Effects of a Change from an Indoor-Based Total Mixed Ration to a Rotational Pasture System Combined With a Moderate Concentrate Feed Supply on Rumen Fermentation of Dairy Cows

 ,
, 
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Experimental Design and Treatments
2.2. Rumen pH and Fluid Composition
i/total number of observations per animal in week i)]/total number of animals in
week i
2.3. Rumen Content
2.4. Rumen Papillae Collection
2.5. VFA Absorption Test
2.6. Statistics
3. Results
3.1. Ration Composition and Weather Data
3.2. Animal Performance
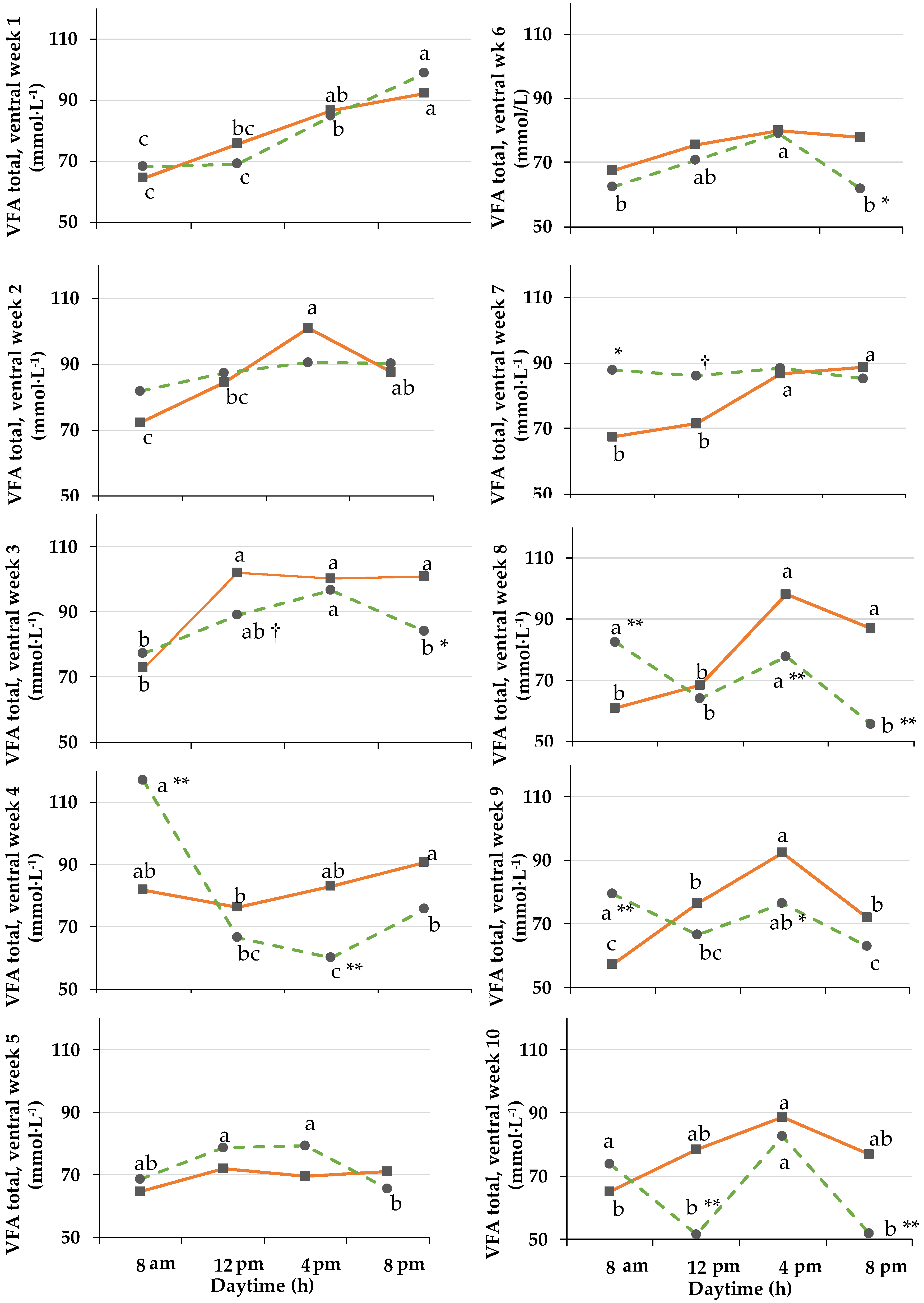
3.3. Rumen pH and Fluid Composition
3.3.1. pH-Sensor Data
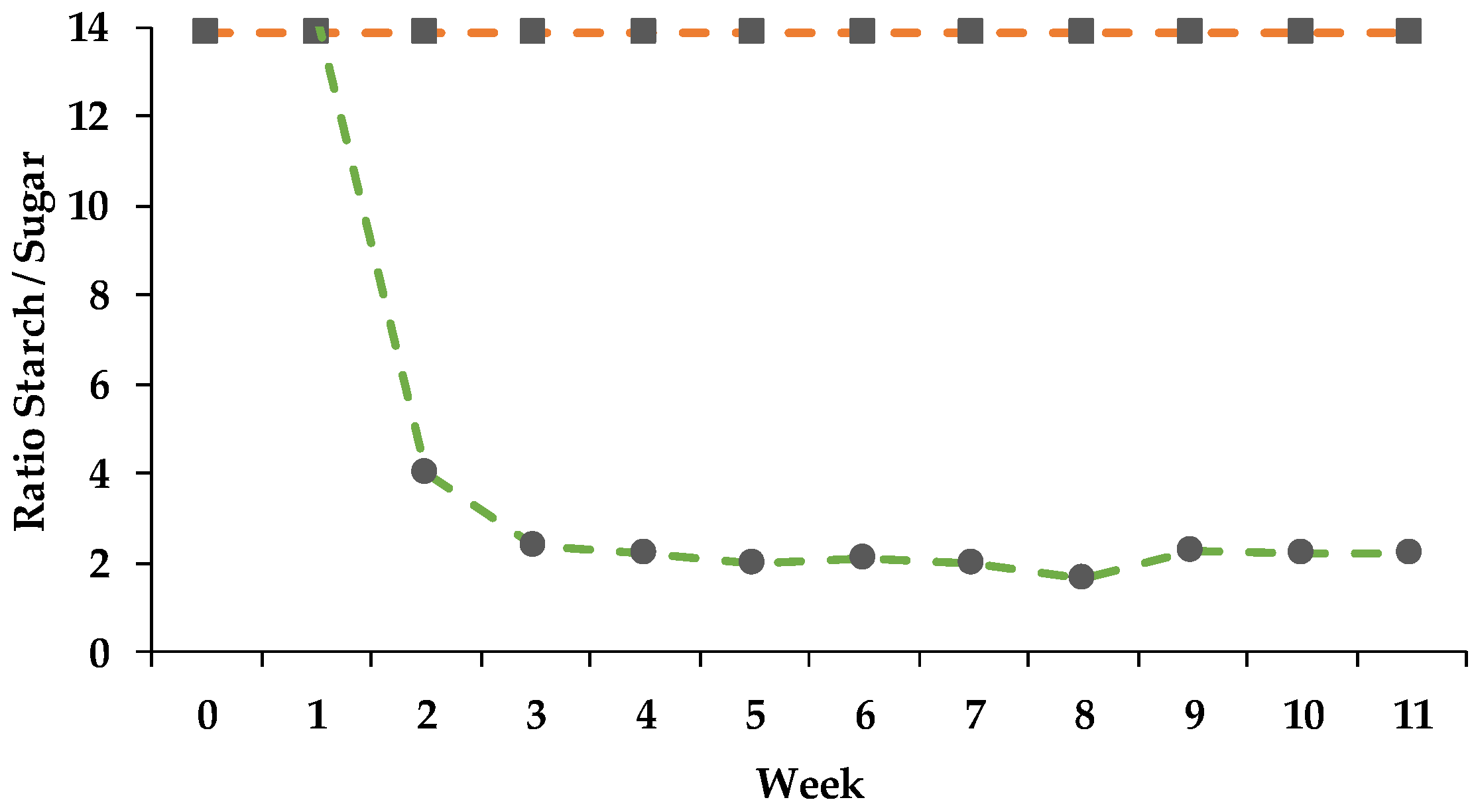
3.3.2. DMI of Starch and Sugar and its Influence on Rumen pH
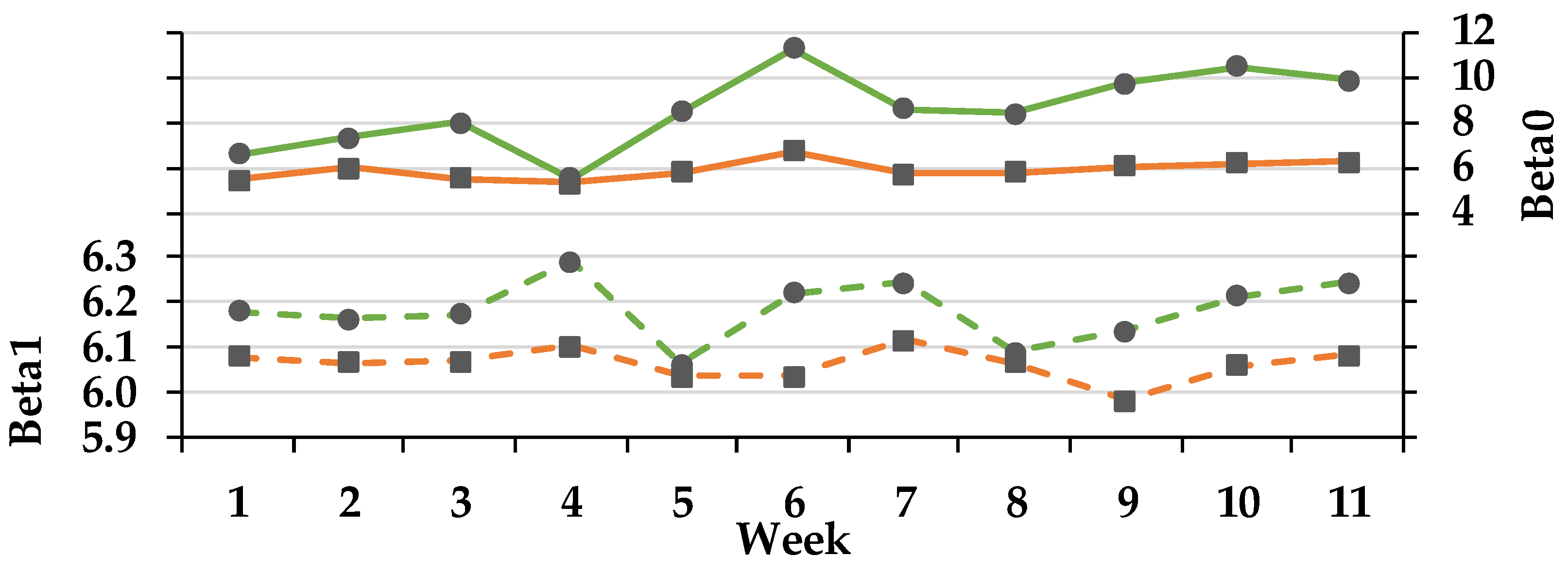
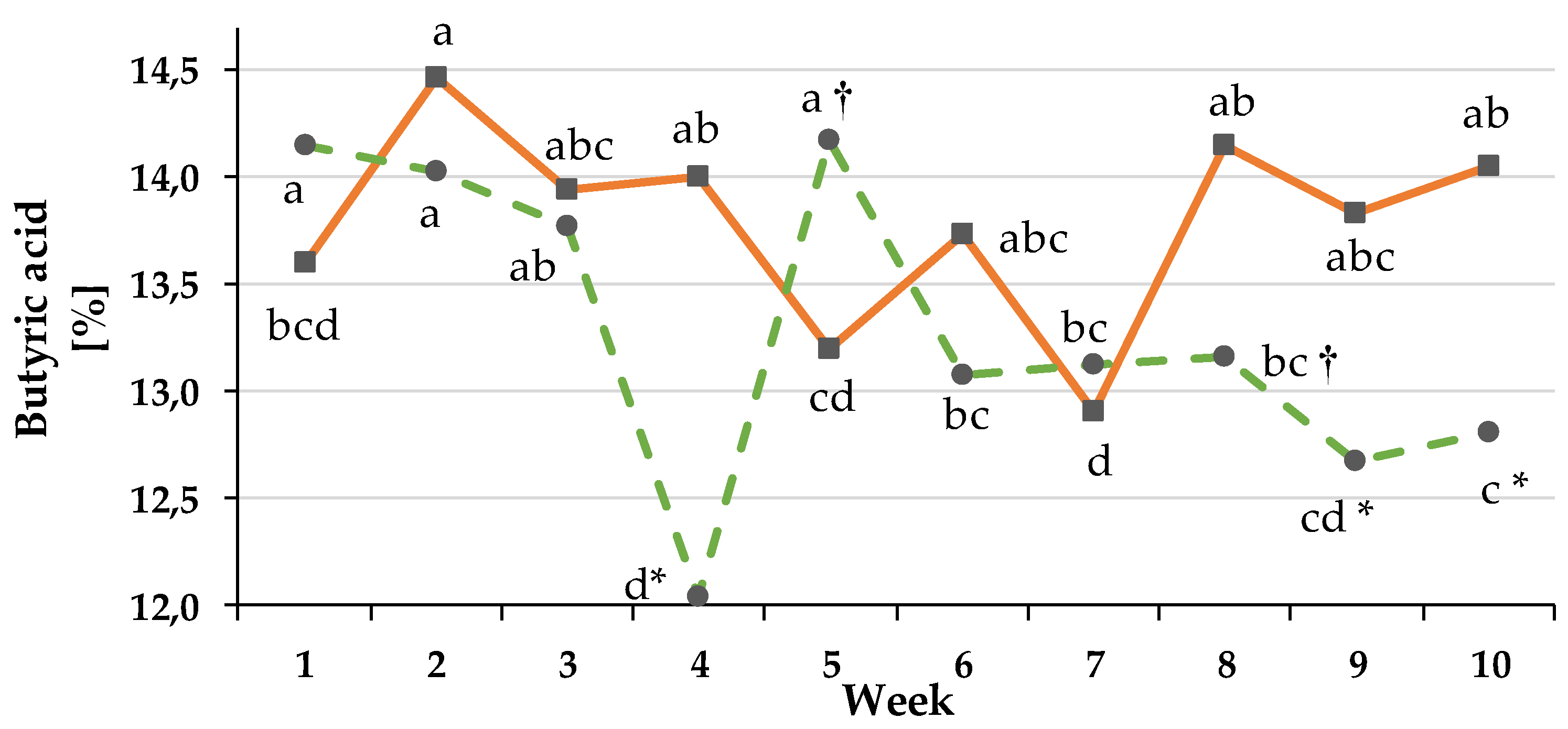
3.3.3. VFA Concentration and Molar Proportions
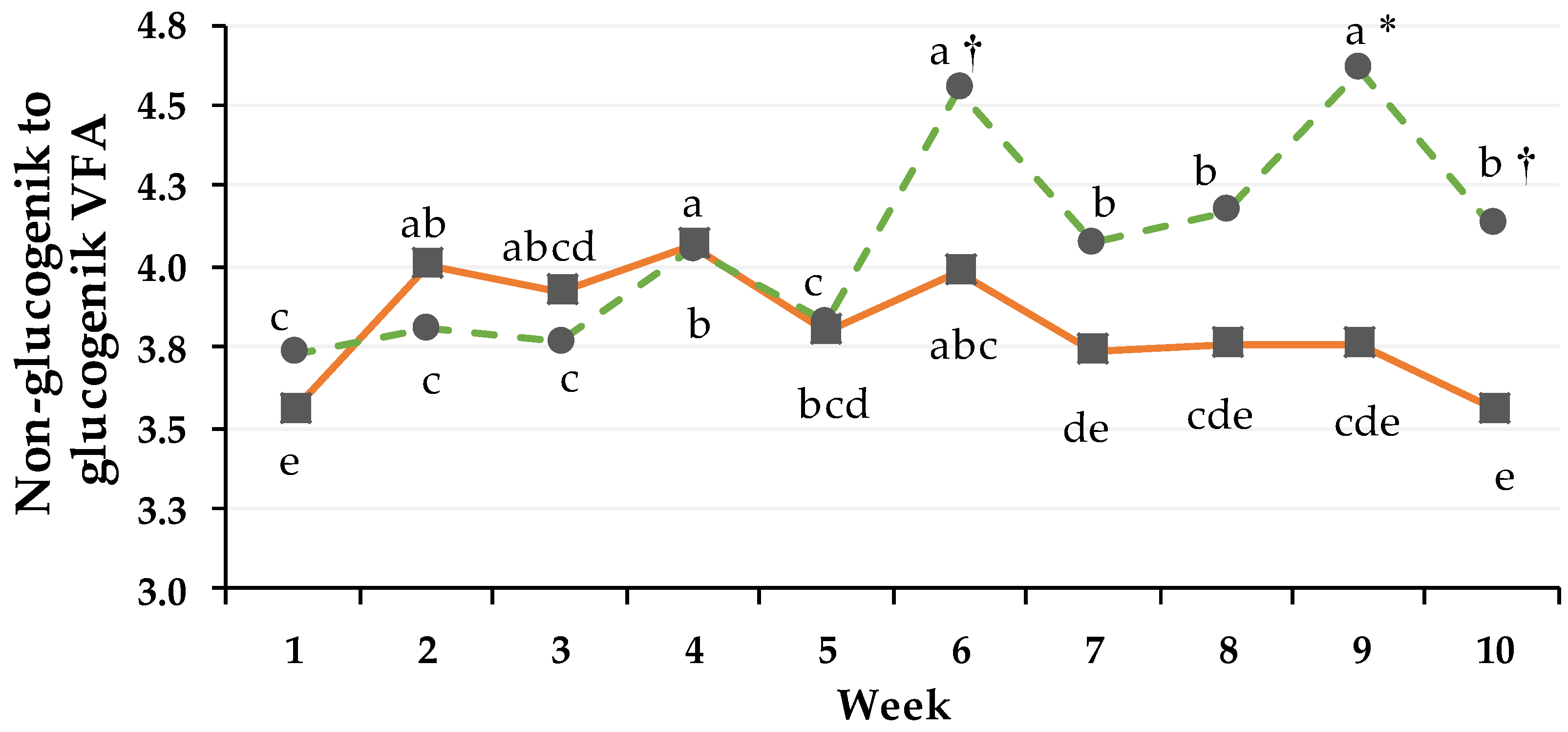
3.4. Non-Glucogenic to Glucogenic Ratio
3.5. Rumen Variables
3.5.1. Rumen Content
3.5.2. Rumen Papillae
3.5.3. Lipopolysaccharide Concentration in Rumen Liquid
3.6. VFA Absorption Test with Buffer Solution
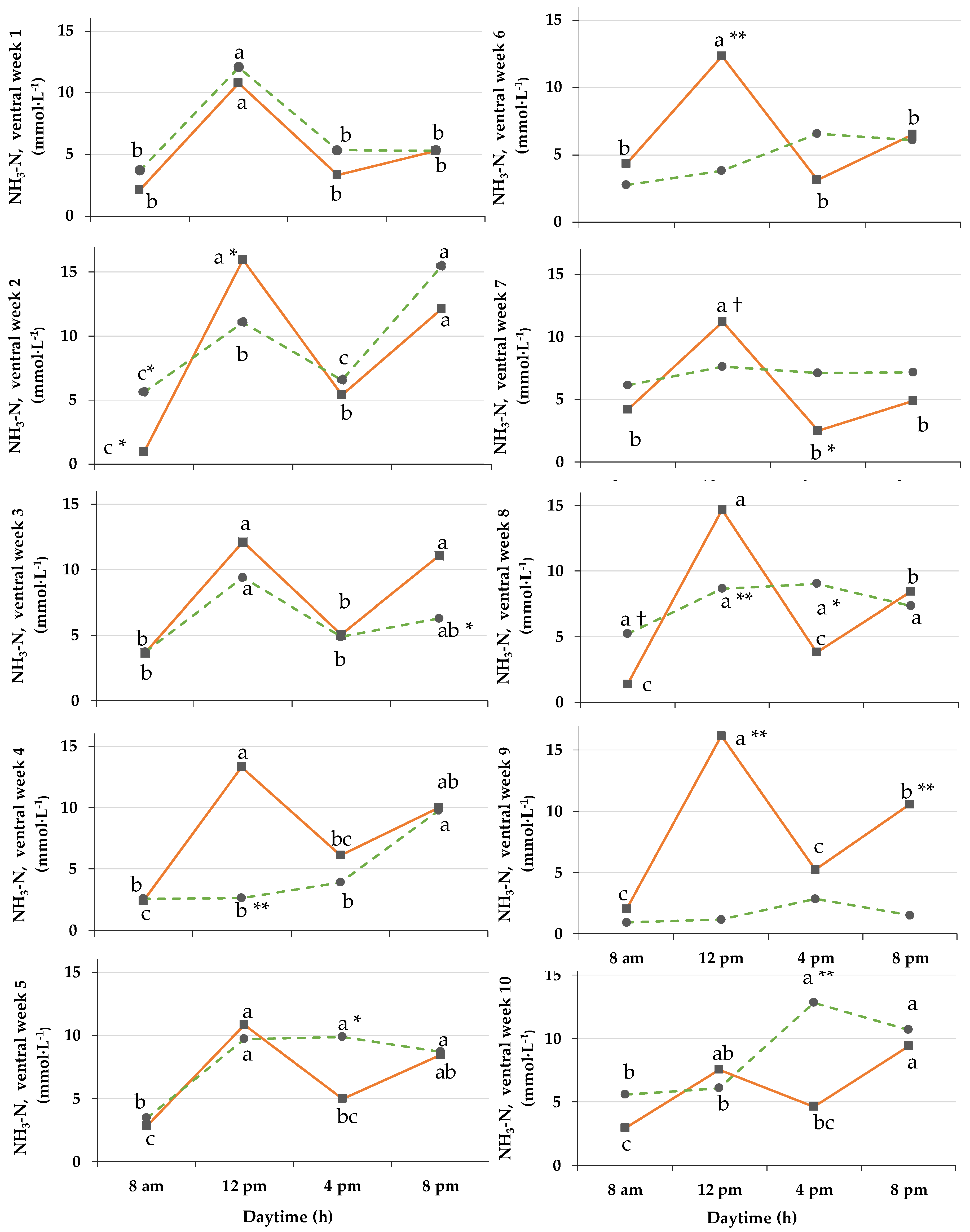
3.7. NH3-N Concentrations
4. Discussion
4.1. Changes of Rumen Papillae Surface Area
4.2. Weekly Pattern of pH during Transition and Fulltime Grazing
4.3. Development of Rumen Fill Grade, Consequences on VFA Production
4.4. Development of NH3-N Concentration in Rumen During Ration Change and Fulltime Grazing
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Appendix A






References
- Schären, M.; Jostmeier, S.; Ruesink, S.; Hüther, L.; Frahm, J.; Bulang, M.; Meyer, U.; Rehage, J.; Isselstein, J.; Breves, G.; et al. The effects of a ration change from a total mixed ration to pasture on health and production of dairy cows. J. Dairy Sci. 2016, 99, 1183–1200. [Google Scholar] [CrossRef] [PubMed]
- Schären, M.; Seyfang, G.M.; Steingass, H.; Dieho, K.; Dijkstra, J.; Hüther, L.; Frahm, J.; Beineke, A.; von Soosten, D.; Meyer, U.; et al. The effects of a ration change from a total mixed ration to pasture on rumen fermentation, volatile fatty acid absorption characteristics, and morphology of dairy cows. J. Dairy Sci. 2016, 99, 3549–3565. [Google Scholar] [CrossRef] [PubMed]
- Hartwiger, J.; Scharen, M.; Gerhards, U.; Huther, L.; Frahm, J.; von Soosten, D.; Kluss, J.; Bachmann, M.; Zeyner, A.; Meyer, U.; et al. Effects of a Change from an Indoor-Based Total Mixed Ration to a Rotational Pasture System Combined with a Moderate Concentrate Feed Supply on the Health and Performance of Dairy Cows. Animals 2018, 8. [Google Scholar] [CrossRef] [PubMed]
- Kennedy, E.; O’Donovan, M.; Murphy, J.P.; Delaby, L.; O’Mara, F. Effects of grass pasture and concentrate-based feeding systems for spring-calving dairy cows in early spring on performance during lactation. Grass Forage Sci. 2005, 60, 310–318. [Google Scholar] [CrossRef]
- Kolver, E.S.; Muller, L.D. Performance and Nutrient Intake of High Producing Holstein Cows Consuming Pasture or a Total Mixed Ration. J. Dairy Sci. 1998, 81, 1403–1411. [Google Scholar] [CrossRef]
- Dohme-Meier, F.; Kaufmann, L.D.; Görs, S.; Junghans, P.; Metges, C.C.; van Dorland, H.A.; Bruckmaier, R.M.; Münger, A. Comparison of energy expenditure, eating pattern and physical activity of grazing and zero-grazing dairy cows at different time points during lactation. Livest. Sci. 2014, 162, 86–96. [Google Scholar] [CrossRef]
- Kolver, E.S.; Roche, J.R.; de Veth, M.J.; Thorne, P.L.; Napper, A.R. Total mixed rations versus pasture diets: Evidence for a genotype x diet on the concentration of amino acid-derived volatiles in milk. Proc. NZSAP 2002, 62, 246–251. [Google Scholar]
- Schären, M.; Kiri, K.; Riede, S.; Gardener, M.; Meyer, U.; Hummel, J.; Urich, T.; Breves, G.; Dänicke, S. Alterations in the Rumen Liquid-, Particle- and Epithelium-Associated Microbiota of Dairy Cows during the Transition from a Silage- and Concentrate-Based Ration to Pasture in Spring. Front. Microbiol. 2017, 8, 744. [Google Scholar] [CrossRef] [PubMed]
- Bannink, A.; France, J.; Lopez, S.; Gerrits, W.J.J.; Kebreab, E.; Tamminga, S.; Dijkstra, J. Modelling the implications of feeding strategy on rumen fermentation and functioning of the rumen wall. Anim. Feed Sci. Technol. 2008, 143, 3–26. [Google Scholar] [CrossRef]
- O’Grady, L.; Doherty, M.L.; Mulligan, F.J. Subacute ruminal acidosis (SARA) in grazing Irish dairy cows. Vet. J. 2008, 176, 44–49. [Google Scholar] [CrossRef] [PubMed]
- Mentschel, J.; Leiser, R.; Mülling, C.; Pfarrer, C.; Claus, R. Butyric acid stimulates rumen mucosa development in the calf mainly by a reduction of apoptosis. Arch. Anim. Nutr. 2001, 55, 85–102. [Google Scholar] [CrossRef]
- Bargo, F.; Muller, L.D.; Delahoy, J.E.; Cassidy, T.W. Performance of high producing dairy cows with three different feeding systems combining pasture and total mixed rations. J. Dairy Sci. 2002, 85, 2948–2963. [Google Scholar] [CrossRef]
- Antonio Silva, J.; Miranda Silva, C.G.; de Paula Sousa, D.; de Paula, N.F.; da Silva Carvalho, A.P.; Gomes Macedo, B.; da Costa Júnior, W.S.; Bianchi-Zanette, I.E.; Viana Costa, R.; Weich Teobaldo, R. Supplementation strategies for dairy cows kept in tropical grass pastures. Semina: Ciências Agrárias 2017, 38, 401–416. [Google Scholar]
- VDLUFA. Verband Deutscher Landwirtschaftlicher Untersuchungs- und Forschungsanstalten. Handbuch der Landwirtschaftlichen Versuchs- und Untersuchungsmethodik (VDLUFA-Methodenbuch), Bd. III Die chemische Untersuchung von Futtermitteln; VDLUFA-Verlag: Darmstadt, Germany, 2012. (In German) [Google Scholar]
- Heublein, C.; Dohme-Meier, F.; Sudekum, K.H.; Bruckmaier, R.M.; Thanner, S.; Schori, F. Impact of cow strain and concentrate supplementation on grazing behaviour, milk yield and metabolic state of dairy cows in an organic pasture-based feeding system. Animal 2017, 11, 1163–1173. [Google Scholar] [CrossRef] [PubMed]
- Geissler, C.; Hoffmann, M.; Hiokel, B. Ein Beitrag zur gaschromatographischen Bestimmung flüchtiger Fettsäuren. Archiv. Anim. Nutr. 1976, 26, 339–344. [Google Scholar] [CrossRef]
- Anonymous. Deutsche Einheitsverfahren zur Wasser-, Abwasser und Schlammuntersuchung. 40. Lieferung; DIN 38406-E5-2; Beuth and Wiley-VCH: Berlin, Germany, 1998; Volume 6. [Google Scholar]
- Abrahamse, P.A.; Dijkstra, J.; Vlaeminck, B.; Tamminga, S. Frequent Allocation of Rotationally Grazed Dairy Cows Changes Grazing Behavior and Improves Productivity. J. Dairy Sci. 2008, 91, 2033–2045. [Google Scholar] [CrossRef] [PubMed]
- Penner, G.B.; Beauchemin, K.A.; Mutsvangwa, T. An Evaluation of the Accuracy and Precision of a Stand-Alone Submersible Continuous Ruminal pH Measurement System. J. Dairy Sci. 2006, 89, 2132–2140. [Google Scholar] [CrossRef]
- Zebeli, Q.; Dijkstra, J.; Tafaj, M.; Steingass, H.; Ametaj, B.N.; Drochner, W. Modeling the adequacy of dietary fiber in dairy cows based on the responses of ruminal pH and milk fat production to composition of the diet. J. Dairy Sci. 2008, 91, 2046–2066. [Google Scholar] [CrossRef] [PubMed]
- Gorniak, T.; Meyer, U.; Sudekum, K.H.; Danicke, S. Impact of mild heat stress on dry matter intake, milk yield and milk composition in mid-lactation Holstein dairy cows in a temperate climate. Arch. Anim. Nutr. 2014, 68, 358–369. [Google Scholar] [CrossRef] [PubMed]
- Kellaway, R.; Colditz, P. The effect of heat stress on growth and nitrogen metabolism in Friesian and F1 Brahman × Friesian heifers. Aust. J. Agric. Res. 1975, 26, 615–622. [Google Scholar] [CrossRef]
- Collier, R.; Zimbelman, R.; Rhoads, R.; Rhoads, M.; Baumgard, L. A re-evaluation of the impact of temperature humidity index (THI) and black globe humidity index (BGHI) on milk production in high producing dairy cows. In Proceedings of the Western Dairy Management Conference, Reno, NV, USA, 11–13 March 2009; pp. 113–125. [Google Scholar]
- Abrahamse, P.A.; Tamminga, S.; Dijkstra, J. Effect of daily movement of dairy cattle to fresh grass in morning or afternoon on intake, grazing behaviour, rumen fermentation and milk production. J. Agric. Sci. 2009, 147, 721–730. [Google Scholar] [CrossRef]
- Gäbel, G.; Aschenbach, J.R.; Müller, F. Transfer of energy substrates across the ruminal epithelium: implications and limitations. Anim. Health Res. Rev. 2007, 3, 15–30. [Google Scholar] [CrossRef]
- Dieho, K.; Bannink, A.; Geurts, I.A.; Schonewille, J.T.; Gort, G.; Dijkstra, J. Morphological adaptation of rumen papillae during the dry period and early lactation as affected by rate of increase of concentrate allowance. J. Dairy Sci. 2016, 99, 2339–2352. [Google Scholar] [CrossRef] [PubMed]
- Ma, L.; Zhao, M.; Xu, J.; Zhao, L.; Bu, D. 1643 Effects of dietary neutral detergent fiber and starch ratio on rumen epithelial cell morphological structure and gene expression in dairy cows. J. Anim. Sci. 2016, 94, 800–801. [Google Scholar] [CrossRef] [Green Version]
- Wang, B.; Wang, D.; Wu, X.; Cai, J.; Liu, M.; Huang, X.; Wu, J.; Liu, J.; Guan, L. Effects of dietary physical or nutritional factors on morphology of rumen papillae and transcriptome changes in lactating dairy cows based on three different forage-based diets. BMC Genom. 2017, 18, 353. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bramley, E.; Lean, I.J.; Fulkerson, W.J.; Stevenson, M.A.; Rabiee, A.R.; Costa, N.D. The definition of acidosis in dairy herds predominantly fed on pasture and concentrates. J. Dairy Sci. 2008, 91, 308–321. [Google Scholar] [CrossRef] [PubMed]
- Oetzel, G.R. Subacute ruminal acidosis in dairy herds: physiology, pathophysiology, milk fat responses, and nutritional management. In Proceedings of the 40th Annual Conference, American Association of Bovine Practitioners, Vancouver, BC, Canada, 20–22 September 2007; pp. 89–119. [Google Scholar]
- Filho, M.H.N.R.; Peyraud, J.L.; Delagarde, R. Foraging behavior and ruminal fermentation of dairy cows grazing ryegrass pasture alone or with white clover. Pesqui. Agropecu Bras. 2012, 47, 458–465. [Google Scholar] [CrossRef] [Green Version]
- Grant, R.J.; Colenbrander, V.F.; Albright, J.L. Effect of Particle Size of Forage and Rumen Cannulation upon Chewing Activity and Laterality in Dairy Cows1. J. Dairy Sci. 1990, 73, 3158–3164. [Google Scholar] [CrossRef]
- Melo, L.Q.; Costa, S.F.; Lopes, F.; Guerreiro, M.C.; Armentano, L.E.; Pereira, M.N. Rumen morphometrics and the effect of digesta pH and volume on volatile fatty acid absorption1. J. Anim. Sci. 2013, 91, 1775–1783. [Google Scholar] [CrossRef] [PubMed]
- Ueda, K.; Mitani, T.; Kondo, S. Effect of water-soluble carbohydrate content in orchardgrass pasture on grazing time and rumen fermentation in dairy cows. Anim. Sci. J. 2016, 87, 1122–1129. [Google Scholar] [CrossRef] [PubMed]
- Gibb, M.J. Chapter 8: Grassland management with emphasis on grazing behaviour. In Fresh Herbage for Dairy Cattle; Dordrecht, T.N., Ed.; Springer: Berlin, Germany, 2007; pp. 141–157. [Google Scholar]
- Taweel, H.Z.; Tas, B.M.; Smit, H.J.; Elgersma, A.; Dijkstra, J.; Tamminga, S. Grazing behaviour, intake, rumen function and milk production of dairy cows offered Lolium perenne containing different levels of water-soluble carbohydrates. Livest. Sci. 2006, 102, 33–41. [Google Scholar] [CrossRef]
- Leng, R.; Steel, J.; Luick, J. Contribution of propionate to glucose synthesis in sheep. Biochem. J. 1967, 103, 785. [Google Scholar] [CrossRef] [PubMed]
- De Menezes, A.B.; Lewis, E.; O’Donovan, M.; O’Neill, B.F.; Clipson, N.; Doyle, E.M. Microbiome analysis of dairy cows fed pasture or total mixed ration diets. FEMS Microbiol. Ecol. 2011, 78, 256–265. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baumgard, L.H.; Rhoads, R.P. Ruminant Nutrition Symposium: Ruminant Production and Metabolic Responses to Heat Stress12. J. Anim. Sci. 2012, 90, 1855–1865. [Google Scholar] [CrossRef] [PubMed]
- Taweel, H.Z.; Tas, B.M.; Dijkstra, J.; Tamminga, S. Intake regulation and grazing behavior of dairy cows under continuous stocking. J. Dairy Sci. 2004, 87, 3417–3427. [Google Scholar] [CrossRef]
- Carruthers, V.R.; Neil, P.G. Milk production and ruminal metabolites from cows offered two pasture diets supplemented with non-structural carbohydrate. N. Z. J. Agric. Res. 1997, 40, 513–521. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Type of Feeding | Item (g/kg of DM, unless otherwise noted) | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| DM (g·kg−1) | Ash | CP | uCP | NEL* | Sugar | Starch | RNB | CF | NDFOM | ADFOM | EE | |
| TMR CG/PG 1 | 471 ± 6 | 66 ± 0 | 158 ± 1 | 152 ± 0.5 | 6.8 ± 0 | 18 ± 0 | 253 ± 2.5 | 1.1 ± 0.3 | 184 ± 0 | 363 ± 0 | 205 ± 1.5 | 37 ± 0.5 |
| Pasture PG | 174 ± 23 | 94 ± 9 | 188 ± 19 | 142 ± 7 | 6.2 ± 0.3 | 114 ± 45 | -- | 6.2 ± 1.9 | 231 ± 34 | 471 ± 40 | 254 ± 37 | 41 ± 5.8 |
| Concentrate PG | 899 | 108 | 93 | 148 | 7.6 | 27 | 579 | 8.9 | 31.5 | 124 | 42 | 30 |
| Variable 1 | Group 2 | Week | p-Value | ||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 0 | 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 11 | PSEM 3 | G | T | G × T | ||
| DMI (kg·d−1) 4 | CG | 19.1 | 19.9 | 21.4 | 20.3 | 20.8 | 21.5 | 22.5 | 21.9 | 21.2 | 21.8 | 22.8 | 20.4 | 1.2 | 0.079 | 0.05 | <0.01 |
| DMI (kg·d−1) 5 | PG | 20.7 | 21.2 | 21.5 | 19.8 | 15.6 | 17.7 | 18.9 | 18.8 | 18.5 | 16.3 | 17.5 | 17.2 | ||||
| Milk yield | CG | 29.3 | 28.3 | 27.5 | 28.8 | 28.9 | 28.9 | 28.6 | 28.1 | 28.4 | 28.0 | 27.3 | 27.2 | 2.7 | 0.523 | <0.01 | <0.01 |
| (kg·d−1) | PG | 29.1 | 29.4 | 29.8 | 29.4 | 26.0 | 27.1 | 25.5 | 25.1 | 24.7 | 22.6 | 23.8 | 22.8 | ||||
| Milk fat | CG | 4.3 | 4.5 | 4.8 | 4.3 | 4.4 | 4.2 | 4.4 | 4.4 | 4.1 | 4.3 | 4.5 | 4.2 | 0.25 | 0.170 | 0.533 | 0.141 |
| content (%) | PG | 3.6 | 4.1 | 3.5 | 3.9 | 4.4 | 4.0 | 4.0 | 4.1 | 4.0 | 3.9 | 4.1 | 4.0 | ||||
| Milk protein | CG | 3.2 | 3.3 | 3.0 | 3.2 | 3.2 | 3.1 | 3.1 | 3.2 | 3.3 | 3.2 | 3.3 | 3.4 | 0.1 | 0.635 | <0.01 | 0.075 |
| content (%) | PG | 3.2 | 3.2 | 3.3 | 3.2 | 3.0 | 3.1 | 3.0 | 3.1 | 3.2 | 3.1 | 3.2 | 3.3 | ||||
| Milk urea | CG | 129 | 139 | 142 | 183 | 211 | 166 | 166 | 177 | 179 | 173 | 180 | 185 | 17 | 0.181 | <0.001 | 0.098 |
| ppm | PG | 160 | 159 | 174 | 210 | 218 | 195 | 214 | 202 | 198 | 170 | 258 | 168 | ||||
| Body weight | CG | 662 | 663 | 658 | 656 | 651 | 660 | 660 | 670 | 670 | 681 | 693 | 695 | 40 | 0.907 | <0.001 | <0.001 |
| (kg) | PG | 685 | 683 | 670 | 664 | 651 | 651 | 640 | 646 | 645 | 658 6 | 670 | 675 | ||||
| Body condition | CG | 2.9 | 2.9 | 3.0 | 3.0 | 3.1 | 3.1 | 3.0 | 3.0 | 3.1 | 3.1 | 3.1 | 2.9 | 0.2 | 0.955 | 0.069 | 0.878 |
| score (scale 1-5) | PG | 2.9 | 3.0 | 3.0 | 3.0 | 3.2 | 3.1 | 3.0 | 2.9 | 3.0 | 3.0 | 3.1 | 3.0 | ||||
| Blood glucose | CG | 59.4 | 63.3 | 64.6 | 58.3 | 63.1 | 63.6 | 59.1 | 51.7 | 61.1 | 62.2 | 47.6 | 54.5 | 2.8 | 0.143 | <0.001 | <0.01 |
| (mg·dL−1) | PG | 56.7 | 60.3 | 60.4 | 56.5 | 56.3 † | 57.6 | 48.8 * | 55.3 | 58.6 | 52.7 * | 58.3 † | 46.9 * | ||||
| Week | p-Value | |||||||
|---|---|---|---|---|---|---|---|---|
| Variable | Group 1 | 0 | 6 | 10 | PSEM 2 | Group | Time | G × T |
| Buffer solution pH 60 min | CG | 7.01 | 6.91 | 7.07 | 0.02 | 0.753 | 0.105 | 0.241 |
| PG | 6.9 | 6.96 | 7.01 | |||||
| Buffer solution C2 60 min (mmol·l−1) | CG | 35.3 | 41.8 | 41.9 | 2.2 | 0.374 | <0.001 | 0.547 |
| PG | 30.8 | 39.4 | 44.4 | |||||
| Buffer solution C3 60 min (mmol·l−1) | CG | 11.9 | 14.2 | 14.5 | 0.91 | 0.273 | <0.001 | 0.639 |
| PG | 10.3 | 13.2 | 14.8 | |||||
| Buffer solution C4 60 min (mmol·l−1) | CG | 5.4 | 6.7 | 7.1 | 0.56 | 0.446 | <0.001 | 0.866 |
| PG | 4.9 | 6.3 | 6.9 | |||||
| Fractional absorption rate C2·h−1 | CG | 0.35 | 0.33 | 0.37 | 0.04 | 0.718 | 0.417 | 0.380 |
| PG | 0.44 | 0.34 | 0.32 | |||||
| Fractional absorption rate C3·h−1 | CG | 0.56 | 0.52 | 0.56 | 0.05 | 0.478 | 0.425 | 0.324 |
| PG | 0.64 | 0.54 | 0.54 | |||||
| Fractional absorption rate C4·h−1 | CG | 0.8 | 0.75 | 0.75 | 0.06 | 0.407 | 0.404 | 0.868 |
| PG | 0.88 | 0.76 | 0.79 | |||||
| Influx of water (L·h−1) | CG | 7.3 | 8.6 | 10.3 | 0.98 | 0.57 | 0.013 | 0.247 |
| PG | 6.9 | 11.0 | 9.6 | |||||
| Fractional Liquid passage rate·h−1 | CG | 0.33 | 0.45 | 0.41 | 0.07 | 0.018 | 0.054 | 0.742 |
| PG | 0.12 | 0.34 | 0.29 | |||||
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hartwiger, J.; Schären, M.; Potthoff, S.; Hüther, L.; Kersten, S.; Von Soosten, D.; Beineke, A.; Meyer, U.; Breves, G.; Dänicke, S. Effects of a Change from an Indoor-Based Total Mixed Ration to a Rotational Pasture System Combined With a Moderate Concentrate Feed Supply on Rumen Fermentation of Dairy Cows. Animals 2018, 8, 205. https://doi.org/10.3390/ani8110205
Hartwiger J, Schären M, Potthoff S, Hüther L, Kersten S, Von Soosten D, Beineke A, Meyer U, Breves G, Dänicke S. Effects of a Change from an Indoor-Based Total Mixed Ration to a Rotational Pasture System Combined With a Moderate Concentrate Feed Supply on Rumen Fermentation of Dairy Cows. Animals. 2018; 8(11):205. https://doi.org/10.3390/ani8110205
Chicago/Turabian StyleHartwiger, Julia, Melanie Schären, Sarah Potthoff, Liane Hüther, Susanne Kersten, Dirk Von Soosten, Andreas Beineke, Ulrich Meyer, Gerhard Breves, and Sven Dänicke. 2018. "Effects of a Change from an Indoor-Based Total Mixed Ration to a Rotational Pasture System Combined With a Moderate Concentrate Feed Supply on Rumen Fermentation of Dairy Cows" Animals 8, no. 11: 205. https://doi.org/10.3390/ani8110205
APA StyleHartwiger, J., Schären, M., Potthoff, S., Hüther, L., Kersten, S., Von Soosten, D., Beineke, A., Meyer, U., Breves, G., & Dänicke, S. (2018). Effects of a Change from an Indoor-Based Total Mixed Ration to a Rotational Pasture System Combined With a Moderate Concentrate Feed Supply on Rumen Fermentation of Dairy Cows. Animals, 8(11), 205. https://doi.org/10.3390/ani8110205