
Dietary Peppermint Extract Inhibits Chronic Heat Stress-Induced Activation of Innate Immunity and Inflammatory Response in the Spleen of Broiler Chickens
Abstract
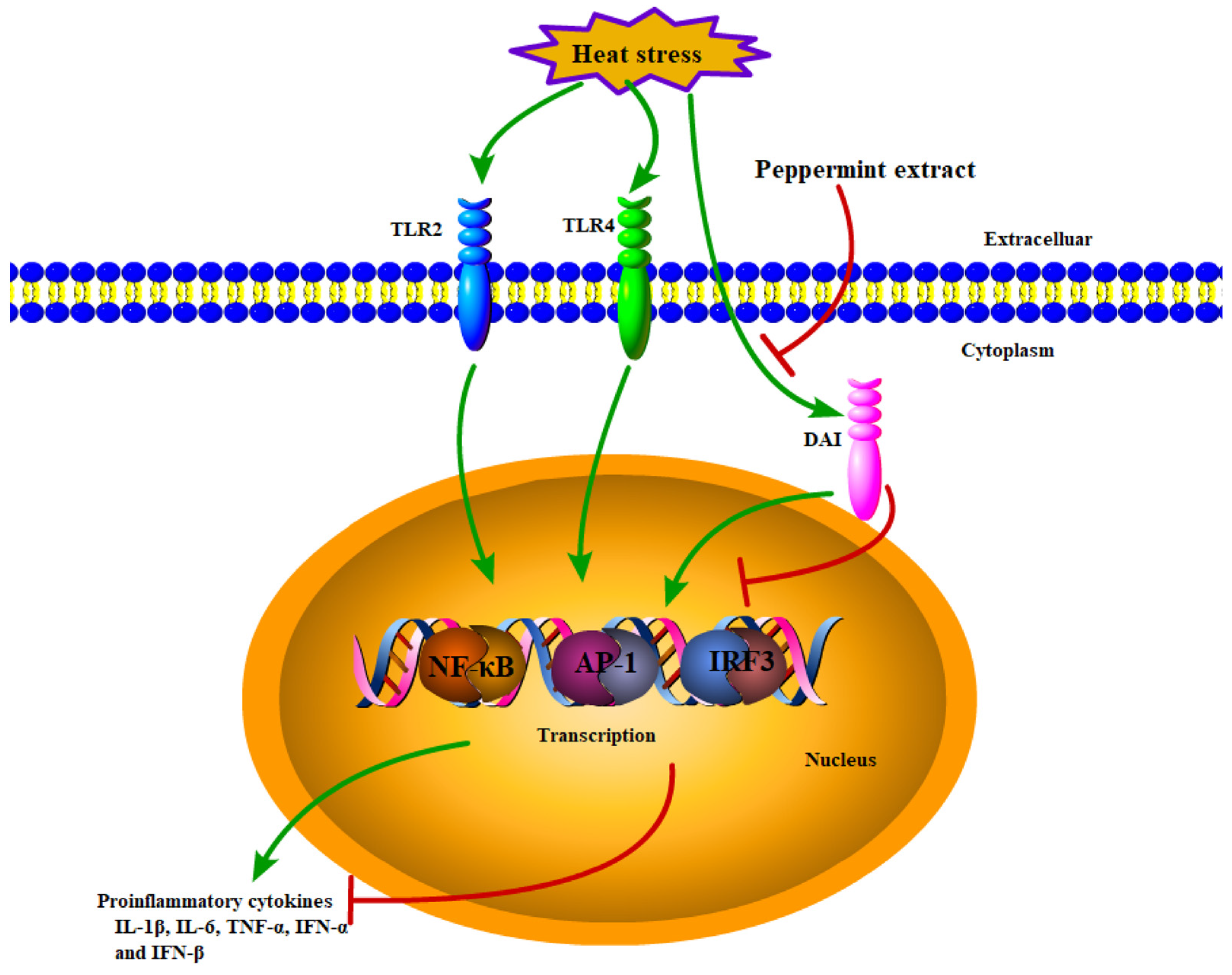
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animal Management
2.2. Sample Collection
2.3. Histochemical Analysis
2.4. Enzyme-Linked Immunosorbent Assay (ELISA) Analysis
2.5. Gene Expression Analysis
2.6. Statistical Analysis
3. Results
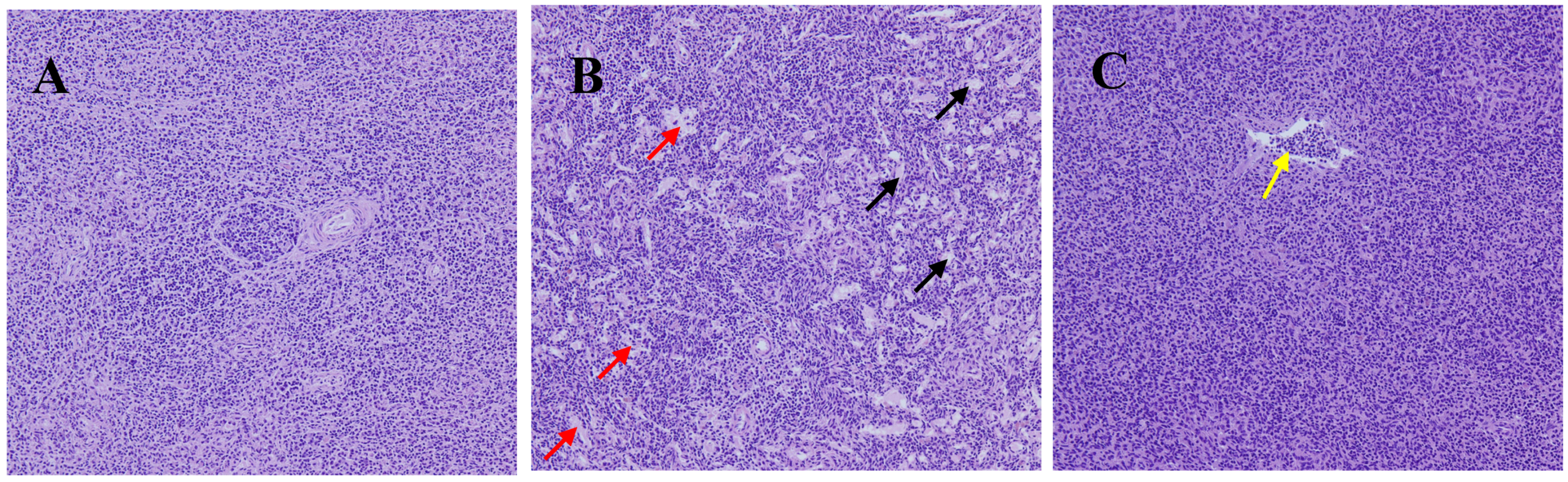
3.1. Peppermint Extract Alleviated the Spleen Tissue Damage Induced by Chronic Heats Stress
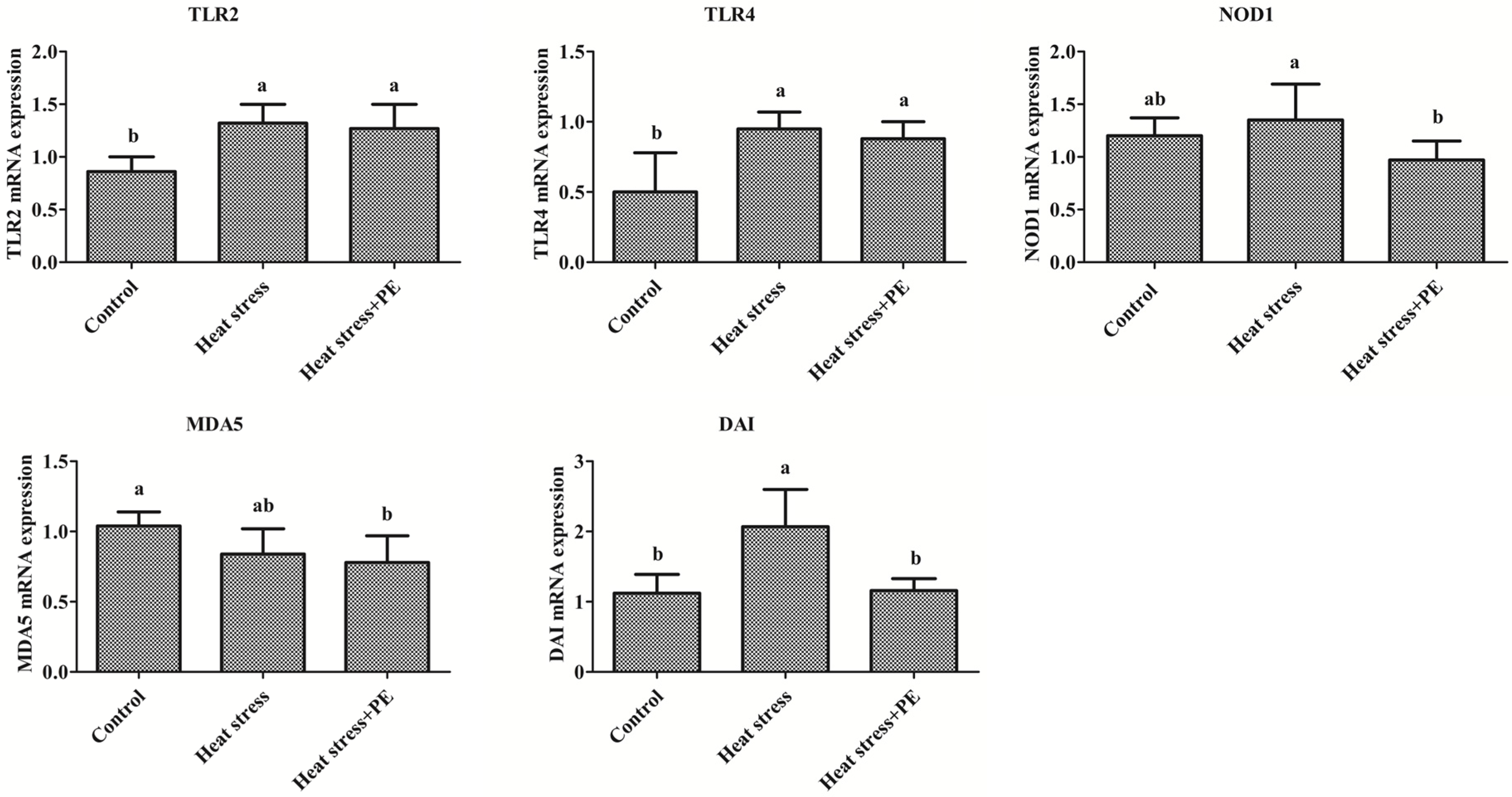
3.2. Peppermint Extract Regulated the Expression of Main Pattern Recognition Receptors under Chronic Heat Stress
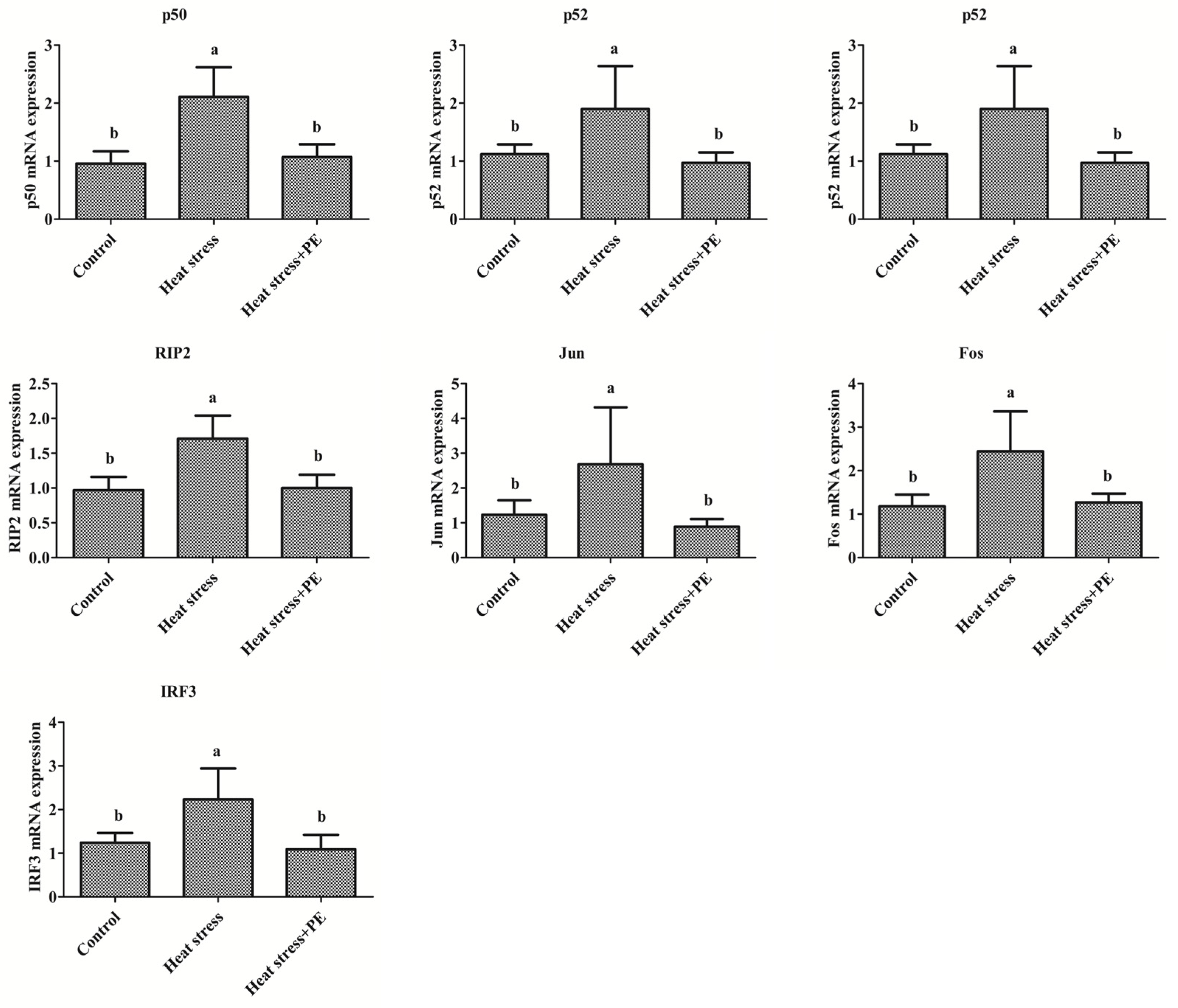
3.3. Peppermint Extract Reduced the Increase of Transcription Factors mRNA Expression Induced by Chronic Heat Stress
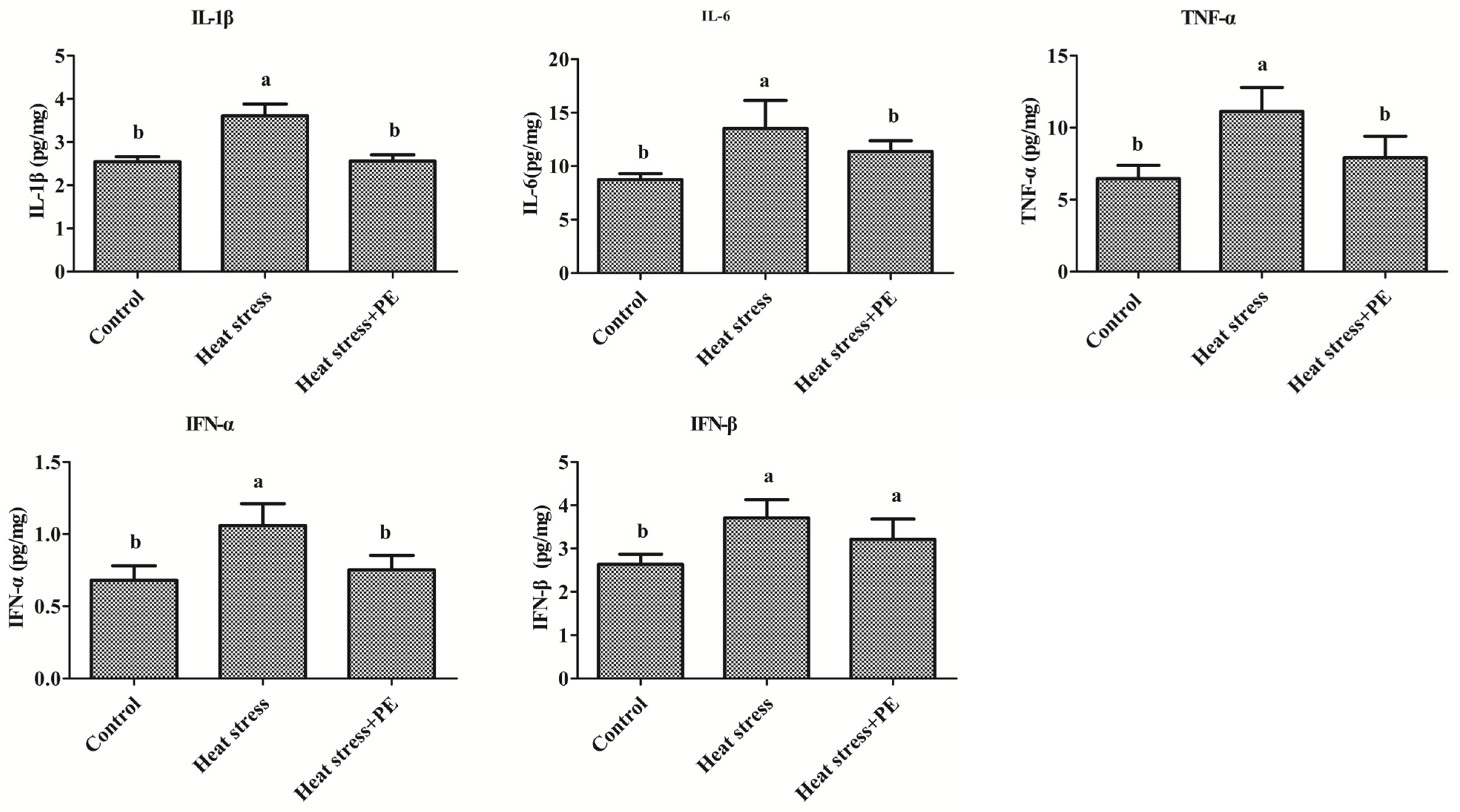
3.4. Peppermint Extract Reduced the Increase of Inflammatory Cytokines Concentration Induced by Chronic Heat Stress
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Lara, L.J.; Rostagno, M.H. Impact of Heat Stress on Poultry Production. Animals 2013, 3, 356–369. [Google Scholar] [CrossRef] [PubMed]
- Rostagno, M.H. Can stress in farm animals increase food safety risk? Foodborne Pathog. Dis. 2009, 6, 767–776. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Z.; Luo, M.; Ma, W.; Ma, S.; Wang, Y.; Zhang, K. Protective Effects of 1,8-Cineole Microcapsules Against Inflammation and Gut Microbiota Imbalance Associated Weight Loss Induced by Heat Stress in Broiler Chicken. Front. Pharmacol. 2020, 11, 585945. [Google Scholar] [CrossRef] [PubMed]
- Bartlett, J.R.; Smith, M.O. Effects of different levels of zinc on the performance and immunocompetence of broilers under heat stress. Poult. Sci. 2003, 82, 1580–1588. [Google Scholar] [CrossRef] [PubMed]
- Zmrhal, V.; Svoradova, A.; Venusova, E.; Slama, P. The Influence of Heat Stress on Chicken Immune System and Mitigation of Negative Impacts by Baicalin and Baicalein. Animals 2023, 13, 2564. [Google Scholar] [CrossRef]
- Inbaraj, S.; Sejian, V.; Bagath, M.; Bhatta, R. Impact of Heat Stress on Immune Responses of Livestock: A Review. Pertanika J. Trop. Agric. Sci. 2016, 39, 459. [Google Scholar]
- Xu, D.; Li, W.; Huang, Y.; He, J.; Tian, Y. The effect of selenium and polysaccharide of Atractylodes macrocephala Koidz. (PAMK) on immune response in chicken spleen under heat stress. Biol. Trace Elem. Res. 2014, 160, 232–237. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Li, M.; Li, X.; Zhou, Y.; Feng, J.; Zhang, M. Effect of relative humidity and intermittent partial heat temperature on immune function of broiler. Chin. J. Anim. Nutr. 2017, 29, 80–87. [Google Scholar]
- Akira, S.; Uematsu, S.; Takeuchi, O. Pathogen recognition and innate immunity. Cell 2006, 124, 783–801. [Google Scholar] [CrossRef]
- Takeuchi, O.; Akira, S. Pattern recognition receptors and inflammation. Cell 2010, 140, 805–820. [Google Scholar] [CrossRef]
- Ma, D.; Liu, Q.; Zhang, M.; Feng, J.; Li, X.; Zhou, Y.; Wang, X. iTRAQ-based quantitative proteomics analysis of the spleen reveals innate immunity and cell death pathways associated with heat stress in broilers (Gallus gallus). J. Proteom. 2019, 196, 11–21. [Google Scholar] [CrossRef] [PubMed]
- Takeda, K.; Akira, S. TLR signaling pathways. Semin. Immunol. 2004, 16, 3–9. [Google Scholar] [CrossRef] [PubMed]
- Berghäll, H.; Sirén, J.; Sarkar, D.; Julkunen, I.; Fisher, P.B.; Vainionpää, R.; Matikainen, S. The interferon-inducible RNA helicase, mda-5, is involved in measles virus-induced expression of antiviral cytokines. Microbes Infect. 2006, 8, 2138–2144. [Google Scholar] [CrossRef] [PubMed]
- Komuro, A.; Horvath, C.M. RNA- and virus-independent inhibition of antiviral signaling by RNA helicase LGP2. J. Virol. 2006, 80, 12332–12342. [Google Scholar] [CrossRef] [PubMed]
- Lin, R.; Lacoste, J.; Nakhaei, P.; Sun, Q.; Yang, L.; Paz, S.; Wilkinson, P.; Julkunen, I.; Vitour, D.; Meurs, E.; et al. Dissociation of a MAVS/IPS-1/VISA/Cardif-IKKepsilon molecular complex from the mitochondrial outer membrane by hepatitis C virus NS3-4A proteolytic cleavage. J. Virol. 2006, 80, 6072–6083. [Google Scholar] [CrossRef] [PubMed]
- Seth, R.B.; Sun, L.; Ea, C.K.; Chen, Z.J. Identification and characterization of MAVS, a mitochondrial antiviral signaling protein that activates NF-kappaB and IRF 3. Cell 2005, 122, 669–682. [Google Scholar] [CrossRef]
- Xu, L.G.; Wang, Y.Y.; Han, K.J.; Li, L.Y.; Zhai, Z.; Shu, H.B. VISA is an adapter protein required for virus-triggered IFN-beta signaling. Mol. Cell. 2005, 19, 727–740. [Google Scholar] [CrossRef]
- Soulat, D.; Bürckstümmer, T.; Westermayer, S.; Goncalves, A.; Bauch, A.; Stefanovic, A.; Hantschel, O.; Bennett, K.L.; Decker, T.; Superti-Furga, G. The DEAD-box helicase DDX3X is a critical component of the TANK-binding kinase 1-dependent innate immune response. EMBO J. 2008, 27, 2135–2146. [Google Scholar] [CrossRef]
- Yu, L.; Zhang, X.; Wu, T.; Su, J.; Wang, Y.; Wang, Y.; Ruan, B.; Niu, X.; Wu, Y. Avian infectious bronchitis virus disrupts the melanoma differentiation associated gene 5 (MDA5) signaling pathway by cleavage of the adaptor protein MAVS. BMC Vet. Res. 2017, 13, 332. [Google Scholar] [CrossRef]
- Upton, J.W.; Kaiser, W.J.; Mocarski, E.S. DAI/ZBP1/DLM-1 Complexes with RIP3 to Mediate Virus-Induced Programmed Necrosis that Is Targeted by Murine Cytomegalovirus vIRA. Cell Host Microbe 2019, 26, 564. [Google Scholar] [CrossRef]
- Udawatte, D.J.; Rothman, A.L. Viral Suppression of RIPK1-Mediated Signaling. mBio 2021, 12, e0172321. [Google Scholar] [CrossRef] [PubMed]
- Ponnusamy, K.; Tzioni, M.M.; Begum, M.; Robinson, M.E.; Caputo, V.S.; Katsarou, A.; Trasanidis, N.; Xiao, X.; Kostopoulos, I.V.; Iskander, D.; et al. The innate sensor ZBP1-IRF3 axis regulates cell proliferation in multiple myeloma. Haematologica 2022, 107, 721–732. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Deng, H.; Cui, H.; Fang, J.; Zuo, Z.; Deng, J.; Li, Y.; Wang, X.; Zhao, L. Inflammatory responses and inflammation-associated diseases in organs. Oncotarget 2018, 9, 7204–7218. [Google Scholar] [CrossRef] [PubMed]
- Lee, D.W.; Ramakrishnan, D.; Valenta, J.; Parney, I.F.; Bayless, K.J.; Sitcheran, R. The NF-κB RelB protein is an oncogenic driver of mesenchymal glioma. PLoS ONE 2013, 8, e57489. [Google Scholar] [CrossRef] [PubMed]
- Karin, M.; Liu, Z.; Zandi, E. AP-1 function and regulation. Curr. Opin. Cell Biol. 1997, 9, 240–246. [Google Scholar] [CrossRef]
- Hamedi, S.; Shomali, T.; Ghaderi, H. Effect of dietary inclusion of Mentha piperita L. on histological and histomorphometrical parameters of the small intestine in broiler chickens. Org. Agric. 2017, 7, 105–110. [Google Scholar] [CrossRef]
- Arab Ameri, S.; Samadi, F.; Dastar, B.; Zarehdaran, S. Efficiency of Peppermint (Mentha piperita) Powder on Performance, Body Temperature and Carcass Characteristics of Broiler Chickens in Heat Stress Condition. Iran. J. Appl. Anim. Sci. 2016, 6, 943–950. [Google Scholar]
- Darabighane, B.; Aghjehgheshlagh, F.M.; Navidshad, B.; Mahdavi, A.; Zarei, A.; Nahashon, S.N. Effects of Peppermint (Mentha piperita) and Aloe vera (Aloe barbadensis) on Ileum Microflora Population and Growth Performance of Broiler Chickens in Comparison with Antibiotic Growth Promoter. Iran. J. Appl. Anim. Sci. 2017, 7, 101–108. [Google Scholar]
- Yarmohammadi Barbarestani, S.; Samadi, F.; Hassani, S.; Asadi, G. Effects of Encapsulated Nano- and Microparticles of Peppermint (Mentha piperita) Alcoholic Extract on the Growth Performance, Blood Parameters and Immune Function of Broilers under Heat Stress Condition. Iran. J. Appl. Anim. Sci. 2017, 7, 669–677. [Google Scholar]
- Mustafa, M.A.G.H.; Mahdi, S.; Bakr, B.N. Using of dried betony and peppermint in broiler diets on performance, hematological and immunity response. Diyala Agric. Sci. J. 2013, 5, 1–9. [Google Scholar]
- Rozza, A.L.; de Faria, F.M.; Brito, A.R.S.; Pellizzon, C.H. The gastroprotective effect of menthol: Involvement of anti-apoptotic, antioxidant and anti-inflammatory activities. PLoS ONE 2014, 9, e86686. [Google Scholar] [CrossRef] [PubMed]
- Shahid, M.; Lee, M.Y.; Yeon, A.; Cho, E.; Sairam, V.; Valdiviez, L.; You, S.; Kim, J. Menthol, a unique urinary volatile compound, is associated with chronic inflammation in interstitial cystitis. Sci. Rep. 2018, 8, 10859. [Google Scholar] [CrossRef] [PubMed]
- Mebius, R.E.; Kraal, G. Structure and function of the spleen. Nat. Rev. Immunol. 2005, 5, 606–616. [Google Scholar] [CrossRef] [PubMed]
- Thaxton, P. Influence of temperature on the immune response of birds. Poult. Sci. 1978, 57, 1430–1440. [Google Scholar] [CrossRef] [PubMed]
- He, S.; Yu, Q.; He, Y.; Hu, R.; Xia, S.; He, J. Dietary resveratrol supplementation inhibits heat stress-induced high-activated innate immunity and inflammatory response in spleen of yellow-feather broilers. Poult. Sci. 2019, 98, 6378–6387. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Zhuang, Y.; Shi, Y.; Xu, Z.; Zhou, C.; Guo, L.; Liu, P.; Wu, C.; Hu, R.; Hu, G.; et al. Effects of N-acetyl-l-cysteine on heat stress-induced oxidative stress and inflammation in the hypothalamus of hens. J. Therm. Biol. 2021, 98, 102927. [Google Scholar] [CrossRef] [PubMed]
- Arab Ameri, S.; Samadi, F.; Dastar, B.; Zerehdaran, S. Effect of Peppermint (Mentha piperita) Powder on Immune Response of Broiler Chickens in Heat Stress. Iran. J. Appl. Anim. Sci. 2016, 6, 435–445. [Google Scholar]
- Ju, X.-H.; Xu, H.-J.; Yong, Y.-H.; An, L.-L.; Xu, Y.-M.; Jiao, P.-R.; Liao, M. Heat stress upregulates the expression of TLR4 and its alternative splicing variant in bama miniature pigs. J. Integr. Agric. 2014, 13, 2479–2487. [Google Scholar] [CrossRef]
- Varasteh, S.; Braber, S.; Akbari, P.; Garssen, J.; Fink-Gremmels, J. Differences in Susceptibility to Heat Stress along the Chicken Intestine and the Protective Effects of Galacto-Oligosaccharides. PLoS ONE 2015, 10, e0138975. [Google Scholar] [CrossRef]
- Zhou, J.; An, H.; Xu, H.; Liu, S.; Cao, X. Heat shock up-regulates expression of Toll-like receptor-2 and Toll-like receptor-4 in human monocytes via p38 kinase signal pathway. Immunology 2005, 114, 522–530. [Google Scholar] [CrossRef]
- Basu, M.; Paichha, M.; Swain, B.; Lenka, S.S.; Singh, S.; Chakrabarti, R.; Samanta, M. Modulation of TLR2, TLR4, TLR5, NOD1 and NOD2 receptor gene expressions and their downstream signaling molecules following thermal stress in the Indian major carp catla (Catla catla). 3 Biotech. 2015, 5, 1021–1030. [Google Scholar] [CrossRef] [PubMed]
- Goudarzi, M.A.; Radfar, M.; Goudarzi, Z. Peppermint as a promising treatment agent in inflammatory conditions: A comprehensive systematic review of literature. Phytother. Res. 2024, 38, 187–195. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Choi, M.K.; Ban, T.; Yanai, H.; Negishi, H.; Lu, Y.; Tamura, T.; Takaoka, A.; Nishikura, K.; Taniguchi, T. Regulation of innate immune responses by DAI (DLM-1/ZBP1) and other DNA-sensing molecules. Proc. Natl. Acad. Sci. USA 2008, 105, 5477–5482. [Google Scholar] [CrossRef] [PubMed]
- Cao, C.; Chowdhury, V.S.; Cline, M.A.; Gilbert, E.R. The Microbiota-Gut-Brain Axis during Heat Stress in Chickens: A Review. Front. Physiol. 2021, 12, 752265. [Google Scholar] [CrossRef]
- Fang, H.; Feng, X.; Xu, T.; Zhong, R.; Lu, D.; Zhang, H.; Shen, W.; Zhao, Y.; Chen, L. Wang. Gut-Spleen Axis: Microbiota via Vascular and Immune Pathways Improve Busulfan-Induced Spleen Disruption. mSphere 2023, 8, e0058122. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | Temperature (°C) | Diet |
|---|---|---|
| Control group | 21 °C | Basal diet |
| Heat stress group | 31 °C | Basal diet |
| Heat stress + PE group | 31 °C | Basal diet + peppermint extract |
| Ingredients (g/kg) | Content (%) | Ingredients (g/kg) | Content (%) |
|---|---|---|---|
| Corn | 56.51 | Calculated nutrient levels | |
| Soybean meal | 35.52 | Metabolizable energy (MJ/kg) | 12.73 |
| Soybean oil | 4.50 | Crude protein (g/kg) | 20.07 |
| NaCl | 0.30 | Available phosphorus (g/kg) | 0.40 |
| Limestone | 1.00 | Calcium (g/kg) | 0.90 |
| Dicalcium phosphate | 1.78 | Lysine (g/kg) | 1.00 |
| DL-Methionine | 0.11 | Methionine (g/kg) | 0.42 |
| Premix 1 | 0.28 | Methionine + cysteine (g/kg) | 0.78 |
| Total | 100.00 |
| Gene Name | Accession Number | Primer Sequences (5′-3′) | Base Number | Product Length (bp) |
|---|---|---|---|---|
| β-actin | NM_205518.1 | Forward: TCCACCGCAAATGCTTCTAA Reverse: GGGGCGTTCGCTCCA | 20 15 | 205 |
| NOD1 | NM_001318438.1 | Forward: AGGAGGTCTCATCAGCGAACATCT Reverse: GCAGCCTCAGCAGAAGAGCATT | 24 22 | 217 |
| TLR2 | NM_204278.1 | Forward: CCTGGTGTTCCTGTTCATCCTCAT Reverse: AGTTGGAGTCGTTCTCACTGTAGG | 24 24 | 173 |
| TLR4 | NM_001030693.1 | Forward: ACGGAAGGCTTTGGTTGGGATT Reverse: GATGTTGCTATCTGGTGCTTGGAA | 22 24 | 184 |
| MDA5 | NM_001193638.1 | Forward: GTGGCTTCAAGTGGCTCAGGAG Reverse: TCTTCTGGCGGCATCTCTTGGA | 22 22 | 107 |
| DAI | NM_205071.1 | Forward: CAAGCGGTTGGTGCCATCATTG Reverse: ATCCTGCCTTGTGCCTTGAACTG | 22 23 | 167 |
| RIP2 | NM_001030943.1 | Forward: AGCCGCACCTGAGGAACAAGA Reverse: CCGTTGCTGGACTGGATGATGAG | 22 24 | 201 |
| Jun | NM_001031289.1 | Forward: AGCCGCACCTGAGGAACAAGA Reverse: CCGTTGCTGGACTGGATGATGAG | 21 23 | 109 |
| Fos | NM_205508.1 | Forward: GACTTAGCAACGACCCATCTTACG Reverse: CAGAACATTCAGACCACCTCAACA | 24 24 | 155 |
| p50 | NM_001347945.1 | Forward: GCCGACACGCAGAGCAAGAT Reverse: ACAACAGCCAGGTTCTCCTTCATT | 20 24 | 248 |
| p65 | NM_205129.1 | Forward: TCATCCACCGCCGCCACATT Reverse: GGCTGAGGAAGGCACTGAAGTC | 20 22 | 232 |
| p52 | NM_001159511.1 | Forward: TCTACCGTGAACTGGAACAGAACA Reverse: CTGGACACTAAGACTGCTGCTATG | 24 24 | 251 |
| RelB | D13794.1 | Forward: CGGCACAGCTTCAGCAACCT Reverse: TCACCACGTTCATATCCACCTCCT | 20 24 | 145 |
| IRF3 | NM_205372.1 | Forward: ACCGCCGTATCTTCCGCATC Reverse: GGTCCTCCAGCAGCATGAACAT | 20 22 | 192 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ma, D.; Zhang, M.; Feng, J. Dietary Peppermint Extract Inhibits Chronic Heat Stress-Induced Activation of Innate Immunity and Inflammatory Response in the Spleen of Broiler Chickens. Animals 2024, 14, 1157. https://doi.org/10.3390/ani14081157
Ma D, Zhang M, Feng J. Dietary Peppermint Extract Inhibits Chronic Heat Stress-Induced Activation of Innate Immunity and Inflammatory Response in the Spleen of Broiler Chickens. Animals. 2024; 14(8):1157. https://doi.org/10.3390/ani14081157
Chicago/Turabian StyleMa, Dandan, Minhong Zhang, and Jinghai Feng. 2024. "Dietary Peppermint Extract Inhibits Chronic Heat Stress-Induced Activation of Innate Immunity and Inflammatory Response in the Spleen of Broiler Chickens" Animals 14, no. 8: 1157. https://doi.org/10.3390/ani14081157
APA StyleMa, D., Zhang, M., & Feng, J. (2024). Dietary Peppermint Extract Inhibits Chronic Heat Stress-Induced Activation of Innate Immunity and Inflammatory Response in the Spleen of Broiler Chickens. Animals, 14(8), 1157. https://doi.org/10.3390/ani14081157