muscle depth, Loin weight | |
| 12 | 27957732 | 28617585 | CA10 | Hemoglobin | |
| 14 | 101123381 | 101254725 | LIPA | Average daily gain, Front leg weight, HDL/LDL ratio, Litter size, Loin muscle area, Monounsaturated fatty acid content, Oleic acid content, Skin thickness, Sperm concentration, Teat number | |
| 14 | 103992037 | 104106576 | IDE | e.g., Abdominal fat percentage, Age at puberty, Age at slaughter, Average backfat thickness, Average daily gain, Backfat at first rib, Backfat at last rib, Backfat between 3rd and 4th last ribs, Body depth, Body height | |
| 14 | 105036770 | 105044765 | RBP4 | Litter size, Total number born alive | |
| 14 | 12437515 | 12563544 | EXTL3 | Fat androstenone level | |
| 8 | 76482022 | 76699351 | FBXW7 | Body mass index | |
| 8 | 31921587 | 32301975 | APBB2 | Stearic acid content | |
| LWT | 2 | 108255930 | 108536881 | PAM | Intramuscular fat content, Loin percentage, Maternal infanticide, Teat number |
| 7 | 26860140 | 26990017 | LRRC1 | Meat colour L | |
| 9 | 48120802 | 48286278 | TECTA | Backfat between 3rd and 4th last ribs | |
| 13 | 177365717 | 179013542 | ROBO2 | Feed efficiency, Linolenic acid content | |
| 14 | 69200215 | 70938204 | CTNNA3 | Teat number | |
| DUR | 6 | 137595524 | 138010444 | SLC44A5 | Diameter of muscle fibers |
| 7 | 27389874 | 27895570 | KHDRBS2 | Loin muscle area, Loin muscle depth, Teat number | |
| KOL | 4 | 61628299 | 61716546 | JPH1 | Intramuscular fat content |
| 14 | 79352396 | 80106258 | KCNMA1 | Meat colour b | |
| WIN | 5 | 18718158 | 18723843 | TARBP2 | Backfat between 3rd and 4th last ribs |
| 8 | 127731903 | 128953331 | CCSER1 | Backfat between 3rd and 4th last ribs, Intramuscular fat content | |
| VIT | 2 | 79766349 | 80141293 | COL23A1 | Front foot size, Hip structure |
| 4 | 79687359 | 79847281 | PRKDC | Feed conversion ratio | |
| 6 | 79849687 | 79958271 | HSPG2 | Days to 113 kg, Marbling | |
| 14 | 79352396 | 80106258 | KCNMA1 | Meat colour b | |
| WAT | 18 | 31027031 | 31125465 | MDFIC | Fat androstenone level |
| WBO | 2 | 15791451 | 15819138 | F2 | e.g., Age at slaughter, Average backfat thickness, Average daily gain, Backfat at last rib, Backfat at rump, Backfat thickness between 3rd and 4th rib, Body weight (end of test), Body weight (weaning) |
| 5 | 4769801 | 4849334 | SHISAL1 | Intramuscular fat content | |
| 5 | 6997444 | 7014422 | POLR3H | Fat androstenone level | |
| 13 | 66316436 | 66452917 | GHRL | Age at slaughter, Average daily gain, Daily feed intake, Days to 100 kg, Feed intake, Loin weight, Marbling | |
| 14 | 21754463 | 22121051 | SPOCK3 | Fat androstenone level |
| Populations | Chr | Start | End | Gene | QTLs |
|---|---|---|---|---|---|
| DUR_EC | 1 | 166173135 | 166310972 | ITGA11 | Obesity index, Teat number |
| 2 | 113774412 | 114206930 | FER | Abdominal circumference, Average backfat thickness, Average daily gain, Backfat at last lumbar, Biceps brachii weight, Body height, Body weight (3 weeks), Carcass weight (hot), Double-bond index | |
| 6 | 80649143 | 80843567 | EPHB2 | Litter weight total | |
| DUR_LIM | 1 | 166173135 | 166310972 | ITGA11 | Obesity index, Teat number |
| 6 | 80649143 | 80843567 | EPHB2 | Litter weight total | |
| DUR_IND | 18 | 40820644 | 41409087 | PDE1C | Backfat at rump |
| 18 | 42030510 | 42046184 | GHRHR | Backfat at last rump, Carcass length, Fat-cuts percentage | |
| DUR_LWT | 2 | 113774412 | 114206930 | FER | e.g., Abdominal circumference, Arachidonic acid content, Aspartate aminotransferase activity, Average backfat thickness, Average daily gain, Backfat at last lumbar, Backfat at mid-back, Backfat at rump, Backfat at tenth rib |
| 7 | 27389874 | 27895570 | KHDRBS2 | Loin muscle area, Loin muscle depth, Teat number | |
| 7 | 30708332 | 30724045 | SNRPC | Loin muscle area, Loin muscle depth | |
| 7 | 30731461 | 30802735 | UHRF1BP1 | Femur length, Hip bone length, Humerus length, Tibia length, Ulna length | |
| 7 | 30812920 | 30995361 | ANKS1A | Femur length, Hip bone length, Humerus length, Tibia length, Ulna length, Galt score (front), Loin muscle area, Loin muscle depth | |
| 7 | 31586555 | 31603617 | ARMC12 | Facial morphology | |
| 7 | 31722990 | 31792904 | SLC26A8 | Facial morphology, Femur length, Humerus length, Tibia length, Ulna length | |
| 14 | 111834168 | 111914304 | PAX2 | Monounsaturated fatty acid to saturated fatty acid ratio, Oleic acid to stearic acid ratio, Palmitoleic acid to palmitic acid ratio, Stearic acid content | |
| DUR_VIT | 2 | 72326714 | 72391648 | VAV1 | Average daily gain, Backfat between 3rd and 4th last rib, Birth weight variability, Body weight (end of test), Conductivity 45 min post-mortem, Fat androstenone level, Intramuscular fat content, Time in feeder per day, pH 24 h postmortem (ham), pH 45 min postmortem |
| DUR_WBO | 1 | 193722164 | 193906565 | ESR2 | Front leg conformation, Gait score (overall), Hind leg conformation, Litter size, Maternal infanticide, Plasma droplet rate, Semen volume, Sperm concentration, Sperm motility, Total number born alive |
| WAT_EC | 1 | 254683885 | 254703225 | AMBP | Conductivity 24 h post-mortem (loin), pH 24 h postmortem (ham), pH 24 h post-mortem (loin), pH 45 min postmortem |
| 3 | 39957269 | 39957727 | NPW | Lean meat percentage | |
| 8 | 37530815 | 37809761 | CORIN | Platelet count | |
| 8 | 37797875 | 37875540 | NFXL1 | Mean corpuscular hemoglobin content, Mean corpuscular volume | |
| 8 | 47473787 | 47601359 | RXFP1 | Red blood cell count | |
| 8 | 71520275 | 71554766 | PPEF2 | Platelet distribution width | |
| 8 | 71573783 | 71603338 | NAAA | Platelet distribution width | |
| 8 | 72543620 | 72679173 | SEPTIN11 | Teat number | |
| 8 | 73502664 | 73958083 | FRAS1 | Teat number | |
| WAT_LIM | 1 | 254683885 | 254703225 | AMBP | Conductivity 24 h post-mortem (loin), pH 24 h postmortem (ham), pH 24 h post-mortem (loin), pH 45 min postmortem |
| 8 | 71520275 | 71554766 | PPEF2 | Platelet distribution width | |
| 8 | 71573783 | 71603338 | NAAA | Platelet distribution width | |
| 14 | 113414264 | 113429936 | PSD | Intramuscular fat content, Oleic acid to stearic acid ratio | |
| 14 | 113464174 | 113478699 | MFSD13A | Oleic acid content, Oleic acid to stearic acid ratio, Stearic acid content | |
| 14 | 113480197 | 113498740 | ACTR1A | Oleic acid content, Stearic acid content | |
| 9 | 45400464 | 45435916 | TMPRSS4 | Backfat between 3rd and 4th last ribs | |
| WAT_LWT | 4 | 105804586 | 105845725 | CSDE1 | Intramuscular fat content |
| 4 | 105868897 | 105893771 | AMPD1 | Juiciness score, Overall impression, sensory panel, Tenderness score | |
| WAT_WBO | 7 | 89120822 | 89168270 | MAX | Meat colour b, Teat number, maximum per side |
| VIT_EC | 18 | 29895878 | 29936233 | TES | Average daily gain, Backfat between 3rd and 4th last rib, Birth weight variability, Body weight (end of test), Conductivity 45 min post-mortem, Fat androstenone level, Intramuscular fat content, Time in feeder per day, pH 24 h postmortem (ham), pH 45 min postmortem, Teat number |
| VIT_LIM | 2 | 72326714 | 72391648 | VAV1 | Average daily gain, Backfat between 3rd and 4th last rib, Birth weight variability, Body weight (end of test), Conductivity 45 min post-mortem, Fat androstenone level, Intramuscular fat content, Time in feeder per day, pH 24 h postmortem (ham), pH 45 min postmortem |
| 18 | 29895878 | 29936233 | TES | Average daily gain, Backfat between 3rd and 4th last rib, Birth weight variability, Body weight (end of test), Conductivity 45 min post-mortem, Fat androstenone level, Intramuscular fat content, Time in feeder per day, pH 24 h postmortem (ham), pH 45 min postmortem, Teat number | |
| VIT_IND | 18 | 29895878 | 29936233 | TES | Average daily gain, Backfat between 3rd and 4th last rib, Birth weight variability, Body weight (end of test), Conductivity 45 min post-mortem, Fat androstenone level, Intramuscular fat content, Time in feeder per day, pH 24 h postmortem (ham), pH 45 min postmortem, Teat number |
| 18 | 31027031 | 31125465 | MDFIC | Fat androstenone level | |
| VIT_WBO | 1 | 193722164 | 193906565 | ESR2 | Front leg conformation, Gait score (overall), Hind leg conformation, Litter size, Maternal infanticide, Plasma droplet rate, Semen volume, Sperm concentration, Sperm motility, Total number born alive |
| Chr | Start | End | Gene | QTLs |
|---|---|---|---|---|
| 5 | 44381009 | 44558744 | FAR2 | Feed conversion ratio |
| 5 | 56817606 | 57005114 | EPS8 | Fat androstenone level |
| 5 | 64519186 | 65002098 | VWF | Litter size |
| 5 | 66665263 | 66838922 | PRMT8 | Teat number |
| 5 | 28300950 | 28605120 | SRGAP1 | Ear area |
| 5 | 29695839 | 29863599 | MSRB3 | Ear area |
| 5 | 97092883 | 97148601 | SLC6A15 | Time in feeder per day |
| 5 | 33858572 | 34033939 | CCT2 | Feed conversion ratio |
| 5 | 34067992 | 34218513 | MYRFL | Feed conversion ratio |
| 5 | 34660029 | 34794321 | PTPRB | Feed conversion ratio |
| 5 | 36274364 | 36658703 | TRHDE | Feed conversion ratio |
| 6 | 97342474 | 97429364 | GNAL | Age at puberty, Arachidic acid content, Average backfat thickness, Average daily gain, Backfat at last lumbar, Backfat at last rib, Backfat at rump, Backfat at tenth rib, Body weight (16 days), Carcass weight (hot), ear area, Feed conversion ratio, Lean meat percentage, Loin muscle area, Loin muscle depth, Oleic acid content, Oleic acid to stearic acid ratio, PH for longissmus dorsi, Stearic acid content, Teat number, Vertebra number, Androstenone laboratory |
| 6 | 108115886 | 108227748 | CABLES1 | Average daily gain, Backfat at rump |
| 6 | 108548837 | 108805693 | LAMA3 | Average daily gain |
| 6 | 75696484 | 75755892 | PADI2 | Fat androstenone level |
| 6 | 112396721 | 112628432 | CDH2 | Average daily gain, Intramuscular fat content, Lean meat percentage, Obesity index |
| 6 | 117631227 | 118334105 | NOL4 | Average backfat thickness |
| 6 | 79849687 | 79958271 | HSPG2 | Days to 113 kg, Marbling |
| 6 | 125890708 | 126043161 | PIK3C3 | Average backfat thickness, Average daily gain, Intramuscular fat content, Loin muscle area |
| 6 | 80649143 | 80843567 | EPHB2 | Litter weight (total) |
| 6 | 137595524 | 138010444 | SLC44A5 | Diameter of muscle fibers |
| 6 | 43933759 | 44069330 | GPI | Average backfat thickness, Body weight (5 weeks), Intramuscular fat content, Osteochondrosis score |
| 6 | 46442857 | 46470075 | ZNF570 | Lean meat percentage |
| 14 | 47946396 | 48040822 | LIMK2 | Fat androstenone level, Melanoma susceptibility |
| 14 | 122777130 | 122828051 | ACSL5 | Fat androstenone level |
| 14 | 123343694 | 123546417 | TCF7L2 | Carcass weight (hot), Number of visits to feeder per day |
| 14 | 133460167 | 133544018 | CHST15 | Teat number |
| 14 | 124760398 | 125010892 | ABLIM1 | Fat androstenone level, Intramuscular fat content |
| 15 | 100868469 | 101211242 | ANKRD44 | Skin thickness |
| 15 | 101623818 | 101957516 | PLCL1 | Skin thickness |
| 15 | 118335925 | 118452296 | XRCC5 | Average backfat thickness, Conductivity 24 h post-mortem (loin), Cooking loss, Fat weight (total), Lean meat percentage, Loin muscle area, Loin muscle depth, Loin weight, PH for longissmusdorsi, Subcutanous fat area, pH 24 h postmortem (ham), pH 24 h post-mortem (ham), pH 24 h post-mortem (loin) |
| 15 | 79935994 | 80025850 | SP3 | Cooking loss, Meat colour b, Shear force, Thawing loss |
| 16 | 32130235 | 32304592 | ITGA1 | Obesity index, Teat number |
| 16 | 32336292 | 32437103 | ITGA2 | Body weight (5 weeks) |
| 16 | 33126494 | 33565542 | ARL15 | Loin muscle area |
| 18 | 34006688 | 34906268 | IMMP2L | Age at puberty |
| 18 | 40820644 | 41409087 | PDE1C | Backfat at rump |
| 18 | 42030510 | 42046184 | GHRHR | Backfat at last rib, Carcass length, Fat-cuts percentage |
| 18 | 51387836 | 51802945 | HECW1 | Teat number |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hlongwane, N.L.; Dzomba, E.F.; Hadebe, K.; van der Nest, M.A.; Pierneef, R.; Muchadeyi, F.C. Identification of Signatures of Positive Selection That Have Shaped the Genomic Landscape of South African Pig Populations. Animals 2024, 14, 236. https://doi.org/10.3390/ani14020236
Hlongwane NL, Dzomba EF, Hadebe K, van der Nest MA, Pierneef R, Muchadeyi FC. Identification of Signatures of Positive Selection That Have Shaped the Genomic Landscape of South African Pig Populations. Animals. 2024; 14(2):236. https://doi.org/10.3390/ani14020236
Chicago/Turabian StyleHlongwane, Nompilo L., Edgar F. Dzomba, Khanyisile Hadebe, Magriet A. van der Nest, Rian Pierneef, and Farai C. Muchadeyi. 2024. "Identification of Signatures of Positive Selection That Have Shaped the Genomic Landscape of South African Pig Populations" Animals 14, no. 2: 236. https://doi.org/10.3390/ani14020236
APA StyleHlongwane, N. L., Dzomba, E. F., Hadebe, K., van der Nest, M. A., Pierneef, R., & Muchadeyi, F. C. (2024). Identification of Signatures of Positive Selection That Have Shaped the Genomic Landscape of South African Pig Populations. Animals, 14(2), 236. https://doi.org/10.3390/ani14020236