
Broiler Spaghetti Meat Abnormalities: Muscle Characteristics and Metabolomic Profiles


 and
and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Ethics Statement
2.2. Animals and Feeding Management
2.3. Sample Collection
2.4. Analysis of Physical Properties and Proximate Composition
2.5. Analysis of Muscle Fiber Morphology
2.6. Liquid Chromatography with Tandem Mass Spectrometry Untargeted Metabolomic Analysis
2.7. Statistical Analysis
3. Results
3.1. Comparison of Physical Properties and Proximate Compositions
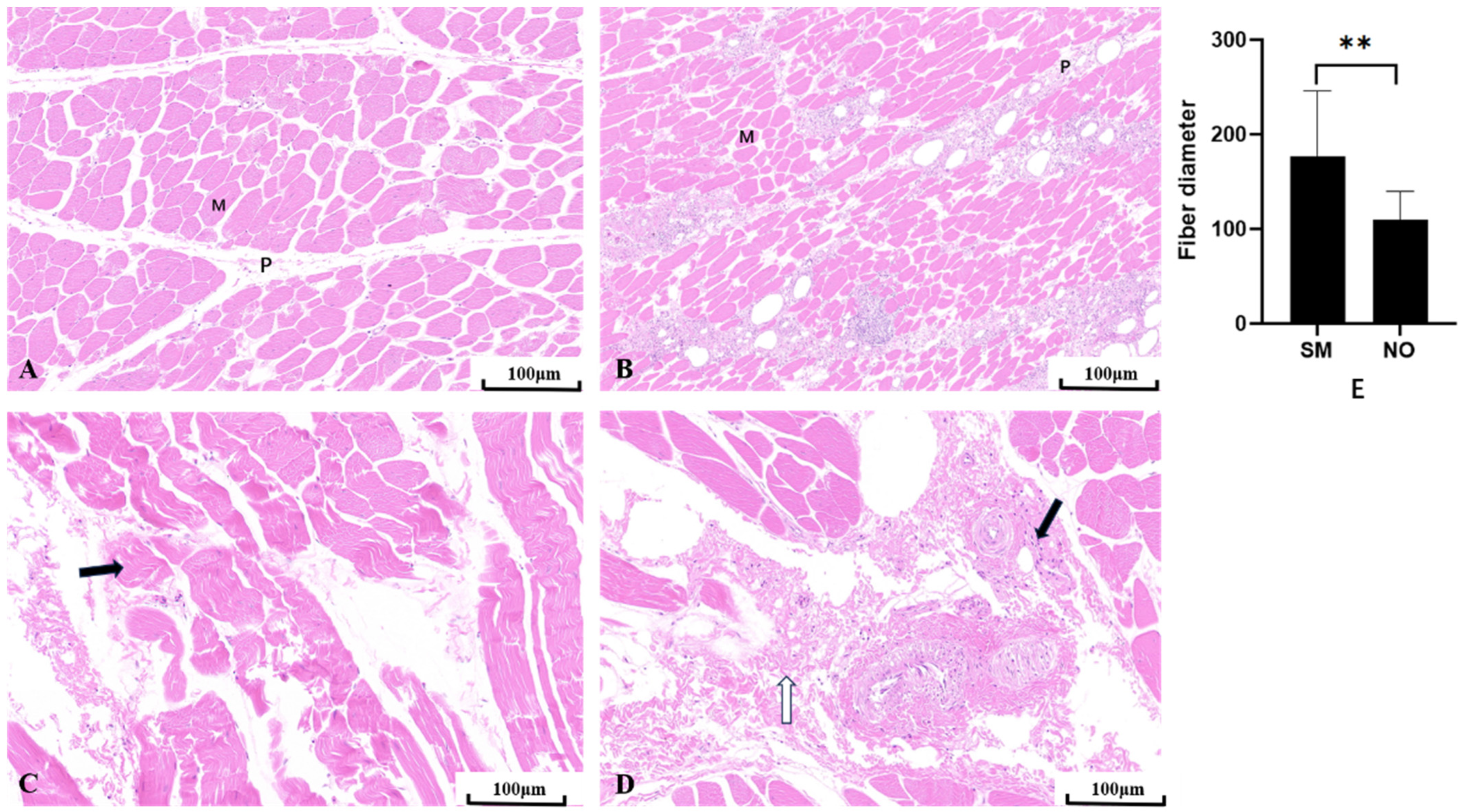
3.2. Comparison of Muscle Fiber Morphologies
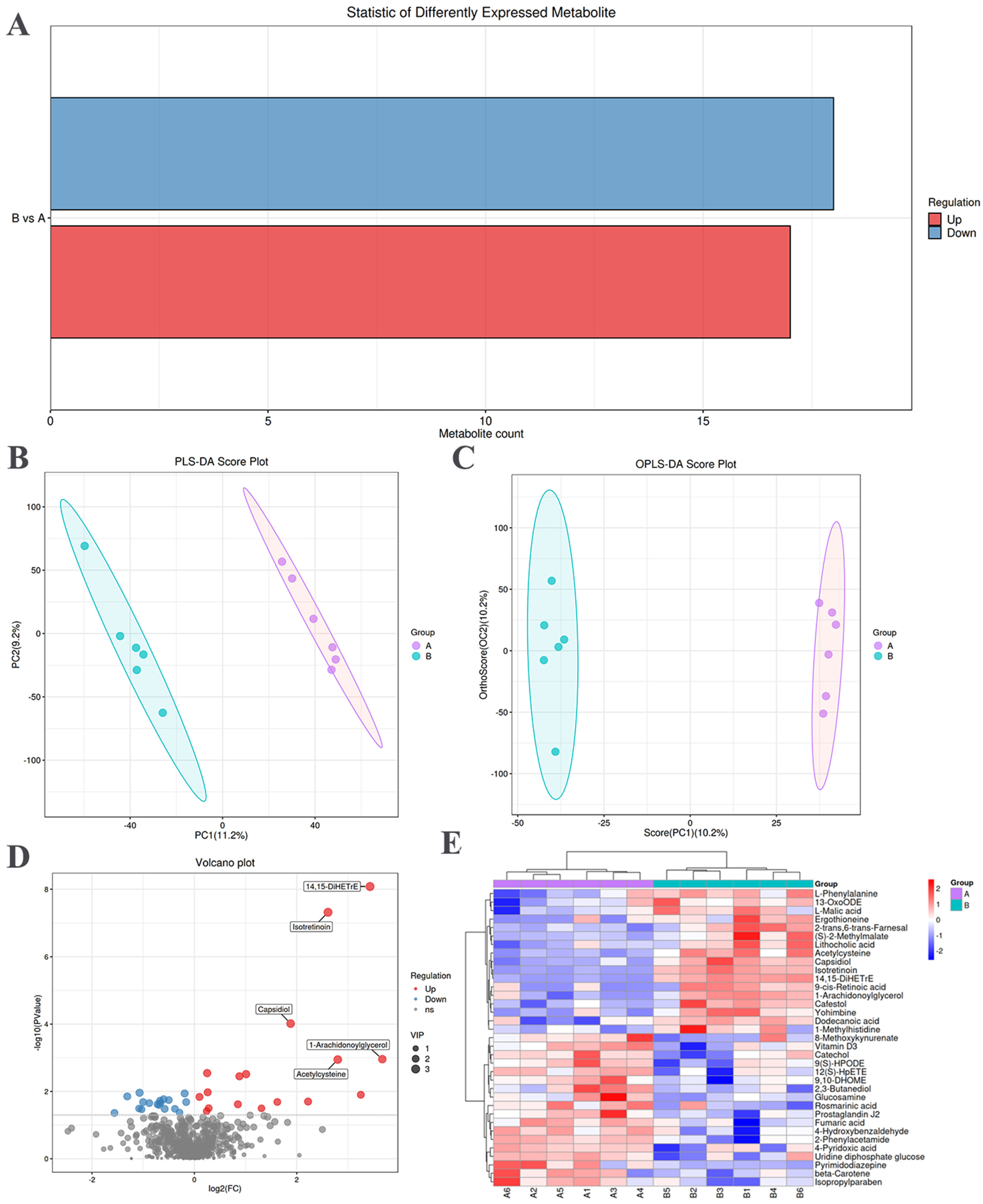
3.3. Identification of Differential Metabolites in SM
3.3.1. Screening of Differential Metabolites
3.3.2. Multivariate Analysis
3.3.3. Hierarchical Clustering Analysis of Differential Metabolites
3.3.4. Kyoto Encyclopedia of Genes and Genomes Enrichment Analysis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Wang, C.; Che, S.; Susta, L.; Barbut, S. Textural and physical properties of breast fillets with myopathies (wooden breast, white striping, spaghetti meat) in Canadian fast-growing broiler chickens. Poult. Sci. 2023, 2, 102309. [Google Scholar] [CrossRef] [PubMed]
- Petracci, M.; Velleman, S.G. Editorial: Avian Muscle Development and Growth Mechanisms: Association With Muscle Myopathies and Meat Quality Volume II. Front. Physiol. 2021, 12, 765515. [Google Scholar] [CrossRef] [PubMed]
- Bailey, R.A. Strategies and opportunities to control breast myopathies: An opinion paper. Front. Physiol. 2023, 14, 1173564. [Google Scholar] [CrossRef] [PubMed]
- Baldi, G.; Yen, C.N.; Daughtry, M.R.; Bodmer, J.; Bowker, B.C.; Zhuang, H.; Petracci, M.; Gerrard, D.E. Exploring the Factors Contributing to the High Ultimate pH of Broiler Pectoralis Major Muscles Affected by Wooden Breast Condition. Front. Physiol. 2020, 11, 343. [Google Scholar] [CrossRef] [PubMed]
- Velleman, S.G.; Petracci, M. Editorial: Avian Muscle Development and Growth Mechanisms: Association With Muscle Myopathies and Meat Quality. Front. Physiol. 2020, 11, 601184. [Google Scholar] [CrossRef] [PubMed]
- Bailey, R.A.; Souza, E.; Avendano, S. Characterising the Influence of Genetics on Breast Muscle Myopathies in Broiler Chickens. Front. Physiol. 2020, 11, 1041. [Google Scholar] [CrossRef] [PubMed]
- Bordignon, F.; Xiccato, G.; Boskovic, M.; Birolo, M.; Trocino, A. Factors Affecting Breast Myopathies in Broiler Chickens and Quality of Defective Meat: A Meta-Analysis. Front. Physiol. 2022, 13, 933235. [Google Scholar] [CrossRef] [PubMed]
- Soglia, F.; Mudalal, S.; Babini, E.; Di, N.M.; Mazzoni, M.; Sirri, F.; Cavani, C.; Petracci, M. Histology, composition, and quality traits of chicken Pectoralis major muscle affected by wooden breast abnormality. Poult. Sci. 2016, 95, 651–659. [Google Scholar] [CrossRef] [PubMed]
- Soglia, F.; Petracci, M.; Davoli, R.; Zappaterra, M. A critical review of the mechanisms involved in the occurrence of growth-related abnormalities affecting broiler chicken breast muscles. Poult. Sci. 2021, 100, 101180. [Google Scholar] [CrossRef]
- Chi, M.Y.; Zhang, H.; Wang, Y.X.; Sun, X.P.; Yang, Q.J.; Guo, C. Silibinin Alleviates Muscle Atrophy Caused by Oxidative Stress Induced by Cisplatin through ERK/FoxO and JNK/FoxO Pathways. Oxid. Med. Cell Longev. 2022, 2022, 5694223. [Google Scholar] [CrossRef]
- Boerboom, G.; Kempen, T.; Navarro, V.A.; Pérez, B.A. Unraveling the cause of white striping in broilers using metabolomics. Poult. Sci. 2018, 97, 3977–3986. [Google Scholar] [CrossRef] [PubMed]
- Peng, M.L.; Han, J.; Li, L.L.; Ma, H.T. Metabolomics reveals the mechanism of (-)-hydroxycitric acid promotion of protein synthesis and inhibition of fatty acid synthesis in broiler chickens. Animal 2018, 12, 774–783. [Google Scholar] [CrossRef] [PubMed]
- NY/T33-2004; Feeding Standard of Chicken. Chinese Academy of Agricultural Sciences: Beijing, China, 2004.
- Maharjan, P.; Hilton, K.; Weil, J.; Suesuttajit, N.; Beitia, A.; Owens, C.M.; Coon, C. Characterizing Woody Breast Myopathy in a Meat Broiler Line by Heat Production, Microbiota, and Plasma Metabolites. Front. Vet. Sci. 2020, 6, 497. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Wang, P.; Xu, X.; Xia, T.; Li, Z.; Zhao, T. Effect of wooden breast myopathy on water-holding capacity and rheological and gelling properties of chicken broiler breast batters. Poult. Sci. 2020, 99, 3742–3751. [Google Scholar] [CrossRef] [PubMed]
- Sun, X.; Koltes, D.A.; Coon, C.N.; Chen, K.; Owens, C.M. Instrumental compression force and meat attribute changes in woody broiler breast fillets during short-term storage. Poult. Sci. 2018, 97, 2600–2606. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Tauler, R. Flexible Implementation of the Trilinearity Constraint in Multivariate Curve Resolution Alternating Least Squares (MCR-ALS) of Chromatographic and Other Type of Data. Molecules 2022, 27, 2338. [Google Scholar] [CrossRef]
- Zelena, E.; Dunn, W.B.; Broadhurst, D.; Francis, M.S.; Carroll, K.M.; Begley, P.; Hagan, S.; Knowles, J.D.; Halsall, A. HUSERMET Consortium; Wilson ID, Kell DB. Development of a robust and repeatable UPLC-MS method for the long-term metabolomic study of human serum. Anal. Chem. 2009, 81, 1357–1364. [Google Scholar] [CrossRef]
- Want, E.J.; Masson, P.; Michopoulos, F.; Wilson, I.D.; Theodoridis, G.; Plumb, R.S.; Shockcor, J.; Loftus, N.; Holmes, E.; Nicholson, J.K. Global metabolic profiling of animal and human tissues via UPLC-MS. Nat. Protoc. 2013, 8, 17–32. [Google Scholar] [CrossRef] [PubMed]
- Thévenot, E.A.; Roux, A.; Xu, Y.; Ezan, E.; Junot, C. Analysis of the Human Adult Urinary Metabolome Variations with Age, Body Mass Index, and Gender by Implementing a Comprehensive Workflow for Univariate and OPLS Statistical Analyses. J. Proteome Res. 2015, 14, 3322–3335. [Google Scholar] [CrossRef]
- Domingo, X.; Siuzdak, G. Metabolomics Data Processing Using XCMS. Methods Mol. Biol. 2020, 2104, 11–24. [Google Scholar]
- Wishart, D.S.; Guo, A.; Oler, E.; Wang, F.; Anjum, A.; Peters, H.; Dizon, R.; Sayeeda, Z.; Tian, S.; Lee, B.L.; et al. HMDB 5.0: The Human Metabolome Database for 2022. Nucleic Acids Res. 2022, 50, 622–631. [Google Scholar] [CrossRef] [PubMed]
- Pang, Z.; Chong, J.; Zhou, G.; Morais, D.A.; Chang, L.; Barrette, M.; Gauthier, C.; Jacques, P.É.; Li, S.; Xia, J. MetaboAnalyst 5.0: Narrowing the gap between raw spectra and functional insights. Nucleic Acids Res. 2021, 49, 388–396. [Google Scholar] [CrossRef] [PubMed]
- Abu, D.; Cairns, J.L.; Boskamp, T.; Marsching, C.; Kessler, T.; Ramallo, C.; Panitz, V.; Sadik, A.; Cordes, J. Spatial probabilistic mapping of metabolite ensembles in mass spectrometry imaging. Nat. Commun. 2023, 14, 1823. [Google Scholar]
- Baéza, E.; Guillier, L.; Petracci, M. Review: Production factors affecting poultry carcass and meat quality attributes. Animal. 2022, 1, 100331. [Google Scholar] [CrossRef] [PubMed]
- Pascual, A.; Trocino, A.; Birolo, M.; Cardazzo, B.; Bordignon, F.; Ballarin, C.; Xiccato, G. Dietary supplementation with sodium butyrate: Growth, gut response at different ages, and meat quality of female and male broiler chickens. Ital. J. Anim. Sci. 2020, 19, 1134–1145. [Google Scholar] [CrossRef]
- Tasoniero, G.; Zhuang, H.; Gamble, G.R.; Bowker, B.C. Effect of spaghetti meat abnormality on broiler chicken breast meat composition and technological quality. Poult. Sci. 2020, 99, 1724–1733. [Google Scholar] [CrossRef]
- Ali, M.; Baek, K.H.; Lee, S.Y.; Kim, H.C.; Park, J.Y.; Jo, C.; Jung, J.H.; Park, H.C.; Nam, K.C. Comparative Meat Qualities of Boston Butt Muscles (M. subscapularis) from Different Pig Breeds Available in Korean Market. Food Sci. Anim. Res. 2021, 41, 71–84. [Google Scholar] [CrossRef] [PubMed]
- Mudalal, S.; Lorenzi, M.; Soglia, F.; Cavani, C.; Petracci, M. Implications of white striping and wooden breast abnormalities on quality traits of raw and marinated chicken meat. Animal 2015, 9, 728–734. [Google Scholar] [CrossRef] [PubMed]
- Griffin, J.R.; Moraes, L.; Wick, M.; Lilburn, M.S. Onset of white striping and progression into wooden breast as defined by myopathic changes underlying Pectoralis major growth. Estimation of growth parameters as predictors for stage of myopathy progression. Avian Pathol. 2018, 47, 12–13. [Google Scholar] [CrossRef]
- Wang, C.; Susta, L.; Barbut, S. Restoring Functionalities in Chicken Breast Fillets with Spaghetti Meat Myopathy by Using Dairy Proteins Gels. Gels 2022, 8, 558. [Google Scholar] [CrossRef]
- Baldi, G.; Soglia, F.; Laghi, L.; Tappi, S.; Rocculi, P.; Tavaniello, S.; Prioriello, D.; Mucci, R.; Maiorano, G.; Petracci, M. Comparison of quality traits among breast meat affected by current muscle abnormalities. Food Res. Int. 2019, 115, 369–376. [Google Scholar] [CrossRef] [PubMed]
- Che, S.; Wang, C.; Varga, C.; Barbut, S.; Susta, L. Prevalence of breast muscle myopathies (spaghetti meat, woody breast, white striping) and associated risk factors in broiler chickens from Ontario Canada. PLoS ONE 2022, 17, 0267019. [Google Scholar] [CrossRef] [PubMed]
- Mazzoni, M.; Soglia, F.; Petracci, M.; Sirri, F.; Lattanzio, G.; Clavenzani, P. Fiber Metabolism, Procollagen and Collagen Type III Immunoreactivity in Broiler Pectoralis Major Affected by Muscle Abnormalities. Animals 2020, 10, 1081. [Google Scholar] [CrossRef] [PubMed]
- Russo, S.; Kwiatkowski, M.; Govorukhina, N.; Bischoff, R.; Melgert, B.N. Meta-Inflammation and Metabolic Reprogramming of Macrophages in Diabetes and Obesity: The Importance of Metabolites. Front. Immunol. 2021, 12, 746151. [Google Scholar] [CrossRef] [PubMed]
- Cockram, P.E.; Kist, M.; Prakash, S.; Chen, S.H.; Wertz, I.E.; Vucic, D. Ubiquitination in the regulation of inflammatory cell death and cancer. Cell Death Differ. 2021, 28, 591–605. [Google Scholar] [CrossRef] [PubMed]
- Trindade, C.A.; Yang, J.; Fonseca, F.; Pham, H.; Napimoga, M.H.; Abdalla, H.B.; Aver, G. Eicosanoid profiles in an arthritis model: Effects of a soluble epoxide hydrolase inhibitor. Biochim. Biophys. Acta Mol. Cell Biol. Lipids 2024, 1869, 159432. [Google Scholar]
- Gartung, A.; Zhao, J.; Chen, S.; Mottillo, E.; VanHecke, G.C.; Ahn, Y.H.; Maddipati, K.R.; Sorokin, A.; Granneman, J.; Lee, M.J. Characterization of Eicosanoids Produced by Adipocyte Lipolysis: Implication of Cyclooxygenase-2 in Adipose Inflammation. J. Biol. Chem. 2016, 291, 16001–16010. [Google Scholar] [CrossRef] [PubMed]
- Caussy, C.; Chuang, J.C.; Billin, A.; Hu, T.; Wang, Y.; Subramanian, G.M.; Djedjos, C.S.; Myers, R.P.; Dennis, E.A.; Loomba, R. Plasma eicosanoids as noninvasive biomarkers of liver fibrosis in patients with nonalcoholic steatohepatitis. Ther. Adv. Gastroenterol. 2020, 13, 175. [Google Scholar] [CrossRef] [PubMed]
- Melnik, B.C. Apoptosis May Explain the Pharmacological Mode of Action and Adverse Effects of Isotretinoin, Including Teratogenicity. Acta Derm. Venereol. 2017, 97, 173–181. [Google Scholar] [CrossRef]
- Yan, E.; Wang, Y.; He, L.; Guo, J.; Zhang, X.; Yin, J. Effects of Dietary L-malic Acid Supplementation on Meat Quality, Antioxidant Capacity and Muscle Fiber Characteristics of Finishing Pigs. Foods 2022, 11, 3335. [Google Scholar] [CrossRef]
- Martínez, R.I.; Chandel, N.S. Mitochondrial TCA cycle metabolites control physiology and disease. Nat. Commun. 2020, 11, 102. [Google Scholar] [CrossRef]
- Chen, K.Y.; Chen, Y.J.; Cheng, C.J.; Jhan, K.Y.; Chiu, C.H.; Wang, L.C. 3-Hydroxybenzaldehyde and 4-Hydroxybenzaldehyde enhance survival of mouse astrocytes treated with Angiostrongylus cantonensis young adults excretory/secretory products. Biomed. J. 2021, 44, 258–266. [Google Scholar] [CrossRef] [PubMed]
- Abe, I.; Oguri, Y.; Verkerke, R.P.; Monteiro, L.B.; Knuth, C.M.; Auger, C.; Qiu, Y.; Westcott, G.P.; Cinti, S.; Shinoda, K.; et al. Lipolysis-derived linoleic acid drives beige fat progenitor cell proliferation. Dev. Cell 2022, 57, 2623–2637. [Google Scholar] [CrossRef] [PubMed]
- Martin, S.A.; Brash, A.R.; Murphy, R.C. The discovery and early structural studies of arachidonic acid. J. Lipid Res. 2016, 57, 1126–1132. [Google Scholar] [CrossRef] [PubMed]
- Lackner, J.; Hess, V.; Stef, L.; Sauerwein, H. Effects of feeding different histidine to lysine ratios on performance, meat quality, and the occurrence of breast myopathies in broiler chickens. Poult. Sci. 2022, 101, 101568. [Google Scholar] [CrossRef] [PubMed]
- Baldi, G.; Soglia, F.; Laghi, L.; Meluzzi, A.; Petracci, M. The role of histidine dipeptides on postmortem acidification of broiler muscles with different energy metabolism. Poult. Sci. 2021, 100, 1299–1307. [Google Scholar] [CrossRef]
- Chacón, P.; Vega, A.; Doukkali, B.; Del, A.; Bellido, V.; Puente, Y.; Alcañiz, L.; Rodríguez, D.; Palacios, R.; Cornejo, J.A. Targeted inhibition of allergen-induced histamine production by neutrophils. FASEB J. 2021, 35, 21483. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Traits | Muscle Condition (M) | p-Value | |
|---|---|---|---|
| NO | SM | ||
| L* | 45.46 ± 3.86 b | 47.85 ± 3.58 a | p = 0.02 |
| a* | 1.61 ± 0.70 | 1.87 ± 1.27 | p = 0.33 |
| b* | 7.93 ± 1.52 b | 9.08 ± 2.83 a | p = 0.04 |
| pH | 5.84 ± 0.33 a | 5.63 ± 0.17 b | p = 0.04 |
| Hydraulic system, n | 0.24 ± 0.05 | 0.25 ± 0.06 | p = 0.87 |
| Shear force, % | 27.48 ± 6.25 a | 18.42 ± 6.78 b | p = 0.00 |
| Moisture, % | 72.75 ± 0.56 | 72.48 ± 0.72 | p = 0.16 |
| Protein, % | 24.93 ± 0.54 a | 24.19 ± 0.44 b | p = 0.00 |
| Fat, % | 1.45 ± 0.43 | 1.82 ± 0.79 | p = 0.06 |
| Collagen 1 | 0.54 ± 0.10 | 0.59 ± 0.11 | p = 0.08 |
| Carcass weight, g | 1864 ± 213.20 | 1857 ± 209.70 | p = 0.98 |
| Unilateral breast, g | 138.41 ± 13.42 | 136.78 ± 12.95 | p = 0.96 |
| Metabolites | MZ | RT (min) | VIP | p | FC |
|---|---|---|---|---|---|
| Upregulated | |||||
| (S)-2-Methylmalate | 149.06 | 258.30 | 2.23 | 0.02 | 4.65 |
| Acetylcysteine | 163.00 | 408.50 | 2.74 | 0.00 | 6.99 |
| Dodecanoic acid | 199.98 | 989.60 | 2.20 | 0.02 | 1.80 |
| Capsidiol | 219.17 | 648.00 | 2.75 | 0.00 | 3.68 |
| 2-trans,6-trans-Farnesal | 221.19 | 918.40 | 2.35 | 0.00 | 1.84 |
| Ergothioneine | 230.10 | 108.10 | 1.92 | 0.03 | 2.48 |
| 13-OxoODE | 277.22 | 810.50 | 1.88 | 0.03 | 1.18 |
| Isotretinoin | 301.22 | 818.70 | 3.04 | 0.00 | 6.10 |
| 9-cis-Retinoic acid | 301.22 | 839.30 | 2.03 | 0.02 | 3.08 |
| Cafestol | 317.21 | 870.20 | 2.42 | 0.00 | 1.19 |
| 14,15-DiHETrE | 321.24 | 743.40 | 3.07 | 0.00 | 10.75 |
| Lithocholic acid | 377.31 | 797.60 | 2.12 | 0.01 | 9.49 |
| 1-Arachidonoylglycerol | 379.28 | 935.00 | 2.56 | 0.00 | 12.73 |
| L-Malic acid | 133.02 | 123.90 | 2.19 | 0.01 | 1.07 |
| L-Phenylalanine | 164.07 | 300.60 | 2.21 | 0.01 | 1.20 |
| 1-Methylhistidine | 168.08 | 107.30 | 2.01 | 0.03 | 1.21 |
| Yohimbine | 353.18 | 740.80 | 2.48 | 0.00 | 2.02 |
| methylnaphthaleneacetic acid | |||||
| Catechol | 111.02 | 909.00 | 1.98 | 0.03 | 0.77 |
| 4-Hydroxybenzaldehyde | 123.04 | 204.50 | 2.09 | 0.02 | 0.66 |
| 2-Phenylacetamide | 136.08 | 204.30 | 2.20 | 0.02 | 0.50 |
| 2,3-Butanediol | 154.99 | 59.90 | 2.06 | 0.02 | 0.61 |
| Glucosamine | 180.09 | 358.40 | 2.08 | 0.02 | 0.54 |
| 8-Methoxykynurenate | 220.06 | 490.50 | 1.90 | 0.04 | 0.82 |
| Pyrimidodiazepine | 221.09 | 424.80 | 2.39 | 0.01 | 0.48 |
| 9(S)-HPODE | 313.24 | 701.10 | 2.09 | 0.02 | 0.63 |
| 12(S)-HpETE | 319.23 | 867.70 | 2.05 | 0.02 | 0.63 |
| Rosmarinic acid | 360.24 | 462.70 | 2.07 | 0.02 | 0.71 |
| Vitamin D3 | 384.35 | 913.40 | 2.11 | 0.02 | 0.89 |
| beta-Carotene | 536.16 | 919.00 | 1.90 | 0.03 | 0.49 |
| Fumaric acid | 115.00 | 74.90 | 2.09 | 0.02 | 0.62 |
| Isopropylparaben | 179.07 | 741.80 | 2.23 | 0.01 | 0.87 |
| 4-Pyridoxic acid | 182.05 | 340.20 | 2.09 | 0.04 | 0.34 |
| 9,10-DHOME | 314.24 | 827.40 | 1.93 | 0.03 | 0.47 |
| Prostaglandin J2 | 315.19 | 682.90 | 2.19 | 0.01 | 0.40 |
| Uridine diphosphate glucose | 565.05 | 80.50 | 2.17 | 0.03 | 0.67 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wu, T.; Liu, P.; Wu, J.; Jiang, Y.; Zhou, N.; Zhang, Y.; Xu, Q.; Zhang, Y. Broiler Spaghetti Meat Abnormalities: Muscle Characteristics and Metabolomic Profiles. Animals 2024, 14, 1236. https://doi.org/10.3390/ani14081236
Wu T, Liu P, Wu J, Jiang Y, Zhou N, Zhang Y, Xu Q, Zhang Y. Broiler Spaghetti Meat Abnormalities: Muscle Characteristics and Metabolomic Profiles. Animals. 2024; 14(8):1236. https://doi.org/10.3390/ani14081236
Chicago/Turabian StyleWu, Teng, Pingping Liu, Jia Wu, Youluan Jiang, Ning Zhou, Yang Zhang, Qi Xu, and Yu Zhang. 2024. "Broiler Spaghetti Meat Abnormalities: Muscle Characteristics and Metabolomic Profiles" Animals 14, no. 8: 1236. https://doi.org/10.3390/ani14081236
APA StyleWu, T., Liu, P., Wu, J., Jiang, Y., Zhou, N., Zhang, Y., Xu, Q., & Zhang, Y. (2024). Broiler Spaghetti Meat Abnormalities: Muscle Characteristics and Metabolomic Profiles. Animals, 14(8), 1236. https://doi.org/10.3390/ani14081236