

Current Status of Trypanosoma grosi and Babesia microti in Small Mammals in the Republic of Korea

Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Ethical Approval
2.2. Small Mammal Sampling
2.3. Molecular Detection of Parasites in Small Mammals
2.4. Sequencing and Phylogenetic Analysis
2.5. Statistical Analysis
3. Results
3.1. Prevalence of Parasitic Protozoa in Small Mammals in the ROK
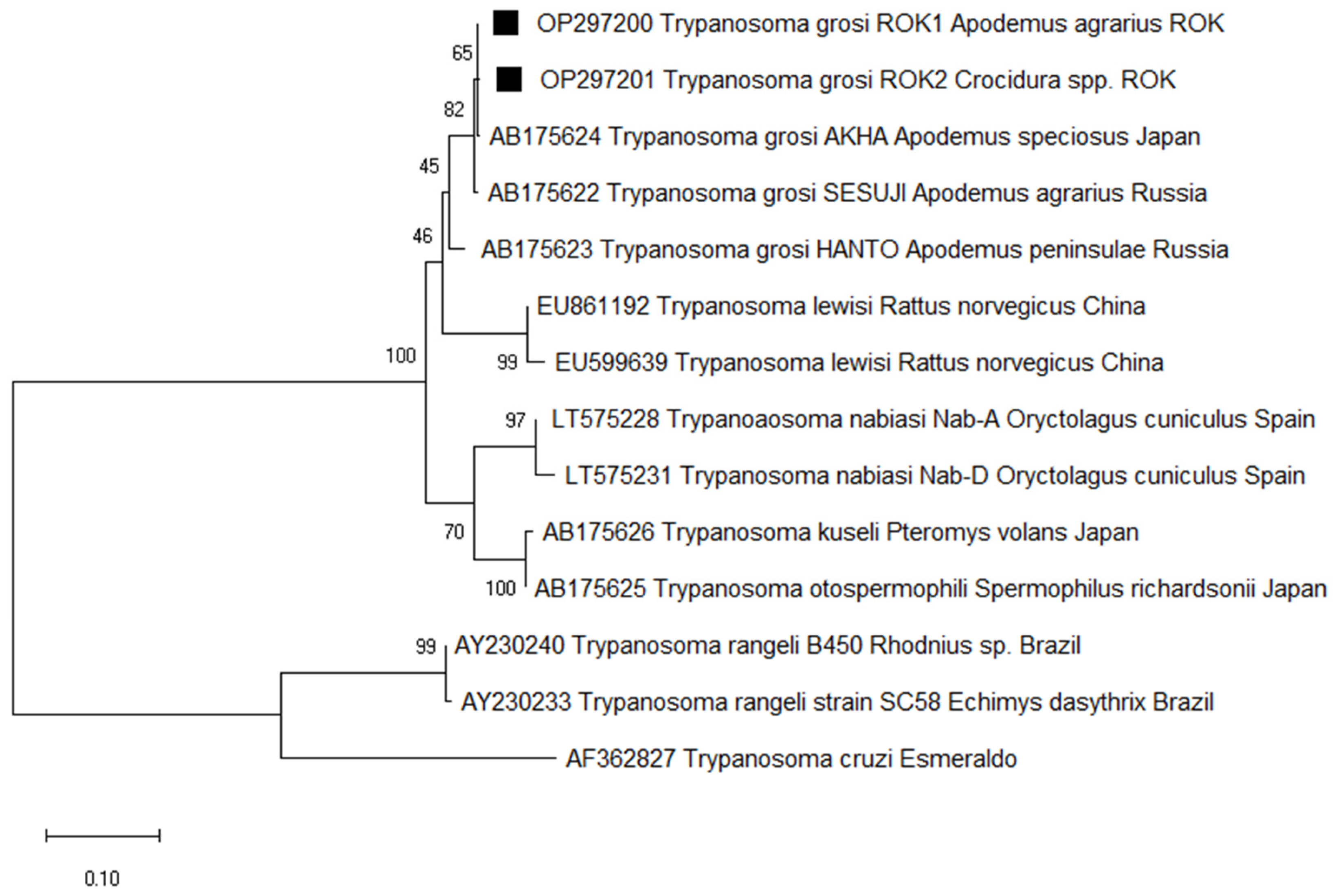
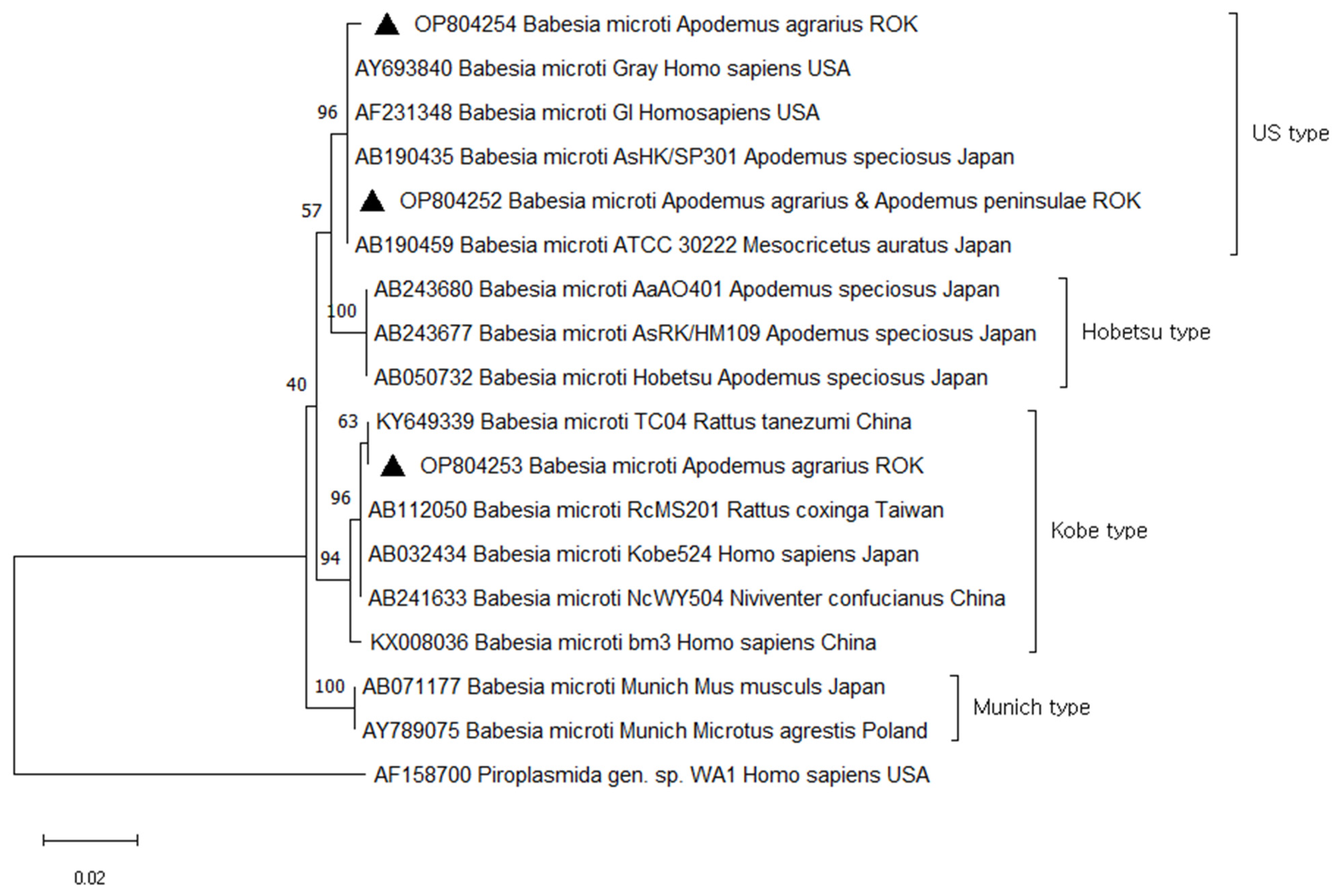
3.2. Prevalence and Phylogenetic Analysis of T. grosi and B. microti in Small Mammals
3.3. Relationships between Season, Ecological Habitat, and T. grosi and B. microti Infections
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Capizzi, D.; Bertolino, S.; Mortelliti, A. Rating the rat: Global patterns and research priorities in impacts and management of rodent pests. Mamm. Rev. 2014, 44, 148–162. [Google Scholar] [CrossRef]
- Han, B.A.; Schmidt, J.P.; Bowden, S.E.; Drake, J.M. Rodent reservoirs of future zoonotic diseases. Proc. Natl. Acad. Sci. USA 2015, 112, 7039–7044. [Google Scholar] [CrossRef] [PubMed]
- Morand, S.; Jittapalapong, S.; Kosoy, M. Rodents as hosts of infectious diseases: Biological and ecological characteristics. Vector Borne Zoonotic Dis. 2015, 15, 1. [Google Scholar] [CrossRef]
- Jahan, N.A.; Lindsey, L.L.; Larsen, P.A. The role of peridomestic rodents as reservoirs for zoonotic foodborne pathogens. Vector-Borne Zoonotic Dis. 2021, 21, 133–148. [Google Scholar] [CrossRef] [PubMed]
- Shimi, A.; Keyhani, M.; Hedayati, K. Studies on salmonellosis in the house mouse, Mus musculus. Lab. Anim. 1979, 13, 33–34. [Google Scholar] [CrossRef] [PubMed]
- Meerburg, B.G. Rodents are a risk factor for the spreading of pathogens on farms. Vet. Microbiol. 2010, 142, 464–465. [Google Scholar] [CrossRef]
- Dahmana, H.; Granjon, L.; Diagne, C.; Davoust, B.; Fenollar, F.; Mediannikov, O. Rodents as hosts of pathogens and related zoonotic disease risk. Pathogens 2020, 9, 202. [Google Scholar] [CrossRef] [PubMed]
- Rabiee, M.H.; Mahmoudi, A.; Siahsarvie, R.; Kryštufek, B.; Mostafavi, E. Rodent-borne diseases and their public health importance in Iran. PLoS Negl. Trop. Dis. 2018, 12, e0006256. [Google Scholar] [CrossRef] [PubMed]
- Seifollahi, Z.; Sarkari, B.; Motazedian, M.H.; Asgari, Q.; Ranjbar, M.J.; Abdolahi Khabisi, S. Protozoan parasites of rodents and their zoonotic significance in Boyer-Ahmad District, Southwestern Iran. Vet. Med. Int. 2016, 2016, 3263868. [Google Scholar] [CrossRef]
- Tijjani, M.; Majid, R.A.; Abdullahi, S.A.; Unyah, N.Z. Detection of rodent-borne parasitic pathogens of wild rats in Serdang, Selangor, Malaysia: A potential threat to human health. Int. J. Parasitol. Parasit. Wildl. 2020, 11, 174–182. [Google Scholar] [CrossRef]
- Pumhom, P.; Pognon, D.; Yangtara, S.; Thaprathorn, N.; Milocco, C.; Douangboupha, B.; Herder, S.; Chaval, Y.; Morand, S.; Jittapalapong, S.; et al. Molecular prevalence of Trypanosoma spp. in wild rodents of Southeast Asia: Influence of human settlement habitat. Epidemiol. Infect. 2014, 142, 1221–1230. [Google Scholar] [CrossRef] [PubMed]
- Bishop, R.; Musoke, A.; Morzaria, S.; Gardner, M.; Nene, V. Theileria: Intracellular protozoan parasites of wild and domestic ruminants transmitted by ixodid ticks. Parasitology 2004, 129, S271–S283. [Google Scholar] [CrossRef] [PubMed]
- Krause, P.J. Human babesiosis. Int. J. Parasitol. 2019, 49, 165–174. [Google Scholar] [CrossRef] [PubMed]
- Uilenberg, G. International collaborative research: Significance of tick-borne hemoparasitic diseases to world animal health. Vet. Parasitol. 1995, 57, 19–41. [Google Scholar] [CrossRef]
- Duh, D.; Punda-Polić, V.; Trilar, T.; Avsic-Zupanc, T. Molecular detection of Theileria sp. in ticks and naturally infected sheep. Vet. Parasitol. 2008, 151, 327–331. [Google Scholar] [CrossRef] [PubMed]
- Homer, M.J.; Aguilar-Delfin, I.; Telford, S.R., 3rd; Krause, P.J.; Persing, D.H. Babesiosis. Clin. Microbiol. Rev. 2000, 13, 451–469. [Google Scholar] [CrossRef] [PubMed]
- Krause, P.J. Babesiosis diagnosis and treatment. Vector Borne Zoonotic Dis. 2003, 3, 45–51. [Google Scholar] [CrossRef] [PubMed]
- Swanson, M.; Pickrel, A.; Williamson, J.; Montgomery, S. Trends in Reported Babesiosis Cases—United States, 2011–2019. MMWR Morb. Mortal Wkly. Rep. 2023, 72, 273–277. [Google Scholar] [CrossRef] [PubMed]
- Young, K.M.; Corrin, T.; Wilhelm, B.; Uhland, C.; Greig, J.; Mascarenhas, M.; Waddell, L.A. Zoonotic Babesia: A scoping review of the global evidence. PLoS ONE 2019, 14, e0226781. [Google Scholar] [CrossRef]
- Hussain, S.; Hussain, A.; Aziz, M.U.; Song, B.; Zeb, J.; George, D.; Li, J.; Sparagano, O. A review of zoonotic babesiosis as an emerging public health threat in Asia. Pathogens 2022, 11, 23. [Google Scholar] [CrossRef]
- Desquesnes, M.; Ravel, S.; Cuny, G. PCR identification of Trypanosoma lewisi, a common parasite of laboratory rats. Kinetoplastid Biol. Dis. 2002, 1, 2. [Google Scholar] [CrossRef]
- Goodrich, I.; McKee, C.; Kosoy, M. Trypanosoma (Herpetosoma) diversity in rodents and lagomorphs of New Mexico with a focus on epizootological aspects of infection in Southern Plains woodrats (Neotoma micropus). PLoS ONE 2020, 15, e0244803. [Google Scholar] [CrossRef]
- Desquesnes, M.; Gonzatti, M.; Sazmand, A.; Thévenon, S.; Bossard, G.; Boulangé, A.; Gimonneau, G.; Truc, P.; Herder, S.; Ravel, S.; et al. A review on the diagnosis of animal trypanosomoses. Parasit. Vector. 2022, 15, 64. [Google Scholar] [CrossRef]
- Nussbaum, K.; Honek, J.; Cadmus, C.M.; Efferth, T. Trypanosomatid parasites causing neglected diseases. Curr. Med. Chem. 2010, 17, 1594–1617. [Google Scholar] [CrossRef]
- Votýpka, J.; d’Avila-Levy, C.M.; Grellier, P.; Maslov, D.A.; Lukeš, J.; Yurchenko, V. New approaches to systematics of Trypanosomatidae: Criteria for taxonomic (re)description. Trends Parasitol. 2015, 31, 460–469. [Google Scholar] [CrossRef] [PubMed]
- Magri, A.; Galuppi, R.; Fioravanti, M. Autochthonous Trypanosoma spp. in European mammals: A brief journey amongst the neglected trypanosomes. Pathogens 2021, 10, 334. [Google Scholar] [CrossRef]
- Hoare, C.A. The trypanosomes of mammals. A zoological monograph. Med. J. Aust. 1972, 1, 140. [Google Scholar] [CrossRef]
- Cox, F.E.G. Protozoan parasites of British small rodents. Mamm. Rev. 1987, 17, 59–66. [Google Scholar] [CrossRef]
- Sato, H.; Osanai, A.; Kamiya, H.; Obara, Y.; Jiang, W.; Zhen, Q.; Chai, J.; Une, Y.; Ito, M. Characterization of SSU and LSU rRNA genes of three Trypanosoma (Herpetosoma) grosi isolates maintained in Mongolian jirds. Parasitology 2005, 130, 157–167. [Google Scholar] [CrossRef]
- Guan, G.; Niu, Q.; Yang, J.; Li, Y.; Gao, J.; Luo, J.; Yin, H. Trypanosoma (Herpetosoma) grosi: First isolation from Chinese striped field mouse (Apodemus agrarius). Parasitol. Int. 2011, 60, 101–104. [Google Scholar] [CrossRef]
- Howie, S.; Guy, M.; Fleming, L.; Bailey, W.; Noyes, H.; Faye, J.A.; Pepin, J.; Greenwood, B.; Whittle, H.; Molyneux, D.; et al. A Gambian infant with fever and an unexpected blood film. PLoS Med. 2006, 3, 1508–1512. [Google Scholar] [CrossRef] [PubMed]
- Sarataphan, N.; Vongpakorn, M.; Nuansrichay, B.; Autarkool, N.; Keowkarnkah, T.; Rodtian, P.; Stich, R.W.; Jittapalapong, S. Diagnosis of a Trypanosoma lewisi-like (Herpetosoma) infection in a sick infant from Thailand. J. Med. Microbiol. 2007, 56, 1118–1121. [Google Scholar] [CrossRef] [PubMed]
- Truc, P.; Büscher, P.; Cuny, G.; Gonzatti, M.I.; Jannin, J.; Joshi, P.; Juyal, P.; Lun, Z.-R.; Mattioli, R.; Pays, E.; et al. Atypical human infections by animal trypanosomes. PLoS Negl. Trop. Dis. 2013, 7, e2256. [Google Scholar] [CrossRef] [PubMed]
- Zamoto, A.; Tsuji, M.; Wei, Q.; Cho, S.H.; Shin, E.H.; Kim, T.S.; Leonova, G.N.; Hagiwara, K.; Asakawa, M.; Kariwa, H.; et al. Epizootiologic survey for Babesia microti among small wild mammals in northeastern Eurasia and a geographic diversity in the beta-tubulin gene sequences. J. Vet. Med. Sci. 2004, 66, 785–792. [Google Scholar] [CrossRef] [PubMed]
- Hong, S.H.; Lee, S.E.; Jeong, Y.I.; Kim, H.C.; Chong, S.T.; Klein, T.A.; Song, J.W.; Gu, S.H.; Cho, S.H.; Lee, W.J. Prevalence and molecular characterizations of Toxoplasma gondii and Babesia microti from small mammals captured in Gyeonggi and Gangwon Provinces, Republic of Korea. Vet. Parasitol. 2014, 205, 512–517. [Google Scholar] [CrossRef]
- Kang, S.W.; Doan, H.T.; Choe, S.E.; Noh, J.H.; Yoo, M.S.; Reddy, K.E.; Kim, Y.H.; Kweon, C.H.; Jung, S.C.; Chang, K.Y. Molecular investigation of tick-borne pathogens in ticks from grazing cattle in Korea. Parasitol. Int. 2013, 62, 276–282. [Google Scholar] [CrossRef] [PubMed]
- Hong, S.H.; Kim, S.Y.; Song, B.G.; Rho, J.R.; Cho, C.R.; Kim, C.N.; Um, T.H.; Kwak, Y.G.; Cho, S.H.; Lee, S.E. Detection and characterization of an emerging type of Babesia sp. similar to Babesia motasi for the first case of human babesiosis and ticks in Korea. Emerg. Microbes Infect. 2019, 8, 869–878. [Google Scholar] [CrossRef]
- Kim, T.Y.; Kim, S.Y.; Kim, T.K.; Lee, H.I.; Cho, S.H.; Lee, W.G.; Kim, H. Molecular evidence of zoonotic Babesia species, other than B. microti, in ixodid ticks collected from small mammals in the Republic of Korea. Vet. Med. Sci. 2021, 7, 2427–2433. [Google Scholar] [CrossRef] [PubMed]
- Bang, M.S.; Kim, C.M.; Pyun, S.H.; Kim, D.M.; Yun, N.R. Molecular investigation of tick-borne pathogens in ticks removed from tick-bitten humans in the southwestern region of the Republic of Korea. PLoS ONE 2021, 16, e0252992. [Google Scholar] [CrossRef]
- Kim, T.Y.; Kim, S.Y.; Seo, J.; Lee, H.I.; Lee, W.G.; Kim, H. Detection of US and Kobe-type Babesia microti in ticks collected from small mammals of the Republic of Korea. Entomol. Res. 2024, 54, e12708. [Google Scholar] [CrossRef]
- Kwon, H.Y.; Im, J.H.; Park, Y.K.; Durey, A.; Lee, J.S.; Baek, J.H. Two Imported Cases of Babesiosis with Complication or Co-Infection with Lyme Disease in Republic of Korea. Korean J. Parasitol. 2018, 56, 609–613. [Google Scholar] [CrossRef] [PubMed]
- Na, Y.J.; Chai, J.Y.; Jung, B.K.; Lee, H.J.; Song, J.Y.; Je, J.H.; Seo, J.H.; Park, S.H.; Choi, J.S.; Kim, M.J. An Imported Case of Severe falciparum Malaria with Prolonged Hemolytic Anemia Clinically Mimicking a Coinfection with Babesiosis. Korean J. Parasitol. 2014, 52, 667–672. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.Y.; Cho, S.H.; Joo, H.N.; Tsuji, M.; Cho, S.R.; Park, I.J.; Chung, G.T.; Ju, J.W.; Cheun, H.I.; Lee, H.W.; et al. First Case of Human Babesiosis in Korea: Detection and Characterization of a Novel Type of Babesia sp. (KO1) Similar to Ovine Babesia. J. Clin. Microbiol. 2007, 45, 2084–2087. [Google Scholar] [CrossRef] [PubMed]
- Okal, M.N.; Odhiambo, B.K.; Otieno, P.; Bargul, J.L.; Masiga, D.; Villinger, J.; Kalayou, S. Anaplasma and Theileria Pathogens in Cattle of Lambwe Valley, Kenya: A Case for Pro-Active Surveillance in the Wildlife–Livestock Interface. Microorganisms 2020, 8, 1830. [Google Scholar] [CrossRef] [PubMed]
- Taylor, L.H.; Welburn, S.C.; Woolhouse, M.E. Theileria annulata: Virulence and transmission from single and mixed clone infections in cattle. Exp. Parasitol. 2002, 100, 186–195. [Google Scholar] [CrossRef] [PubMed]
- Egan, S.L.; Taylor, C.L.; Austen, J.M.; Banks, P.B.; Northover, A.S.; Ahlstrom, L.A.; Ryan, U.M.; Irwin, P.J.; Oskam, C.L. Haemoprotozoan surveillance in peri-urban native and introduced wildlife from Australia. Curr. Res. Parasitol. Vector-Borne Dis. 2021, 1, 100052. [Google Scholar] [CrossRef]
- Mangombi, J.B.; N’dilimabaka, N.; Lekana-Douki, J.B.; Banga, O.; Maghendji-Nzondo, S.; Bourgarel, M.; Leroy, E.; Fenollar, F.; Mediannikov, O. First investigation of pathogenic bacteria, protozoa and viruses in rodents and shrews in context of forest-savannah-urban areas interface in the city of Franceville (Gabon). PLoS ONE 2021, 16, e0248244. [Google Scholar] [CrossRef]
- Sakka, H.; Quéré, J.P.; Kartavtseva, I.; Pavlenko, M.; Chelomina, G.; Atopkin, D.; Bogdanov, A.; Michaux, J. Comparative phylogeography of four Apodemus species (Mammalia: Rodentia) in the Asian Far East: Evidence of Quaternary climatic changes in their genetic structure. Biol. J. Linn. Soc. 2010, 100, 797–821. [Google Scholar] [CrossRef]
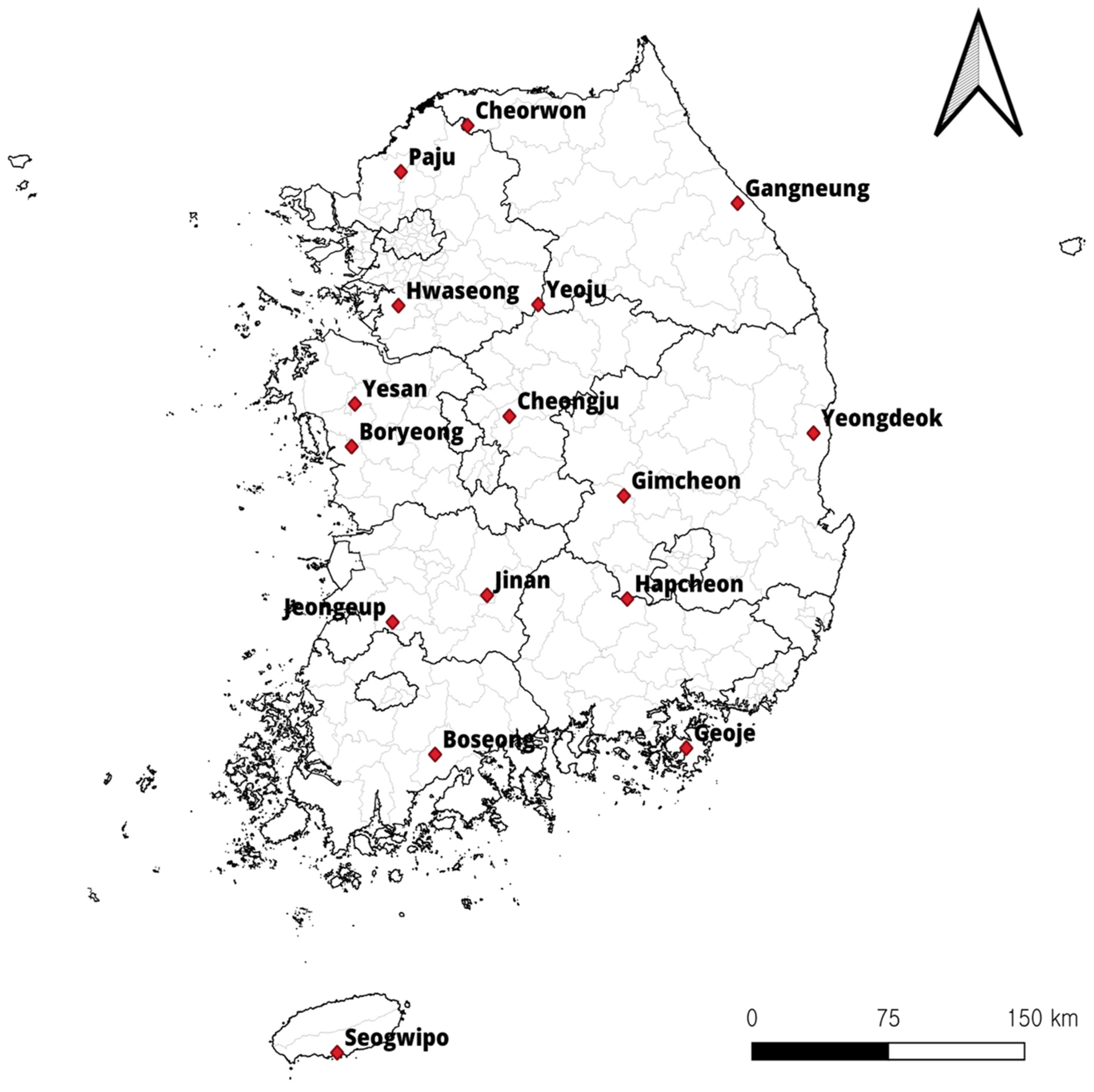
 ) in the Republic of Korea (ROK). Map was created using the Free and Open Source QGIS (QGIS 3.28.10. Geographic Information System, http://www.qgis.org, accesed on 8 September 2023).
) in the Republic of Korea (ROK). Map was created using the Free and Open Source QGIS (QGIS 3.28.10. Geographic Information System, http://www.qgis.org, accesed on 8 September 2023).
) in the Republic of Korea (ROK). Map was created using the Free and Open Source QGIS (QGIS 3.28.10. Geographic Information System, http://www.qgis.org, accesed on 8 September 2023).
) in the Republic of Korea (ROK). Map was created using the Free and Open Source QGIS (QGIS 3.28.10. Geographic Information System, http://www.qgis.org, accesed on 8 September 2023).



{kind=link}
{kind=link}
{kind=link}
| Site | No. Tested | No. T. grosi- Positive (%) | No. B. microti- Positive (%) | B. microti Genotype |
|---|---|---|---|---|
| Gangneung-si | 43 | 11 (25.6) | 14 (32.6) §§ | US type |
| Hwaseong-si | 33 | 12 (36.4) | 1 (3.0) | US type |
| Seogwipo-si | 31 | 15 (48.4) § | 2 (6.5) | Kobe type |
| Jinan-gun | 24 | 5 (20.8) | 2 (8.3) | US type |
| Cheorwon-gun | 23 | 6 (64.7) | 1 (4.3) | US type |
| Geoje-si | 23 | 6 (26.1) | 0 (0.0) | |
| Yeoju-si | 22 | 3 (13.6) | 0 (0.0) | |
| Yeongdeok-gun | 20 | 4 (20.0) | 2 (10.0) | US type |
| Paju-si | 19 | 6 (31.6) | 6 (31.6) | US type |
| Jeongeup-si | 18 | 1 (5.6) | 2 (11.1) | US type |
| Gimcheon-si | 18 | 1 (5.6) | 0 (0.0) | |
| Boseong-gun | 17 | 1 (5.9) | 1 (5.9) | US type |
| Hapcheon-gun | 16 | 1 (6.3) | 2 (12.5) | US type |
| Yesan-si | 14 | 2 (14.3) | 0 (0.0) | |
| Boryeong-si | 8 | 0 (0.0) | 0 (0.0) | |
| Cheongju-si | 8 | 0 (0.0) | 0 (0.0) | |
| Total | 331 | 79 (23.9) | 33 (10.0) |
| Small Mammal Species | No. Tested | No. of T. grosi- Positive (%) | No. of B. microti- Positive (%) | No. of Coinfection (%) |
|---|---|---|---|---|
| Apodemus agrarius | 290 | 77 (26.5) | 32 (11.0) | 6 (2.0) |
| Crocidura sp. | 23 | 2 (8.7) | 0 | 0 |
| Apodemus peninsulae | 2 | 0 | 1 (50.0) | 0 |
| Craseomys regulus | 4 | 0 | 0 | 0 |
| Craseomys rufocanus | 1 | 0 | 0 | 0 |
| Cricetulus triton | 1 | 0 | 0 | 0 |
| Micromys minutus | 6 | 0 | 0 | 0 |
| Microtus fortis | 2 | 0 | 0 | 0 |
| Myodes regulus | 1 | 0 | 0 | 0 |
| Rattus norvegicus | 1 | 0 | 0 | 0 |
| Total | 331 | 79 (23.9) | 33 (10.0) | 6 (1.8) |
| Categories | No. Tested | No. of T. grosi- Positive (%) | p-Value | No. of B. microti- Positive (%) | p-Value | |
|---|---|---|---|---|---|---|
| Season | Spring | 174 | 46 (26.4) | 0.248 | 17 (9.8) | 0.898 |
| Autumn | 157 | 33 (21.0) | 16 (10.2) | |||
| Ecological habitat | Reservoir | 70 | 8 (11.4) | 0.006 | 9 (12.8) | 0.363 |
| Other | 261 | 71(27.2) | 24(9.2) | |||
| Small mammal species | A. agrarius | 290 | 77 (26.5) | 0.0014 | 32 (11.0) | 0.098 |
| Other | 41 | 2 (4.8) | 1 (2.4) | |||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, H.J.; Han, B.; Lee, H.-I.; Ju, J.-W.; Shin, H.-I. Current Status of Trypanosoma grosi and Babesia microti in Small Mammals in the Republic of Korea. Animals 2024, 14, 989. https://doi.org/10.3390/ani14070989
Kim HJ, Han B, Lee H-I, Ju J-W, Shin H-I. Current Status of Trypanosoma grosi and Babesia microti in Small Mammals in the Republic of Korea. Animals. 2024; 14(7):989. https://doi.org/10.3390/ani14070989
Chicago/Turabian StyleKim, Hyun Jung, BoGyeong Han, Hee-Il Lee, Jung-Won Ju, and Hyun-Il Shin. 2024. "Current Status of Trypanosoma grosi and Babesia microti in Small Mammals in the Republic of Korea" Animals 14, no. 7: 989. https://doi.org/10.3390/ani14070989
APA StyleKim, H. J., Han, B., Lee, H. -I., Ju, J. -W., & Shin, H. -I. (2024). Current Status of Trypanosoma grosi and Babesia microti in Small Mammals in the Republic of Korea. Animals, 14(7), 989. https://doi.org/10.3390/ani14070989