

Comparative Transcriptome Analysis of the Hepatopancreas from Macrobrachium rosenbergii Exposed to the Heavy Metal Copper

Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Experimental Animal
2.2. Cu Exposure Experiment and Sample Collection
2.3. RNA Extraction, Library Construction, and RNA Sequencing
2.4. Transcriptome Assembly and Functional Annotation Analysis
2.5. Identification of Differentially Expressed Genes (DEGs) and Functional Enrichment Analysis
2.6. Simple Sequence Repeat (SSR) and Single-Nucleotide Polymorphism (SNP) Detection
2.7. Validation of RNA-Seq Results with Quantitative Real-Time PCR (qRT-PCR)
3. Results
3.1. RNA Sequencing and De Novo Assembly
3.2. Functional Annotation and Classification of the Transcriptome
3.3. Identification of Differentially Expressed Genes (DEGs)
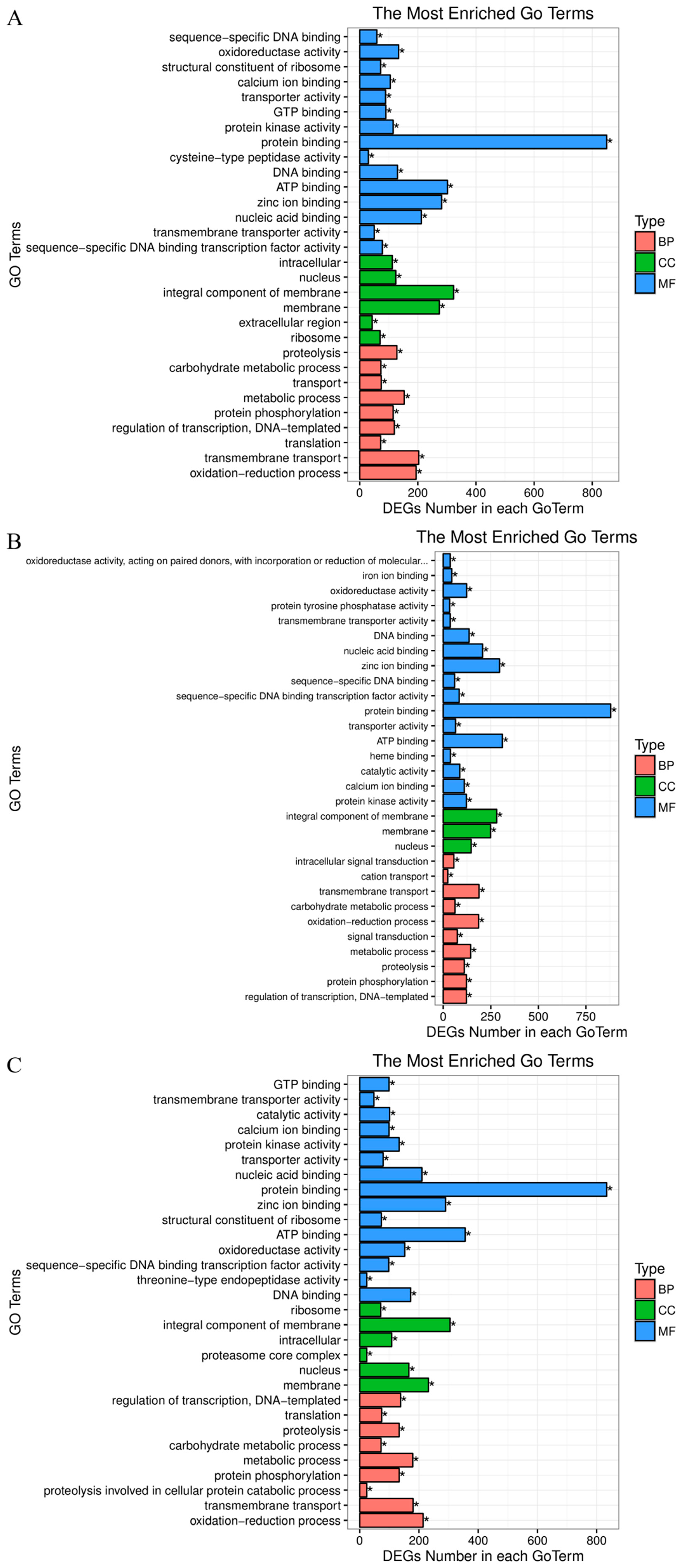
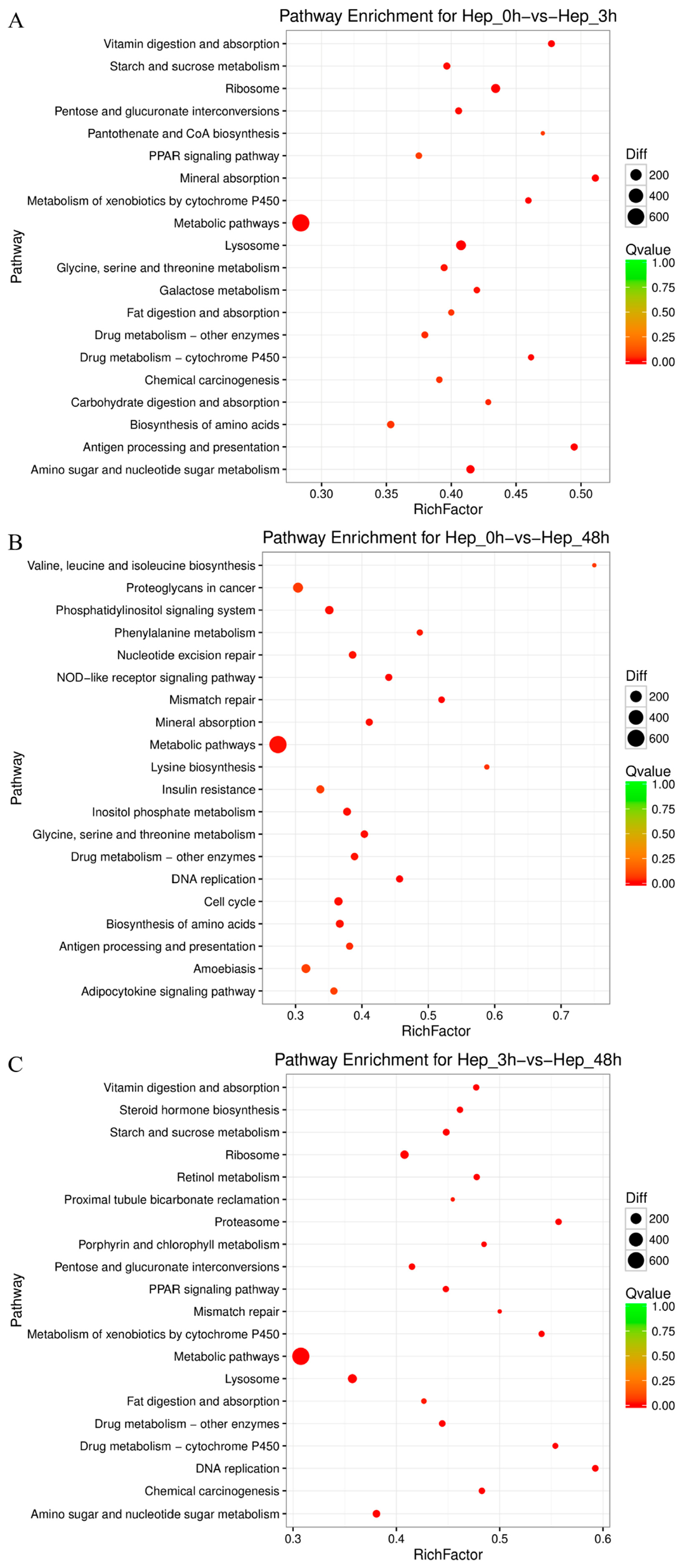
3.4. GO and KEGG Enrichment Analyses of DEGs
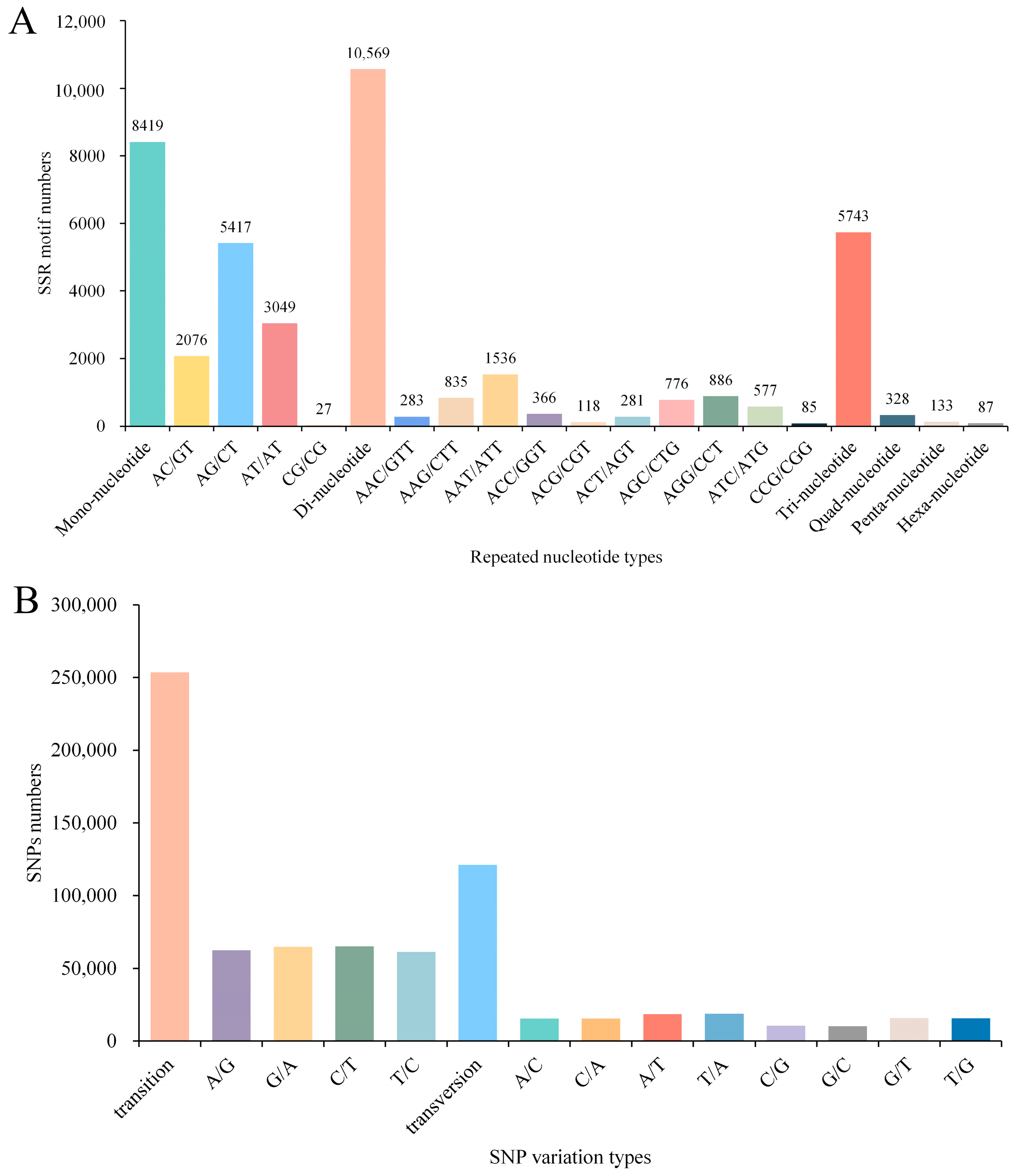
3.5. Detection of SSR and SNP Loci in the Transcriptome
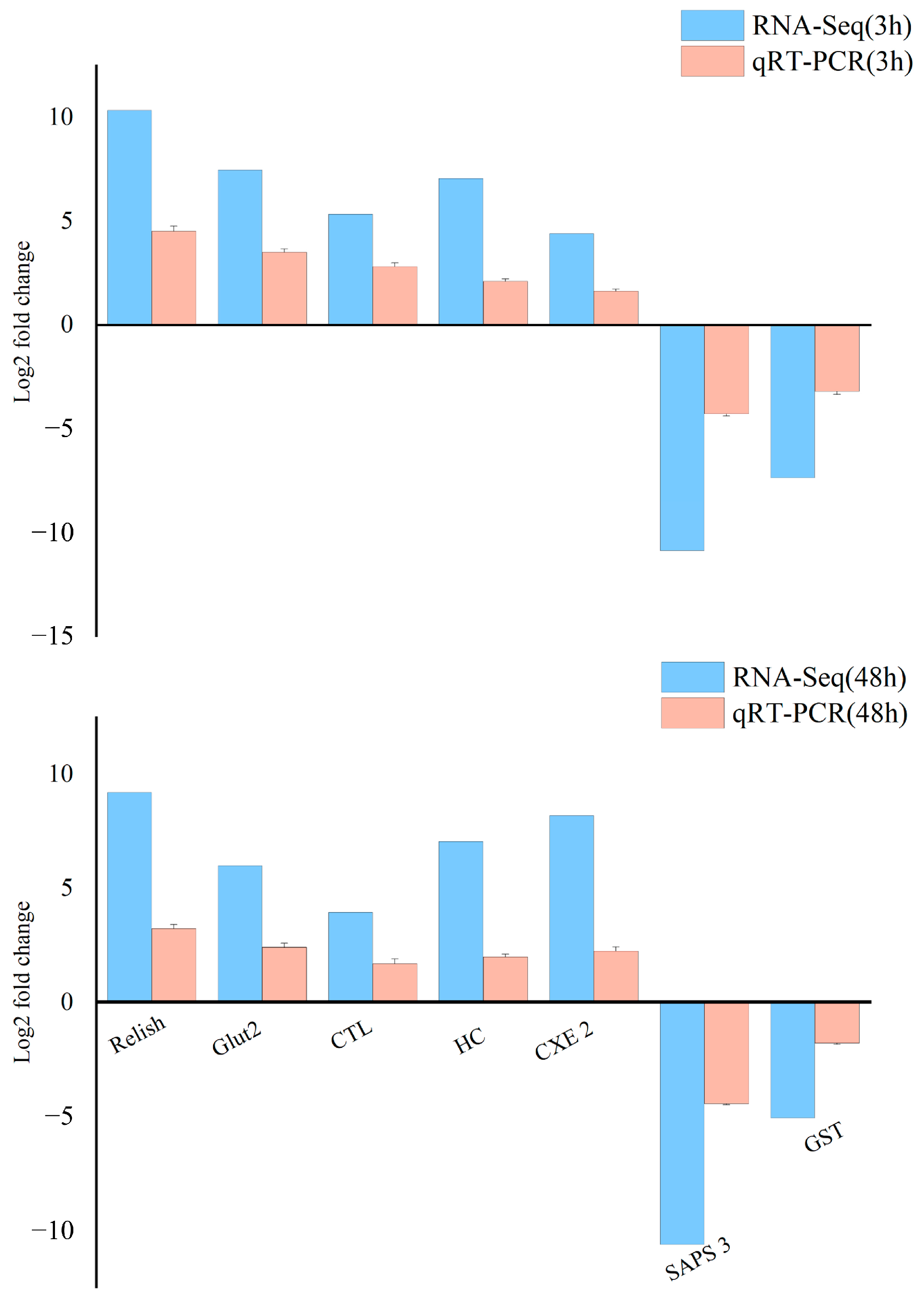
3.6. The qRT-PCR Validation of Differentially Expressed Genes
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Dawood, M. Dietary Cu requirements for aquatic animals: A review. Biol. Trace Elem. Res. 2022, 200, 5273–5282. [Google Scholar] [CrossRef] [PubMed]
- Bian, C.; Zheng, Z.; Su, J.; Chang, S.; Yu, H.; Bao, J.; Xin, Y.; Jiang, X. Copper homeostasis and cuproptosis in tumor pathogenesis and therapeutic strategies. Front. Pharmacol. 2023, 14, 1271613. [Google Scholar] [CrossRef] [PubMed]
- Qian, D.; Xu, C.; Chen, C.; Qin, J.; Chen, L.; Li, E. Toxic effect of chronic waterborne copper exposure on growth, immunity, anti-oxidative capacity and gut microbiota of Pacific white shrimp Litopenaeus vannamei. Fish. Shellfish Immunol. 2020, 100, 445–455. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Liu, H.; Liber, K.; Jiang, T.; Yang, J. Copper-induced ionoregulatory disturbance, histopathology, and transcriptome responses in freshwater mussel (Anodonta woodiana) gills. Fishes 2023, 8, 368. [Google Scholar] [CrossRef]
- Guo, H.; Li, K.; Wang, W.; Wang, C.; Shen, Y. Effects of copper on hemocyte apoptosis, ROS production, and gene expression in white shrimp Litopenaeus vannamei. Biol. Trace Elem. Res. 2017, 179, 318–326. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Guan, T.; Wang, G.; Gu, J.; Wu, N.; Zhu, C.; Wang, H.; Li, J. Effects of copper on gill function of juvenile oriental river prawn (Macrobrachium nipponense): Stress and toxic mechanism. Aquat. Toxicol. 2023, 261, 106631. [Google Scholar] [CrossRef] [PubMed]
- García-Villada, L.; Rico, M.; Altamirano, M.; Sánchez-Martín, L.; López-Rodas, V.; Costas, E. Occurrence of Cu resistant mutants in the toxic cyanobacteria Microcystis aeruginosa: Characterisation and future implications in the use of Cu sulphate as algaecide. Water Res. 2004, 38, 2207–2213. [Google Scholar] [CrossRef]
- Zhuang, W.; Zhou, F. Distribution, source and pollution assessment of heavy metals in the surface sediments of the Yangtze River Estuary and its adjacent East China Sea. Mar. Pollut. Bull. 2021, 164, 112002. [Google Scholar] [CrossRef] [PubMed]
- Guo, H.; Xian, J.; Zheng, P.; Lu, Y.; Wang, L.; Zhang, X.; Li, J.; Zhang, Z.; Wang, A. Dietary copper affects antioxidant status of shrimp (Penaeus monodon) reared in low salinity water. Aquac. Rep. 2022, 22, 100979. [Google Scholar] [CrossRef]
- Xing, K.; Liu, Y.; Yan, C.; Zhou, Y.; Zhang, R.; Sun, Y.; Zhang, J. Transcriptomic analysis of Neocaridina denticulate sinensis hepatopancreas indicates immune changes after Cu exposure. Fish. Shellfish Immunol. 2022, 121, 23–30. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Zhang, M.; Yu, W.; Li, J.; Kong, D. Ecotoxicological risk ranking of 19 metals in the lower Yangtze River of China based on their threats to aquatic wildlife. Sci. Total Environ. 2022, 812, 152370. [Google Scholar] [CrossRef] [PubMed]
- Lan, X.; Peng, X.; Du, T.; Xia, Z.; Gao, Q.; Tang, Q.; Yi, S.; Yang, G. Alterations of the gut microbiota and metabolomics associated with the different growth performances of Macrobrachium rosenbergii families. Animals 2023, 13, 1539. [Google Scholar] [CrossRef] [PubMed]
- Tu, H.; Peng, X.; Yao, X.; Tang, Q.; Xia, Z.; Li, J.; Yang, G.; Yi, S. Integrated transcriptomic and metabolomic analyses reveal low-temperature tolerance mechanism in giant freshwater prawn Macrobrachium rosenbergii. Animals 2023, 13, 1605. [Google Scholar] [CrossRef] [PubMed]
- Zhong, Z.; Chen, G.; Tu, H.; Yao, X.; Peng, X.; Lan, X.; Tang, Q.; Yi, S.; Xia, Z.; Cai, M.; et al. Transcriptomic analysis and functional gene expression in different stages of gonadal development of Macrobrachium rosenbergii. Fishes 2023, 8, 94. [Google Scholar] [CrossRef]
- Liang, J.; Abdullah, A.L.B.; Wang, H.; Liu, G.; Han, M. Change in energy-consuming strategy, nucleolar metabolism and physical defense in Macrobrachium rosenbergii after acute and chronic polystyrene nanoparticles exposure. Aquat. Toxicol. 2023, 263, 106711. [Google Scholar] [CrossRef] [PubMed]
- Farhadi, A.; Tang, S.; Huang, M.; Yu, Q.; Xu, C.; Li, E. Identification of key immune and stress related genes and pathways by comparative analysis of the gene expression profile under multiple environmental stressors in pacific white shrimp (Litopenaeus vannamei). Fish. Shellfish Immunol. 2023, 135, 108695. [Google Scholar] [CrossRef]
- Zhang, R.; Zhao, Z.; Li, M.; Luo, L.; Wang, S.; Guo, K.; Xu, W. Effects of saline-alkali stress on the tissue structure, antioxidation, immunocompetence and metabolomics of Eriocheir sinensis. Sci. Total Environ. 2023, 871, 162109. [Google Scholar] [CrossRef]
- Kouba, A.; Buric, M.; Kozák, P. Bioaccumulation and effects of heavy metals in crayfish: A review. Water Air Soil. Poll. 2010, 211, 5–16. [Google Scholar] [CrossRef]
- Abbas, M.; Abd El-Aziz, M.; Kaddah, M.; Hassan, A.K.; El-Naggar, H.A.; Radwan, M.; El-Tabakh, M.; Afifi, M.A.; Bashar, M. Bioaccumulation, biosedimentation, and health hazards of elements in crayfish, Procambarus clarkii from El-Rahawi Drain and El-Qanatir in the River Nile, Egypt. Biol. Trace Elem. Res. 2023, 201, 3050–3059. [Google Scholar] [CrossRef] [PubMed]
- Sang, W.; Liu, J.; Xing, Y.; Feng, C.; Hu, Q.; Lang, Y.; Li, X.; Bao, J.; Jiang, H. Transcriptome analysis of hepatopancreas of Chinese grass shrimp, Palaemonetes sinensis, infected by Enterocytospora artemiae. Fish. Shellfish Immunol. 2023, 133, 108557. [Google Scholar] [CrossRef]
- Zhang, L.; Song, Z.; Zhou, Y.; Zhong, S.; Yu, Y.; Liu, T.; Gao, X.; Li, L.; Kong, C.; Wang, X.; et al. The accumulation of toxic elements (Pb, Hg, Cd, As, and Cu) in red swamp crayfish (Procambarus clarkii) in Qianjiang and the associated risks to human health. Toxics 2023, 11, 635. [Google Scholar] [CrossRef]
- Wu, Y.; Tian, W.; Cheng, C.; Yang, L.; Ye, Q.; Li, W.; Jiang, J. Effects of cadmium exposure on metabolism, antioxidant defense, immune function, and the hepatopancreas transcriptome of Cipangopaludina cathayensis. Ecotoxicol. Environ. Saf. 2023, 264, 115416. [Google Scholar] [CrossRef] [PubMed]
- Reddy, R.; Pillai, B.R.; Adhikari, S. Bioaccumulation of copper in post-larvae and juveniles of freshwater prawn Macrobrachium rosenbergii (de man) exposed to sub-lethal levels of copper sulfate. Aquaculture 2006, 252, 356–360. [Google Scholar] [CrossRef]
- Li, N.; Zhao, Y.; Yang, J. Impact of waterborne copper on the structure of gills and hepatopancreas and its impact on the content of metallothionein in juvenile giant freshwater prawn Macrobrachium rosenbergii (Crustacea: Decapoda). Arch. Environ. Con Tox. 2007, 52, 73–79. [Google Scholar] [CrossRef] [PubMed]
- Li, N.; Ma, Q.; Yang, J.; Zhao, Y.; Brochon, J.C. Effects of water-borne copper on the gills and hepatopancreas of Macrobrachium rosenbergii. Integr. Zool. 2008, 3, 299–310. [Google Scholar] [CrossRef]
- Pabuwal, V.; Boswell, M.; Pasquali, A.; Wise, S.S.; Kumar, S.; Shen, Y.; Garcia, T.; LaCerte, C., Jr.; Wise, J.P.; Wise, J.P.S.; et al. Transcriptomic analysis of cultured whale skin cells exposed to hexavalent chromium [Cr (VI)]. Aquat. Toxicol. 2013, 134, 74–81. [Google Scholar] [CrossRef] [PubMed]
- Jin, J.; Wang, Y.; Wu, Z.; Hergazy, A.; Lan, J.; Zhao, L.; Liu, X.; Chen, N.; Lin, L. Transcriptomic analysis of liver from grass carp (Ctenopharyngodon idellus) exposed to high environmental ammonia reveals the activation of antioxidant and apoptosis pathways. Fish. Shellfish Immunol. 2017, 63, 444–451. [Google Scholar] [CrossRef] [PubMed]
- Guo, H.; Miao, Y.; Xian, J.; Qian, K.; Wang, A. Expression profile of antioxidant enzymes in hemocytes from freshwater prawn Macrobrachium rosenbergii exposed to an elevated level of copper. Bull. Environ. Contam. Toxicol. 2015, 95, 447–451. [Google Scholar] [CrossRef] [PubMed]
- Grabherr, M.G.; Haas, B.J.; Yassour, M.; Levin, J.Z.; Thompson, D.A.; Amit, I.; Adiconis, X.; Fan, L.; Raychowdhury, R.; Zeng, Q.; et al. Full-length transcriptome assembly from RNA-Seq data without a reference genome. Nat. Biotechnol. 2011, 29, 130–644. [Google Scholar] [CrossRef] [PubMed]
- Pertea, G.; Huang, X.; Liang, F.; Antonescu, V.; Sultana, R.; Karamycheva, S.; Lee, Y.; White, J.; Cheung, F.; Parvizi, B. TIGR Gene Indices clustering tools (TGICL): A software system for fast clustering of large EST datasets. Bioinformatics 2003, 19, 651–652. [Google Scholar] [CrossRef] [PubMed]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef] [PubMed]
- Conesa, A.; Gotz, S.; Garcia-Gomez, J.M.; Terol, J.; Talon, M.; Robles, M. Blast2GO: A universal tool for annotation, visualization and analysis in functional genomics research. Bioinformatics 2005, 21, 3674–3676. [Google Scholar] [CrossRef] [PubMed]
- Xie, C.; Mao, X.; Huang, J.; Ding, Y.; Wu, J.; Dong, S.; Kong, L.; Gao, G.; Li, C.; Wei, L. KOBAS 2.0: A web server for annotation and identification of enriched pathways and diseases. Nucleic Acids Res. 2011, 39, W316–W322. [Google Scholar] [CrossRef] [PubMed]
- Li, B.; Dewey, C.N. RSEM: Accurate transcript quantification from RNA-Seq data with or without a reference genome. BMC Bioinform. 2011, 12, 323. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Feng, Z.; Wang, X.; Wang, X.; Zhang, X. DEGseq: An R package for identifying differentially expressed genes from RNA-seq data. Bioinformatics 2010, 26, 136–138. [Google Scholar] [CrossRef]
- Anders, S.; Huber, W. Differential expression analysis for sequence count data. Genome Biol. 2010, 11, R106. [Google Scholar] [CrossRef] [PubMed]
- Young, M.; Wakefield, M.J.; Smyth, G.K.; Oshlack, A. Gene ontology analysis for RNA-seq: Accounting for selection bias. Genome Biol. 2010, 11, R14. [Google Scholar] [CrossRef]
- Thiel, T.; Michalek, W.; Varshney, R.K.; Graner, A. Exploiting EST databases for the development and characterization of gene-derived SSR-markers in barley (Hordeum vulgare L.). Theor. Appl. Genet. 2003, 106, 411–422. [Google Scholar] [CrossRef] [PubMed]
- McKenna, A.; Hanna, M.; Banks, E.; Sivachenko, A.; Cibulskis, K.; Kernytsky, A.; Garimella, K.; Altshuler, D.; Gabriel, S.; Daly, M.; et al. The genome analysis toolkit: A MapReduce framework for analyzing next-generation DNA sequencing data. Genome Res. 2010, 20, 1297–1303. [Google Scholar] [CrossRef] [PubMed]
- Zhou, T.; Liu, J.; Chan, S.; Wang, W. Molecular characterization and expression dynamics of three key genes in the PI3K-AKT pathway reveal its involvement in the immunotoxicological responses of the giant river prawn Macrobrachium rosenbergii to acute ammonia and nitrite stress. Ecotox. Environ. Safe 2021, 208, 111767. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Guo, H.; Deng, M.; Liang, J.; Lu, W.; Shen, Y. Gill transcriptome alterations in Macrobrachium rosenbergii under copper exposure. Chemosphere 2019, 233, 796–808. [Google Scholar] [CrossRef]
- Xing, K.; Liu, Y.; Yan, C.; Zhou, Y.; Sun, Y.; Su, N.; Yang, F.; Xie, S.; Zhang, J. Transcriptome analysis of Neocaridina denticulate sinensis under copper exposure. Gene 2021, 764, 145098. [Google Scholar] [CrossRef]
- Habib, Y.J.; Wan, H.; Sun, Y.; Shi, J.; Yao, C.; Lin, J.; Ge, H.; Wang, Y.; Zhang, Z. Genome-wide identification of toll-like receptors in pacific white shrimp (Litopenaeus vannamei) and expression analysis in response to Vibrio parahaemolyticus invasion. Aquaculture 2021, 532, 735996. [Google Scholar] [CrossRef]
- Cho, H.; Park, K.H.; Jang, Y.; Cho, Y.; Heo, Y.K.; Kim, M.; Kim, Y.B. Identification and characterization of a toll-like receptor gene from Macrobrachium nipponense. Fish. Shellfish Immunol. 2021, 108, 109–115. [Google Scholar] [CrossRef]
- Li, Y.; Yuan, W.; Xu, Q.; Liu, H.; Dai, X. The regulation of immune responses against white spot syndrome virus or Vibrio alginolyticus in toll-like receptors silenced giant freshwater prawn (Macrobrachium rosenbergii). Fish. Shellfish Immunol. 2020, 107, 84–94. [Google Scholar] [CrossRef]
- Xu, M.; Wu, J.; Ge, D.; Wu, C.; Chi, C.; Lv, Z.; Liao, Z.; Liu, H. A novel toll-like receptor from Mytilus coruscus is induced in response to stress. Fish. Shellfish Immunol. 2018, 78, 331–337. [Google Scholar] [CrossRef]
- Shen, D.; Tong, M.; Guo, J.; Mei, X.; Xia, D.; Qiu, Z.; Zhao, Q. A pattern recognition receptor C-type lectin-S6 (CTL-S6) is involved in the immune response in the Silkworm (Lepidoptera: Bombycidae). J. Insect Sci. 2021, 21, 1–8. [Google Scholar] [CrossRef]
- Zhang, X.; Wang, X.; Huang, Y.; Hui, K.; Shi, Y.; Wang, W.; Ren, Q. Cloning and characterization of two different ficolins from the giant freshwater prawn Macrobrachium rosenbergii. Dev. Comp. Immunol. 2014, 44, 359–369. [Google Scholar] [CrossRef]
- Luo, J.; Chen, Y.; Huang, Y.; Feng, J.; Yuan, Y.; Jian, J.; Cai, S.; Yang, S. A novel C-type lectin for Litopenaeus vannamei involved in the innate immune response against Vibrio infection. Fish. Shellfish Immunol. 2023, 135, 108621. [Google Scholar] [CrossRef]
- Zhang, Z.; Zhang, C.; Dai, X.; Zhang, R.; Cao, X.; Wang, K.; Huang, X.; Ren, Q. Two relish isoforms produced by alternative splicing participate in the regulation of antimicrobial peptides expression in Procambarus clarkii intestine. Fish. Shellfish Immunol. 2020, 99, 107–118. [Google Scholar] [CrossRef]
- Ruan, Z.; Wan, Z.; Yang, L.; Li, W.; Wang, Q. JAK/STAT signalling regulates antimicrobial activities in Eriocheir sinensis. Fish. Shellfish Immunol. 2019, 84, 491–501. [Google Scholar] [CrossRef]
- Shi, Y.; Jin, M.; Ma, F.; Huang, Y.; Huang, X.; Feng, J.; Zhao, L.; Chen, Y.; Ren, Q. Involvement of relish gene from Macrobrachium rosenbergii in the expression of anti-microbial peptides. Dev. Comp. Immunol. 2015, 52, 236–244. [Google Scholar] [CrossRef]
- Coates, C.J.; Decker, H. Immunological properties of oxygen-transport proteins: Hemoglobin, hemocyanin and hemerythrin. Cell Mol. Life Sci. 2017, 74, 293–317. [Google Scholar] [CrossRef]
- Zhang, X.; Huang, C.; Qin, Q. Antiviral properties of hemocyanin isolated from shrimp Penaeus monodon. Antivir. Res. 2004, 61, 93–99. [Google Scholar] [CrossRef]
- Coates, C.J.; Nairn, J. Diverse immune functions of hemocyanins. Dev. Comp. Immunol. 2014, 45, 43–55. [Google Scholar] [CrossRef]
- Minghetti, M.; Leaver, M.J.; Carpenè, E.; George, S.G. Copper transporter 1, metallothionein and glutathione reductase genes are differentially expressed in tissues of sea bream (Sparus aurata) after exposure to dietary or waterborne copper. Comp. Biochem. Phys. C. 2008, 147, 450–459. [Google Scholar] [CrossRef]
- Zhao, D.; Zhang, X.; Liu, D.; Ru, S. Cu accumulation, detoxification and tolerance in the red swamp crayfish Procambarus clarkii. Ecotoxicol. Environ. Saf. 2019, 175, 201–207. [Google Scholar] [CrossRef]
- Wei, H.; Wang, S.; Xu, E.; Liu, J.; Li, X.; Wang, Z. Synergistic toxicity of microcystin-LR and Cu to Zebrafish (Danio rerio). Sci. Total Environ. 2020, 713, 136393. [Google Scholar] [CrossRef]
- Ma’rifah, F.; Saputri, M.R.; Soegianto, A.; Irawan, B.; Putranto, T.W.C. The change of metallothionein and oxidative response in gills of the Oreochromis niloticus after exposure to copper. Animals 2019, 9, 353. [Google Scholar] [CrossRef]
- Markossian, K.A.; Kurganov, B.I. Copper chaperones, intracellular copper trafficking proteins. Function, structure, and mechanism of action. Biochemistry 2003, 68, 827–837. [Google Scholar] [CrossRef]
- Linz, R.; Lutsenko, S. Copper-transporting ATPases ATP7A and ATP7B: Cousins, not twins. J. Bioenerg. Biomembr. 2007, 39, 403–407. [Google Scholar] [CrossRef]
- Guo, H.; Chen, T.; Liang, Z.; Fan, L.; Shen, Y.; Zhou, D. iTRAQ and PRM-based comparative proteomic profiling in gills of white shrimp Litopenaeus vannamei under copper stress. Chemosphere 2021, 263, 128270. [Google Scholar] [CrossRef]
- Zang, Y.; Zhang, X.; Huang, P.; Ame, K.H.; Shen, H. Cloning and expression of two carboxylesterases, and their activity modulation in Chinese mitten crab Eriocheir sinensis under pesticide exposer. Turk. J. Fish. Aquat. Sci. 2020, 20, 521–529. [Google Scholar] [CrossRef]
- Spencer, N.Y.; Engelhardt, J.F. The basic biology of redoxosomes in cytokine-mediated signal transduction and implications for disease-specific therapies. Biochemistry 2014, 53, 1551–1564. [Google Scholar] [CrossRef]
- Ni, Y.; Zhang, J.; Zhu, W.; Duan, Y.; Bai, H.; Luan, C. Echinacoside inhibited cardiomyocyte pyroptosis and improved heart function of HF rats induced by isoproterenol via suppressing NADPH/ROS/ER stress. J. Cell Mol. Med. 2022, 26, 5414–5425. [Google Scholar] [CrossRef]
- Zhao, D.; Zhang, X.; Li, X.; Ru, S.; Wang, Y.; Yin, J.; Liu, D. Oxidative damage induced by copper in testis of the red swamp crayfish Procambarus clarkii and its underlying mechanisms. Aquat. Toxicol. 2019, 207, 120–131. [Google Scholar] [CrossRef]
- Ou, H.; Liu, J. Role of Nrf2-Keap1 signaling in the antioxidant defense response induced by low salinity in the kuruma shrimp (Marsupenaeus japonicus). Aquac. Int. 2022, 30, 2793–2811. [Google Scholar] [CrossRef]
- Cheng, C.; Ma, H.; Deng, Y.; Feng, J.; Chen, X.; Guo, Z. The role of Mu-type glutathione S-transferase in the mud crab (Scylla paramamosain) during ammonia stress. Comp. Biochem. Physiol. C Toxicol. Pharmacol. 2020, 227, 108642. [Google Scholar] [CrossRef]
- Liu, K.; Yeh, M.S.; Kou, G.; Cheng, W.; Lo, C.F. Identification and cloning of a selenium-dependent glutathione peroxidase from tiger shrimp, Penaeus monodon, and its transcription following pathogen infection and related to the molt stages. Dev. Comp. Immunol. 2010, 34, 935–944. [Google Scholar] [CrossRef]
- Gu, Z.; Ren, Y.; Liang, C.; Zhang, X.; Geng, Y.; Xu, D.; Ye, N. Glutathione s-transferase (GST) identified from giant kelp Macrocystis pyrifera increases the copper tolerance of Synechococcus elongatus pcc 7942. J. Ocean Univ. China 2023, 22, 777–789. [Google Scholar] [CrossRef]
- Zhang, X.; Zhang, W.; Zhao, L.; Zheng, L.; Wang, B.; Song, C.; Liu, S. Mechanisms of gills response to cadmium exposure in greenfin horse-faced filefish (Thamnaconus septentrionalis): Oxidative stress, immune response, and energy metabolism. Animals 2024, 7, 561. [Google Scholar] [CrossRef]
- Kim, B.; Rhee, J.; Jeong, C.; Seo, J.S.; Park, G.S.; Lee, Y.; Lee, J. Heavy metals induce oxidative stress and trigger oxidative stress-mediated heat shock protein (hsp) modulation in the intertidal copepod Tigriopus Japonicus. Comp. Biochem. Physiol. C Toxicol. Pharmacol. 2014, 166, 65–74. [Google Scholar] [CrossRef] [PubMed]
- Jin, Y.; Clark, A.B.; Slebos, R.; Al-Refai, H.; Taylor, J.A.; Kunkel, T.A.; Resnick, M.A.; Gordenin, D.A. Cadmium is a mutagen that acts by inhibiting mismatch repair. Nat. Genet. 2003, 34, 326–329. [Google Scholar] [CrossRef]
- Chang, M.; Wang, W.; Wang, A.; Tian, T.; Wang, P.; Zheng, Y.; Liu, Y. Effects of cadmium on respiratory burst, intracellular Ca2+ and DNA damage in the white shrimp Litopenaeus vannamei. Comp. Biochem. Physiol. C Pharmacol. Toxicol. Endocrinol. 2009, 149, 581–586. [Google Scholar] [CrossRef]
- Hsu, T.; Tsai, H.; Huang, K.; Luan, M.; Hsieh, C. Sublethal levels of cadmium down-regulate the gene expression of DNA mismatch recognition protein MutS homolog 6 (MSH6) in zebrafish (Danio rerio) embryos. Chemosphere 2010, 81, 748–754. [Google Scholar] [CrossRef]
- Olave, M.C.; Graham, R.P. Mismatch repair deficiency: The what, how and why it is important. Gene Chromosome Canc. 2022, 61, 314–321. [Google Scholar] [CrossRef]
- Huang, D.; Qiao, X.; Liang, Q.; Wei, W.; Kong, J.; Kang, C.; Liu, Y.; Wang, W. Molecular characterization and function analysis of a nucleotide excision repair gene Rad23 from Litopenaeus Vannamei after Vibrio alginolyticus challenge. Fish. Shellfish Immun. 2018, 83, 190–204. [Google Scholar] [CrossRef]
- Kim, B.M.; Rhee, J.S.; Choi, I.Y.; Lee, Y.M. Transcriptional profiling of antioxidant defense system and heat shock protein (Hsp) families in the cadmium- and copper-exposed marine ciliate Euplotes crassu. Genes Genom. 2018, 40, 85–98. [Google Scholar] [CrossRef]
- Liang, C.; Liu, J.; Cao, F.; Li, Z.; Chen, T. Transcriptomic analyses of the acute ammonia stress response in the hepatopancreas of the kuruma shrimp (Marsupenaeus japonicus). Aquaculture 2019, 513, 734328. [Google Scholar] [CrossRef]
- Ren, X.; Xu, Y.; Zhang, Y.; Wang, X.; Liu, P.; Li, J. Comparative accumulation and transcriptomic analysis of juvenile Marsupenaeus japonicus under cadmium or copper exposure. Chemosphere 2020, 249, 126157. [Google Scholar] [CrossRef] [PubMed]
- Tang, D.; Shi, X.; Guo, H.; Bai, Y.; Shen, C.; Zhang, Y.; Wang, Z. Comparative transcriptome analysis of the gills of Procambarus clarkii provides novel insights into the immune-related mechanism of copper stress tolerance. Fish. Shellfish Immunol. 2020, 96, 32–40. [Google Scholar] [CrossRef]
- Zaib, S.; Hayyat, A.; Ali, N.; Gul, A.; Naveed, M.; Khan, I. Role of mitochondrial membrane potential and lactate dehydrogenase a in apoptosis. Anticancer Agents Med. Chem. 2022, 22, 2048–2062. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Tang, N.; Hadden, T.J.; Rishi, A.K. Akt, FoxO and regulation of apoptosis. Biochim. Biophys. Acta Mol. Cell Res. 2011, 1813, 1978–1986. [Google Scholar] [CrossRef] [PubMed]
- Shukla, S.; Rizvi, F.; Raisuddin, S.; Kakkar, P. FoxO proteins’ nuclear retention and BH3-only protein Bim induction evoke mitochondrial dysfunction-mediated apoptosis in berberine-treated HepG2 cells. Free Radic. Biol. Med. 2014, 76, 185–199. [Google Scholar] [CrossRef] [PubMed]
- Kopeina, G.S.; Zhivotovsky, B. Caspase-2 as a master regulator of genomic stability. Trends Cell Biol. 2021, 31, 712–720. [Google Scholar] [CrossRef] [PubMed]
- Qin, S.; Xu, C.; Li, S.; Wang, X.; Sun, X.; Wang, P.; Zhang, B.; Ren, H. Hyperthermia induces apoptosis by targeting survivin in esophageal cancer. Oncol. Rep. 2015, 34, 2656–2664. [Google Scholar] [CrossRef]
- Schwerdtle, T.; Hamann, I.; Jahnke, G.; Walter, I.; Richter, C.; Parsons, J.L.; Dianov, G.L.; Hartwig, A. Impact of copper on the induction and repair of oxidative DNA damage, poly (ADP-ribosyl) ation and PARP-1 activity. Mol. Nutr. Food Res. 2007, 51, 201–210. [Google Scholar] [CrossRef] [PubMed]
- Xue, Q.; Kang, R.; Klionsky, D.J.; Tang, D.; Liu, J.; Chen, X. Copper metabolism in cell death and autophagy. Autophagy 2023, 19, 2175–2195. [Google Scholar] [CrossRef] [PubMed]
- Sokolova, I.M.; Lannig, G. Interactive effects of metal pollution and temperature on metabolism in aquatic ectotherms: Implications of global climate change. Clim. Res. 2008, 37, 181–201. [Google Scholar] [CrossRef]
- Tang, D.; Liu, R.; Shi, X.; Shen, C.; Bai, Y.; Tang, B.; Wang, Z. Toxic effects of metal copper stress on immunity, metabolism and pathologic changes in chinese mitten crab (Eriocheir japonica sinensis). Ecotoxicology 2021, 30, 632–642. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.; Jeon, M.J.; Won, E.J.; Yoo, J.W.; Lee, Y.M. Effect of heavy metals on the energy metabolism in the brackish water flea Diaphanosoma celebensis. Ecotoxicol. Environ. Saf. 2023, 262, 115189. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Reid, M.A.; Hanse, E.A.; Li, H.; Li, Y.; Ruiz, B.I.; Fan, Q.; Kong, M. SAPS3 subunit of protein phosphatase 6 is an AMPK inhibitor and controls metabolic homeostasis upon dietary challenge in male mice. Nat. Commun. 2023, 14, 1368. [Google Scholar] [CrossRef] [PubMed]
- Rencurel, F.; Waeber, G.; Antoine, B.; Rocchiccioli, F.; Maulard, P.; Girard, J.; Leturque, A. Requirement of glucose metabolism for regulation of glucose transporter type 2 (Glut2) gene expression in liver. Biochem. J. 1996, 314 Pt 3, 903–909. [Google Scholar] [CrossRef] [PubMed]
- Patil, A.; Dwivedi, P.S.R.; Gaonkar, S.N.; Kumbhar, V.; Shankar Madiwalar, V.; Khanal, P.; Patil, B.M. Glut-2 mediated glucose uptake analysis of Duranta repens: In-silico and In-vitro approach. J. Diabetes Metab. Disord. 2022, 21, 419–427. [Google Scholar] [CrossRef] [PubMed]
- Kong, L.; Bai, J.; Li, Q. Comparative assessment of genomic SSR, EST-SSR and EST-SNP markers for evaluation of the genetic diversity of wild and cultured Pacific Oyster, Crassostrea gigas Thunberg. Aquaculture 2014, 420, S85–S91. [Google Scholar] [CrossRef]
- Wang, H.; Lv, J.; Zeng, Q.; Liu, Y.; Xing, Q.; Wang, S.; Hu, J.; Bao, L. Genetic differentiation and selection signatures in two bay scallop (Argopecten irradians) breeds revealed by whole-genome resequencing analysis. Aquaculture 2021, 543, 736944. [Google Scholar] [CrossRef]
- Lu, Y.; Zheng, P.; Zhang, Z.; Zhang, X.; Li, J.; Wang, D.; Xu, J.; Xian, J.; Wang, A. Hepatopancreas transcriptome alterations in red claw crayfish (Cherax quadricarinatus) under microcystin-LR (MC-LR) stress. Aquac. Rep. 2023, 29, 101478. [Google Scholar] [CrossRef]
- Cao, J.; Wu, L.; Jin, M.; Li, T.; Hui, K.; Ren, Q. Transcriptome profiling of the Macrobrachium rosenbergii lymphoid organ under the white spot syndrome virus challenge. Fish. Shellfish Immunol. 2017, 67, 27–39. [Google Scholar] [CrossRef] [PubMed]
- Ding, Z.; Jin, M.; Ren, Q. Transcriptome analysis of Macrobrachium rosenbergii intestines under the white spot syndrome virus and poly (I:C) challenges. PLoS ONE 2018, 13, e0204626. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name | Gene ID | Sequence (5′-3′) | Product Length (bp) | Tm (°C) |
|---|---|---|---|---|
| Relish | CL2513.Contig3_All | F: AGGAGGAGGAAGAGGAAGAGGAGAG | 164 | 61 |
| R: TGGCACTGAAGGCTCATCTGGAA | 61 | |||
| Glut 2 | CL1337.Contig2_All | F: TGGCTATTTCTCTGGCGGCATTG | 120 | 60 |
| R: CGATGGAGGCGGAAGTGATGAAC | 60 | |||
| CXE2 | CL426.Contig5_All | F: ATTGTCAGAGGCAAATGGCAGTGT | 141 | 59 |
| R: CGAAGACTTCCAGTTGGTGGTTCT | 59 | |||
| HC | CL2.Contig14_All | F: GGATGCTCTTGCCCAAGGGTAAG | 116 | 60 |
| R: AGTGCCGCCGTGCTCTGATT | 60 | |||
| GST | CL355.Contig7_All | F: CTTGAAGACCCGAGCCACTATTGAT | 166 | 58 |
| R: GAAACCATTGAGCCATCCCAAAGC | 58 | |||
| SAPS 3 | CL281.Contig5_All | F: AGAATGGATACATCTTGGGCTCCCT | 128 | 59 |
| R: GGTATGGCATCACAGGCAGCAAT | 59 | |||
| CTL | CL3394.Contig2_All | F: AGGGCTACTGGATCTGGGTGGA | 138 | 60 |
| R: TCCGAGGGTAACTTTCTTCCGAGT | 60 | |||
| EF-1 | F: GAGGAAGATTGAACGCAAGA | 152 | 60 | |
| R: TTAAGGATGCCAGTCTCCAC | 60 |
| Summary | 0 h | 3 h | 48 h | Total |
|---|---|---|---|---|
| Total raw reads | 65,384,862 | 58,978,912 | 55,185,928 | 179,549,702 |
| Total clean reads | 65,117,218 | 58,693,126 | 54,982,780 | 178,793,124 |
| Total clean read ratio (%) | 99.59 | 99.52 | 99.63 | 99.58 |
| Number of unigenes | 88,579 | 99,729 | 102,062 | 85,984 |
| Mean length of unigenes (bp) | 697.92 | 735.61 | 693.59 | 1077.23 |
| N50 length of unigenes (bp) | 1704 | 1954 | 1782 | 2559 |
| N90 length of unigenes (bp) | 232 | 237 | 226 | 351 |
| Gene ID | Gene | Description | log2FC (3 h) | Regulation | log2FC (48 h) | Regulation |
|---|---|---|---|---|---|---|
| Oxidation–reduction | ||||||
| CL1542.Contig3_All | NOX | NADPH oxidase | 3.9906 | up | 4.2492 | up |
| CL5985.Contig3_All | GPx | glutathione peroxidase-like | 2.5799 | up | −8.2860 | down |
| CL355.Contig7_All | GST | glutathione S-transferase | −7.3801 | down | −5.0590 | down |
| Energy metabolism | ||||||
| CL281.Contig5_All | SAPS 3 | serine/threonine–protein phosphatase 6 regulatory subunit 3 | −10.9208 | down | −10.6095 | down |
| Unigene17066_All | FAS | fatty acid synthase | −1.2389 | down | −2.6630 | down |
| CL441.Contig3_All | ACC | acetyl-CoA carboxylase | 3.0290 | up | −3.2607 | down |
| CL800.Contig2_All | SCD 1 | delta-9 acyl-CoA desaturase | −1.0850 | down | −1.8534 | down |
| CL1337.Contig2_All | Glut 2 | glucose transporter 2 | 7.4867 | up | 6.0060 | up |
| Apoptosis | ||||||
| CL4973.Contig1_All | Cyt C | cytochrome c | - | - | 2.7752 | up |
| CL1823.Contig6_All | FOXO | forkhead box protein O-like | 3.5861 | up | 2.3504 | up |
| CL7764.Contig1_All | survivin | survivin | −2.4683 | down | −4.5185 | down |
| CL1421.Contig1_All | Casp 2 | caspase 2 | 1.3839 | up | 1.2932 | up |
| Copper homeostasis | ||||||
| CL2846.Contig2_All | Ctr 1 | High-affinity copper uptake protein 1 | 1.3229 | up | 1.6525 | up |
| Unigene9795_All | COX 17 | cytochrome c oxidase copper chaperone | 1.5825 | up | 1.6470 | up |
| Unigene13182_All | Cu-MT | copper-induced metallothionein | −1.2944 | down | 4.4623 | up |
| CL3799.Contig3_All | ATP 7A | copper-transporting ATPase 1-like | - | - | 4.8592 | up |
| Detoxification | ||||||
| CL1886.Contig2_All | CYP450 | cytochrome P450 | - | - | 5.7927 | up |
| CL426.Contig5_All | CXE 2 | JHE-like carboxylesterase 2 | 4.3916 | up | 8.2016 | up |
| Immune response | ||||||
| CL2.Contig14_All | HC | hemocyanin | 7.0572 | up | 7.0599 | up |
| CL3394.Contig2_All | CTL | C-type lectin | 5.3396 | up | 3.9633 | up |
| CL4909.Contig1_All | FLP 2 | ficolin-like protein 2 | 1.7688 | up | 2.0525 | up |
| CL5288.Contig1_All | Domeless | domeless | 2.1386 | up | 2.1734 | up |
| CL607.Contig1_All | Stat | signal transducer and activator of transcription | 4.3341 | up | 1.5504 | up |
| CL4523.Contig2_All | TLR | Toll-like receptor | 3.8945 | up | 3.0839 | up |
| CL2513.Contig3_All | Relish | relish | 10.3394 | up | 9.2218 | up |
| DNA damage repair | ||||||
| CL4136.Contig1_All | DDB1 | DNA damage-binding protein 1 | −1.1834 | down | −1.3557 | down |
| Unigene25307_All | MLH1 | MutL Homolog 1 | - | - | −4.1634 | down |
| Unigene8072_All | MSH2 | MutS Homolog 2 | - | - | −2.0912 | down |
| Unigene15503_All | MSH6 | MutS Homolog 6 | - | - | −1.8564 | down |
| Unigene362_All | DDB2 | DNA damage-binding protein 2 | - | - | −1.8626 | down |
| Unigene27679_All | RFC2 | replication factor C subunit 2-like | - | - | −2.1294 | down |
| Unigene768_All | RFC4 | replication factor C subunit 4 | - | - | −2.1227 | down |
| CL6139.Contig1_All | RFC5 | replication factor C subunit 5-like | - | - | −2.5631 | down |
| Unigene17725_All | POLE2 | DNA polymerase epsilon subunit 2 | - | - | −3.4375 | down |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, J.; Bao, Z.; Guo, J.; Su, X.; Zou, Y.; Guo, H. Comparative Transcriptome Analysis of the Hepatopancreas from Macrobrachium rosenbergii Exposed to the Heavy Metal Copper. Animals 2024, 14, 1117. https://doi.org/10.3390/ani14071117
Zhang J, Bao Z, Guo J, Su X, Zou Y, Guo H. Comparative Transcriptome Analysis of the Hepatopancreas from Macrobrachium rosenbergii Exposed to the Heavy Metal Copper. Animals. 2024; 14(7):1117. https://doi.org/10.3390/ani14071117
Chicago/Turabian StyleZhang, Jiayuan, Zhiming Bao, Jieyu Guo, Xianbin Su, Yongfeng Zou, and Hui Guo. 2024. "Comparative Transcriptome Analysis of the Hepatopancreas from Macrobrachium rosenbergii Exposed to the Heavy Metal Copper" Animals 14, no. 7: 1117. https://doi.org/10.3390/ani14071117
APA StyleZhang, J., Bao, Z., Guo, J., Su, X., Zou, Y., & Guo, H. (2024). Comparative Transcriptome Analysis of the Hepatopancreas from Macrobrachium rosenbergii Exposed to the Heavy Metal Copper. Animals, 14(7), 1117. https://doi.org/10.3390/ani14071117