
Periparturient Mineral Metabolism: Implications to Health and Productivity

{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Risk Factors for Periparturient Mineral Imbalance
3. Gastrointestinal Absorption and Homeostatic Mechanisms Maintaining Blood Calcium
4. Inflammation and Redistribution of Minerals
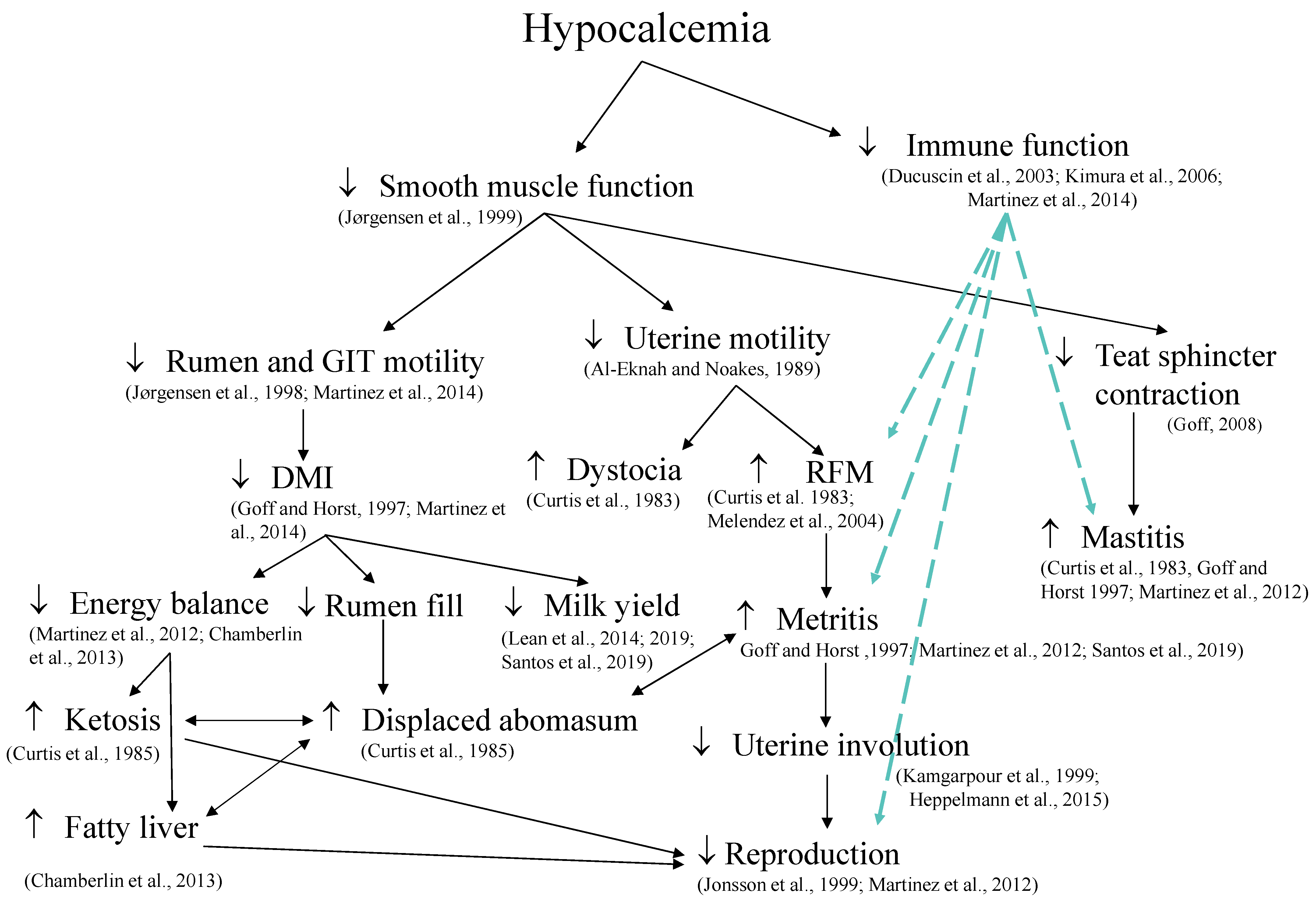
5. Mechanisms Linking Hypocalcemia and Health in Dairy Cows
6. Methods to Prevent Periparturient Mineral Imbalance
6.1. Alterations in Acid-Base Balance
6.2. Low Ca Diets and Sequestering Agents
6.3. Sources of Vitamin D
6.4. Oral Calcium after Calving
7. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Neto, A.V.; Lima, I.; Lopes, F.; Lopera, C.; Zimpel, R.; Sinedino, L.; Jeong, K.; Galvão, K.; Thatcher, W.; Nelson, C.; et al. Use of calcitriol to maintain postpartum blood calcium and improve immune function in dairy cows. J. Dairy Sci. 2017, 100, 5805–5823. [Google Scholar] [CrossRef] [PubMed]
- Lopera, C.; Zimpel, R.; Vieira-Neto, A.; Lopes, F.R.; Ortiz, W.; Poindexter, M.; Faria, B.N.; Gambarini, M.L.; Block, E.; Nelson, C.D.; et al. Effects of level of dietary cation-anion difference and duration of prepartum feeding on performance and metabolism of dairy cows. J. Dairy Sci. 2018, 101, 7907–7929. [Google Scholar] [CrossRef] [PubMed]
- Rodney, R.M.; Martinez, N.; Block, E.; Hernandez, L.L.; Celi, P.; Nelson, C.D.; Santos, J.E.P.; Lean, I.J. Effects of prepartum dietary cation-anion difference and source of vitamin D in dairy cows: Vitamin D, mineral and bone metabolism. J. Dairy Sci. 2018, 101, 2519–2543. [Google Scholar] [CrossRef] [PubMed]
- House, W.A.; Bell, A.W. Mineral accretion in the fetus and adnexa during late gestation in Holstein cows. J. Dairy Sci. 1993, 76, 2999–3010. [Google Scholar] [CrossRef] [PubMed]
- NRC. Nutrient Requirements of Dairy Cattle, 7th ed.; National Academies Press: Washington, DC, USA, 2001. [Google Scholar] [CrossRef]
- Zimpel, R.; Poindexter, M.B.; Neto, A.V.; Block, E.; Nelson, C.D.; Staples, C.R.; Thatcher, W.W.; Santos, J.E.P. Effect of dietary cation-anion difference on acid-base status and dry matter intake in dry pregnant cows. J. Dairy Sci. 2018, 101, 8461–8475. [Google Scholar] [CrossRef] [PubMed]
- Visek, W.; Monroe, R.; Swanson, E.; Comar, C. Calcium metabolism in dairy cows as studied with Ca45. J. Diary Sci. 1953, 36, 373–384. [Google Scholar] [CrossRef]
- Megahed, A.A.; Hiew, M.W.; El Badawy, S.A.; Constable, P.D. Plasma calcium concentrations are decreased at least 9 hours before parturition in multiparous Holstein-Friesian cattle in a herd fed an acidogenic diet during late gestation. J. Dairy Sci. 2018, 101, 1365–1378. [Google Scholar] [CrossRef] [PubMed]
- Goff, J.P.; Kimura, K.; Horst, R.L. Effect of mastectomy on milk fever, energy, and vitamins A, E, and β-carotene status at parturition. J. Dairy Sci. 2002, 85, 1427–1436. [Google Scholar] [CrossRef] [PubMed]
- Reinhardt, T.A.; Lippolis, J.D.; McCluskey, B.J.; Goff, J.P.; Horst, R.L. Prevalence of subclinical hypocalcemia in dairy herds. Vet. J. 2011, 188, 122–124. [Google Scholar] [CrossRef]
- McArt, J.; Neves, R. Association of transient, persistent, or delayed subclinical hypocalcemia with early lactation disease, removal, and milk yield in Holstein cows. J. Dairy Sci. 2020, 103, 690–701. [Google Scholar] [CrossRef]
- Martinez, N.; Sinedino, L.D.; Bisinotto, R.S.; Daetz, R.; Lopera, C.; Risco, C.A.; Galvão, K.N.; Thatcher, W.W.; Santos, J.E.P. Effects of oral calcium supplementation on mineral and acid-base status, energy metabolites, and health of postpartum dairy cows. J. Dairy Sci. 2016, 99, 8397–8416. [Google Scholar] [CrossRef] [PubMed]
- Kimura, K.; Reinhardt, T.; Goff, J. Parturition and hypocalcemia blunts calcium signals in immune cells of dairy cattle. J. Dairy Sci. 2006, 89, 2588–2595. [Google Scholar] [CrossRef]
- Martinez, N.; Sinedino, L.; Bisinotto, R.; Ribeiro, E.; Gomes, G.; Lima, F.; Greco, L.; Risco, C.; Galvão, K.; Taylor-Rodriguez, D.; et al. Effect of induced subclinical hypocalcemia on physiological responses and neutrophil function in dairy cows. J. Dairy Sci. 2014, 97, 874–887. [Google Scholar] [CrossRef] [PubMed]
- Goff, J.P. Macromineral physiology and application to the feeding of the dairy cow for prevention of milk fever and other periparturient mineral disorders. Anim. Feed Sci. Technol. 2006, 126, 237–257. [Google Scholar] [CrossRef]
- Martinez, N.; Risco, C.; Lima, F.; Bisinotto, R.; Greco, L.; Ribeiro, E.; Maunsell, F.; Galvão, K.; Santos, J.E.P. Evaluation of peripartal calcium status, energetic profile, and neutrophil function in dairy cows at low or high risk of developing uterine disease. J. Dairy Sci. 2012, 95, 7158–7172. [Google Scholar] [CrossRef] [PubMed]
- DeGaris, P.J.; Lean, I.J. Milk fever in dairy cows: A review of pathophysiology and control principles. Vet. J. 2008, 176, 58–69. [Google Scholar] [CrossRef]
- Vagg, M.; Payne, J. The effect of ammonium chloride induced acidosis on calcium metabolism in ruminants. Br. Vet. J. 1970, 126, 531–537. [Google Scholar] [CrossRef] [PubMed]
- Horst, R.L.; Goff, J.P.; Reinhardt, T.A. Advancing age results in reduction of intestinal and bone 1,25-dihydroxyvitamin D receptor. Endocrinology 1990, 126, 1053–1057. [Google Scholar] [CrossRef] [PubMed]
- Liesegang, A.; Singer, K.; Boos, A. Vitamin D receptor amounts across different segments of the gastrointestinal tract in Brown Swiss and Holstein Frisian cows of different age. J. Anim. Physiol. Anim. Nutr. 2008, 92, 316–323. [Google Scholar] [CrossRef] [PubMed]
- Hanai, H.; Brennan, D.P.; Cheng, L.; Goldman, M.E.; Chorev, M.; Levine, M.A.; Sacktor, B.; Liang, C.T. Downregulation of parathyroid hormone receptors in renal membranes from aged rats. Am. J. Physiol. Physiol. 1990, 259, F444–F450. [Google Scholar] [CrossRef] [PubMed]
- Armbrecht, H.J.; Boltz, M.; Strong, R.; Richardson, A.; Bruns, M.E.H.; Christakos, S. Expression of calbindin-D decreases with age in intestine and kidney. Endocrinology 1989, 125, 2950–2956. [Google Scholar] [CrossRef]
- Suttle, N.F.; Field, A.C. Studies on magnesium in ruminant nutrition. 8. Effect of increased intakes of potassium and water on the metabolism of magnesium, phosphorus, sodium, potassium and calcium in sheep. Br. J. Nutr. 1967, 21, 819–831. [Google Scholar] [CrossRef] [PubMed]
- Leonhard-Marek, S.; Martens, H. Effects of potassium on magnesium transport across rumen epithelium. Am. J. Physiol. 1996, 271, G1034–G1038. [Google Scholar] [CrossRef] [PubMed]
- Estep, H.; Shaw, W.A.; Watlington, C.; Hobe, R.; Holland, W.; Tucker, S.G. Hypocalcemia due to hypomagnesemia and reversible parathyroid hormone unresponsiveness. J. Clin. Endocrinol. Metab. 1969, 29, 842–848. [Google Scholar] [CrossRef] [PubMed]
- Nissenson, R.A.; Strewler, G.J.; Williams, R.D.; Leung, S.C. Activation of the parathyroid hormone receptor-adenylate cyclase system in osteosarcoma cells by a human renal carcinoma factor. Cancer Res. 1985, 45, 5358–5363. [Google Scholar] [PubMed]
- Dunlay, R.; Hruska, K. PTH receptor coupling to phospholipase C is an alternate pathway of signal transduction in bone and kidney. Am. J. Physiol. Physiol. 1990, 258, F223–F231. [Google Scholar] [CrossRef]
- Maguire, M.E. Hormone-sensitive magnesium transport and magnesium regulation of adenylate cyclase. Trends Pharmacol. Sci. 1984, 5, 73–77. [Google Scholar] [CrossRef]
- Connolly, T.M.; Bross, T.E.; Majerus, P.W. Isolation of a phosphomonoesterase from human platelets that specifically hydrolyzes the 5-phosphate of inositol 1,4,5-triphosphate. J. Biol. Chem. 1985, 13, 7868–7874. [Google Scholar] [CrossRef]
- Anast, C.S.; Mohs, J.M.; Kaplan, S.L.; Burns, T.W. Evidence for parathyroid failure in magnesium deficiency. Science 1972, 177, 606–608. [Google Scholar] [CrossRef] [PubMed]
- Goff, J.P. The monitoring, prevention, and treatment of milk fever and subclinical hypocalcemia in dairy cows. Vet. J. 2008, 176, 50–57. [Google Scholar] [CrossRef] [PubMed]
- Baker, R.M.; Boston, R.C.; Boyes, T.E.; Leaver, D.D. Variations in the response of sheep to experimental magnesium deficiency. Res. Vet. Sci. 1979, 26, 129–133. [Google Scholar] [CrossRef] [PubMed]
- Weiss, W.P. Macromineral digestion by lactating dairy cows: Factors affecting digestibility of magnesium. J. Dairy Sci. 2004, 87, 2167–2171. [Google Scholar] [CrossRef] [PubMed]
- Martens, H.; Schweigel, M. Pathophysiology of grass tetany and other hypomagnesemias. Vet. Clin. N. Am. Food Anim. Pract. 2000, 16, 339–368. [Google Scholar] [CrossRef] [PubMed]
- Ram, L.; Schonewille, J.T.; Martens, H.; Klooster, A.T.V.; Beynen, A.C. Magnesium absorption by wethers fed potassium bicarbonate in combination with different dietary magnesium concentrations. J. Dairy Sci. 1998, 81, 2485–2492. [Google Scholar] [CrossRef] [PubMed]
- Lean, I.J.; DeGaris, P.J.; McNeil, D.M.; Block, E. Hypocalcemia in dairy cows: Meta-analysis and dietary cation anion difference theory revisited. J. Dairy Sci. 2006, 89, 669–684. [Google Scholar] [CrossRef] [PubMed]
- Van De Braak, A.E.; Klooster, A.T.V.; Malestein, A. Influence of a deficient supply of magnesium during the dry period on the rate of calcium mobilization by dairy cows at parturition. Res. Vet. Sci. 1987, 42, 101–108. [Google Scholar] [CrossRef] [PubMed]
- Knowlton, K.; Herbein, J. Phosphorus partitioning during early lactation in dairy cows fed diets varying in phosphorus content. J. Dairy Sci. 2002, 85, 1227–1236. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.; Zhou, J.; Tang, W.; Jiang, X.; Rowe, D.W.; Quarles, L.D. Pathogenic role of Fgf23 in Hyp mice. Am. J. Physiol. Metab. 2006, 291, E38–E49. [Google Scholar] [CrossRef] [PubMed]
- Gattineni, J.; Bates, C.M.; Twombley, K.; Dwarakanath, V.; Robinson, M.L.; Goetz, R.; Mohammadi, M.; Baum, M. FGF23 decreases renal NaPi-2a and NaPi-2c expression and induces hypophosphatemia in vivo predominantly via FGF receptor 1. Am. J. Physiol. Physiol. 2009, 297, F282–F291. [Google Scholar] [CrossRef] [PubMed]
- Shimada, T.; Kakitani, M.; Yamazaki, Y.; Hasegawa, H.; Takeuchi, Y.; Fujita, T.; Fukumoto, S.; Tomizuka, K.; Yamashita, T. Targeted ablation of Fgf23 demonstrates an essential physiological role of FGF23 in phosphate and vitamin D metabolism. J. Clin. Investig. 2004, 113, 561–568. [Google Scholar] [CrossRef] [PubMed]
- Cohrs, I.; Wilkens, M.; Grünberg, W. Short communication: Effect of dietary phosphorus deprivation in late gestation and early lactation on the calcium homeostasis of periparturient dairy cows. J. Dairy Sci. 2018, 101, 9591–9598. [Google Scholar] [CrossRef] [PubMed]
- Peterson, A.; Orth, M.; Goff, J.; Beede, D. Periparturient responses of multiparous Holstein cows fed different dietary phosphorus concentrations prepartum. J. Dairy Sci. 2005, 88, 3582–3594. [Google Scholar] [CrossRef] [PubMed]
- Santos, J.E.P.; Lean, I.J.; Golder, H.; Block, E. Meta-analysis of the effects of prepartum dietary cation-anion difference on performance and health of dairy cows. J. Dairy Sci. 2019, 102, 2134–2154. [Google Scholar] [CrossRef] [PubMed]
- Bacchetta, J.; Sea, J.L.; Chun, R.F.; Lisse, T.S.; Wesseling-Perry, K.; Gales, B.; Adams, J.S.; Salusky, I.B.; Hewison, M. FGF23 inhibits extra-renal synthesis of 1,25-dihydroxyvitamin D in human monocytes. J. Bone Miner. Res. 2012, 28, 46–55. [Google Scholar] [CrossRef]
- Nelson, C.; Reinhardt, T.; Thacker, T.; Beitz, D.; Lippolis, J. Modulation of the bovine innate immune response by production of 1α,25-dihydroxyvitamin D3 in bovine monocytes. J. Dairy Sci. 2010, 93, 1041–1049. [Google Scholar] [CrossRef] [PubMed]
- Roche, J.; Berry, D. Periparturient climatic, animal, and management factors influencing the incidence of milk fever in grazing systems. J. Dairy Sci. 2006, 89, 2775–2783. [Google Scholar] [CrossRef] [PubMed]
- Heuer, C.; Schukken, Y.; Dobbelaar, P. Postpartum body condition score and results from the first test day milk as predictors of disease, fertility, yield, and culling in commercial dairy herds. J. Dairy Sci. 1999, 82, 295–304. [Google Scholar] [CrossRef] [PubMed]
- Leno, B.M.; Neves, R.C.; Louge, I.M.; Curler, M.D.; Thomas, M.J.; Overton, T.R.; McArt, J.A. Differential effects of a single dose of oral calcium based on postpartum calcium concentration in Holstein cows. J. Dairy Sci. 2018, 101, 3285–3302. [Google Scholar] [CrossRef] [PubMed]
- Vieira-Neto, A.; Negro, G.; Zimpel, R.; Poindexter, M.; Lopes, F., Jr.; Thatcher, W.; Nelson, C.; Santos, J.E.P. Effects of injectable calcitriol on mineral metabolism and postpartum health and performance in dairy cows. J. Dairy Sci. 2021, 104, 683–701. [Google Scholar] [CrossRef] [PubMed]
- Sejersen, H.; Sørensen, M.T.; Larsen, T.; Bendixen, E.; Ingvartsen, K.L. Liver protein expression in dairy cows with high liver triglycerides in early lactation. J. Dairy Sci. 2012, 95, 2409–2421. [Google Scholar] [CrossRef] [PubMed]
- McLeod, J.F.; Cooke, N.E. The vitamin D-binding protein, α-fetoprotein, albumin multigene family: Detection of transcripts in multiple tissues. J. Biol. Chem. 1989, 264, 21760–21769. [Google Scholar] [CrossRef] [PubMed]
- Nykjaer, A.; Dragun, D.; Walther, D.; Vorum, H.; Jacobsen, C.; Herz, J.; Melsen, F.; Christensen, E.I.; Willnow, T.E. An endocytic pathway essential for renal uptake and activation of the steroid 25-(OH) vitamin D3. Cell 1999, 96, 507–515. [Google Scholar] [CrossRef] [PubMed]
- Schröder, B.; Breves, G. Mechanisms and regulation of calcium absorption from the gastrointestinal tract in pigs and ruminants: Comparative aspects with special emphasis on hypocalcemia in dairy cows. Anim. Health Res. Rev. 2006, 7, 31–41. [Google Scholar] [CrossRef] [PubMed]
- Khorasani, G.; Armstrong, D. Calcium, phosphorus, and magnesium absorption and secretion in the bovine digestive tract as influenced by dietary concentration of these elements. Livest. Prod. Sci. 1992, 31, 271–286. [Google Scholar] [CrossRef]
- Frömter, E.; Diamond, J. Route of passive ion permeation in epithelia. Nat. New Biol. 1972, 235, 9–13. [Google Scholar] [CrossRef] [PubMed]
- Farquhar, M.G.; Palade, G.E. Junctional complexes in various epithelia. J. Cell Biol. 1963, 17, 375–412. [Google Scholar] [CrossRef] [PubMed]
- Günzel, D.; Yu, A.S.L. Claudins and the modulation of tight junction permeability. Physiol. Rev. 2013, 93, 525–569. [Google Scholar] [CrossRef] [PubMed]
- Fujita, H.; Sugimoto, K.; Inatomi, S.; Maeda, T.; Osanai, M.; Uchiyama, Y.; Yamamoto, Y.; Wada, T.; Kojima, T.; Yokozaki, H.; et al. Tight junction proteins claudin-2 and -12 are critical for vitamin D-dependent Ca2+ absorption between enterocytes. Mol. Biol. Cell 2008, 19, 1912–1921. [Google Scholar] [CrossRef]
- Elfers, K.; Marr, I.; Wilkens, M.R.; Breves, G.; Langeheine, M.; Brehm, R.; Muscher-Banse, A.S. Expression of tight junction properties and cadherin 17 in the small intestine of young goats offered a reduced N and/or Ca diet. PLoS ONE 2016, 11, e0154311. [Google Scholar] [CrossRef] [PubMed]
- Tudpor, K.; Teerapornpuntakit, J.; Jantarajit, W.; Krishnamra, N.; Charoenphandhu, N. 1,25-dihydroxyvitamin D3 rapidly stimulates the solvent drag-induced paracellular calcium transport in the duodenum of female rats. J. Physiol. Sci. 2008, 58, 297–307. [Google Scholar] [CrossRef]
- Kutuzova, G.D.; DeLuca, H.F. Gene expression profiles in rat intestine identify pathways for 1,25-dihydroxyvitamin D3 stimulated calcium absorption and clarify its immunomodulatory properties. Arch. Biochem. Biophys. 2004, 432, 152–166. [Google Scholar] [CrossRef] [PubMed]
- Schröder, B.; Wilkens, M.R.; Ricken, G.E.; Leonhard-Marek, S.; Fraser, D.R.; Breves, G. Calcium transport in bovine rumen epithelium as affected by luminal Ca concentrations and Ca sources. Physiol. Rep. 2015, 3, e12615. [Google Scholar] [CrossRef] [PubMed]
- Hyde, M.L.; Wilkens, M.R.; Fraser, D.R. In vivo measurement of strontium absorption from the rumen of dairy cows as an index of calcium absorption capacity. J. Dairy Sci. 2019, 102, 5699–5705. [Google Scholar] [CrossRef] [PubMed]
- Höller, H.; Breves, G.; Gerdes, H.; Kocabatmaz, M. Flux of calcium across the sheep rumen wall in vivo and in vitro. Q. J. Exp. Physiol. 1988, 73, 609–618. [Google Scholar] [CrossRef] [PubMed]
- Peng, J.-B.; Chen, X.-Z.; Berger, U.V.; Vassilev, P.M.; Tsukaguchi, H.; Brown, E.M.; Hediger, M.A. Molecular cloning and characterization of a channel-like transporter mediating intestinal calcium absorption. J. Biol. Chem. 1999, 274, 22739–22746. [Google Scholar] [CrossRef] [PubMed]
- Hoenderop, J.G.J.; van der Kemp, A.W.C.M.; Hartog, A.; van de Graaf, S.F.; van Os, C.H.; Willems, P.H.; Bindels, R.J.M. Molecular identification of the apical Ca2+ channel in 1,25-dihydroxyvitamin D3-responsive epithelia. J. Biol. Chem. 1999, 274, 8375–8378. [Google Scholar] [CrossRef] [PubMed]
- Perret, C.; Desplan, C.; Thomasset, M. Cholecalcin (a 9-kDa cholecalciferol-induced calcium-binding protein) messenger RNA: Distribution and induction by calcitriol in the rat digestive tract. Eur. J. Biochem. 1985, 150, 211–217. [Google Scholar] [CrossRef] [PubMed]
- Taylor, A.N.; McIntosh, J.E.; Bourdeau, J.E. Immunocytochemical localization of vitamin D-dependent calcium-binding protein in renal tubules of rabbit, rat, and chick. Kidney Int. 1982, 21, 765–773. [Google Scholar] [CrossRef] [PubMed]
- Bredderman, P.J.; Wasserman, R.H. Chemical composition, affinity for calcium, and some related properties of the vitamin D dependent calcium-binding protein. Biochemistry 1974, 13, 1687–1694. [Google Scholar] [CrossRef] [PubMed]
- Schachter, D.; Rosen, S.M. Active transport of Ca45 by the small intestine and its dependence on vitamin D. Am. J. Physiol. Content 1959, 196, 357–362. [Google Scholar] [CrossRef] [PubMed]
- Favus, M.J.; Angeid-Backman, E.; Breyer, M.D.; Coe, F.L. Effects of trifluoperazine, ouabain, and ethacrynic acid on intestinal calcium transport. Am. J. Physiol. Liver Physiol. 1983, 244, G111–G115. [Google Scholar] [CrossRef] [PubMed]
- Ghijsen, W.E.; De Jong, M.D.; Van Os, C.H. Kinetic properties of Na+/Ca2+ exchange in basolateral plasma membranes of rat small intestine. Biochim. Biophys. Acta 1983, 730, 85–94. [Google Scholar] [CrossRef] [PubMed]
- Clausen, M.V.; Hilbers, F.; Poulsen, H. The structure and function of the Na,K-ATPase isoforms in health and disease. Front. Physiol. 2017, 8, 371. [Google Scholar] [CrossRef] [PubMed]
- Brown, E.M.; Gamba, G.; Riccardi, D.; Lombardi, M.; Butters, R.; Kifor, O.; Sun, A.; Hediger, M.A.; Lytton, J.; Hebert, S.C. Cloning and characterization of an extracellular Ca2+-sensing receptor from bovine parathyroid. Nature 1993, 366, 575–580. [Google Scholar] [CrossRef] [PubMed]
- Morrissey, J.; Cohn, D. Secretion and degradation of parathormone as a function of intracellular maturation of hormone pools. J. Cell Biol. 1979, 83, 521–528. [Google Scholar] [CrossRef] [PubMed]
- van Abel, M.; Hoenderop, J.G.; van der Kemp, A.W.; Friedlaender, M.M.; van Leeuwen, J.P.; Bindels, R.J. Coordinated control of renal Ca2+ transport proteins by parathyroid hormone. Kidney Int. 2005, 68, 1708–1721. [Google Scholar] [CrossRef] [PubMed]
- Matsumoto, N.; Hemmi, A.; Yamato, H.; Ohnishi, R.; Segawa, H.; Ohno, S.; Miyamoto, K. Immunohistochemical analyses of parathyroid hormone-dependent downregulation of renal type II Na-Pi co-transporters by cryobiopsy. J. Med. Investig. 2010, 57, 138–145. [Google Scholar] [CrossRef] [PubMed]
- O’Brien, C.A.; Nakashima, T.; Takayanagi, H. Osteocyte control of osteoclastogenesis. Bone 2013, 54, 258–263. [Google Scholar] [CrossRef] [PubMed]
- Fu, Q.; Jilka, R.L.; Manolagas, S.C.; O’Brien, C.A. Parathyroid hormone stimulates receptor activator of NFkB ligand and inhibits osteoprotegerin expression via protein kinase A activation of cAMP-response element-binding protein. J. Biol. Chem. 2002, 277, 48868–48875. [Google Scholar] [CrossRef] [PubMed]
- Picotto, G.; Massheimer, V.; Boland, R. Parathyroid hormone stimulates calcium influx and the cAMP messenger system in rat enterocytes. Am. J. Physiol. Physiol. 1997, 273, C1349–C1353. [Google Scholar] [CrossRef] [PubMed]
- Nemere, I.; Norman, A.W. Parathyroid hormone stimulates calcium transport in perfused duodena from normal chicks: Comparison with the rapid (transcaltachic) effect of 1,25-dihydroxyvitamin D3. Endocrinology 1986, 119, 1406–1408. [Google Scholar] [CrossRef] [PubMed]
- Hymøller, L.; Jensen, S. Vitamin D(3) synthesis in the entire skin surface of dairy cows despite hair coverage. J. Dairy Sci. 2010, 93, 2025–2029. [Google Scholar] [CrossRef] [PubMed]
- Hanewald, K.H.; Rappoldt, M.P.; Roborgh, J.R. The antirachitic activity of previtamin D3. Recl. Trav. Chim. Pays-Bas Belg. 1961, 80, 1003–1014. [Google Scholar] [CrossRef]
- Ponchon, G.; Kennan, A.L.; DeLuca, H.F. Activation of vitamin D by the liver. J. Clin. Investig. 1969, 48, 2032–2037. [Google Scholar] [CrossRef] [PubMed]
- Fraser, D.R.; Kodicek, E. Unique biosynthesis by kidney of a biologically active vitamin D metabolite. Nature 1970, 228, 764–766. [Google Scholar] [CrossRef] [PubMed]
- Trang, H.M.; Cole, D.E.; Rubin, L.A.; Andreas, P.; Shirley, S.; Reinhold, V. Evidence that vitamin D3 increases serum 25-hydroxyvitamin D more efficiently than does vitamin D2. Am. J. Clin. Nutr. 1998, 68, 854–858. [Google Scholar] [CrossRef] [PubMed]
- Hymøller, L.; Jensen, S. Vitamin D2 impairs utilization of vitamin D3 in high-yielding dairy cows in a cross-over supplementation regimen. J. Dairy Sci. 2011, 94, 3462–3466. [Google Scholar] [CrossRef] [PubMed]
- Bhattacharyya, M.H.; DeLuca, H. Subcellular location of rat liver calciferol-25-hydroxylase. Arch. Biochem. Biophys. 1974, 160, 58–62. [Google Scholar] [CrossRef] [PubMed]
- Nelson, C.D.; Lippolis, J.D.; Reinhardt, T.A.; Sacco, R.E.; Powell, J.L.; Drewnoski, M.E.; O’Neil, M.; Beitz, D.C.; Weiss, W.P. Vitamin D status of dairy cattle: Outcomes of current practices in the dairy industry. J. Dairy Sci. 2016, 99, 10150–10160. [Google Scholar] [CrossRef] [PubMed]
- Fraser, D.R.; Kodicek, E. Regulation of 25-hydroxycholecalciferol-1-hydroxylase activity in kidney by parathyroid hormone. Nat. New Biol. 1973, 241, 163–166. [Google Scholar] [CrossRef] [PubMed]
- Poindexter, M.B.; Kweh, M.F.; Zimpel, R.; Zuniga, J.; Lopera, C.; Zenobi, M.G.; Jiang, Y.; Engstrom, M.; Celi, P.; Santos, J.E.P.; et al. Feeding supplemental 25-hydroxyvitamin D3 increases serum mineral concentrations and alters mammary immunity of lactating dairy cows. J. Dairy Sci. 2020, 103, 805–822. [Google Scholar] [CrossRef] [PubMed]
- Dahlbäck, H.; Wikvall, K. 25-Hydroxylation of vitamin D3 in rat liver: Roles of mitochondrial and microsomal cytochrome P-450. Biochem. Biophys. Res. Commun. 1987, 142, 999–1005. [Google Scholar] [CrossRef] [PubMed]
- Casas, E.; Leach, R.J.; Reinhardt, T.A.; Thallman, R.M.; Lippolis, J.D.; Bennett, G.L.; Kuehn, L.A. A genomewide association study identified CYP2J2 as a gene controlling serum vitamin D status in beef cattle. J. Anim. Sci. 2013, 91, 3549–3556. [Google Scholar] [CrossRef] [PubMed]
- Baran, D.T.; Milne, M.L. 1,25-Dihydroxyvitamin D increases hepatocyte cytosolic calcium levels: A potential regulator of vitamin D-25 hydroxylase. J. Clin. Investig. 1986, 77, 1622–1626. [Google Scholar] [CrossRef] [PubMed]
- Corlett, S.; Chaudhary, M.; Tomlinson, S.; Care, A. The involvement of intracellular calcium ion concentration and calmodulin in the 25-hydroxylation of cholecalciferol in ovine and rat liver. Cell Calcium 1987, 8, 247–258. [Google Scholar] [CrossRef] [PubMed]
- Haussler, M.R.; Whitfield, G.K.; Kaneko, I.; Haussler, C.A.; Hsieh, D.; Hsieh, J.-C.; Jurutka, P.W. Molecular mechanisms of vitamin D action. Calcif. Tissue Int. 2013, 92, 77–98. [Google Scholar] [CrossRef] [PubMed]
- Carlberg, C.; Seuter, S. Dynamics of nuclear receptor target gene regulation. Chromosoma 2010, 119, 479–484. [Google Scholar] [CrossRef] [PubMed]
- Pansu, D.; Bellaton, C.; Roche, C.; Bronner, F. Duodenal and ileal calcium absorption in the rat and effects of vitamin D. Am. J. Physiol. Liver Physiol. 1983, 244, G695–G700. [Google Scholar] [CrossRef] [PubMed]
- Van Cromphaut, S.J.; Dewerchin, M.; Hoenderop, J.G.J.; Stockmans, I.; Van Herck, E.; Kato, S.; Bindels, R.J.M.; Collen, D.; Carmeliet, P.; Bouillon, R.; et al. Duodenal calcium absorption in vitamin D receptor-knockout mice: Functional and molecular aspects. Proc. Natl. Acad. Sci. USA 2001, 98, 13324–13329. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.M.; Riley, E.M.; Meyer, M.B.; Benkusky, N.A.; Plum, L.A.; DeLuca, H.F.; Pike, J.W. 1,25-dihydroxyvitamin D3 controls a cohort of vitamin D receptor target genes in the proximal intestine that is enriched for calcium-regulating components. J. Biol. Chem. 2015, 290, 18199–18215. [Google Scholar] [CrossRef] [PubMed]
- Khuituan, P.; Wongdee, K.; Jantarajit, W.; Suntornsaratoon, P.; Krishnamra, N.; Charoenphandhu, N. Fibroblast growth factor-23 negates 1,25(OH)2D3-induced intestinal calcium transport by reducing the transcellular and paracellular calcium fluxes. Arch. Biochem. Biophys. 2013, 536, 46–52. [Google Scholar] [CrossRef]
- Li, Y.C.; Pirro, A.E.; Demay, M.B. Analysis of vitamin D-dependent calcium-binding protein messenger ribonucleic acid expression in mice lacking the vitamin D receptor. Endocrinology 1998, 139, 847–851. [Google Scholar] [CrossRef] [PubMed]
- Yamagishi, N.; Miyazaki, M.; Naito, Y. The expression of genes for transepithelial calcium-transporting proteins in the bovine duodenum. Vet. J. 2006, 171, 363–366. [Google Scholar] [CrossRef] [PubMed]
- Li, F.; Li, C.; Chen, Y.; Liu, J.; Zhang, C.; Irving, B.; Fitzsimmons, C.; Plastow, G.; Guan, L.L. Host genetics influence the rumen microbiota and heritable rumen microbial features associate with feed efficiency in cattle. Microbiome 2019, 7, 92. [Google Scholar] [CrossRef] [PubMed]
- Malmuthuge, N.; Liang, G.; Guan, L.L. Regulation of rumen development in neonatal ruminants through microbial metagenomes and host transcriptomes. Genome Biol. 2019, 20, 172. [Google Scholar] [CrossRef] [PubMed]
- Zhao, K.; Chen, Y.H.; Penner, G.B.; Oba, M.; Guan, L.L. Transcriptome analysis of ruminal epithelia revealed potential regulatory mechanisms involved in host adaptation to gradual high fermentable dietary transition in beef cattle. BMC Genom. 2017, 18, 976. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Li, C.; Li, F.; Wang, X.; Zhang, X.; Liu, T.; Nian, F.; Yue, X.; Li, F.; Pan, X.; et al. Effects of early feeding on the host rumen transcriptome and bacterial diversity in lambs. Sci. Rep. 2016, 6, 32479. [Google Scholar] [CrossRef] [PubMed]
- Wilkens, M.R.; Kunert-Keil, C.; Brinkmeier, H.; Schröder, B. Expression of calcium channel TRPV6 in ovine epithelial tissue. Vet. J. 2009, 182, 294–300. [Google Scholar] [CrossRef] [PubMed]
- Schrapers, K.T.; Sponder, G.; Liebe, F.; Liebe, H.; Stumpff, F. The bovine TRPV3 as a pathway for the uptake of Na+, Ca2+, and NH4+. PLoS ONE 2018, 13, e0193519. [Google Scholar] [CrossRef] [PubMed]
- Kitazawa, S.; Kajimoto, K.; Kondo, T.; Kitazawa, R. Vitamin D3 supports osteoclastogenesis via functional vitamin D response element of human RANKL gene promoter. J. Cell. Biochem. 2003, 89, 771–777. [Google Scholar] [CrossRef] [PubMed]
- Kondo, T.; Kitazawa, R.; Maeda, S.; Kitazawa, S. 1α,25 dihydroxyvitamin D3 rapidly regulates the mouse osteoprotegerin gene through dual pathways. J. Bone Miner. Res. 2004, 19, 1411–1419. [Google Scholar] [CrossRef] [PubMed]
- Shapiro, L.H.; Venta, P.J.; Yu, Y.L.; Tashian, R.E. Carbonic anhydrase II is induced in HL-60 cells by 1,25-dihydroxyvitamin D3: A model for osteoclast gene regulation. FEBS Lett. 1989, 249, 307–310. [Google Scholar] [CrossRef] [PubMed]
- Dimke, H.; Hoenderop, J.G.; Bindels, R.J. Hereditary tubular transport disorders: Implications for renal handling of Ca2+ and Mg2+. Clin. Sci. 2009, 118, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Seldin, D.W. Renal handling of calcium. Nephron 1999, 81, 2–7. [Google Scholar] [CrossRef] [PubMed]
- Yeh, B.-I.; Sun, T.-J.; Lee, J.Z.; Chen, H.-H.; Huang, C.-L. Mechanism and molecular determinant for regulation of rabbit transient receptor potential type 5 (TRPV5) channel by extracellular pH. J. Biol. Chem. 2003, 278, 51044–51052. [Google Scholar] [CrossRef] [PubMed]
- Hoenderop, J.G.J.; Müller, D.; van der Kemp, A.W.C.M.; Hartog, A.; Suzuki, M.; Ishibashi, K.; Imai, M.; Sweep, F.; Willems, P.H.G.M.; van Os, C.H.; et al. Calcitriol controls the epithelial calcium channel in kidney. J. Am. Soc. Nephrol. 2001, 12, 1342–1349. [Google Scholar] [CrossRef] [PubMed]
- Friedman, P.A.; Gesek, F.A. Vitamin D3 accelerates PTH-dependent calcium transport in distal convoluted tubule cells. Am. J. Physiol. Physiol. 1993, 265, F300–F308. [Google Scholar] [CrossRef] [PubMed]
- Sheldon, I.M.; Cronin, J.; Goetze, L.; Donofrio, G.; Schuberth, H.-J. Defining postpartum uterine disease and the mechanisms of infection and immunity in the female reproductive tract in cattle. Biol. Reprod. 2009, 81, 1025–1032. [Google Scholar] [CrossRef] [PubMed]
- McDougall, S.; Arthur, D.; Bryan, M.; Vermunt, J.; Weir, A. Clinical and bacteriological response to treatment of clinical mastitis with one of three intramammary antibiotics. N. Z. Vet. J. 2007, 55, 161–170. [Google Scholar] [CrossRef] [PubMed]
- Raetz, C.R.H.; Whitfield, C. Lipopolysaccharide endotoxins. Annu. Rev. Biochem. 2002, 71, 635–700. [Google Scholar] [CrossRef] [PubMed]
- Chow, J.C.; Young, D.W.; Golenbock, D.T.; Christ, W.J.; Gusovsky, F. Toll-like receptor-4 mediates lipopolysaccharide-induced signal transduction. J. Biol. Chem. 1999, 274, 10689–10692. [Google Scholar] [CrossRef] [PubMed]
- Andreakos, E.; Sacre, S.M.; Smith, C.; Lundberg, A.; Kiriakidis, S.; Stonehouse, T.; Monaco, C.; Feldmann, M.; Foxwell, B.M. Distinct pathway of LPS-induced NF-kB activation and cytokine production in human myeloid and nonmyeloid cells defined by selective utilization of MyD88 and Mal/TIRAP. Immunobiology 2004, 103, 2229–2237. [Google Scholar] [CrossRef]
- Waldron, M.; Nonnecke, B.; Nishida, T.; Horst, R.; Overton, T. Effect of lipopolysaccharide infusion on serum micromineral and vitamin D concentrations in dairy cows. J. Dairy Sci. 2003, 86, 3440–3446. [Google Scholar] [CrossRef] [PubMed]
- Kvidera, S.; Horst, E.; Abuajamieh, M.; Mayorga, E.; Fernandez, M.S.; Baumgard, L. Glucose requirements of an activated immune system in lactating Holstein cows. J. Dairy Sci. 2017, 100, 2360–2374. [Google Scholar] [CrossRef]
- Holowaychuk, M.; Birkenheuer, A.; Li, J.; Marr, H.; Boll, A.; Nordone, S. Hypocalcemia and hypovitaminosis D in dogs with induced endotoxemia. J. Vet. Intern. Med. 2012, 26, 244–251. [Google Scholar] [CrossRef] [PubMed]
- Shinozuka, Y.; Kawai, K.; Sato, R.; Higashitani, A.; Hamamoto, Y.; Okita, M.; Isobe, N. Blood ionized calcium levels and acute-phase blood glucose kinetics in goats after intramammary infusion of lipopolysaccharide. J. Vet. Med. Sci. 2018, 80, 242–246. [Google Scholar] [CrossRef] [PubMed]
- Toribio, R.E.; Kohn, C.W.; Hardy, J.; Rosol, T.J. Alterations in serum parathyroid hormone and electrolyte concentrations and urinary excretion of electrolytes in horses with induced endotoxemia. J. Vet. Intern. Med. 2005, 19, 223–231. [Google Scholar] [CrossRef] [PubMed]
- Hisaeda, K.; Koshiishi, T.; Sasaki, A.; Shinozuka, Y.; Isobe, N.; Kawai, K. Changes in ionized calcium concentration in the blood of dairy cows with peracute coliform mastitis. J. Vet. Med. Sci. 2020, 82, 457–462. [Google Scholar] [CrossRef] [PubMed]
- Gennaro, R.; Pozzan, T.; Romeo, D. Monitoring of cytosolic free Ca2+ in C5a-stimulated neutrophils: Loss of receptor-modulated Ca2+ stores and Ca2+ uptake in granule-free cytoplasts. Proc. Natl. Acad. Sci. USA 1984, 81, 1416–1420. [Google Scholar] [CrossRef] [PubMed]
- Lioudyno, M.I.; Kozak, J.A.; Penna, A.; Safrina, O.; Zhang, S.L.; Sen, D.; Roos, J.; Stauderman, K.A.; Cahalan, M.D. Orai1 and STIM1 move to the immunological synapse and are up-regulated during T cell activation. Proc. Natl. Acad. Sci. USA 2008, 105, 2011–2016. [Google Scholar] [CrossRef] [PubMed]
- Kobayashi, S.D.; Malachowa, N.; DeLeo, F.R. Neutrophils and bacterial immune evasion. J. Innate Immun. 2018, 10, 432–441. [Google Scholar] [CrossRef] [PubMed]
- Hermant, B.; Bibert, S.; Concord, E.; Dublet, B.; Weidenhaupt, M.; Vernet, T.; Gulino-Debrac, D. Identification of proteases involved in the proteolysis of vascular endothelium cadherin during neutrophil transmigration. J. Biol. Chem. 2003, 278, 14002–14012. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Zhao, X.; Ma, S. Secretion of 92 kDa gelatinase (MMP-9) by bovine neutrophils. Vet. Immunol. Immunopathol. 1999, 67, 247–258. [Google Scholar] [CrossRef] [PubMed]
- Ohtsuka, H.; Ohki, K.; Tajima, M.; Yoshino, T.; Takahashi, K. Evaluation of blood acid-base balance after experimental administration of endotoxin in adult cow. J. Vet. Med. Sci. 1997, 59, 483–485. [Google Scholar] [CrossRef] [PubMed]
- Raulo, S.; Sorsa, T.; Tervahartiala, T.; Latvanen, T.; Pirilä, E.; Hirvonen, J.; Maisi, P. Increase in milk metalloproteinase activity and vascular permeability in bovine endotoxin-induced and naturally occurring Escherichia coli mastitis. Vet. Immunol. Immunopathol. 2002, 85, 137–145. [Google Scholar] [CrossRef] [PubMed]
- Carlstedt, F.; Eriksson, M.; Kiiski, R.; Larsson, A.; Lind, L. Hypocalcemia during porcine endotoxemic shock: Effects of calcium administration. Crit. Care Med. 2000, 28, 2909–2914. [Google Scholar] [CrossRef] [PubMed]
- Rebollar, E.; Arruebo, M.P.; Plaza, M.A.; Murillo, M.D. Effect of lipopolysaccharide on rabbit small intestine muscle contractility in vitro: Role of prostaglandins. Neurogastroenterol. Motil. 2002, 14, 633–642. [Google Scholar] [CrossRef] [PubMed]
- Weisbrodt, N.W.; Pressley, T.A.; Li, Y.-F.; Zembowicz, M.J.; Higham, S.C.; Zembowicz, A.; Lodato, R.F.; Moody, F.G. Decreased ileal muscle contractility and increased NOS II expression induced by lipopolysaccharide. Am. J. Physiol. Liver Physiol. 1996, 271, G454–G460. [Google Scholar] [CrossRef] [PubMed]
- Collares, E.F. Effect of bacterial lipopolysaccharide on gastric emptying of liquids in rats. Braz. J. Med. Biol. Res. 1997, 30, 207–211. [Google Scholar] [CrossRef] [PubMed]
- Barona, I.; Fagundes, D.S.; Gonzalo, S.; Grasa, L.; Arruebo, M.P.; Plaza, M.; Murillo, M.D. Role of TLR4 and MAPK in the local effect of LPS on intestinal contractility. J. Pharm. Pharmacol. 2011, 63, 657–662. [Google Scholar] [CrossRef] [PubMed]
- Lodato, R.F.; Khan, A.R.; Zembowicz, M.J.; Weisbrodt, N.W.; Pressley, T.A.; Li, Y.-F.; Lodato, J.A.; Zembowicz, A.; Moody, F.G. Roles of IL-1 and TNF in the decreased ileal muscle contractility induced by lipopolysaccharide. Am. J. Physiol. Liver Physiol. 1999, 276, G1356–G1362. [Google Scholar] [CrossRef] [PubMed]
- Eades, S.C. Endotoxemia in dairy cattle: Role of eicosanoids in reticulorumen stasis. J. Dairy Sci. 1993, 76, 414–420. [Google Scholar] [CrossRef] [PubMed]
- Plaza, M.A.; Fioramonti, J.; Bueno, L. Role of central interleukin-1β in gastrointestinal motor disturbances induced by lipopolysaccharide in sheep. Dig. Dis. Sci. 1997, 42, 242–250. [Google Scholar] [CrossRef] [PubMed]
- Meurer, M.; Höcherl, K. Endotoxaemia differentially regulates the expression of renal Ca2+ transport proteins in mice. Acta Physiol. 2018, 225, e13175. [Google Scholar] [CrossRef] [PubMed]
- Woudenberg-Vrenken, T.E.; Bindels, R.J.M.; Hoenderop, J.G.J. The role of transient receptor potential channels in kidney disease. Nat. Rev. Nephrol. 2009, 5, 441–449. [Google Scholar] [CrossRef]
- Curtis, C.R.; Erb, H.N.; Sniffen, C.J.; Smith, R.D.; Powers, P.A.; Smith, M.C.; White, M.E.; Hillman, R.B.; Pearson, E.J. Association of parturient hypocalcemia with eight periparturient disorders in Holstein cows. J. Am. Vet. Med. Assoc. 1983, 183, 559–561. [Google Scholar] [PubMed]
- Melendez, P.; Donovan, G.A.; Risco, C.A.; Goff, J.P. Plasma mineral and energy metabolite concentrations in dairy cows fed an anionic prepartum diet that did or did not have retained fetal membranes after parturition. Am. J. Vet. Res. 2004, 65, 1071–1076. [Google Scholar] [CrossRef] [PubMed]
- Risco, C.A.; Reynolds, J.P.; Hird, D. Uterine prolapse and hypocalcemia in dairy cows. J. Am. Vet. Med. Assoc. 1984, 185, 1517–1519. [Google Scholar] [PubMed]
- Massey, C.D.; Wang, C.; Donovan, G.A.; Beede, D.K. Hypocalcemia at parturition as a risk factor for left displacement of the abomasum in dairy cows. J. Am. Vet. Med. Assoc. 1993, 203, 852–853. [Google Scholar] [CrossRef] [PubMed]
- Jørgensen, R.J.; Nyengaard, N.R.; Daniel, R.C.; Mellau, L.S.; Enemark, J.M. Induced hypocalcaemia by Na2EDTA infusion. A review. Zentralblatt fur Veterinarmedizin Reihe A 1999, 46, 389–407. [Google Scholar] [CrossRef] [PubMed]
- Jørgensen, R.J.; Nyengaard, N.R.; Ham, S.; Enemark, J.M.; Andersen, P.H. Rumen motility during induced hyper- and hypocalcaemia. Acta Vet. Scand. 1998, 39, 331–338. [Google Scholar] [CrossRef] [PubMed]
- Al-Eknah, M.M.; Noakes, D.E. A preliminary study on the effect of induced hypocalcaemia and nifedipine on uterine activity in the parturient cow. J. Vet. Pharmacol. Ther. 1989, 12, 237–239. [Google Scholar] [CrossRef]
- Lean, I.; Santos, J.E.P.; Block, E.; Golder, H. Effects of prepartum dietary cation-anion difference intake on production and health of dairy cows: A meta-analysis. J. Dairy Sci. 2019, 102, 2103–2133. [Google Scholar] [CrossRef] [PubMed]
- Ducusin, R.; Uzuka, Y.; Satoh, E.; Otani, M.; Nishimura, M.; Tanabe, S.; Sarashina, T. Effects of extracellular Ca2+ on phagocytosis and intracellular Ca2+ concentrations in polymorphonuclear leukocytes of postpartum dairy cows. Res. Vet. Sci. 2003, 75, 27–32. [Google Scholar] [CrossRef] [PubMed]
- Lean, I.J.; DeGaris, P.J.; Celi, P.; McNeill, D.M.; Rodney, R.M.; Fraser, D.R. Influencing the future: Interactions of skeleton, energy, protein and calcium during late gestation and early lactation. Anim. Prod. Sci. 2014, 54, 1177–1189. [Google Scholar] [CrossRef]
- Chamberlin, W.G.; Middleton, J.R.; Spain, J.N.; Johnson, G.C.; Ellersieck, M.R.; Pithua, P. Subclinical hypocalcemia, plasma biochemical parameters, lipid metabolism, postpartum disease, and fertility in postparturient dairy cows. J. Dairy Sci. 2013, 96, 7001–7013. [Google Scholar] [CrossRef] [PubMed]
- Curtis, C.R.; Erb, H.N.; Sniffen, C.J.; Smith, R.D.; Kronfeld, D.S. Path analysis of dry period nutrition, postpartum metabolic and reproductive disorders, and mastitis in Holstein cows. J. Dairy Sci. 1985, 68, 2347–2360. [Google Scholar] [CrossRef] [PubMed]
- Goff, J.P.; Horst, R.L. Physiological changes at parturition and their relationship to metabolic disorders. J. Dairy Sci. 1997, 80, 1260–1268. [Google Scholar] [CrossRef] [PubMed]
- Heppelmann, M.; Krach, K.; Krueger, L.; Benz, P.; Herzog, K.; Piechotta, M.; Hoedemaker, M.; Bollwein, H. The effect of metritis and subclinical hypocalcemia on uterine involution in dairy cows evaluated by sonomicrometry. J. Reprod. Dev. 2015, 61, 565–569. [Google Scholar] [CrossRef] [PubMed]
- Jonsson, N.N.; Pepper, P.M.; Daniel, R.C.W.; McGowan, M.C.; Fulkerson, W. Association between non-parturient post-partum hypocalcaemia and the interval from calving to first ovulation in Holstein-Friesian cows. Anim. Sci. 1999, 69, 377–383. [Google Scholar] [CrossRef]
- Kamgarpour, R.; Daniel, R.C.; Fenwick, D.C.; Mcguigan, K.; Murphy, G. Post partum subclinical hypocalcaemia and effects on ovarian function and uterine involution in a dairy herd. Vet. J. 1999, 158, 59–67. [Google Scholar] [CrossRef] [PubMed]
- Bréchard, S.; Tschirhart, E.J. Regulation of superoxide production in neutrophils: Role of calcium influx. J. Leukoc. Biol. 2008, 84, 1223–1237. [Google Scholar] [CrossRef] [PubMed]
- Burgos, R.; Conejeros, I.; Hidalgo, M.; Werling, D.; Hermosilla, C. Calcium influx, a new potential therapeutic target in the control of neutrophils-dependent inflammatory diseases in bovines. Vet. Immunol. Immunopathol. 2011, 143, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Prakriya, M.; Lewis, R.S. Store-operated calcium channels. Physiol. Rev. 2015, 95, 1383–1436. [Google Scholar] [CrossRef] [PubMed]
- Parekh, A.B.; Putney, J.W., Jr. Store-operated calcium channels. Physiol. Rev. 2005, 85, 757–810. [Google Scholar] [CrossRef] [PubMed]
- Steinckwich, N.; Frippiat, J.-P.; Stasia, M.-J.; Erard, M.; Boxio, R.; Tankosic, C.; Doignon, I.; Nüsse, O. Potent inhibition of store-operated Ca2+ influx and superoxide production in HL60 cells and polymorphonuclear neutrophils by the pyrazole derivative BTP2. J. Leukoc. Biol. 2007, 81, 1054–1064. [Google Scholar] [CrossRef] [PubMed]
- Yu, Y.; De Waele, C.; Chadee, K. Calcium-dependent interleukin-8 gene expression in T84 human colonic epithelial cells. Inflamm. Res. 2001, 50, 220–226. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.M.; An, J. Cytokines, inflammation and pain. Int. Anesthesiol. Clin. 2007, 45, 27–37. [Google Scholar] [CrossRef]
- Crabtree, G.R. Calcium, calcineurin, and the control of transcription. J. Biol. Chem. 2001, 276, 2313–2316. [Google Scholar] [CrossRef] [PubMed]
- Shaw, K.T.; Ho, A.M.; Raghavan, A.; Kim, J.; Jain, J.; Park, J.; Sharma, S.; Rao, A.; Hogan, P.G. Immunosuppressive drugs prevent a rapid dephosphorylation of transcription factor NFAT1 in stimulated immune cells. Proc. Natl. Acad. Sci. USA 1995, 92, 11205–11209. [Google Scholar] [CrossRef] [PubMed]
- Timmerman, L.A.; Clipstone, N.A.; Ho, S.N.; Northrop, J.P.; Crabtree, G.R. Rapid shuttling of NF-AT in discrimination of Ca2+ signals and immunosuppression. Nature 1996, 383, 837–840. [Google Scholar] [CrossRef] [PubMed]
- Fric, J.; Zelante, T.; Wong, A.Y.W.; Mertes, A.; Yu, H.-B.; Ricciardi-Castagnoli, P. NFAT control of innate immunity. Blood 2012, 120, 1380–1389. [Google Scholar] [CrossRef] [PubMed]
- Ender, F.; Dishington, I.W.; Helgebostad, A. Parturient paresis and related forms of hypocalcemic disorders induced experimentally in dairy cows. Acta Vet. Scand. 1962, 3, 5–52. [Google Scholar]
- Ender, F.; Dishington, I.W.; Helgebostad, A. Calcium balance studies in dairy cows under experimental induction and prevention of hypocalcaemic paresis puerperalis. Z. Tierernahr. Futtermittelkd. 1971, 28, 233–256. [Google Scholar] [CrossRef] [PubMed]
- Charbonneau, E.; Pellerin, D.; Oetzel, G.R. Impact of lowering dietary cation-anion difference in nonlactating dairy cows: A meta-analysis. J. Dairy Sci. 2006, 89, 537–548. [Google Scholar] [CrossRef] [PubMed]
- Horst, R.; Goff, J.; Reinhardt, T.; Buxton, D. Strategies for preventing milk fever in dairy cattle. J. Dairy Sci. 1997, 80, 1269–1280. [Google Scholar] [CrossRef] [PubMed]
- Block, E. Manipulating dietary anions and cations for prepartum dairy cows to reduce incidence of milk fever. J. Dairy Sci. 1984, 67, 2939–2948. [Google Scholar] [CrossRef] [PubMed]
- Zimpel, R.; Marinho, M.N.; Almeida, K.; Ruiz, A.R.; Perdomo, M.; Poindexter, M.; Vieira-Neto, A.; Arshad, U.; Husnain, A.; Nelson, C.; et al. Prepartum level of dietary cation-anion difference fed to nulliparous cows: Acid-base balance, mineral metabolism, and health responses. J. Dairy Sci. 2021, 104, 12580–12599. [Google Scholar] [CrossRef] [PubMed]
- Zimpel, R.; Almeida, K.; Marinho, M.N.; Ruiz, A.R.; Perdomo, M.; Poindexter, M.; Husnain, A.; Vieira-Neto, A.; Silva, A.; Arshad, U.; et al. Prepartum level of dietary cation-anion difference fed to nulliparous cows: Lactation and reproductive responses. J. Dairy Sci. 2021, 104, 11699–11714. [Google Scholar] [CrossRef] [PubMed]
- Constable, P.D. A simplified strong ion model for acid-base equilibria: Application to horse plasma. J. Appl. Physiol. 1997, 83, 297–311. [Google Scholar] [CrossRef] [PubMed]
- Bigner, D.; Goff, J.; Faust, M.; Burton, J.; Tyler, H.; Horst, R. Acidosis effects on insulin response during glucose tolerance tests in Jersey cows. J. Dairy Sci. 1996, 79, 2182–2188. [Google Scholar] [CrossRef]
- Vieira-Neto, A.; Zimpel, R.; Lopes, F.; Scheffler, T.; Block, E.; Thatcher, W.; Santos, J.E.P. Duration and degree of diet-induced metabolic acidosis prepartum alter tissues responses to insulin in dairy cows. J. Dairy Sci. 2021, 104, 1660–1679. [Google Scholar] [CrossRef] [PubMed]
- Oberleithner, H.; Greger, R.; Lang, F. The effect of respiratory and metabolic acid-base changes on ionized calcium concentration: In vivo and in vitro experiments in man and rat. Eur. J. Clin. Investig. 1982, 12, 451–455. [Google Scholar] [CrossRef] [PubMed]
- Lopez, I.; Aguilera-Tejero, E.; Felsenfeld, A.J.; Estepa, J.C.; Rodriguez, M. Direct effect of acute metabolic and respiratory acidosis on parathyroid hormone secretion in the dog. J. Bone Miner. Res. 2002, 17, 1691–1700. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; McDonnell, E.H.; Sedor, F.A.; Toffaletti, J.G. pH effects on measurements of ionized calcium and ionized magnesium in blood. Arch. Pathol. Lab. Med. 2002, 126, 947–950. [Google Scholar] [CrossRef] [PubMed]
- Goff, J.; Liesegang, A.; Horst, R. Died-induced pseudohypoparathyroidism: A hypocalcemia and milk fever risk factor. J. Dairy Sci. 2014, 97, 1520–1528. [Google Scholar] [CrossRef]
- Vieira-Neto, A.; Leão, I.; Prim, J.; Silva, A.; Marinho, M.N.; Zimpel, R.; Etheve, S.; Nelson, C.; Santos, J.E.P. Effect of duration of exposure to diets differing in DCAD on Ca metabolism after a parathyroid hormone challenge in dairy cows. J. Dairy Sci. 2021, 104, 1018–1038. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez, E.; Bach, A.; Devant, M.; Aris, A. Is calcitonin an active hormone in the onset and prevention of hypocalcemia in dairy cattle? J. Dairy Sci. 2016, 99, 3023–3030. [Google Scholar] [CrossRef] [PubMed]
- Bushinsky, D.A.; Goldring, J.M.; Coe, F.L. Cellular contribution to pH-mediated calcium flux in neonatal mouse calvariae. Am. J. Physiol. 1985, 248, F785–F789. [Google Scholar] [CrossRef] [PubMed]
- Krieger, N.S.; Parker, W.R.; Alexander, K.M.; Bushinsky, D.A. Prostaglandins regulate acid-induced cell-mediated bone resorption. Am. J. Physiol. Physiol. 2000, 279, F1077–F1082. [Google Scholar] [CrossRef] [PubMed]
- Frick, K.K.; Bushinsky, D.A. Metabolic acidosis stimulates RANKL RNA expression in bone through a cyclo-oxygenase-dependent mechanism. J. Bone Miner. Res. 2003, 18, 1317–1325. [Google Scholar] [CrossRef] [PubMed]
- Wilkens, M.; Praechter, C.; Breves, G.; Schröder, B. Stimulating effects of a diet negative in dietary cation-anion difference on calcium absorption from the rumen in sheep. J. Anim. Physiol. Anim. Nutr. 2016, 100, 156–166. [Google Scholar] [CrossRef]
- Weich, W.; Block, E.; Litherland, N.B. Extended negative dietary cation-anion difference feeding does not negatively affect postpartum performance of multiparous dairy cows. J. Dairy Sci. 2013, 96, 5780–5792. [Google Scholar] [CrossRef] [PubMed]
- Wu, Z.; Bernard, J.K.; Zanzalari, K.P.; Chapman, J.D. Effect of feeding a negative dietary cation-anion difference diet for an extended time prepartum on postpartum serum and urine metabolites and performance. J. Dairy Sci. 2014, 97, 7133–7143. [Google Scholar] [CrossRef] [PubMed]
- DeGaris, P.; Lean, I.; Rabiee, A.; Heuer, C. Effects of increasing days of exposure to prepartum transition diets on milk production and milk composition in dairy cows. Aust. Vet. J. 2008, 86, 341–351. [Google Scholar] [CrossRef] [PubMed]
- Vieira-Neto, A.; Duarte, G.; Zimpel, R.; Thatcher, W.; Santos, J.E.P. Days in the prepartum group are associated with subsequent performance in Holstein cows. J. Dairy Sci. 2021, 104, 5964–5978. [Google Scholar] [CrossRef] [PubMed]
- Goings, R.; Jacobson, N.; Beitz, D.; Littledike, E.; Wiggers, K. Prevention of parturient paresis by a prepartum, calcium-deficient diet. J. Dairy Sci. 1974, 57, 1184–1188. [Google Scholar] [CrossRef] [PubMed]
- Kichura, T.S.; Horst, R.L.; Beitz, D.C.; Littledike, E.T. Relationships between prepartal dietary calcium and phosphorus, vitamin D metabolism, and parturient paresis in dairy cows. J. Nutr. 1982, 112, 480–487. [Google Scholar] [CrossRef] [PubMed]
- Green, H.; Horst, R.; Beitz, D.; Littledike, E. Vitamin D metabolites in plasma of cows fed a prepartum low-calcium diet for prevention of parturient hypocalcemia. J. Dairy Sci. 1981, 64, 217–226. [Google Scholar] [CrossRef] [PubMed]
- NASEM Dairy (National Academies of Sciences, Engineering, and Medicine). Nutrient Requirements of Dairy Cattle, 8th ed.; The National Academies Press: Washington, DC, USA, 2021. [Google Scholar] [CrossRef]
- Thilsing-Hansen, T.; Jørgensen, R. Hot topic: Prevention of parturient paresis and subclinical hypocalcemia in dairy cows by zeolite A administration in the dry period. J. Dairy Sci. 2001, 84, 691–693. [Google Scholar] [CrossRef] [PubMed]
- Thilsing, T.; Jørgensen, R.J.; Poulsen, H.D. In vitro binding capacity of zeolite A to calcium, phosphorus and magnesium in rumen fluid as influenced by changes in pH. J. Vet. Med. 2006, 53, 57–64. [Google Scholar] [CrossRef] [PubMed]
- Kerwin, A.; Ryan, C.; Leno, B.; Jakobsen, M.; Theilgaard, P.; Barbano, D.; Overton, T. Effects of feeding synthetic zeolite A during the prepartum period on serum mineral concentration, oxidant status, and performance of multiparous Holstein cows. J. Dairy Sci. 2019, 102, 5191–5207. [Google Scholar] [CrossRef] [PubMed]
- Allen, V.G. Influence of dietary aluminum on nutrient utilization in ruminants. J. Anim. Sci. 1984, 59, 836–844. [Google Scholar] [CrossRef] [PubMed]
- Horst, R.; Goff, J.; Reinhardt, T. Calcium and vitamin D metabolism in the dairy cow. J. Dairy Sci. 1994, 77, 1936–1951. [Google Scholar] [CrossRef]
- Wilkens, M.; Oberheide, I.; Schröder, B.; Azem, E.; Steinberg, W.; Breves, G. Influence of the combination of 25-hydroxyvitamin D3 and a diet negative in cation-anion difference on peripartal calcium homeostasis of dairy cows. J. Dairy Sci. 2012, 95, 151–164. [Google Scholar] [CrossRef] [PubMed]
- Martinez, N.; Rodney, R.M.; Block, E.; Hernandez, L.L.; Nelson, C.D.; Lean, I.J.; Santos, J.E.P. Effects of prepartum dietary cation-anion difference and source of vitamin D in dairy cows: Health and reproductive responses. J. Dairy Sci. 2018, 101, 2563–2578. [Google Scholar] [CrossRef] [PubMed]
- Martinez, N.; Rodney, R.M.; Block, E.; Hernandez, L.L.; Nelson, C.D.; Lean, I.J.; Santos, J.E.P. Effects of prepartum dietary cation-anion difference and source of vitamin D in dairy cows: Lactation performance and energy metabolism. J. Dairy Sci. 2018, 101, 2544–2562. [Google Scholar] [CrossRef] [PubMed]
- Poindexter, M.; Zimpel, R.; Vieira-Neto, A.; Husnain, A.; Silva, A.; Faccenda, A.; de Avila, A.S.; Celi, P.; Cortinhas, C.; Santos, J.E.P.; et al. Effect of prepartum source and amount of vitamin D supplementation on lactation performance of dairy cows. J. Dairy Sci. 2023, 106, 974–989. [Google Scholar] [CrossRef] [PubMed]
- Weiss, W.; Azem, E.; Steinberg, W.; Reinhardt, T. Effect of feeding 25-hydroxyvitamin D3 with a negative cation-anion difference diet on calcium and vitamin D status of periparturient cows and their calves. J. Dairy Sci. 2015, 98, 5588–5600. [Google Scholar] [CrossRef] [PubMed]
- Gast, D.; Horst, R.; Jorgensen, N.; DeLuca, H. Potential use of 1,25-dihydroxycholecalciferol for prevention of parturient paresis. J. Dairy Sci. 1979, 62, 1009–1013. [Google Scholar] [CrossRef] [PubMed]
- Hove, K.; Kristiansen, T. Prevention of parturient hypocalcemia: Effect of a single oral dose of 1,25-dihydroxyvitamin D3. J. Dairy Sci. 1982, 65, 1934–1940. [Google Scholar] [CrossRef] [PubMed]
- Goff, J.P.; Horst, R.L.; Beitz, D.C.; Littledike, E.T. Use of 24-F-1,25-dihydroxyvitamin D3 to prevent parturient paresis in dairy cows. J. Dairy Sci. 1988, 71, 1211–1219. [Google Scholar] [CrossRef] [PubMed]
- USDA. Dairy 2014, “Dairy Cattle Management Practices in the United States, 2014” USDA–APHIS–VS–CEAH–NAHMS. Fort Collins, CO#692.0216. 2016. Available online: https://www.aphis.usda.gov/sites/default/files/dairy-trends-hlth-mngmnt-1991-2014.pdf (accessed on 1 September 2023).
- Goff, J.P.; Horst, R.L. Oral administration of calcium salts for treatment of hypocalcemia in cattle. J. Dairy Sci. 1993, 76, 101–108. [Google Scholar] [CrossRef] [PubMed]
- Martinez, N.; Sinedino, L.D.; Bisinotto, R.S.; Daetz, R.; Risco, C.A.; Galvão, K.N.; Thatcher, W.W.; Santos, J.E.P. Effects of oral calcium supplementation on productive and reproductive performance in dairy cows. J. Dairy Sci. 2016, 99, 8417–8430. [Google Scholar] [CrossRef] [PubMed]
- Domino, A.R.; Korzec, H.C.; McArt, J.A. Field trial of 2 calcium supplements on early lactation health and production in multiparous Holstein cows. J. Dairy Sci. 2017, 100, 9681–9690. [Google Scholar] [CrossRef] [PubMed]
- Valldecabres, A.; Branco-Lopes, R.; Bernal-Córdoba, C.; Silva-Del-Río, N. Production and reproduction responses for dairy cattle supplemented with oral calcium bolus after calving: Systematic review and meta-analysis. JDS Commun. 2023, 4, 9–13. [Google Scholar] [CrossRef] [PubMed]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vieira-Neto, A.; Lean, I.J.; Santos, J.E.P. Periparturient Mineral Metabolism: Implications to Health and Productivity. Animals 2024, 14, 1232. https://doi.org/10.3390/ani14081232
Vieira-Neto A, Lean IJ, Santos JEP. Periparturient Mineral Metabolism: Implications to Health and Productivity. Animals. 2024; 14(8):1232. https://doi.org/10.3390/ani14081232
Chicago/Turabian StyleVieira-Neto, Achilles, Ian J. Lean, and José Eduardo P. Santos. 2024. "Periparturient Mineral Metabolism: Implications to Health and Productivity" Animals 14, no. 8: 1232. https://doi.org/10.3390/ani14081232
APA StyleVieira-Neto, A., Lean, I. J., & Santos, J. E. P. (2024). Periparturient Mineral Metabolism: Implications to Health and Productivity. Animals, 14(8), 1232. https://doi.org/10.3390/ani14081232