Intravenous Infusions of Glycerol Versus Propylene Glycol for the Regulation of Negative Energy Balance in Sheep: A Randomized Trial



Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animal Studies and Experimental Design
2.2. Plasma Biochemical Analysis
2.3. Statistical Analysis
3. Results
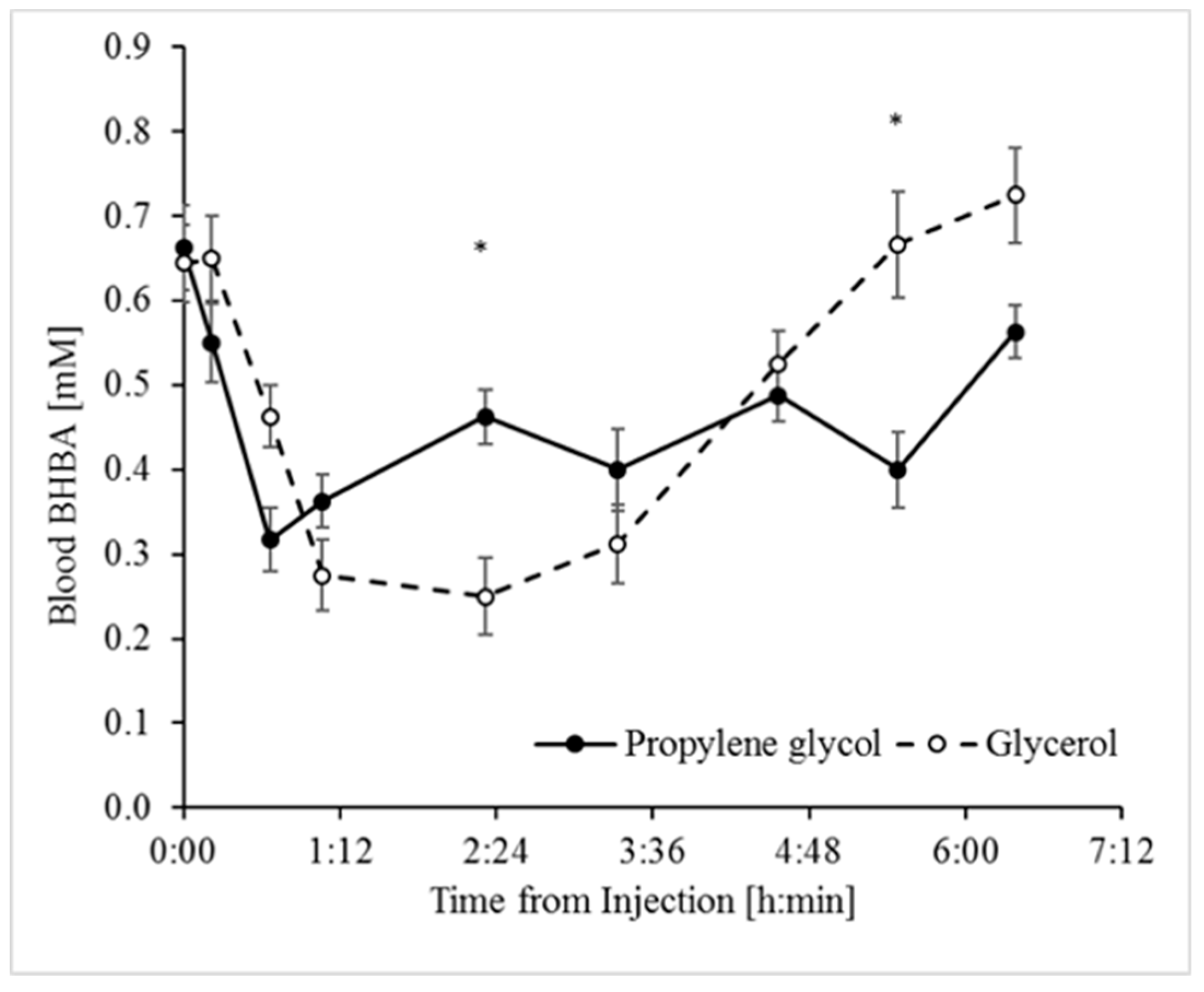
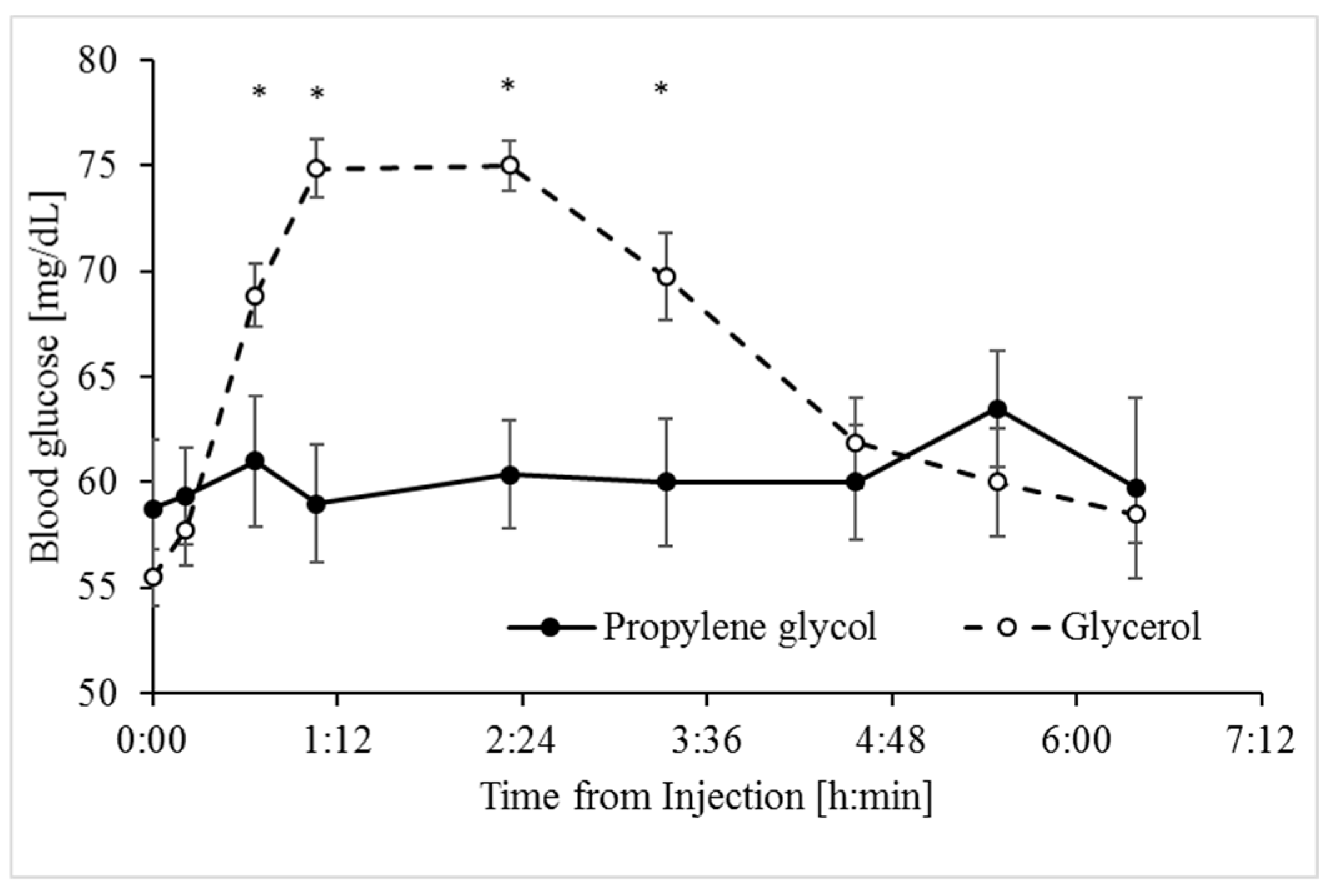
3.1. Glycerol and Propylene Glycol Were Both Anti-Ketogenic, but Only Glycerol was Glucogenic
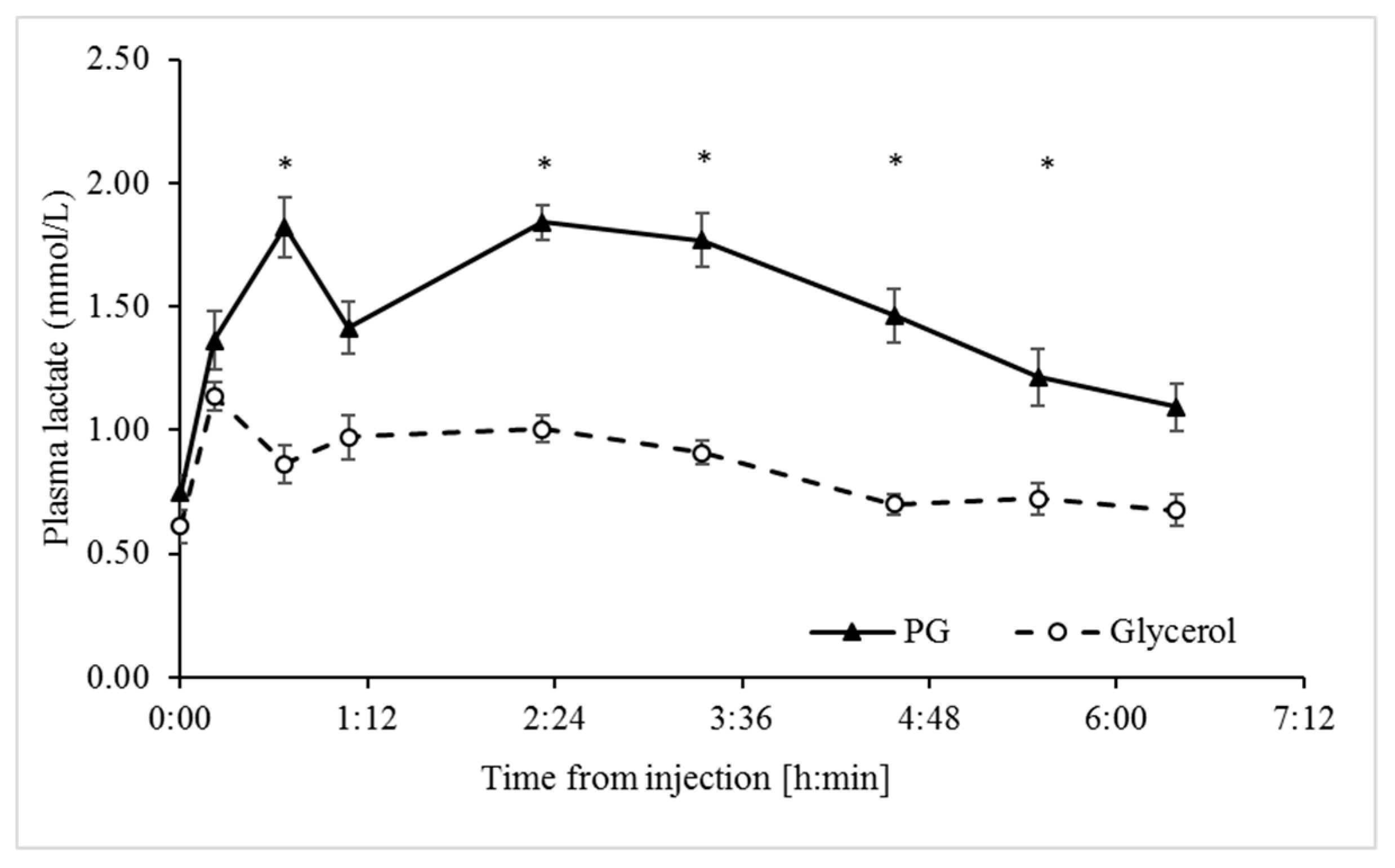
3.2. Unlike Glycerol, Propylene Glycol Increased Circulating Lactate Concentrations
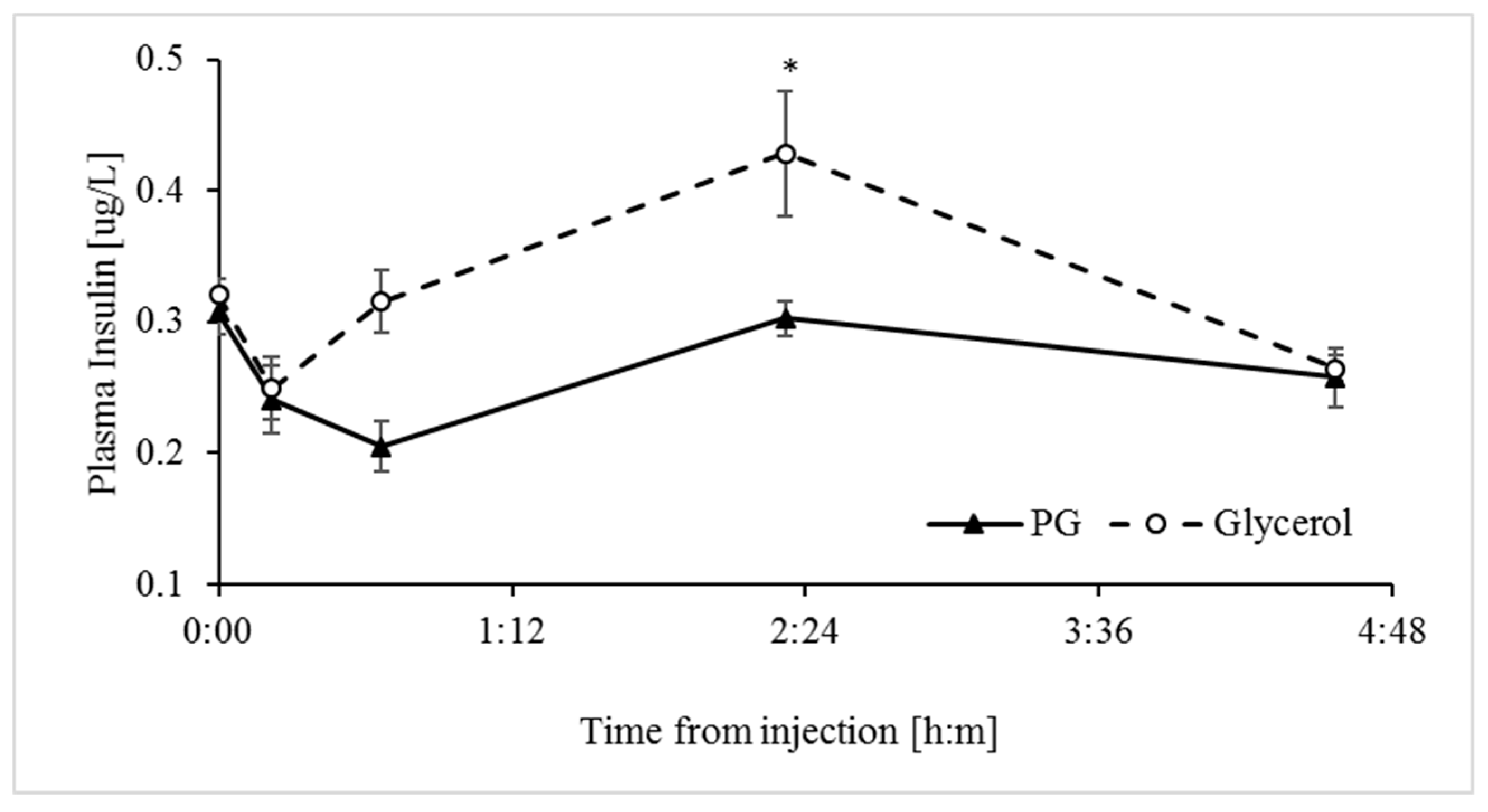
3.3. Glycerol Stimulates Greater Release of Insulin into Plasma than Propylene Glycol
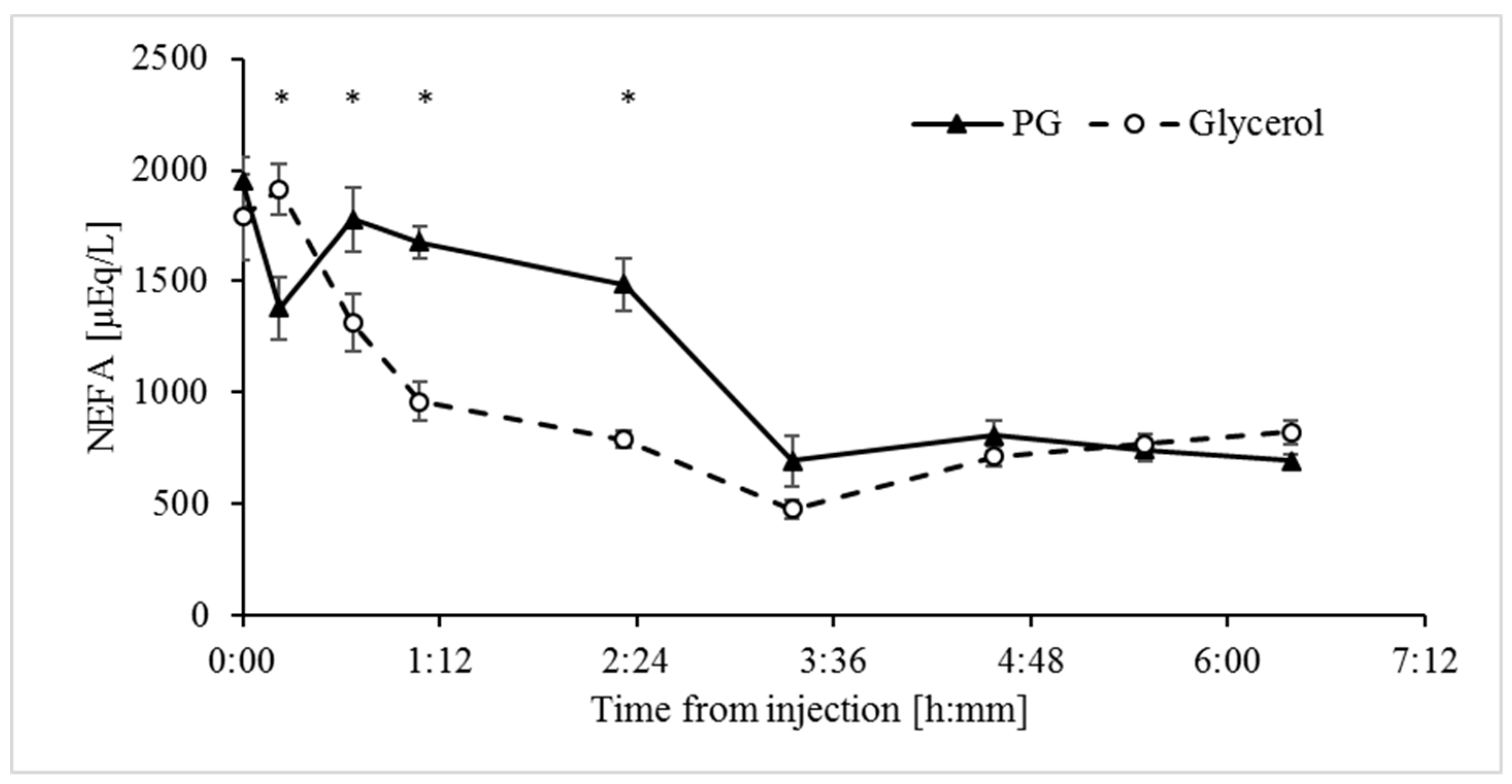
3.4. Both Glycerol and Propylene Glycol are Anti-Lipolytic, but the Response to Glycerol is Faster
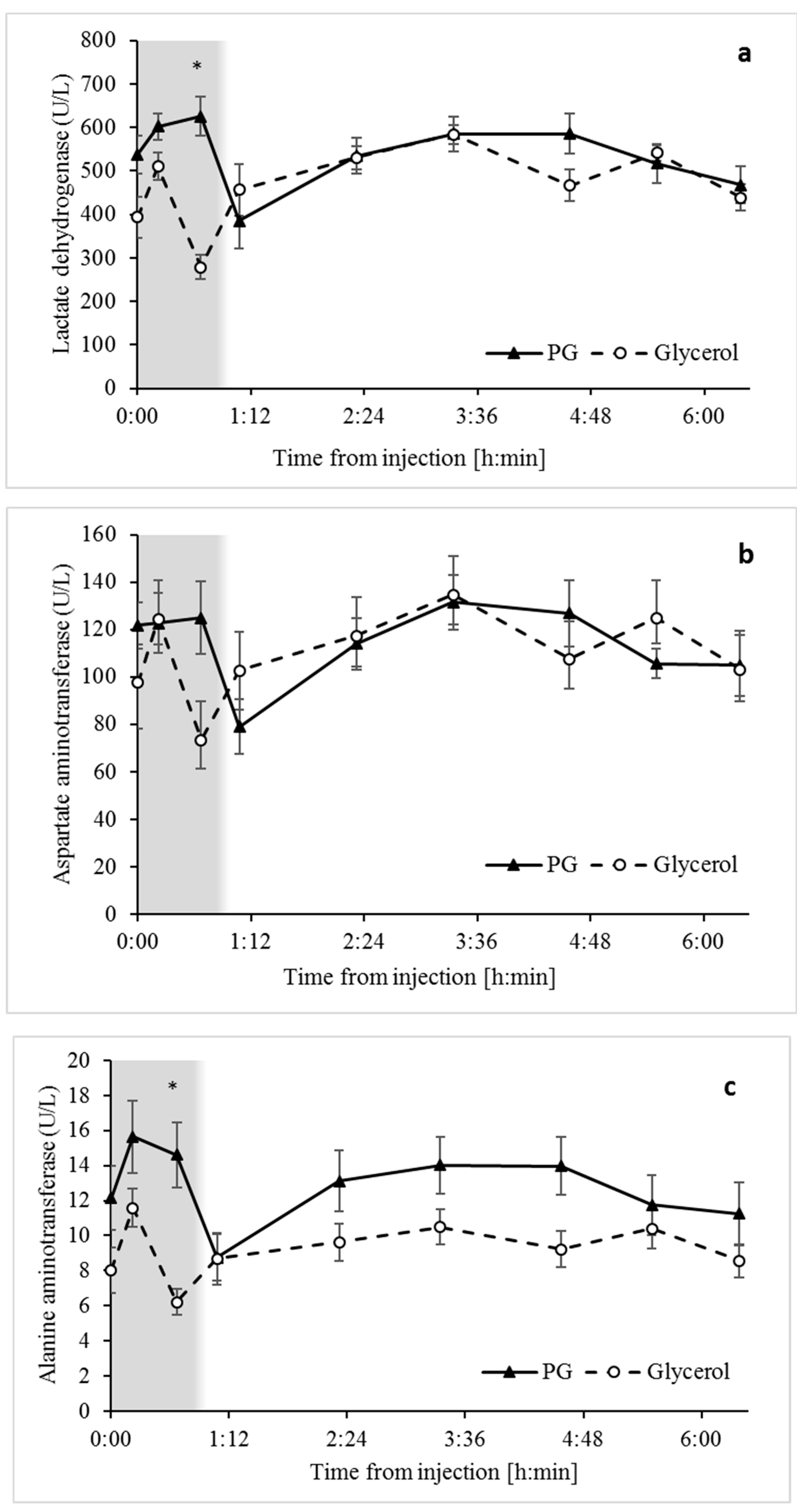
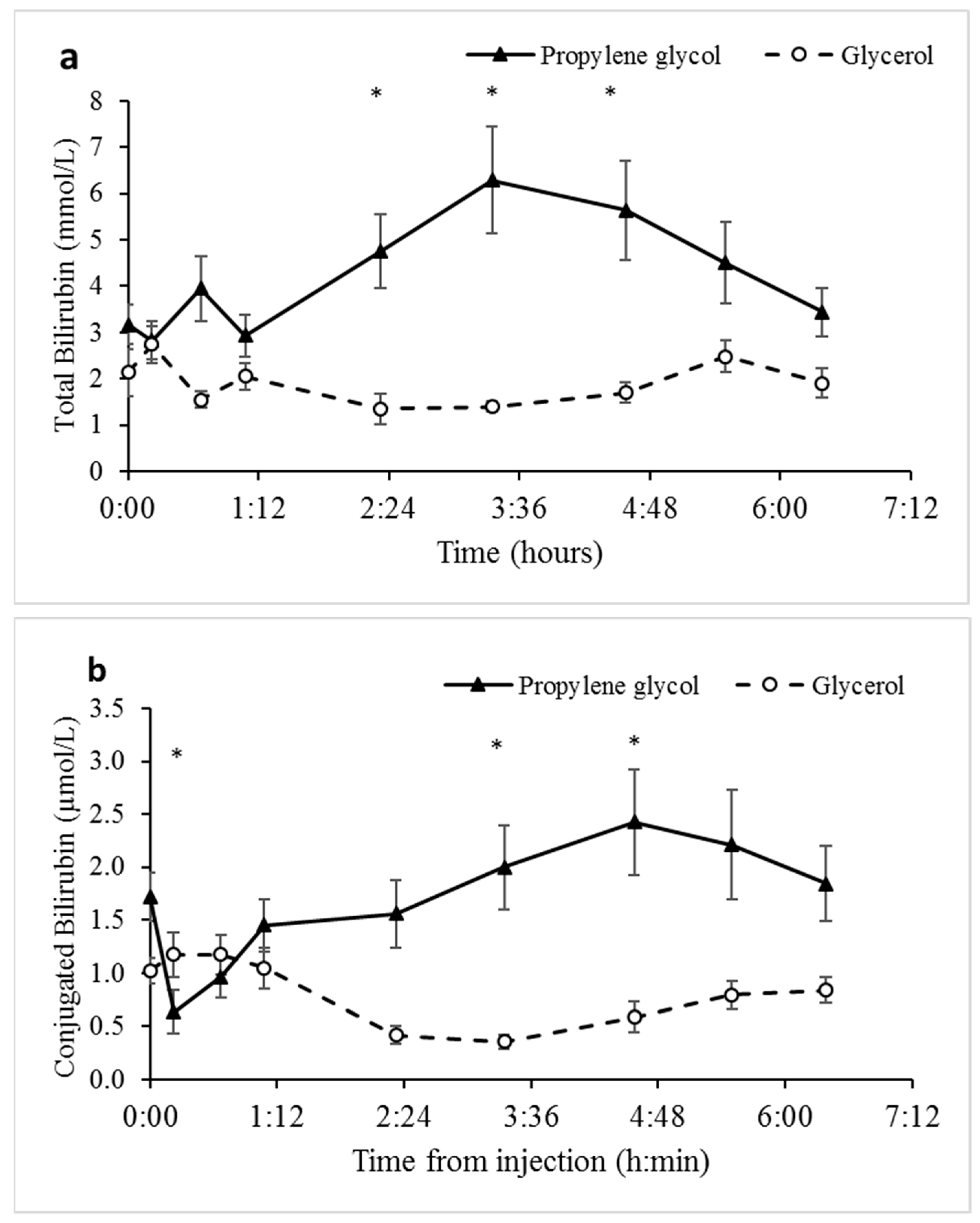
3.5. Biomarkers of Liver Function Indicate that Unlike Glycerol, Propylene Glycol may Induce Tissue Damage and Hemolysis
4. Discussion
5. Conclusions
6. Patent
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- McCue, M.D. Starvation physiology: Reviewing the different strategies animals use to survive a common challenge. Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 2010, 156, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Bauman, D.E.; Bruce Currie, W. Partitioning of Nutrients During Pregnancy and Lactation: A Review of Mechanisms Involving Homeostasis and Homeorhesis. J. Dairy Sci. 1980, 63, 1514–1529. [Google Scholar] [CrossRef]
- Rook, J.S. Pregnancy toxemia of ewes, does, and beef cows. Vet. Clin. North Am. Food Anim. Pract. 2000, 16, 293–317. [Google Scholar] [CrossRef]
- Rukkwamsuk, T.; Geelen, M.J.H.; Kruip, T.A.M.; Wensing, T. Interrelation of Fatty Acid Composition in Adipose Tissue, Serum, and Liver of Dairy Cows During the Development of Fatty Liver Postpartum. J. Dairy Sci. 2000, 83, 52–59. [Google Scholar] [CrossRef]
- Moallem, U.; Rozov, A.; Gootwine, E.; Honig, H. Plasma concentrations of key metabolites and insulin in late-pregnant ewes carrying 1 to 5 fetuses. J. Anim. Sci. 2012, 90, 318–324. [Google Scholar] [CrossRef] [PubMed]
- Zamir, S.; Rozov, A.; Gootwine, E. Treatment of pregnancy toxaemia in sheep with flunixin meglumine. Vet. Rec. 2009, 165, 265–266. [Google Scholar] [CrossRef] [PubMed]
- Schulz, L.M.; Riese, R.L. Pregnancy Toxemia in the Ewe. Iowa State Univ. Vet. 1983, 45, 11–15. [Google Scholar]
- Banchero, G.E.; Milton, J.T.; Lindsay, D.R.; Martin, G.B.; Quintans, G. Colostrum production in ewes: A review of regulation mechanisms and of energy supply. Animal 2015, 9, 831–837. [Google Scholar] [CrossRef]
- Moallem, U.; Rosov, A.; Honig, H.; Ofir, I.; Livshits, L.; Gootwine, E. Molasses-based supplement improved the metabolic status of late-pregnant ewes bearing multiple fetuses. Anim. Feed Sci. Technol. 2016, 219, 83–93. [Google Scholar] [CrossRef]
- Bowman, J.G.P.; Sowell, B.F.; Paterson, J.A. Liquid supplementation for ruminants fed low-quality forage diets: A review. Anim. Feed Sci. Technol. 1995, 55, 105–138. [Google Scholar] [CrossRef]
- Nielsen, N.; Ingvartsen, K. Propylene glycol for dairy cowsA review of the metabolism of propylene glycol and its effects on physiological parameters, feed intake, milk production and risk of ketosis. Anim. Feed Sci. Technol. 2004, 115, 191–213. [Google Scholar] [CrossRef]
- Ferraro, S.M.; Mendoza, G.D.; Miranda, L.A.; Gutiérrez, C.G. In vitro ruminal fermentation of glycerol, propylene glycol and molasses combined with forages and their effect on glucose and insulin blood plasma concentrations after an oral drench in sheep. Anim. Feed Sci. Technol. 2016, 213, 74–80. [Google Scholar] [CrossRef]
- Trabue, S.; Scoggin, K.; Tjandrakusuma, S.; Rasmussen, M.A.; Reilly, P.J. Ruminal fermentation of propylene glycol and glycerol. J. Agric. Food Chem. 2007, 55, 7043–7051. [Google Scholar] [CrossRef]
- Johnson, R. The treatment of ketosis with glycerol and propilenglicol. Cornell Vet. 1954, 44, 6. [Google Scholar] [PubMed]
- Rukkwamsuk, T.; Rungruang, S.; Choothesa, A.; Wensing, T. Effect of propylene glycol on fatty liver development and hepatic fructose 1,6 bisphosphatase activity in periparturient dairy cows. Livest. Prod. Sci. 2005, 95, 95–102. [Google Scholar] [CrossRef]
- Christensen, J.O.; Grummer, R.R.; Rasmussen, F.E.; Bertics, S.J. Effect of method of delivery of propylene glycol on plasma metabolites of feed-restricted cattle. J. Dairy Sci. 1997, 80, 563–568. [Google Scholar] [CrossRef]
- Rémond, B.; Souday, E.; Jouany, J.P. In vitro and in vivo fermentation of glycerol by rumen microbes. Anim. Feed Sci. Technol. 1993, 41, 121–132. [Google Scholar] [CrossRef]
- Chung, Y.-H.; Rico, D.E.; Martinez, C.M.; Cassidy, T.W.; Noirot, V.; Ames, A.; Varga, G.A. Effects of Feeding Dry Glycerin to Early Postpartum Holstein Dairy Cows on Lactational Performance and Metabolic Profiles. J. Dairy Sci. 2007, 90, 5682–5691. [Google Scholar] [CrossRef]
- Czerkawski, J.W.; Breckenridge, G. Dissimilation of 1,2-propanediol by rumen micro-organisms. Br. J. Nutr. 1973, 29, 317–330. [Google Scholar] [CrossRef] [Green Version]
- Kristensen, N.B.; Raun, B.M.L. Ruminal and intermediary metabolism of propylene glycol in lactating Holstein cows. J. Dairy Sci. 2007, 90, 4707–4717. [Google Scholar] [CrossRef]
- Werner Omazic, A.; Kronqvist, C.; Zhongyan, L.; Martens, H.; Holtenius, K. The fate of glycerol entering the rumen of dairy cows and sheep. J. Anim. Physiol. Anim. Nutr. (Berl) 2015, 99, 258–264. [Google Scholar] [CrossRef] [PubMed]
- Piantoni, P.; Allen, M.S. Evaluation of propylene glycol and glycerol infusions as treatments for ketosis in dairy cows. J. Dairy Sci. 2015, 98, 5429–5439. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gootwine, E.; Reicher, S.; Rozov, A. Prolificacy and lamb survival at birth in Awassi and Assaf sheep carrying the FecB (Booroola) mutation. Anim Reprod Sci 2008, 108, 402–411. [Google Scholar] [CrossRef] [PubMed]
- Gootwine, E. Mini review: Breeding Awassi and Assaf sheep for diverse management conditions. Trop. Anim. Health Prod. 2011, 43, 1289–1296. [Google Scholar] [CrossRef] [PubMed]
- Panousis, N.; Brozos, C.; Karagiannis, I.; Giadinis, N.D.; Lafi, S.; Kritsepi-Konstantinou, M. Evaluation of Precision Xceed® meter for on-site monitoring of blood β-hydroxybutyric acid and glucose concentrations in dairy sheep. Res. Vet. Sci. 2012, 93, 435–439. [Google Scholar] [CrossRef]
- Pichler, M.; Damberger, A.; Schwendenwein, I.; Gasteiner, J.; Drillich, M.; Iwersen, M. Thresholds of whole-blood β-hydroxybutyrate and glucose concentrations measured with an electronic hand-held device to identify ovine hyperketonemia. J. Dairy Sci. 2014, 97, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Matthews, J.N.; Altman, D.G.; Campbell, M.J.; Royston, P. Analysis of serial measurements in medical research. BMJ 1990, 300, 230–235. [Google Scholar] [CrossRef] [PubMed]
- Bell, A.W. Regulation of organic nutrient metabolism during transition from late pregnancy to early lactation. J Anim Sci 1995, 73, 2804–2819. [Google Scholar] [CrossRef] [PubMed]
- Adewuyi, A.A.; Gruys, E.; van Eerdenburg, F.J. Non esterified fatty acids (NEFA) in dairy cattle. A review. Vet Q 2005, 27, 117–126. [Google Scholar] [CrossRef] [Green Version]
- Wroblewski, F.; Gregory, K.F. Lactic Dehydrogenase Isozymes and Their Distribution in Normal Tissues and Plasma and in Disease States*. Ann. N. Y. Acad. Sci. 1961, 94, 912–932. [Google Scholar] [CrossRef]
- Kwo, P.Y.; Cohen, S.M.; Lim, J.K. ACG Clinical Guideline: Evaluation of Abnormal Liver Chemistries. Am. J. Gastroenterol. 2017, 112, 18–35. [Google Scholar] [CrossRef] [PubMed]
- Barcellini, W.; Fattizzo, B. Clinical Applications of Hemolytic Markers in the Differential Diagnosis and Management of Hemolytic Anemia. Dis. Markers 2015, 2015, 635670. [Google Scholar] [CrossRef] [PubMed]
- Maurer, M.; Peinhopf, W.; Gottschalk, J.; Einspanier, A.; Koeller, G.; Wittek, T. Effects of different dosages of propylene glycol in dry cows and cows in early lactation. J. Dairy Res. 2017, 84, 375–384. [Google Scholar] [CrossRef] [PubMed]
- Sauer, F.D.; Erfle, J.D.; Fisher, L.J. Propylene Glycol and Glycerol As a Feed Additive for Lactating Dairy Cows: An Evaluation of Blood Metabolite Parameters. Can. J. Anim. Sci. 1973, 53, 265–271. [Google Scholar] [CrossRef]
- White, H.M. The Role of TCA Cycle Anaplerosis in Ketosis and Fatty Liver in Periparturient Dairy Cows. Animals 2015, 5, 793–802. [Google Scholar] [CrossRef] [PubMed]
- Baird, G.D.; Heitzman, R.J. Mode of action of a glucocorticoid on bovine intermediary metabolism. Possible role in controlling hepatic ketogenesis. BBA—Gen. Subj. 1971, 252, 184–198. [Google Scholar] [CrossRef]
- Brockman, R.P. Roles for insulin and glucagon in the development of ruminant ketosis—A review. Can. Vet. J. La. Rev. Vet. Can. 1979, 20, 121–126. [Google Scholar]
- Sengupta, S.; Peterson, T.R.; Laplante, M.; Oh, S.; Sabatini, D.M. mTORC1 controls fasting-induced ketogenesis and its modulation by ageing. Nature 2010, 468, 1100–1104. [Google Scholar] [CrossRef]
- Brockman, R.P. Insulin and glucagon responses in plasma to intraportal infusions of propionate and butyrate in sheep (Ovis aries). Comp. Biochem. Physiol.—Part A Physiol. 1982, 73, 237–238. [Google Scholar] [CrossRef]
- West, C.E.; Passey, R.F. Effect of glucose load and of insulin on the metabolism of glucose and of palmitate in sheep. Biochem. J. 1967, 102, 58–64. [Google Scholar] [CrossRef] [Green Version]
- Brockman, R.P.; Laarveld, B. Hormonal regulation of metabolism in ruminants; a review. Livest. Prod. Sci. 1986, 14, 313–334. [Google Scholar] [CrossRef]
- McGarry, J.D.; Foster, D.W. The regulation of ketogenesis from oleic acid and the influence of antiketogenic agents. J. Biol. Chem. 1971, 246, 6247–6253. [Google Scholar] [PubMed]
- McGarry, J.D.; Foster, D.W. Regulation of hepatic fatty acid oxidation and ketone body production. Annu. Rev. Biochem. 1980, 49, 395–420. [Google Scholar] [CrossRef] [PubMed]
- Christopher, M.M.; Eckfeldt, J.H.; Eaton, J.W. Propylene glycol ingestion causes D-lactic acidosis. Lab. Invest. 1990, 62, 114–118. [Google Scholar] [PubMed]
- Lomax, M.A.; Baird, G.D. Blood flow and nutrient exchange across the liver and gut of the dairy cow. Br. J. Nutr. 1983, 49, 481–496. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ruddick, J.A. Toxicology, metabolism, and biochemistry of 1,2-propanediol. Toxicol. Appl. Pharmacol. 1972, 21, 102–111. [Google Scholar] [CrossRef]
- POTTER, B.J. Haemoglobinuria caused by propylene glycol in sheep. Br. J. Pharmacol. Chemother. 1958, 13, 385–389. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment | p-Value | |||||||
|---|---|---|---|---|---|---|---|---|
| Glycerol (Mean) | PG (Mean) | SEM | Treatment | Time | Treatment × Time | Animal Effect | Pretreatment | |
| BHBA, mM | 0.49 | 0.46 | 0.03 | 0.643 | 0.0001 | 0.0001 | 0.0492 | 0.0336 |
| Glucose, mg/dL | 69.0 | 59.0 | 1.14 | 0.0001 | 0.0006 | 0.0005 | 0.1760 | 0.0029 |
| Lactate, mmol/L | 0.89 | 1.48 | 0.075 | 0.0002 | 0.0001 | 0.0001 | 0.0307 | 0.8167 |
| Insulin *, µg/L | 0.31 | 0.25 | 0.01 | 0.0078 | 0.0057 | 0.0884 | 1 | 0.4360 |
| NEFA, µEq/L | 982 | 1144 | 10.7 | 0.047 | 0.0001 | 0.0001 | 0.09 | 0.0781 |
| LDH, U/L | 498.7 | 515.7 | 24.4 | 0.648 | 0.0174 | 0.0013 | 0.121 | 0.0178 |
| AST, U/L | 117.5 | 107.1 | 6.86 | 0.312 | 0.0008 | 0.0018 | 0.029 | 0.0003 |
| ALT, U/L | 10.9 | 11.3 | 0.62 | 0.710 | 0.0003 | 0.0005 | 0.0401 | 0.0001 |
| Total Bilirubin, µmol/L | 2.25 | 3.93 | 0.35 | 0.0074 | 0.0123 | 0.0001 | 0.05 | 0.0095 |
| Conjugated Bilirubin, µmol/L | 1.05 | 1.39 | 0.15 | 0.174 | 0.0283 | 0.0001 | 0.067 | 0.0033 |
| Treatment | SEM | p-Value | ||||
|---|---|---|---|---|---|---|
| Glycerol | PG | Treatment | Pretreatment | Body Weight | ||
| (Mean) | (Mean) | |||||
| BHBA | ||||||
| Delta % | 61 | 57.6 | 7.02 | 0.746 | 0.215 | 1 |
| AUC, min × mM | 79.1 | 64.1 | 15 | 0.494 | 0.887 | NA |
| Glucose | ||||||
| Delta % | 42.3 | 11.2 | 2.69 | 0.0001 | 0.0002 | 0.0102 |
| AUC, min × mg/dL * | 3178.2 | 239 | 377.3 | 0.0001 | 0.065 | NA |
| Lactate | ||||||
| Delta % | 87.1 | 192.8 | 11.1 | 0.0001 | 0.0001 | 0.0227 |
| AUC, min × mmol/L | 71.8 | 239.1 | 19.2 | 0.0001 | 0.236 | NA |
| Insulin | ||||||
| Delta % | 41.7 | 7.2 | 10.3 | 0.0399 | 0.113 | 0.735 |
| *AUC, min × µg/L | 14. 6 | −5.1 | 3.75 | 0.0075 | 0.718 | NA |
| NEFA | ||||||
| Delta % | 73.2 | 71.4 | 2.67 | 0.692 | 0.0286 | 0.518 |
| AUC, min × µEq/L | 178194.2 | 62670.9 | 15064.5 | 0.0001 | 0.0001 | NA |
| Total Bilirubin | ||||||
| Delta % | 59.1 | 155.8 | 27.2 | 0.033 | 0.0109 | 0.285 |
| AUC, min × µmol/L | −92.78 | 505.4 | 147.1 | 0.0161 | 0.6839 | NA |
| Conjugated Bilirubin | ||||||
| Delta % | 33.7 | 61.8 | 23.8 | 0.464 | 0.383 | 0.582 |
| AUC, min × µmol/L | 51.8 | 25 | 49 | 0.735 | 0.258 | NA |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kalyesubula, M.; Rosov, A.; Alon, T.; Moallem, U.; Dvir, H. Intravenous Infusions of Glycerol Versus Propylene Glycol for the Regulation of Negative Energy Balance in Sheep: A Randomized Trial. Animals 2019, 9, 731. https://doi.org/10.3390/ani9100731
Kalyesubula M, Rosov A, Alon T, Moallem U, Dvir H. Intravenous Infusions of Glycerol Versus Propylene Glycol for the Regulation of Negative Energy Balance in Sheep: A Randomized Trial. Animals. 2019; 9(10):731. https://doi.org/10.3390/ani9100731
Chicago/Turabian StyleKalyesubula, Mugagga, Alexander Rosov, Tamir Alon, Uzi Moallem, and Hay Dvir. 2019. "Intravenous Infusions of Glycerol Versus Propylene Glycol for the Regulation of Negative Energy Balance in Sheep: A Randomized Trial" Animals 9, no. 10: 731. https://doi.org/10.3390/ani9100731
APA StyleKalyesubula, M., Rosov, A., Alon, T., Moallem, U., & Dvir, H. (2019). Intravenous Infusions of Glycerol Versus Propylene Glycol for the Regulation of Negative Energy Balance in Sheep: A Randomized Trial. Animals, 9(10), 731. https://doi.org/10.3390/ani9100731