
Curcumin Mitigates Oxidative Damage in Broiler Liver and Ileum Caused by Aflatoxin B1-Contaminated Feed through Nrf2 Signaling Pathway

Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Chemicals
2.2. Animal and Feeding Management
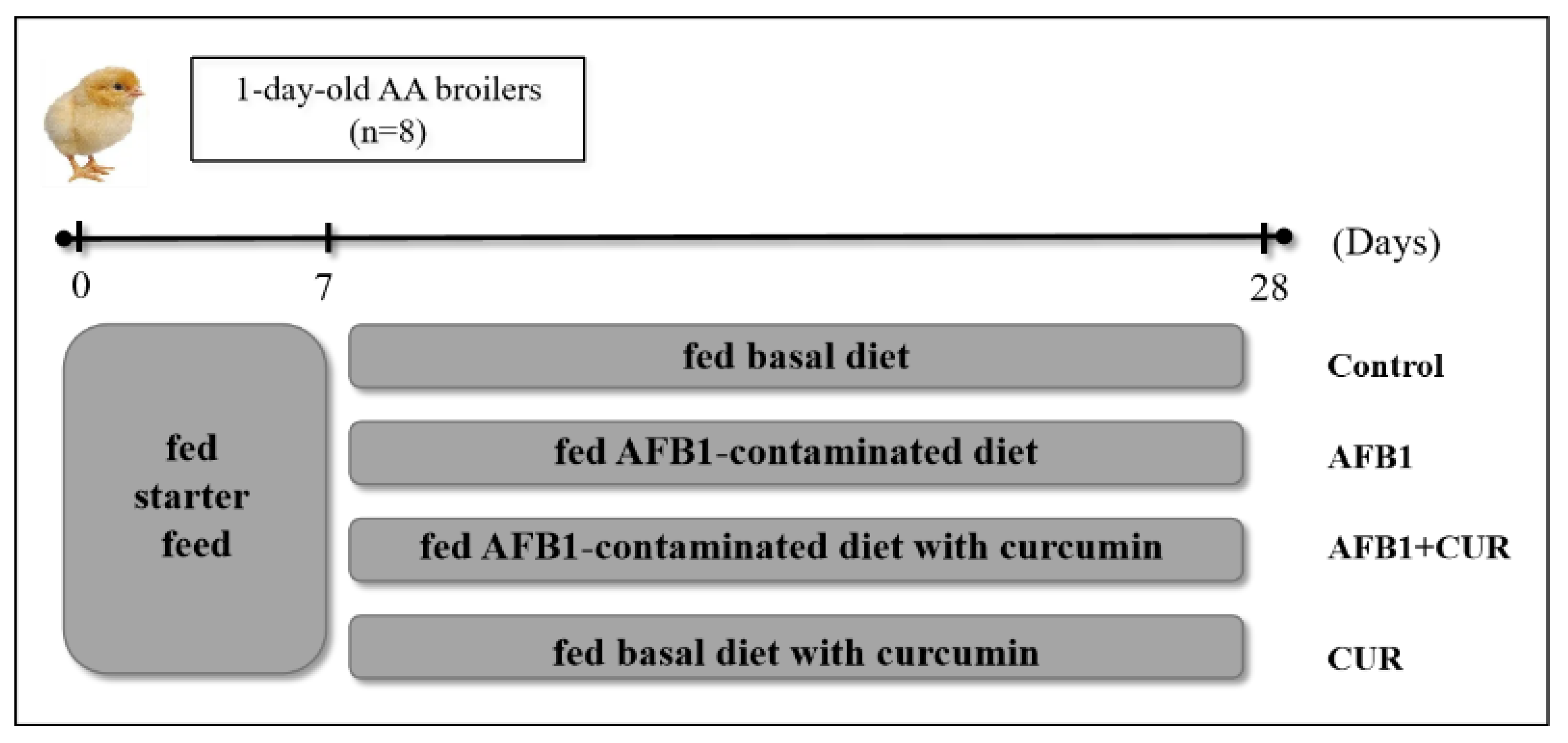
2.3. Experimental Design
2.4. Sample Collection
2.5. Organ Index Measurement
2.6. Histological Analysis
2.7. Determination of Antioxidant Indicators
2.8. Determination of Intestinal Permeability
2.9. Transcriptional Analyze
2.10. Statistical Analysis
3. Result
3.1. Effect of CUR on the Growth Performance of Broilers Exposed to AFB1
3.2. Effect of CUR on the Organ Weight and Organ Indexes of Broilers Exposed to AFB1
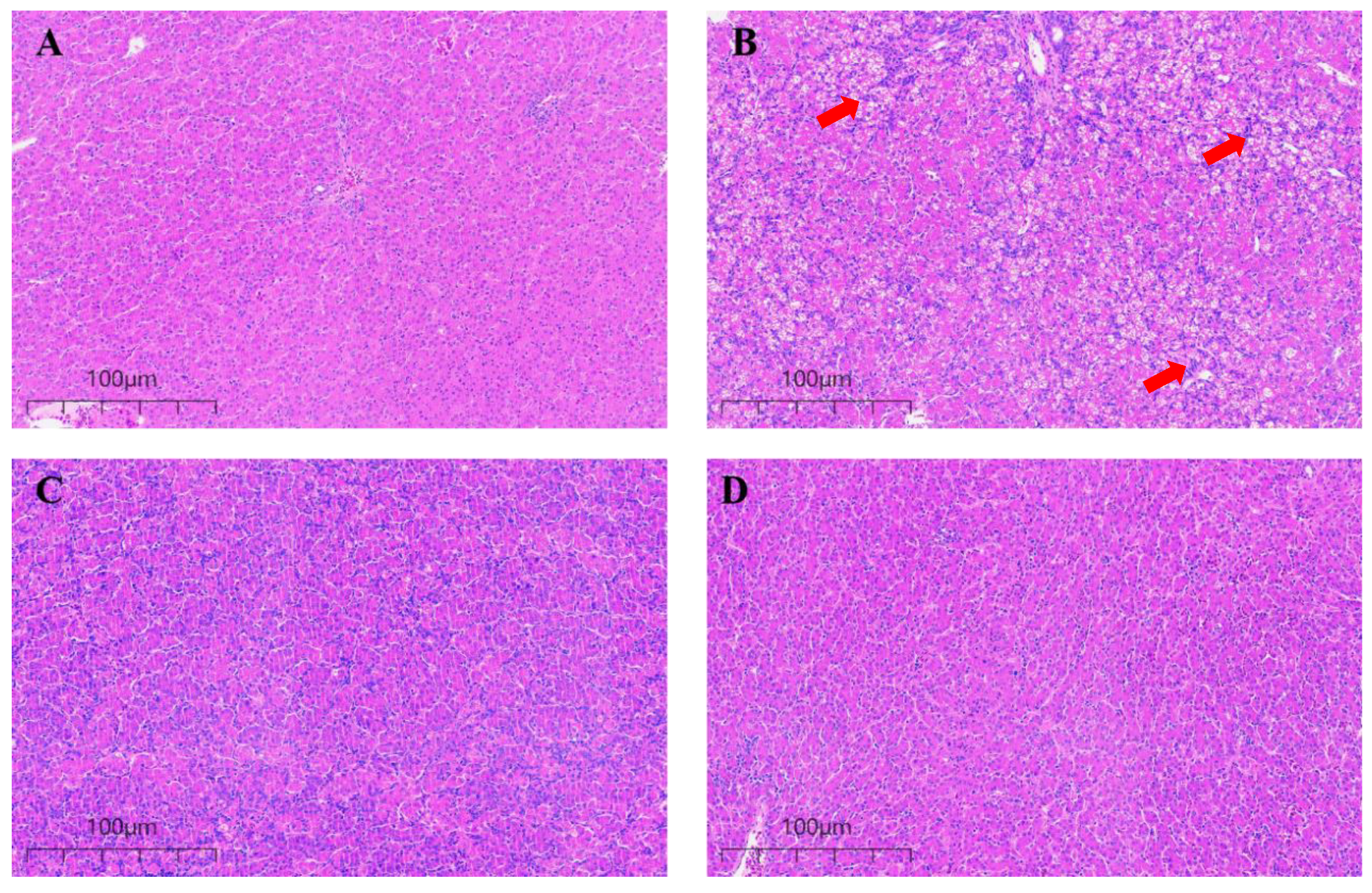
3.3. Effect of CUR on the Microstructure of Broiler Liver Exposed to AFB1
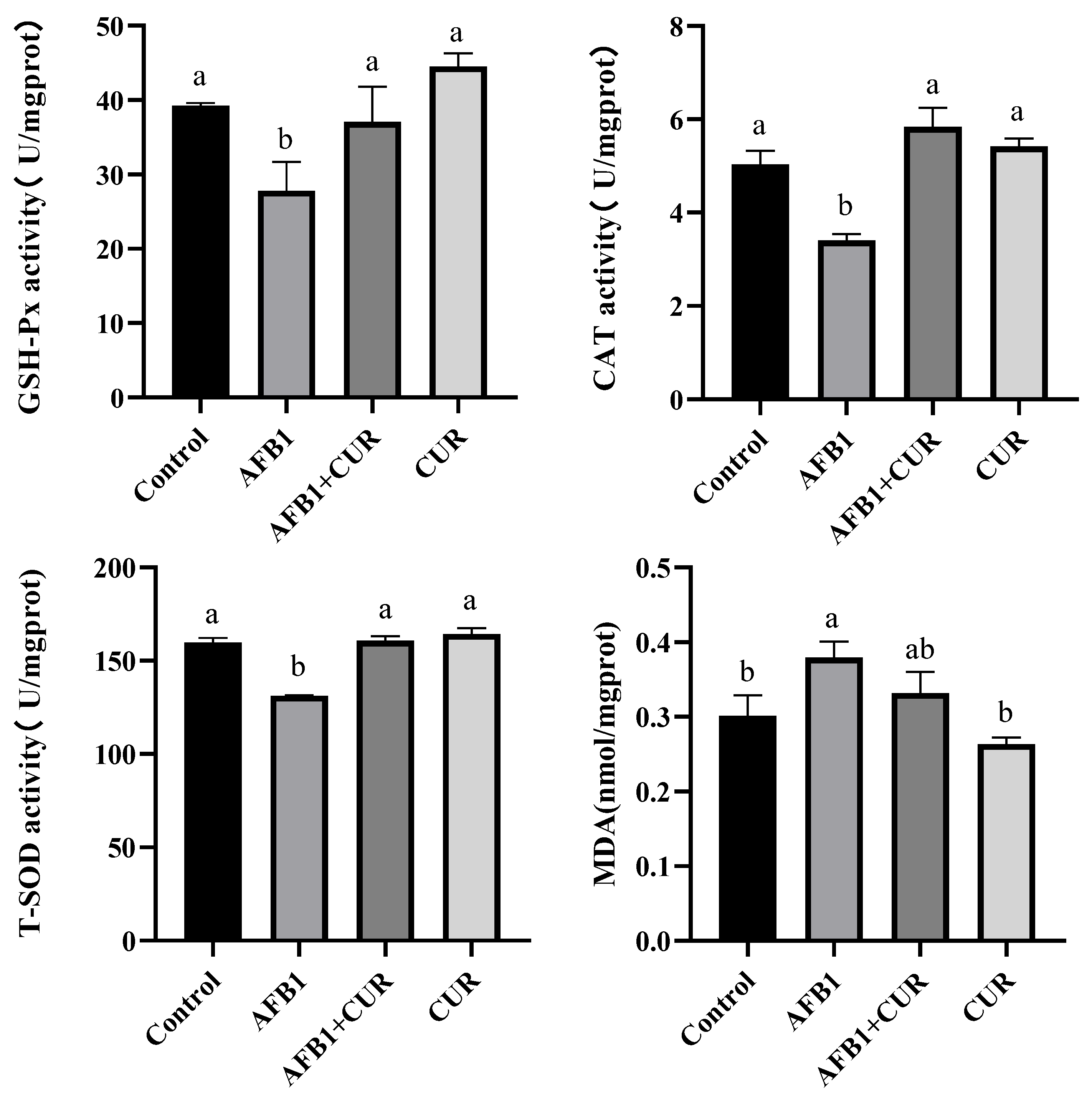
3.4. Effect of CUR on the Antioxidant Indicators of Broiler Liver Exposed to AFB1
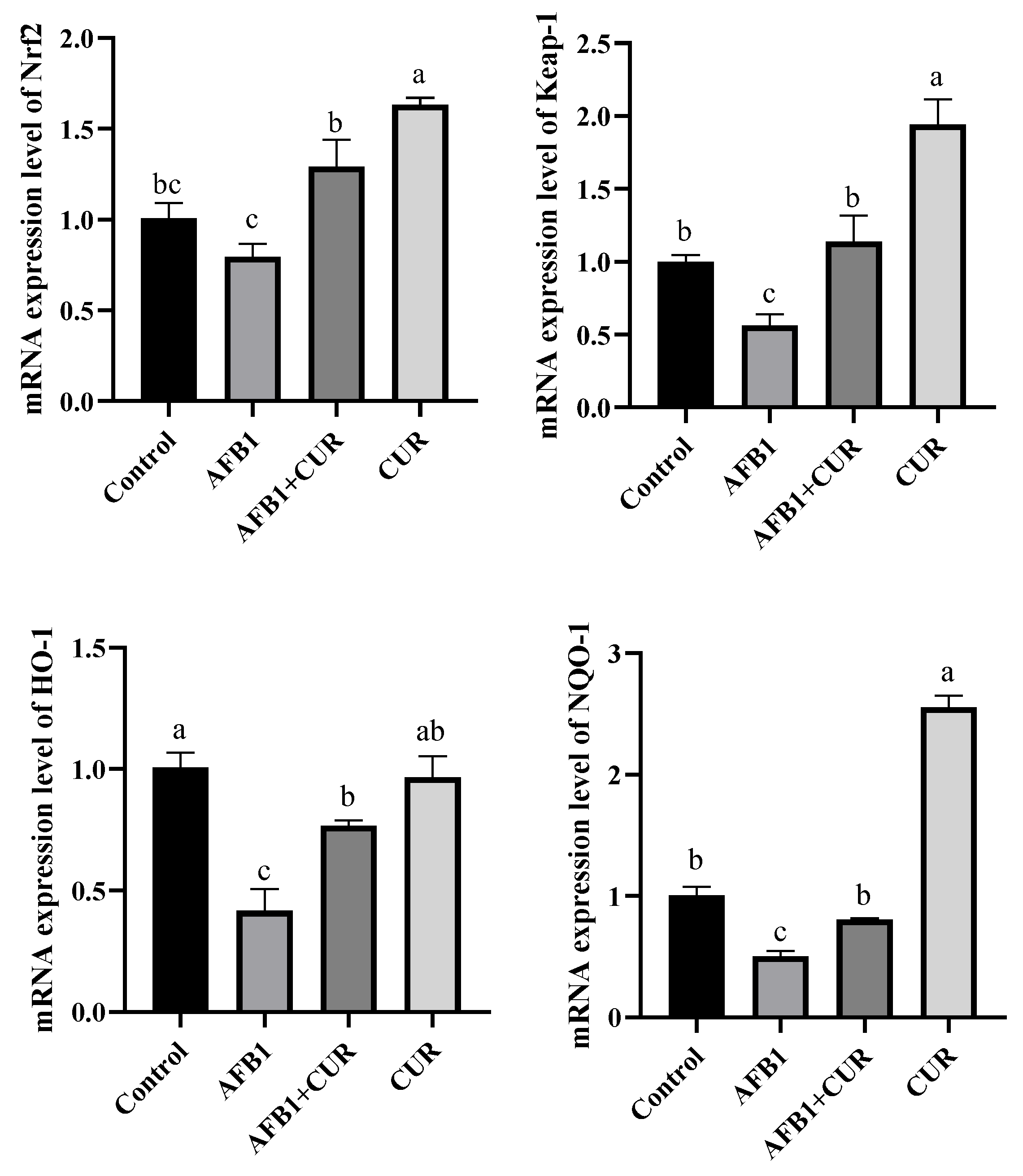
3.5. Expression Levels of Nrf2 Pathway Genes in Liver
3.6. Effect of CUR on the Relative Weight and Length of Broiler Intestine Exposed to AFB1
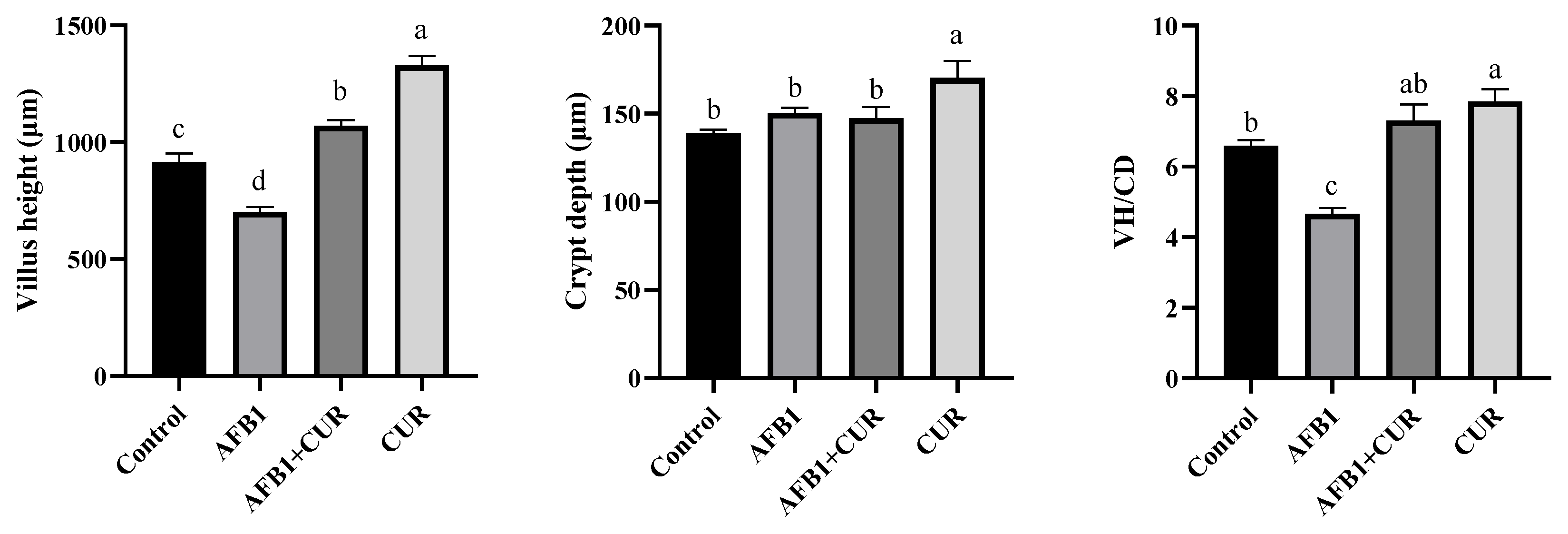
3.7. Effect of CUR on the Ileum Morphology of Broilers Exposed to AFB1
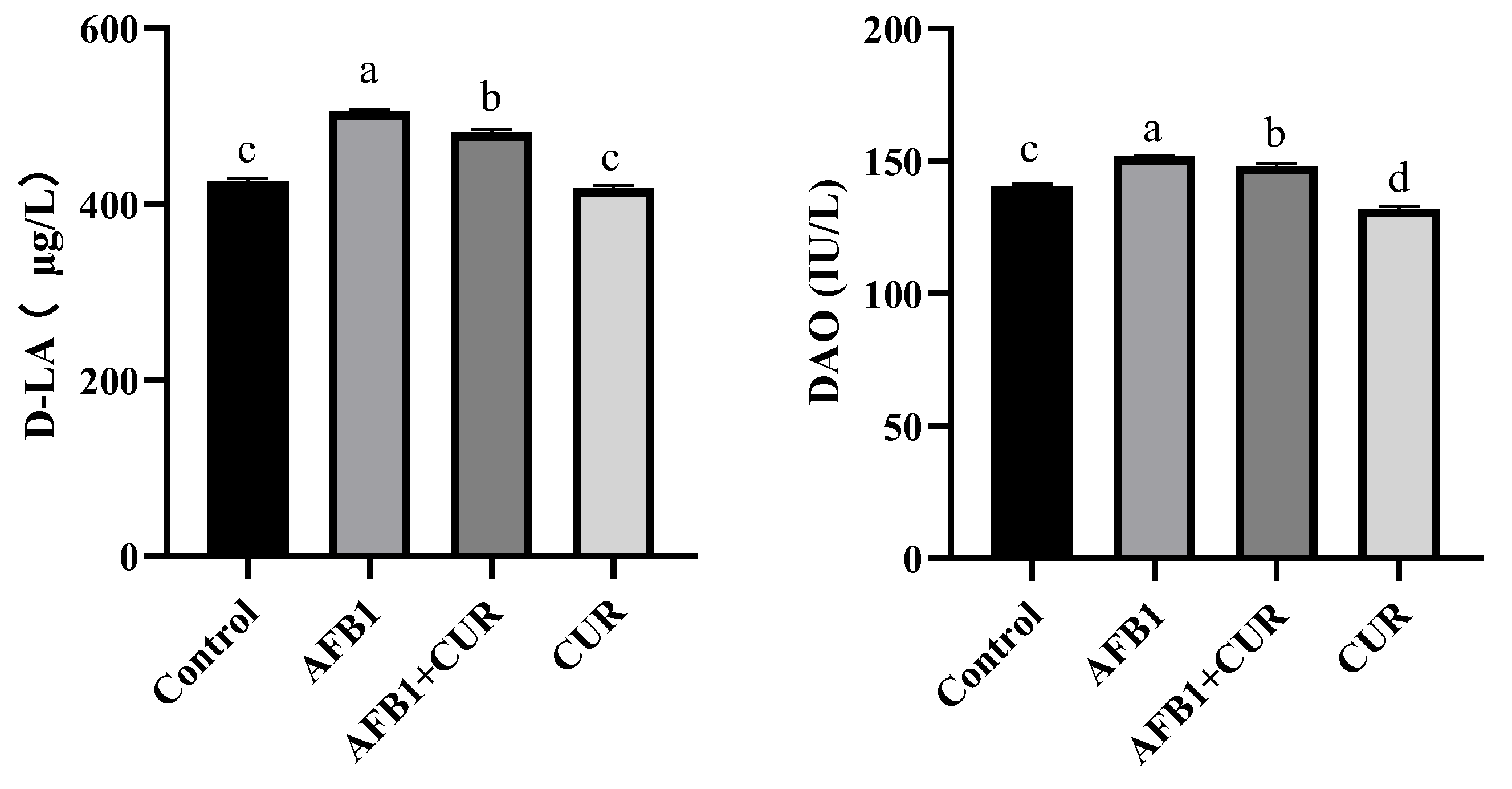
3.8. Effect of CUR on the Intestinal Permeability of Broilers Exposed to AFB1
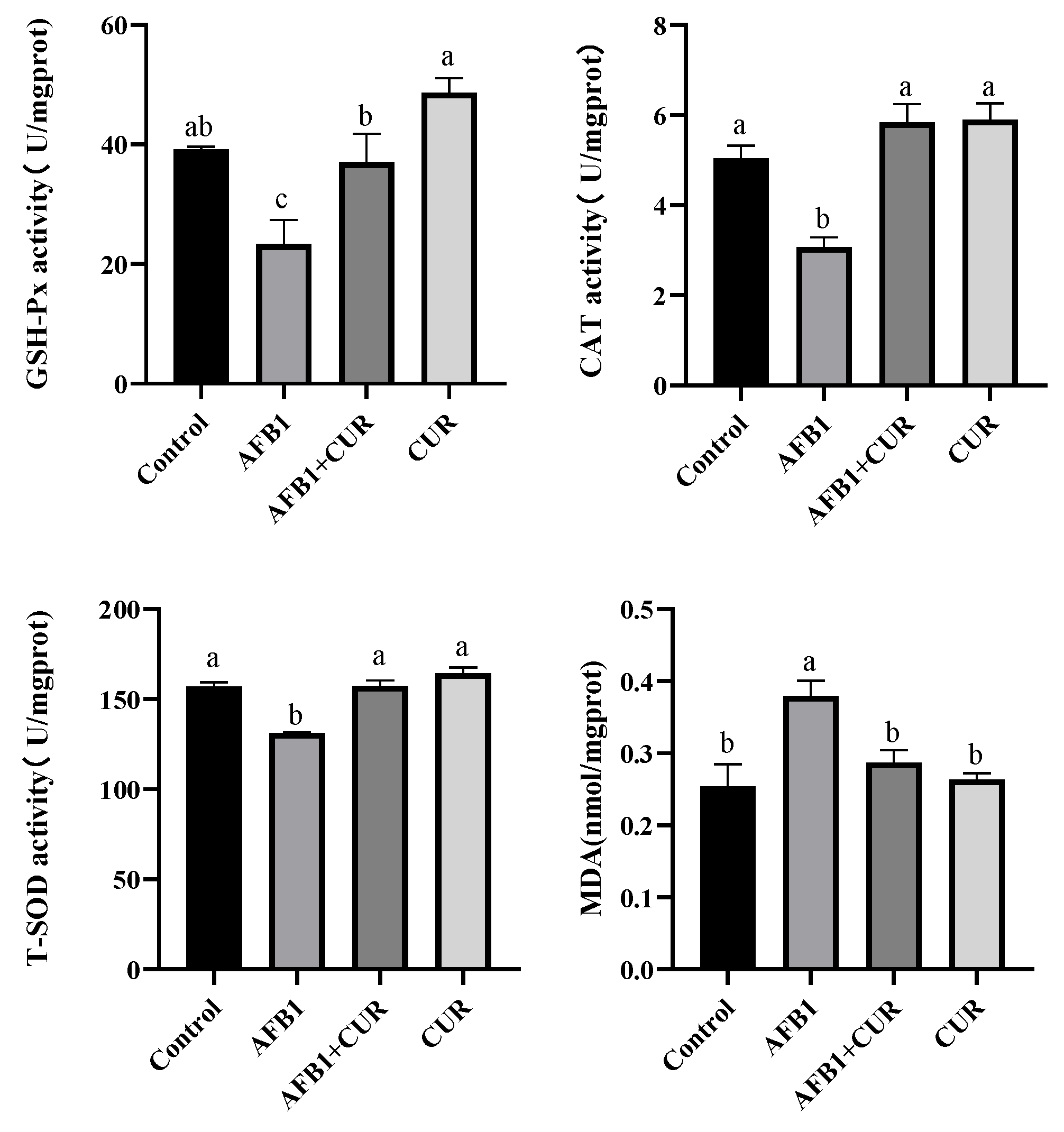
3.9. Effect of CUR on the Antioxidant Indicators of Broiler Ileum Exposed to AFB1
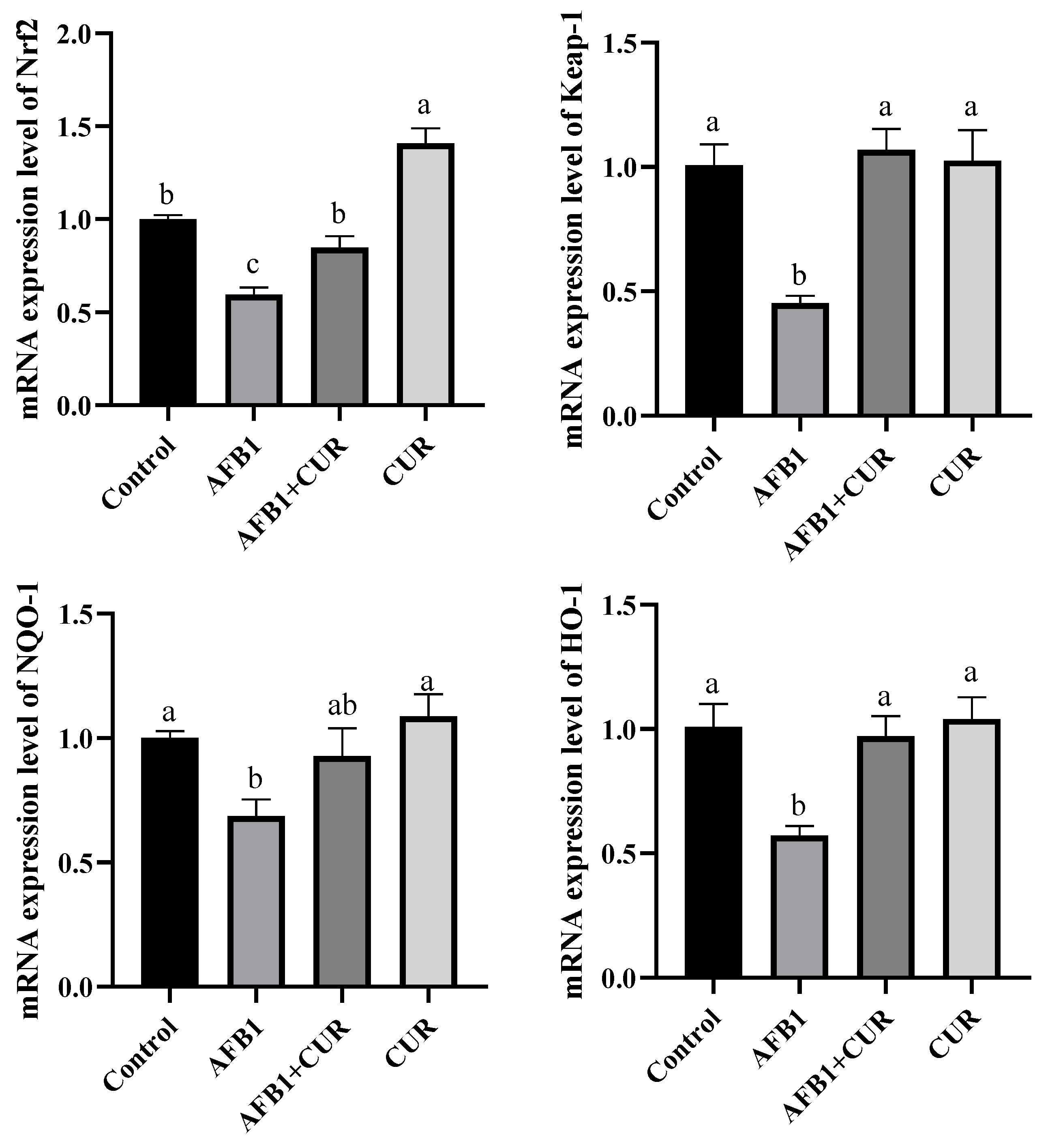
3.10. Effect of CUR on the Expression of Nrf2 Pathway Genes in Broiler Ileum Exposed to AFB1
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Fan, T.; Xie, Y.; Ma, W. Research progress on the protection and detoxification of phytochemicals against aflatoxin B-induced liver toxicity. Toxicon 2021, 195, 58–68. [Google Scholar] [CrossRef]
- Wang, Y.; Wang, X.; Li, Q. Aflatoxin B in poultry liver: Toxic mechanism. Toxicon 2023, 233, 107262. [Google Scholar] [CrossRef]
- Li, Q.; Zhang, M.; Sun, J.; Li, Y.; Zu, S.; Xiang, Y.; Jin, X. Porcine β-defensin-2 alleviates aflatoxin B1 induced intestinal mucosal damage via ROS-Erk signaling pathway. Sci. Total Environ. 2023, 905, 167201. [Google Scholar] [CrossRef]
- Yang, H.; Wang, Y.; Jin, S.; Pang, Q.; Shan, A.; Feng, X. Dietary resveratrol alleviated lipopolysaccharide-induced ileitis through Nrf2 and NF-κB signaling pathways in ducks (Anas platyrhynchos). J. Anim. Physiol. Anim. Nutr. 2022, 106, 1306–1320. [Google Scholar] [CrossRef]
- Xiao, S.; Wu, Y.; Gao, S.; Zhou, M.; Liu, Z.; Xiong, Q.; Jiang, L.; Yuan, G.; Li, L.; Yang, L. Deciphering the Hazardous Effects of AFB1 and T-2 Toxins: Unveiling Toxicity and Oxidative Stress Mechanisms in PK15 Cells and Mouse Kidneys. Toxins 2023, 15, 503. [Google Scholar] [CrossRef] [PubMed]
- Frangiamone, M.; Yemelin, A.; Cimbalo, A.; Font, G.; Thines, E.; Manyes, L. Fermented whey modulated AFB1 and OTA-induced hepatotoxicity and nephrotoxicity in vivo. A relative and absolute quantification about sex differences. Toxicol. Mech. Methods 2023, 33, 529–540. [Google Scholar] [CrossRef] [PubMed]
- Cao, W.; Yu, P.; Yang, K.; Cao, D. Aflatoxin B1: Metabolism, toxicology, and its involvement in oxidative stress and cancer development. Toxicol. Mech. Methods 2022, 32, 395–419. [Google Scholar] [CrossRef]
- Mughal, M.; Xi, P.; Yi, Z.; Jing, F. Aflatoxin B1 invokes apoptosis via death receptor pathway in hepatocytes. Oncotarget 2017, 8, 8239–8249. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Sang, R.; Zhao, X.; Li, C.; Wang, W.; Wang, M.; Ge, B.; Zhang, X. Research Note: Taraxasterol alleviates aflatoxin B1-induced oxidative stress in chicken primary hepatocytes. Poult. Sci. 2023, 102, 102286. [Google Scholar] [CrossRef]
- Wang, F.; Zuo, Z.; Chen, K.; Gao, C.; Yang, Z.; Zhao, S.; Li, J.; Song, H.; Peng, X.; Fang, J.; et al. Histopathological Injuries, Ultrastructural Changes, and Depressed TLR Expression in the Small Intestine of Broiler Chickens with Aflatoxin B1. Toxins 2018, 10, 131. [Google Scholar] [CrossRef]
- Sarker, M.; Wan, X.; Yang, H.; Wang, Z. Dietary Lycopene Supplementation Could Alleviate Aflatoxin B Induced Intestinal Damage through Improving Immune Function and Anti-Oxidant Capacity in Broilers. Animals 2021, 11, 3165. [Google Scholar] [CrossRef] [PubMed]
- Tao, W.; Zhu, W.; Nabi, F.; Li, Z.; Liu, J. Penthorum chinense Pursh compound flavonoids supplementation alleviates Aflatoxin B1-induced liver injury via modulation of intestinal barrier and gut microbiota in broiler. Ecotoxicol. Environ. Saf. 2023, 255, 114805. [Google Scholar] [CrossRef] [PubMed]
- Kensler, T.W.; Wakabayashi, N.; Biswal, S. Cell survival responses to environmental stresses via the Keap1-Nrf2-ARE pathway. Annu. Rev. Pharmacol. Toxicol. 2007, 47, 89–116. [Google Scholar] [CrossRef]
- Loeber, C.C.; Buechner, A.E. Dissertatio Inauguralis Medica de Curcuma officinarum Ejusque Genuinis Virtutibus. Diss Inaug Halae, eds Præs, Büchnero AE. Ph.D. Thesis, Halae Magdeburgicae, Halle, Germany, 1748; pp. 1–28. [Google Scholar]
- Cheng, A.L.; Hsu, C.H.; Lin, J.K.; Hsu, M.M.; Ho, Y.F.; Shen, T.S.; Ko, J.Y.; Lin, J.T.; Lin, B.R.; Ming-Shiang, W.; et al. Phase I clinical trial of curcumin, a chemopreventive agent, in patients with high-risk or pre-malignant lesions. Anticancer Res. 2001, 21, 2895–2900. [Google Scholar] [PubMed]
- Pulido-Moran, M.; Moreno-Fernandez, J.; Ramirez-Tortosa, C.; Ramirez-Tortosa, M. Curcumin and Health. Molecules 2016, 21, 264. [Google Scholar] [CrossRef]
- Park, J.H.; Lee, B.M.; Kim, H.S. Potential protective roles of curcumin against cadmium-induced toxicity and oxidative stress. J. Toxiocol. Environ. Health Part B Crit. Rev. 2020, 24, 95–118. [Google Scholar] [CrossRef] [PubMed]
- Yadav, S.; Teng, P.; Souza Dos Santos, T.; Gould, R.; Craig, S.; Lorraine Fuller, A.; Pazdro, R.; Kim, W. The effects of different doses of curcumin compound on growth performance, antioxidant status, and gut health of broiler chickens challenged with Eimeria species. Poult. Sci. 2020, 99, 5936–5945. [Google Scholar] [CrossRef]
- Daneshyar, M. Effect of dietary turmeric on antioxidant properties of thigh meat in broiler chickens after slaughter. Anim. Sci. J. 2012, 83, 599–604. [Google Scholar] [CrossRef]
- Leyva-Diaz, A.; Hernandez-Patlan, D.; Solis-Cruz, B.; Adhikari, B.; Kwon, Y.; Latorre, J.; Hernandez-Velasco, X.; Fuente-Martinez, B.; Hargis, B.; Lopez-Arellano, R.; et al. Evaluation of curcumin and copper acetate against Salmonella typhimurium infection, intestinal permeability, and cecal microbiota composition in broiler chickens. J. Anim. Sci. Biotechnol. 2021, 12, 23. [Google Scholar] [CrossRef]
- Liu, X.; Guan, P.; Yu, C.; Yang, H.; Shan, A.; Feng, X. Curcumin alleviated lipopolysaccharide-induced lung injury via regulating the Nrf2-ARE and NF-κB signaling pathways in ducks. J. Sci. Food Agric. 2022, 102, 6603–6611. [Google Scholar] [CrossRef]
- Nasir, R.; Naeem, M.; Yan, R.; Xiang, Z.; Tian, W. Effect of Dietary Supplementation of Curcumin on Growth Performance, Intestinal Morphology and Nutrients Utilization of Broiler Chicks. J. Poult. Sci. 2013, 50, 44–52. [Google Scholar] [CrossRef]
- Sarker, M.; Wan, X.; Yang, H.; Wang, Z. AflatoxinB (AFB) and its toxic effect on the broilers intestine: A review. Vet. Med. Sci. 2023, 9, 1646–1655. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.; Ibtisham, F.; Niu, Y.; Wang, Z.; Li, G.; Zhao, Y.; Nawab, A.; Xiao, M.; An, L. Curcumin inhibits heat-induced oxidative stress by activating the MAPK-Nrf2/ARE signaling pathway in chicken fibroblasts cells. J. Therm. Biol. 2019, 79, 112–119. [Google Scholar] [CrossRef] [PubMed]
- Liu, M.; Lu, Y.; Gao, P.; Xie, X.; Li, D.; Yu, D.; Yu, M. Effect of curcumin on laying performance, egg quality, endocrine hormones, and immune activity in heat-stressed hens. Poult. Sci. 2020, 99, 2196–2202. [Google Scholar] [CrossRef] [PubMed]
- Nawab, A.; Tang, S.; Li, G.; An, L.; Wu, J.; Liu, W.; Xiao, M. Dietary curcumin supplementation effects on blood immunological profile and liver enzymatic activity of laying hens after exposure to high temperature conditions. J. Therm. Biol. 2020, 90, 102573. [Google Scholar] [CrossRef] [PubMed]
- Nawab, A.; Li, G.; An, L.; Wu, J.; Chao, L.; Xiao, M.; Zhao, Y.; Birmani, M.; Ghani, M. Effect of curcumin supplementation on TLR4 mediated non-specific immune responses in liver of laying hens under high-temperature conditions. J. Therm. Biol. 2019, 84, 384–397. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Liu, R.; Wei, G.; Guo, G.; Yu, H.; Zhang, Y.; Ishfaq, M.; Fazilani, S.; Zhang, X. Curcumin protects against Aflatoxin B1-induced liver injury in broilers via the modulation of long non-coding RNA expression. Ecotoxicol. Environ. Saf. 2021, 208, 111725. [Google Scholar] [CrossRef]
- American Veterinary Medical Association. AVMA Guidelines for the Euthanasia of Animals; American Veterinary Medical Association: Schaumburg, IL, USA, 2020. [Google Scholar]
- Trebak, F.; Alaoui, A.; Alexandre, D.; El Ouezzani, S.; Anouar, Y.; Chartrel, N.; Magoul, R. Impact of aflatoxin B1 on hypothalamic neuropeptides regulating feeding behavior. Neurotoxicology 2015, 49, 165–173. [Google Scholar] [CrossRef]
- Szkudelska, K.; Drzymała, H.; Szkudelski, T.; Bukowska, K.; Nogowski, L. Lack of the effect of mycotoxins-aflatoxin B1 and ochratoxin A on some functions of rat adipocytes. Toxicol. In Vitro 2005, 19, 771–777. [Google Scholar] [CrossRef]
- Wang, T.; Li, X.; Liao, G.; Wang, Z.; Han, X.; Gu, J.; Mu, X.; Qiu, J.; Qian, Y. AFB1 Triggers Lipid Metabolism Disorders through the PI3K/Akt Pathway and Mediates Apoptosis Leading to Hepatotoxicity. Foods 2024, 13, 163. [Google Scholar] [CrossRef]
- Osborne, D.; Huff, W.; Hamilton, P.; Burmeister, H. Comparison of ochratoxin, aflatoxin, and T-2 toxin for their effects on selected parameters related to digestion and evidence for specific metabolism of carotenoids in chickens. Poult. Sci. 1982, 61, 1646–1652. [Google Scholar] [CrossRef]
- Santos, R.; Velkers, F.; Vernooij, J.; Star, L.; Heerkens, J.; van Harn, J.; de Jong, I. Nutritional interventions to support broiler chickens during Eimeria infection. Poult. Sci. 2022, 101, 101853. [Google Scholar] [CrossRef]
- Hafez, M.; El-Kazaz, S.; Alharthi, B.; Ghamry, H.; Alshehri, M.; Sayed, S.; Shukry, M.; El-Sayed, Y. The Impact of Curcumin on Growth Performance, Growth-Related Gene Expression, Oxidative Stress, and Immunological Biomarkers in Broiler Chickens at Different Stocking Densities. Animals 2022, 12, 958. [Google Scholar] [CrossRef]
- Shannon, T.; Ledoux, D.; Rottinghaus, G.; Shaw, D.; Dakovic, A.; Markovic, M. The efficacy of raw and concentrated bentonite clay in reducing the toxic effects of aflatoxin in broiler chicks. Poult. Sci. 2017, 96, 1651–1658. [Google Scholar] [CrossRef]
- Ke, P. Diverse Functions of Autophagy in Liver Physiology and Liver Diseases. Int. J. Mol. Sci. 2019, 20, 300. [Google Scholar] [CrossRef]
- Ali Rajput, S.; Sun, L.; Zhang, N.; Mohamed Khalil, M.; Gao, X.; Ling, Z.; Zhu, L.; Khan, F.; Zhang, J.; Qi, D. Ameliorative Effects of Grape Seed Proanthocyanidin Extract on Growth Performance, Immune Function, Antioxidant Capacity, Biochemical Constituents, Liver Histopathology and Aflatoxin Residues in Broilers Exposed to Aflatoxin B1. Toxins 2017, 9, 371. [Google Scholar] [CrossRef] [PubMed]
- Cheung, E.; Vousden, K. The role of ROS in tumour development and progression. Nat. Rev. Cancer 2022, 22, 280–297. [Google Scholar] [CrossRef] [PubMed]
- Salem, R.; El-Habashi, N.; Fadl, S.; Sakr, O.; Elbialy, Z. Effect of probiotic supplement on aflatoxicosis and gene expression in the liver of broiler chicken. Environ. Toxicol. Pharmacol. 2018, 60, 118–127. [Google Scholar] [CrossRef] [PubMed]
- Cheng, X.; Liang, J.; Wu, D.; Guo, X.; Cao, H.; Zhang, C.; Liu, P.; Hu, R.; Hu, G.; Zhuang, Y. Blunting ROS/TRPML1 pathway protects AFB1-induced porcine intestinal epithelial cells apoptosis by restoring impaired autophagic flux. Ecotoxicol. Environ. Saf. 2023, 257, 114942. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Liu, T.; Koci, M.; Wang, Y.; Fu, Y.; Ma, M.; Ma, Q.; Zhao, L. Chlorogenic Acid Alleviated AFB1-Induced Hepatotoxicity by Regulating Mitochondrial Function, Activating Nrf2/HO-1, and Inhibiting Noncanonical NF-κB Signaling Pathway. Antioxidants 2023, 12, 2027. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Zhou, X.; Liu, M.; Zang, H.; Zhang, R.; Yang, H.; Jin, S.; Qi, X.; Shan, A.; Feng, X. Quality of chicken breast meat improved by dietary pterostilbene referring to up-regulated antioxidant capacity and enhanced protein structure. Food Chem. 2023, 405, 134848. [Google Scholar] [CrossRef]
- Ulasov, A.; Rosenkranz, A.; Georgiev, G.; Sobolev, A. Nrf2/Keap1/ARE signaling: Towards specific regulation. Life Sci. 2022, 291, 120111. [Google Scholar] [CrossRef]
- Miyauchi, E.; Morita, H.; Tanabe, S. Lactobacillus rhamnosus alleviates intestinal barrier dysfunction in part by increasing expression of zonula occludens-1 and myosin light-chain kinase in vivo. J. Dairy Sci. 2009, 92, 2400–2408. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Zhu, H. Curcumin Improved Intestinal Epithelial Barrier Integrity by Up-Regulating ZO-1/Occludin/Claudin-1 in Septic Rats. Evid.-Based Complement. Altern. Med. ECAM 2022, 2022, 2884522. [Google Scholar] [CrossRef]
- Jin, S.; Yang, H.; Jiao, Y.; Pang, Q.; Wang, Y.; Wang, M.; Shan, A.; Feng, X. Anas platyrhynchos Dietary Curcumin Alleviated Acute Ileum Damage of Ducks (Anas platyrhynchos) Induced by AFB1 through Regulating Nrf2-ARE and NF-κB Signaling Pathways. Foods 2021, 10, 1370. [Google Scholar] [CrossRef] [PubMed]
- Goodarzi, N.; Akbari Bazm, M.; Poladi, S.; Rashidi, F.; Mahmoudi, B.; Abumandour, M. Histology of the small intestine in the common pheasant (Phasianus colchicus): A scanning electron microscopy, histochemical, immunohistochemical, and stereological study. Microsc. Res. Tech. 2021, 84, 2388–2398. [Google Scholar] [CrossRef] [PubMed]
- Robert, H.; Payros, D.; Pinton, P.; Théodorou, V.; Mercier-Bonin, M.; Oswald, I. Impact of mycotoxins on the intestine: Are mucus and microbiota new targets? J. Toxicol. Environ. Health Part B Crit. Rev. 2017, 20, 249–275. [Google Scholar] [CrossRef]
- Hamai, Y.; Hihara, J.; Emi, M.; Ibuki, Y.; Kurokawa, T.; Yoshikawa, T.; Hirohata, R.; Ohsawa, M.; Kitasaki, N.; Okada, M. Prospective Randomized Trial of Early Postoperative Enteral and Total Parenteral Nutrition for Treating Esophageal Cancer. Anticancer Res. 2021, 41, 6237–6246. [Google Scholar] [CrossRef]
- Vötterl, J.; Klinsoda, J.; Zebeli, Q.; Hennig-Pauka, I.; Kandler, W.; Metzler-Zebeli, B. Dietary Phytase and Lactic Acid-Treated Cereal Grains Differently Affected Calcium and Phosphorus Homeostasis from Intestinal Uptake to Systemic Metabolism in a Pig Model. Nutrients 2020, 12, 1542. [Google Scholar] [CrossRef]
- Diaz de Barboza, G.; Guizzardi, S.; Moine, L.; Tolosa de Talamoni, N. Oxidative stress, antioxidants and intestinal calcium absorption. World J. Gastroenterol. 2017, 23, 2841–2853. [Google Scholar] [CrossRef]
- Singh, R.; Chandrashekharappa, S.; Bodduluri, S.; Baby, B.; Hegde, B.; Kotla, N.; Hiwale, A.; Saiyed, T.; Patel, P.; Vijay-Kumar, M.; et al. Enhancement of the gut barrier integrity by a microbial metabolite through the Nrf2 pathway. Nat. Commun. 2019, 10, 89. [Google Scholar] [CrossRef] [PubMed]
- Yang, H.; Yu, C.; Yin, Z.; Guan, P.; Jin, S.; Wang, Y.; Feng, X. Curcumin: A potential exogenous additive for the prevention of LPS-induced duck ileitis by the alleviation of inflammation and oxidative stress. J. Sci. Food Agric. 2023, 103, 1550–1560. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ingredients, % | Days 7 to 21 | Days 22 to 28 |
|---|---|---|
| Corn | 58.5 | 61.15 |
| Soybean meal | 30.0 | 26.3 |
| Corn protein meal | 4.06 | 4.33 |
| Soybean oil | 2.70 | 3.80 |
| Limestone | 1.33 | 1.26 |
| Calcium bicarbonate | 1.60 | 1.52 |
| L-Lysine, 99% | 0.20 | 0.14 |
| DL-methionine, 98% | 0.21 | 0.10 |
| Sodium chloride | 0.30 | 0.30 |
| Choline chloride | 0.10 | 0.10 |
| Premix 1 | 1.00 | 1.00 |
| Nutrient levels | ||
| ME (MJ/kg) | 12.54 | 12.96 |
| Crude protein (%) | 21.50 | 20.09 |
| Calcium (%) | 1.06 | 0.91 |
| Total phosphorus (%) | 0.73 | 0.69 |
| Effective phosphorus | 0.45 | 0.43 |
| Lys (%) | 1.15 | 1.01 |
| Met (%) | 0.55 | 0.43 |
| Met+ Cys (%) | 0.91 | 0.77 |
| Thr (%) | 0.80 | 0.73 |
| Genes | Primer Sequence (5′ → 3′) | Product Size (bp) | Accession Number |
|---|---|---|---|
| Nrf2 | F: GGGACGGTGACACAGGAACAAC | 93 | XM_046921130.1 |
| R: TCCACAGCGGGAAATCAGAAAGATC | |||
| Keap1 | F: CGCTTTCTTCAGGGGTAGCA | 170 | NM_205117.2 |
| R: AGTTCGGTGCAGAAGAGGTG | |||
| HO-1 | F: GCTGGGAAGGAGAGTGAGAGGAC | 107 | XM_046921508.1 |
| R: GCGACTGTGGTGGCGATGAAG | |||
| NQO1 | F: CGAGTGCTTTGTCTACGAGATGGAG | 102 | NM_001277620.2 |
| R: AGGTCAGCCGCTTCAATCTTCTTC | |||
| β-actin | F: CTGTGCCCATCTATGAAGGCTA | 139 | NM_205518.2 |
| R: ATTTCTCTCTCGGCTGTGGTG |
| Items | Treatments | SEM | p-Value | |||
|---|---|---|---|---|---|---|
| Control | AFB1 | AFB1+CUR | CUR | |||
| 1 D BW (g) | 38.50 | 38.50 | 37.50 | 38.67 | 0.333 | 0.355 |
| 28 D BW (g) | 936.82 b | 714.14 d | 849.00c | 1001.59 a | 22.663 | <0.001 |
| 1-28D | ||||||
| ADG (g/bird) | 32.08 b | 24.13 d | 29.00 c | 34.39 a | 0.808 | <0.001 |
| ADFI (g/bird) | 48.60 ab | 46.85 c | 48.14 bc | 50.08 a | 0.342 | 0.003 |
| F/G (g/g) | 1.52 c | 1.94 a | 1.66 b | 1.46 d | 0.040 | <0.001 |
| Items | Treatments | SEM | p-Value | |||
|---|---|---|---|---|---|---|
| Control | AFB1 | AFB1+CUR | CUR | |||
| Heart weight (g) | 5.91 a | 4.61 b | 5.63 a | 63.88 a | 0.179 | 0.001 |
| Heart index (%) | 0.64 | 0.69 | 0.61 | 0.63 | 0.016 | 0.318 |
| Liver weight (g) | 20.66 a | 15.93 b | 21.13 a | 20.46 a | 0.546 | <0.001 |
| Liver index (%) | 2.23 b | 2.44 a | 2.28 ab | 2.15 b | 0.038 | 0.042 |
| Spleen weight (g) | 0.99 | 0.84 | 1.01 | 0.98 | 0.042 | 0.473 |
| Spleen index (%) | 0.11 | 0.13 | 0.11 | 0.10 | 0.005 | 0.284 |
| Lungs weight (g) | 6.20 a | 4.20 b | 5.80 a | 5.89 a | 0.210 | <0.001 |
| Lungs index (%) | 0.67 | 0.62 | 0.63 | 0.59 | 0.017 | 0.358 |
| Kidney weight (g) | 5.20 | 4.56 | 5.46 | 5.44 | 0.223 | 0.468 |
| Kidney index (%) | 0.57 | 0.68 | 0.59 | 0.54 | 0.028 | 0.286 |
| Pancreas weight (g) | 2.73 a | 1.93 b | 2.65 a | 2.63 a | 0.083 | <0.001 |
| Pancreas index (%) | 0.30 | 0.29 | 0.29 | 0.26 | 0.007 | 0.345 |
| Thymus weight (g) | 2.95 | 1.87 | 3.03 | 3.31 | 0.056 | 0.003 |
| Thymus index (%) | 0.32 | 0.27 | 0.33 | 0.32 | 0.014 | 0.438 |
| Bursa of Fabricius weight (g) | 1.85 a | 1.00 b | 1.76 a | 1.88 a | 1.846 | 0.005 |
| Bursa of Fabricius index (%) | 0.20 | 0.15 | 0.19 | 0.19 | 0.203 | 0.371 |
| Items | Treatments | SEM | p-Value | |||
|---|---|---|---|---|---|---|
| Control | AFB1 | AFB1+CUR | CUR | |||
| Intestinal weight and relative weight | ||||||
| Intestine weight (g) | 45.27 a | 35.30 b | 45.47 a | 48.17 a | 1.273 | <0.001 |
| Intestine index (%) | 4.91 | 5.08 | 4.92 | 4.73 | 0.085 | 0.562 |
| Relative length(cm/kg·BW) | ||||||
| Duodenum relative length | 19.73 a | 17.79 b | 18.27 b | 19.75 a | 0.239 | <0.001 |
| Jejunum relative length | 54.69 a | 47.01 b | 51.44 ab | 54.45 a | 1.11 | 0.038 |
| Ileum relative length | 54.07 ab | 47.53 c | 51.33 b | 55.79 a | 0.87 | <0.001 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, J.; Sun, X.; Chai, X.; Jiao, Y.; Sun, J.; Wang, S.; Yu, H.; Feng, X. Curcumin Mitigates Oxidative Damage in Broiler Liver and Ileum Caused by Aflatoxin B1-Contaminated Feed through Nrf2 Signaling Pathway. Animals 2024, 14, 409. https://doi.org/10.3390/ani14030409
Zhang J, Sun X, Chai X, Jiao Y, Sun J, Wang S, Yu H, Feng X. Curcumin Mitigates Oxidative Damage in Broiler Liver and Ileum Caused by Aflatoxin B1-Contaminated Feed through Nrf2 Signaling Pathway. Animals. 2024; 14(3):409. https://doi.org/10.3390/ani14030409
Chicago/Turabian StyleZhang, Jingyang, Xue Sun, Xuehong Chai, Yihan Jiao, Jing Sun, Shenao Wang, Hao Yu, and Xingjun Feng. 2024. "Curcumin Mitigates Oxidative Damage in Broiler Liver and Ileum Caused by Aflatoxin B1-Contaminated Feed through Nrf2 Signaling Pathway" Animals 14, no. 3: 409. https://doi.org/10.3390/ani14030409
APA StyleZhang, J., Sun, X., Chai, X., Jiao, Y., Sun, J., Wang, S., Yu, H., & Feng, X. (2024). Curcumin Mitigates Oxidative Damage in Broiler Liver and Ileum Caused by Aflatoxin B1-Contaminated Feed through Nrf2 Signaling Pathway. Animals, 14(3), 409. https://doi.org/10.3390/ani14030409