
NOTCH1 as a Negative Regulator of Avian Adipocyte Differentiation: Implications for Fat Deposition

Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Cell Culture
2.2. Construction, Amplification, and Purification of Plasmids
2.3. RNA Interference
2.4. Transfections
2.5. DAPT Treatment
2.6. Adipogenic Differentiation of ICP1 Cells
2.7. Oil Red O Staining and Quantification
2.8. RNA Extraction, cDNA Synthesis, and RT-qPCR Assay
2.9. Cell Fractionation Assay
2.10. Western Blot Analysis
2.11. Transcription Factor Binding Sites Prediction
2.12. Dual-Luciferase Reporter Assay
2.13. RNA-seq Analysis
2.14. Statistical Analysis
3. Results
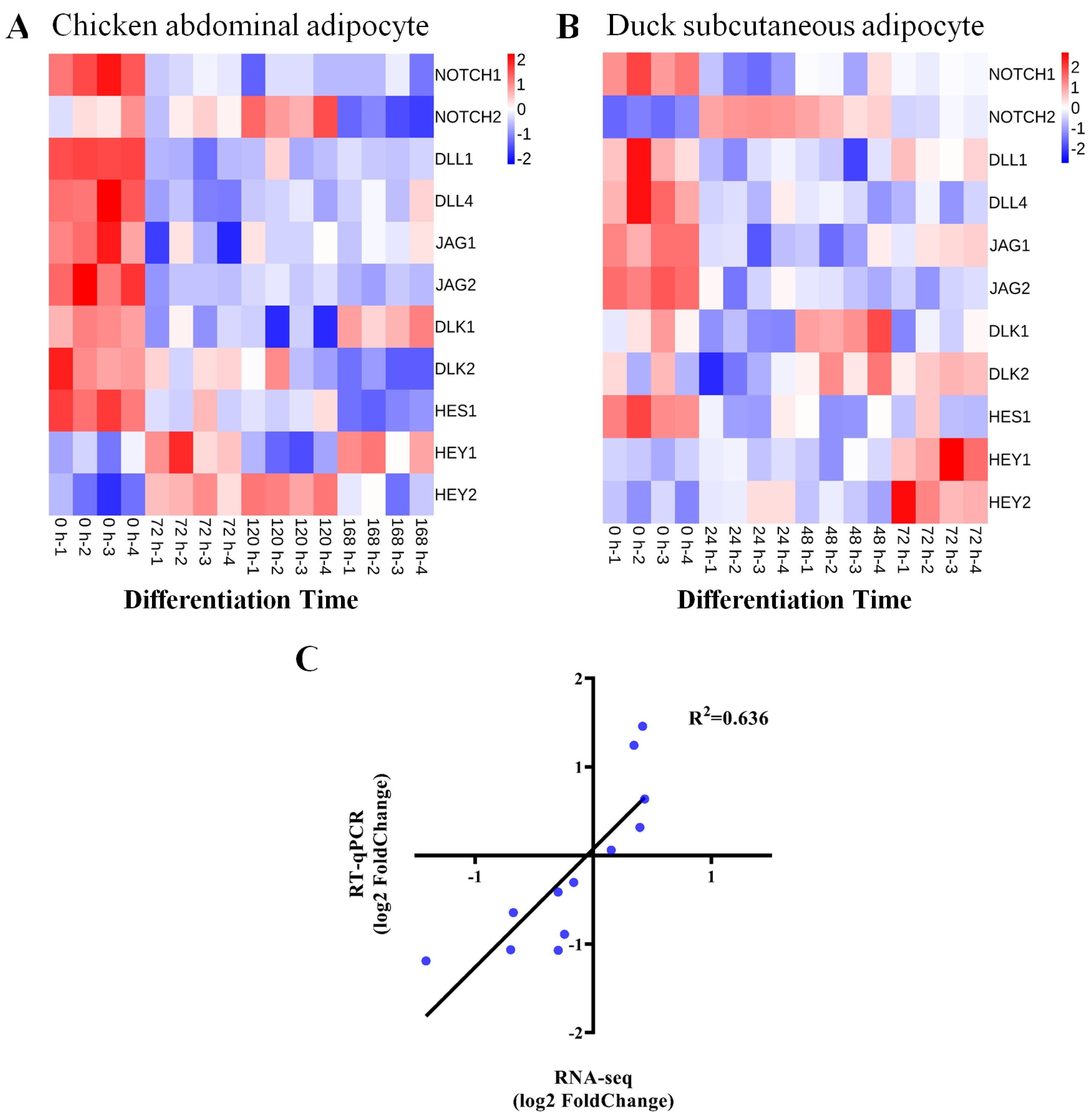
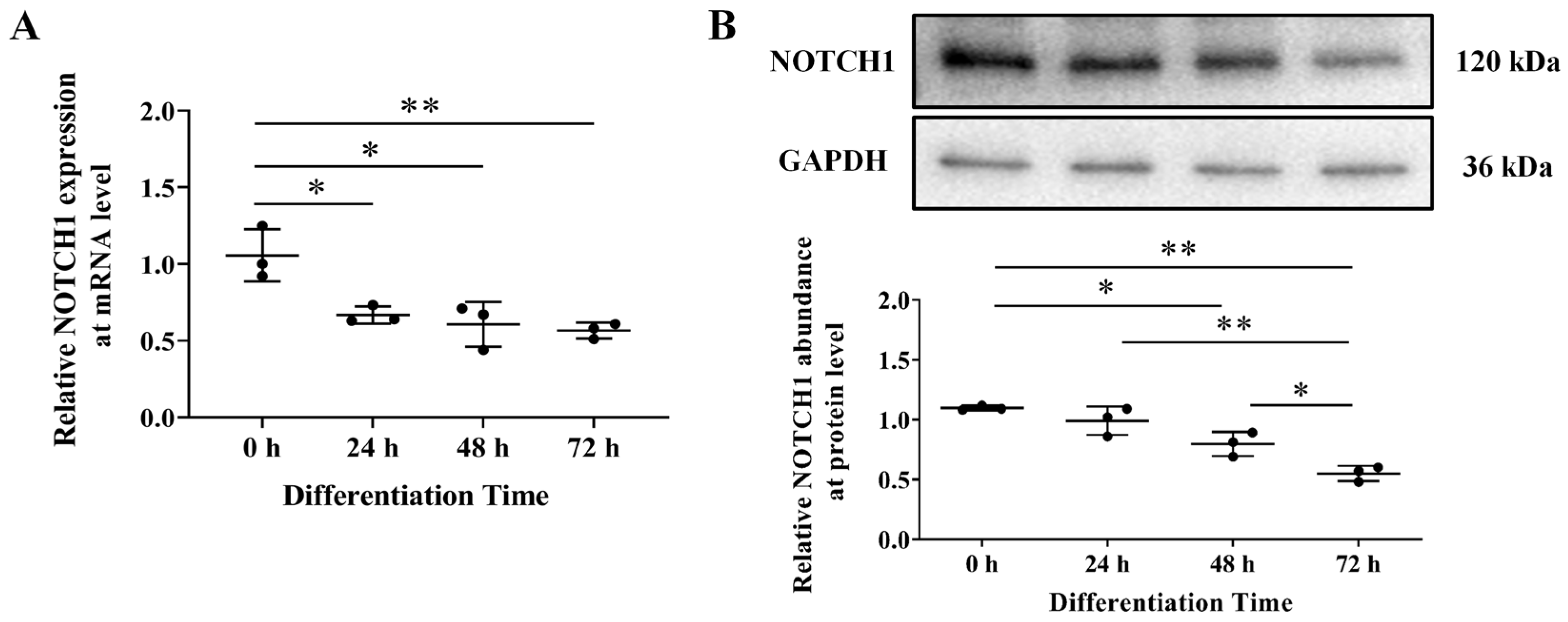
3.1. NOTCH1 Expression during Avian Adipocyte Differentiation
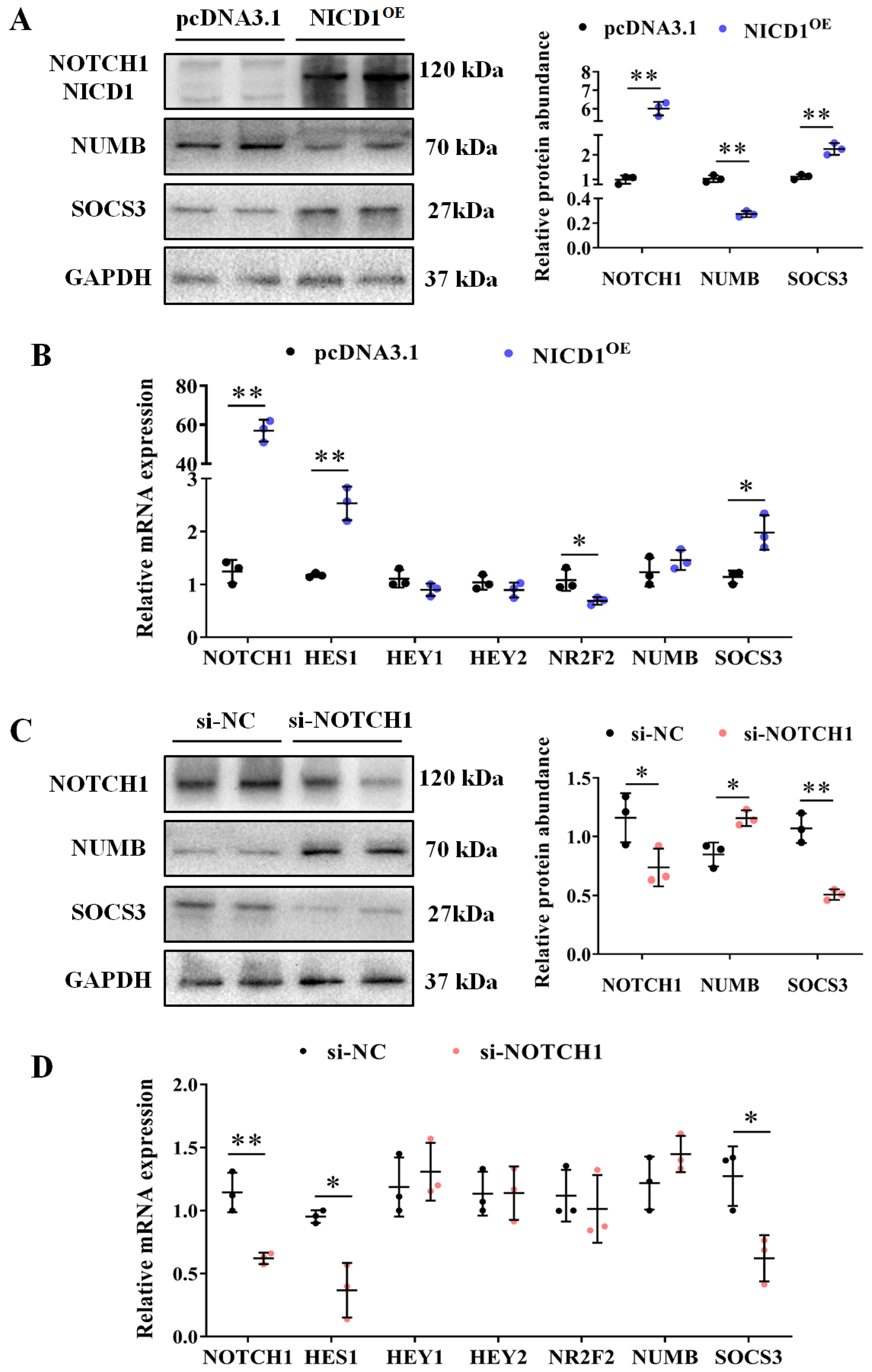
3.2. NOTCH1 Regulates Expression of Signaling-Related Genes
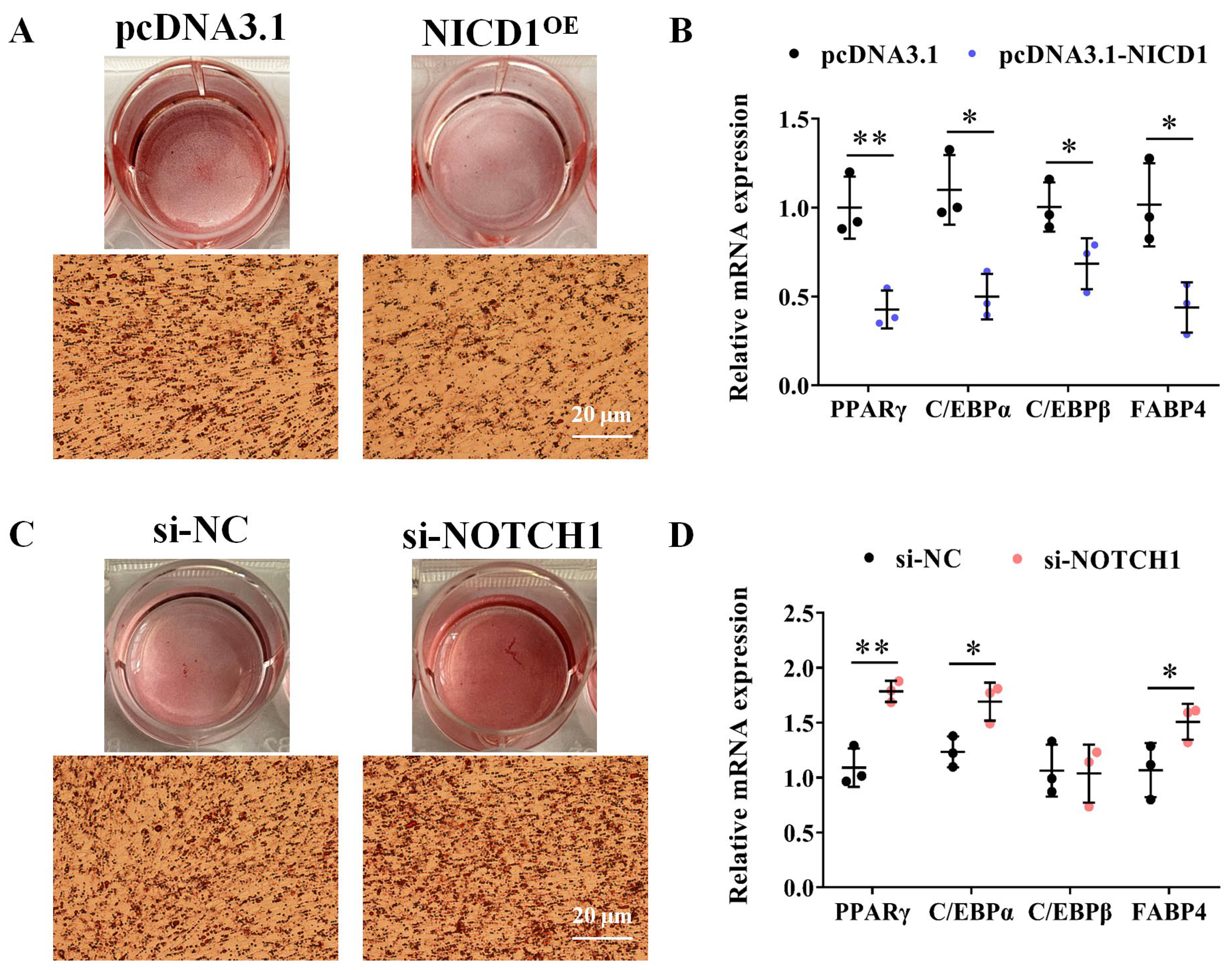
3.3. NOTCH1 Inhibits Avian Adipocyte Differentiation
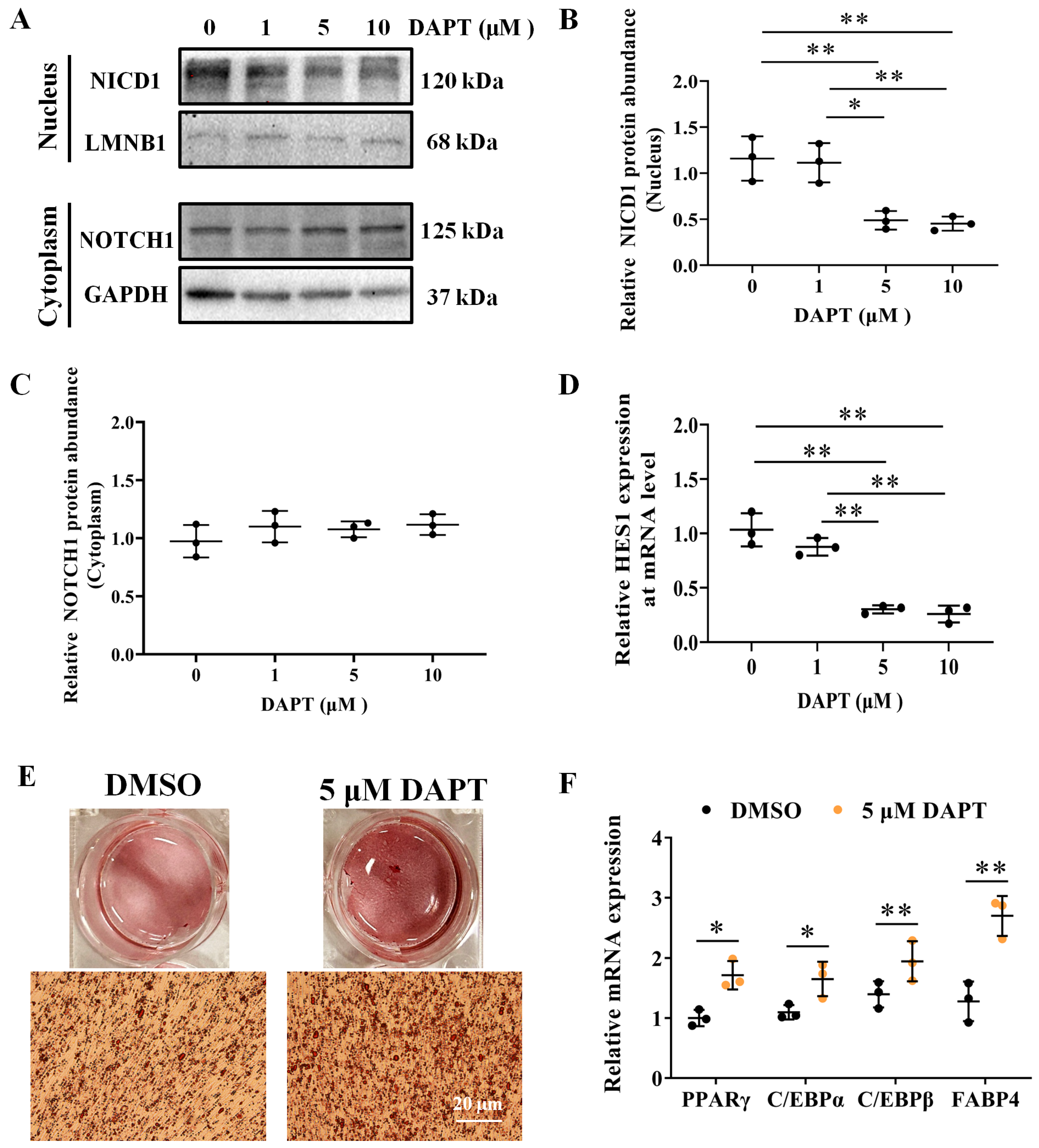
3.4. DAPT, an Inhibitor of NOTCH Signaling, Promotes Adipocyte Differentiation
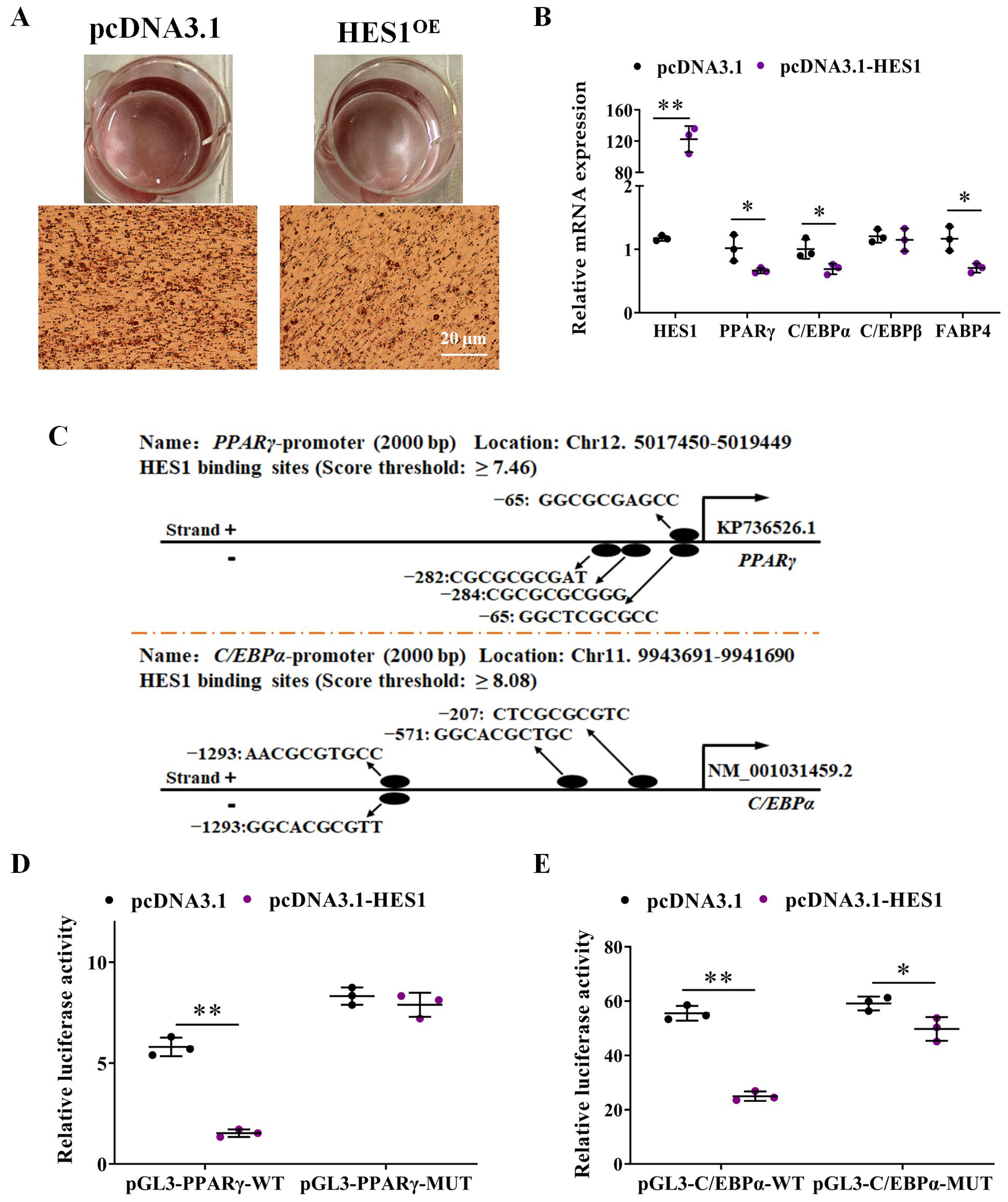
3.5. HES1 Inhibits the Expression of PPARγ and C/EBPα by Directly Binding to Their Upstream Region
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Baeza, E.; Guillier, L.; Petracci, M. Review: Production factors affecting poultry carcass and meat quality attributes. Animal 2018, 16 (Suppl. 1), 100331. [Google Scholar] [CrossRef]
- Mathew, H.; Castracane, V.D.; Mantzoros, C. Adipose tissue and reproductive health. Metabolism 2018, 86, 18–32. [Google Scholar] [CrossRef]
- Wang, G.; Kim, W.K.; Cline, M.A.; Gilbert, E.R. Factors affecting adipose tissue development in chickens: A review. Poult. Sci. 2017, 96, 3687–3699. [Google Scholar] [CrossRef]
- Ghaben, A.L.; Scherer, P.E. Adipogenesis and metabolic health. Nat. Rev. Mol. Cell Biol. 2019, 20, 242–258. [Google Scholar] [CrossRef]
- Schwalie, P.C.; Dong, H.; Zachara, M.; Russeil, J.; Alpern, D.; Akchiche, N.; Caprara, C.; Sun, W.; Schlaudraff, K.U.; Soldati, G.; et al. A stromal cell population that inhibits adipogenesis in mammalian fat depots. Nature 2018, 559, 103–108. [Google Scholar] [CrossRef] [PubMed]
- Mu, F.; Jing, Y.; Ning, B.; Huang, J.; Cui, T.; Guo, Y.; You, X.; Yan, X.; Li, H.; Wang, N. Peroxisome proliferator-activated receptor gamma isoforms differentially regulate preadipocyte proliferation, apoptosis, and differentiation in chickens. Poult. Sci. 2020, 99, 6410–6421. [Google Scholar] [CrossRef] [PubMed]
- Sun, D.; Li, X.; Yin, Z.; Hou, Z. The Full-Length Transcriptome Provides New Insights into the Transcript Complexity of Abdominal Adipose and Subcutaneous Adipose in Pekin Ducks. Front. Physiol. 2021, 12, 767739. [Google Scholar] [CrossRef] [PubMed]
- Donati, G.; Proserpio, V.; Lichtenberger, B.M.; Natsuga, K.; Sinclair, R.; Fujiwara, H.; Watt, F.M. Epidermal Wnt/beta-catenin signaling regulates adipocyte differentiation via secretion of adipogenic factors. Proc. Natl. Acad. Sci. USA 2014, 111, E1501–E1509. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Zhao, Q.; Li, X.; Yin, Z.; Chen, S.; Wu, S.; Yang, N.; Hou, Z. MYOD1 inhibits avian adipocyte differentiation via miRNA-206/KLF4 axis. J. Anim. Sci. Biotechnol. 2021, 12, 55. [Google Scholar] [CrossRef]
- Wang, Z.; Yin, Z.T.; Zhang, F.; Li, X.Q.; Chen, S.R.; Yang, N.; Porter, T.E.; Hou, Z. Dynamics of transcriptome changes during subcutaneous preadipocyte differentiation in ducks. BMC Genom. 2019, 20, 688. [Google Scholar] [CrossRef] [PubMed]
- Zohorsky, K.; Mequanint, K. Designing Biomaterials to Modulate Notch Signaling in Tissue Engineering and Regenerative Medicine. Tissue Eng. Part B Rev. 2021, 27, 383–410. [Google Scholar] [CrossRef] [PubMed]
- Kopan, R.; Ilagan, M.X. The canonical Notch signaling pathway. unfolding the activation mechanism. Cell 2009, 137, 216–233. [Google Scholar] [CrossRef] [PubMed]
- Meurette, O.; Mehlen, P. Notch Signaling in the Tumor Microenvironment. Cancer Cell 2018, 34, 536–548. [Google Scholar] [CrossRef] [PubMed]
- Zhou, B.; Lin, W.; Long, Y.; Yang, Y.; Zhang, H.; Wu, K.; Chu, Q. Notch signaling pathway: Architecture, disease, and therapeutics. Signal Transduct. Target. Ther. 2022, 7, 95. [Google Scholar] [CrossRef]
- Singh, S.B.; Coffman, C.N.; Carroll-Portillo, A.; Varga, M.G.; Lin, H.C. Notch Signaling Pathway Is Activated by Sulfate Reducing Bacteria. Front. Cell. Infect. Microbiol. 2021, 11, 695299. [Google Scholar] [CrossRef]
- Chen, X.; Qin, J.; Cheng, C.M.; Tsai, M.J.; Tsai, S.Y. COUP-TFII is a major regulator of cell cycle and Notch signaling pathways. Mol. Endocrinol. 2012, 26, 1268–1277. [Google Scholar] [CrossRef]
- Miao, L.; Lu, Y.; Nusrat, A.; Abdelnasser, H.Y.; Datta, S.; Zhou, B.; Schwartz, R.J.; Wu, M. The Spatiotemporal Expression of Notch1 and Numb and Their Functional Interaction during Cardiac Morphogenesis. Cells 2021, 10, 2192. [Google Scholar] [CrossRef]
- Esteves de Lima, J.; Bonnin, M.A.; Birchmeier, C.; Duprez, D. Muscle contraction is required to maintain the pool of muscle progenitors via YAP and NOTCH during fetal myogenesis. eLife 2016, 5, e15593. [Google Scholar] [CrossRef]
- Zuo, Q.; Zhang, C.; Jin, K.; Jing, J.; Sun, C.; Ahmed, M.F.; Song, J.; Zhang, Y.; Chen, G.; Li, B. NICD-mediated notch transduction regulates the different fate of chicken primordial germ cells and spermatogonial stem cells. Cell Biosci. 2018, 8, 40. [Google Scholar] [CrossRef]
- Ross, D.A.; Rao, P.K.; Kadesch, T. Dual roles for the Notch target gene Hes-1 in the differentiation of 3T3-L1 preadipocytes. Mol. Cell. Biol. 2004, 24, 3505–3513. [Google Scholar] [CrossRef] [PubMed]
- Nichols, A.M.; Pan, Y.; Herreman, A.; Hadland, B.K.; De Strooper, B.; Kopan, R.; Huppert, S.S. Notch pathway is dispensable for adipocyte specification. Genesis 2004, 40, 40–44. [Google Scholar] [CrossRef]
- Song, B.Q.; Chi, Y.; Li, X.; Du, W.J.; Han, Z.B.; Tian, J.J.; Li, J.J.; Chen, F.; Wu, H.H.; Han, L.X.; et al. Inhibition of Notch Signaling Promotes the Adipogenic Differentiation of Mesenchymal Stem Cells Through Autophagy Activation and PTEN-PI3K/AKT/mTOR Pathway. Cell. Physiol. Biochem. 2015, 36, 1991–2002. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Zhang, T.; Wu, C.; Wang, S.; Wang, Y.; Li, H.; Wang, N. Immortalization of chicken preadipocytes by retroviral transduction of chicken TERT and TR. PLoS ONE 2017, 12, e0177348. [Google Scholar] [CrossRef] [PubMed]
- Ross, D.A.; Kadesch, T. The notch intracellular domain can function as a coactivator for LEF-1. Mol. Cell. Biol. 2001, 21, 7537–7544. [Google Scholar] [CrossRef]
- Dovey, H.F.; John, V.; Anderson, J.P.; Chen, L.Z.; de Saint Andrieu, P.; Fang, L.Y.; Freedman, S.B.; Folmer, B.; Goldbach, E.; Holsztynska, E.J.; et al. Functional gamma-secretase inhibitors reduce beta-amyloid peptide levels in brain. J. Neurochem. 2001, 76, 173–181. [Google Scholar] [CrossRef]
- Shang, Z.; Guo, L.; Wang, N.; Shi, H.; Wang, Y.; Li, H. Oleate promotes differentiation of chicken primary preadipocytes in vitro. Biosci. Rep. 2014, 34, e00093. [Google Scholar] [CrossRef] [PubMed]
- Ramirez-Zacarias, J.L.; Castro-Munozledo, F.; Kuri-Harcuch, W. Quantitation of adipose conversion and triglycerides by staining intracytoplasmic lipids with Oil red O. Histochemistry 1992, 97, 493–497. [Google Scholar] [CrossRef]
- Kurien, B.T.; Scofield, R.H. Western blotting. Methods 2006, 38, 283–293. [Google Scholar] [CrossRef]
- Castro-Mondragon, J.A.; Riudavets-Puig, R.; Rauluseviciute, I.; Lemma, R.B.; Turchi, L.; Blanc-Mathieu, R.; Lucas, J.; Boddie, P.; Khan, A.; Manosalva Perez, N.; et al. JASPAR 2022: The 9th release of the open-access database of transcription factor binding profiles. Nucleic Acids Res. 2022, 50, D165–D173. [Google Scholar] [CrossRef]
- Kim, D.; Paggi, J.M.; Park, C.; Bennett, C.; Salzberg, S.L. Graph-based genome alignment and genotyping with HISAT2 and HISAT-genotype. Nat. Biotechnol. 2019, 37, 907–915. [Google Scholar] [CrossRef]
- Anders, S.; Pyl, P.T.; Huber, W. HTSeq—A Python framework to work with high-throughput sequencing data. Bioinformatics 2015, 31, 166–169. [Google Scholar] [CrossRef] [PubMed]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; He, C.; Ran, L.; Wang, Y.; Xiong, Y.; Wang, Y.; Zhu, J.; Lin, Y. miR-130b duplex (miR-130b-3p/miR-130b-5p) negatively regulates goat intramuscular preadipocyte lipid droplets accumulation by inhibiting Kruppel-like factor 3 expression. J. Anim. Sci. 2023, 101, skad184. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Tan, J.; Du, H.; Liu, X.; Wang, S.; Wu, S.; Yuan, Z.; Zhu, X. Notch1 Inhibits Rosiglitazone-Induced Adipogenic Differentiation in Primary Thymic Stromal Cells. Front. Pharmacol. 2018, 9, 1284. [Google Scholar] [CrossRef] [PubMed]
- Lei, T.; Bi, Y.; Gao, M.J.; Gao, S.M.; Zhou, L.L.; Zheng, H.L.; Chen, X.D. HES1 inhibits adipogenesis of porcine mesenchymal stem cells via transcriptional repression of FAD24. Domest. Anim. Endocrinol. 2013, 45, 28–32. [Google Scholar] [CrossRef] [PubMed]
- Duan, K.; Sun, Y.; Zhang, X.; Zhang, T.; Zhang, W.; Zhang, J.; Wang, G.; Wang, S.; Leng, L.; Li, H.; et al. Identification and characterization of transcript variants of chicken peroxisome proliferator-activated receptor gamma. Poult. Sci. 2015, 94, 2516–2527. [Google Scholar] [CrossRef] [PubMed]
- Jing, Y.; Mu, F.; Xing, X.; Huang, J.; Lou, M.; Xu, H.; Ning, B.; Lou, Y.; Gao, Z.; Luo, H.; et al. Knockout and Restoration Reveal Differential Functional Roles of PPARgamma1 and PPARgamma2 in Chicken Adipogenesis. J. Agric. Food Chem. 2022, 70, 14959–14973. [Google Scholar] [CrossRef]
- Aranguren, X.L.; Beerens, M.; Coppiello, G.; Wiese, C.; Vandersmissen, I.; Lo Nigro, A.; Verfaillie, C.M.; Gessler, M.; Luttun, A. COUP-TFII orchestrates venous and lymphatic endothelial identity by homo- or hetero-dimerisation with PROX1. J. Cell Sci. 2013, 126 Pt 5, 1164–1175. [Google Scholar] [CrossRef]
- Zhu, F.; Yin, Z.T.; Wang, Z.; Smith, J.; Zhang, F.; Martin, F.; Ogeh, D.; Hincke, M.; Lin, F.B.; Burt, D.W.; et al. Three chromosome-level duck genome assemblies provide insights into genomic variation during domestication. Nat. Commun. 2021, 12, 5932. [Google Scholar] [CrossRef]
- Hwang, I.; Kim, J.B. Two Faces of White Adipose Tissue with Heterogeneous Adipogenic Progenitors. Diabetes Metab. J. 2019, 43, 752–762. [Google Scholar] [CrossRef]
- Nahmgoong, H.; Jeon, Y.G.; Park, E.S.; Choi, Y.H.; Han, S.M.; Park, J.; Ji, Y.; Sohn, J.H.; Han, J.S.; Kim, Y.Y.; et al. Distinct properties of adipose stem cell subpopulations determine fat depot-specific characteristics. Cell Metab. 2022, 34, 458–472e456. [Google Scholar] [CrossRef] [PubMed]
- Macotela, Y.; Emanuelli, B.; Mori, M.A.; Gesta, S.; Schulz, T.J.; Tseng, Y.H.; Kahn, C.R. Intrinsic differences in adipocyte precursor cells from different white fat depots. Diabetes 2012, 61, 1691–1699. [Google Scholar] [CrossRef] [PubMed]
- Ferrero, R.; Rainer, P.; Deplancke, B. Toward a Consensus View of Mammalian Adipocyte Stem and Progenitor Cell Heterogeneity. Trends Cell Biol. 2020, 30, 937–950. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Symbol | Accession Numbers | Sequence (5′–3′) | Length (bp) |
|---|---|---|---|
| NOTCH1 | XM_046928731.1 | F: AACGCTGTGGATGATCTGGG | 131 |
| R: AGCGGGGTCTCCTCCTTATT | |||
| PPARγ | NM_001001460 | F: GTGCAATCAAAATGGAGCC | 170 |
| R: CTTACAACCTTCACATGCAT | |||
| FABP4 | NM_204290 | F: ATGTGCGACCAGTTTGT | 143 |
| R: TCACCATTGATGCTGATAG | |||
| C/EBPα | NM_001031459 | F: GGAGCAAGCCAACTTCTACG | 181 |
| R: GTCGATGGAGTGCTCGTTCT | |||
| C/EBPβ | NM_205253 | F: CGCCCGCCTTTAAATCCATG | 151 |
| R: GGGCTGAAGTCAATGGCTCT | |||
| HES1 | XM_040679737 | F: GGACGCGCTGAAGAAGGATA | 203 |
| R: CTTCGCAGGTGGAGAGGAAC | |||
| HEY1 | XM_040665051 | F: TGGCTGAAGTGGCTCGATAC | 175 |
| R: TGAGGGTGATGTCCAAAGGC | |||
| HEY2 | XM_040669098 | F: GGGCAGCGAGAACAACTACT | 125 |
| R: CCCGGCGCCTTTTCTCTATA | |||
| SOCS3 | NM_204600.2 | F: GCCTCAAGACGTTCAGCTCT | 196 |
| R: GTCTTGACGCTGAGGGTGAA | |||
| NUMB | AF176086 | F: ACAAAACCCGTGACAGTGGT | 168 |
| R: GGCACGGACAGTCTTTGAGA | |||
| NR2F2 | XM_046924463 | F: TGACCTGGAGCGAGTTGTTC | 185 |
| R: GTCGACATGCAACGCTTTCA | |||
| GAPDH | NM_204305 | F: CCTCTCTGGCAAAGTCCAAG | 200 |
| R: CATCTGCCCATTTGATGTTG |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, Z.; Su, Y.; Zhao, M.; Ma, Z.; Li, J.; Hou, Z.; Li, H. NOTCH1 as a Negative Regulator of Avian Adipocyte Differentiation: Implications for Fat Deposition. Animals 2024, 14, 585. https://doi.org/10.3390/ani14040585
Wang Z, Su Y, Zhao M, Ma Z, Li J, Hou Z, Li H. NOTCH1 as a Negative Regulator of Avian Adipocyte Differentiation: Implications for Fat Deposition. Animals. 2024; 14(4):585. https://doi.org/10.3390/ani14040585
Chicago/Turabian StyleWang, Zheng, Yue Su, Mingyu Zhao, Zhenhua Ma, Jianhui Li, Zhuocheng Hou, and Huifeng Li. 2024. "NOTCH1 as a Negative Regulator of Avian Adipocyte Differentiation: Implications for Fat Deposition" Animals 14, no. 4: 585. https://doi.org/10.3390/ani14040585
APA StyleWang, Z., Su, Y., Zhao, M., Ma, Z., Li, J., Hou, Z., & Li, H. (2024). NOTCH1 as a Negative Regulator of Avian Adipocyte Differentiation: Implications for Fat Deposition. Animals, 14(4), 585. https://doi.org/10.3390/ani14040585