Effect of Feeding Cold-Pressed Sunflower Cake on Ruminal Fermentation, Lipid Metabolism and Bacterial Community in Dairy Cows

 , , , and
, , , and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals and Experimental Design
2.2. Sampling and Measurements
2.3. Sample Handling and Laboratory Procedures
2.3.1. Feed
2.3.2. Purine Derivatives Determination
2.3.3. Short Chain Fatty Acid Determination
2.3.4. Ruminal Fatty Acid Profile Determination
2.3.5. DNA Extraction and Illumina Library Generation
2.4. Calculations and Statistical Analysis
3. Results
3.1. Rumen Fatty Cid Composition
3.2. Ruminal Fermentation and Microbial Protein Synthesis
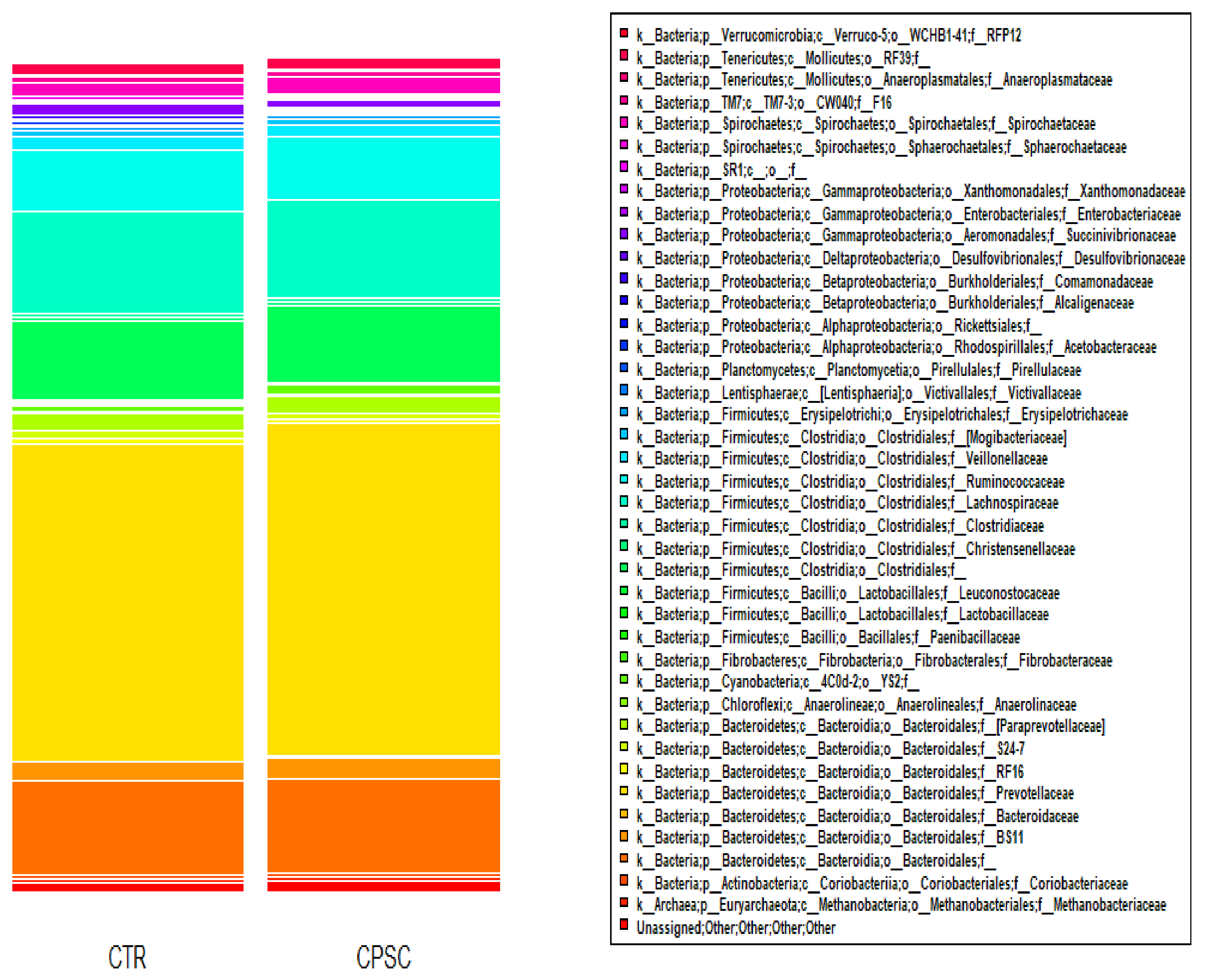
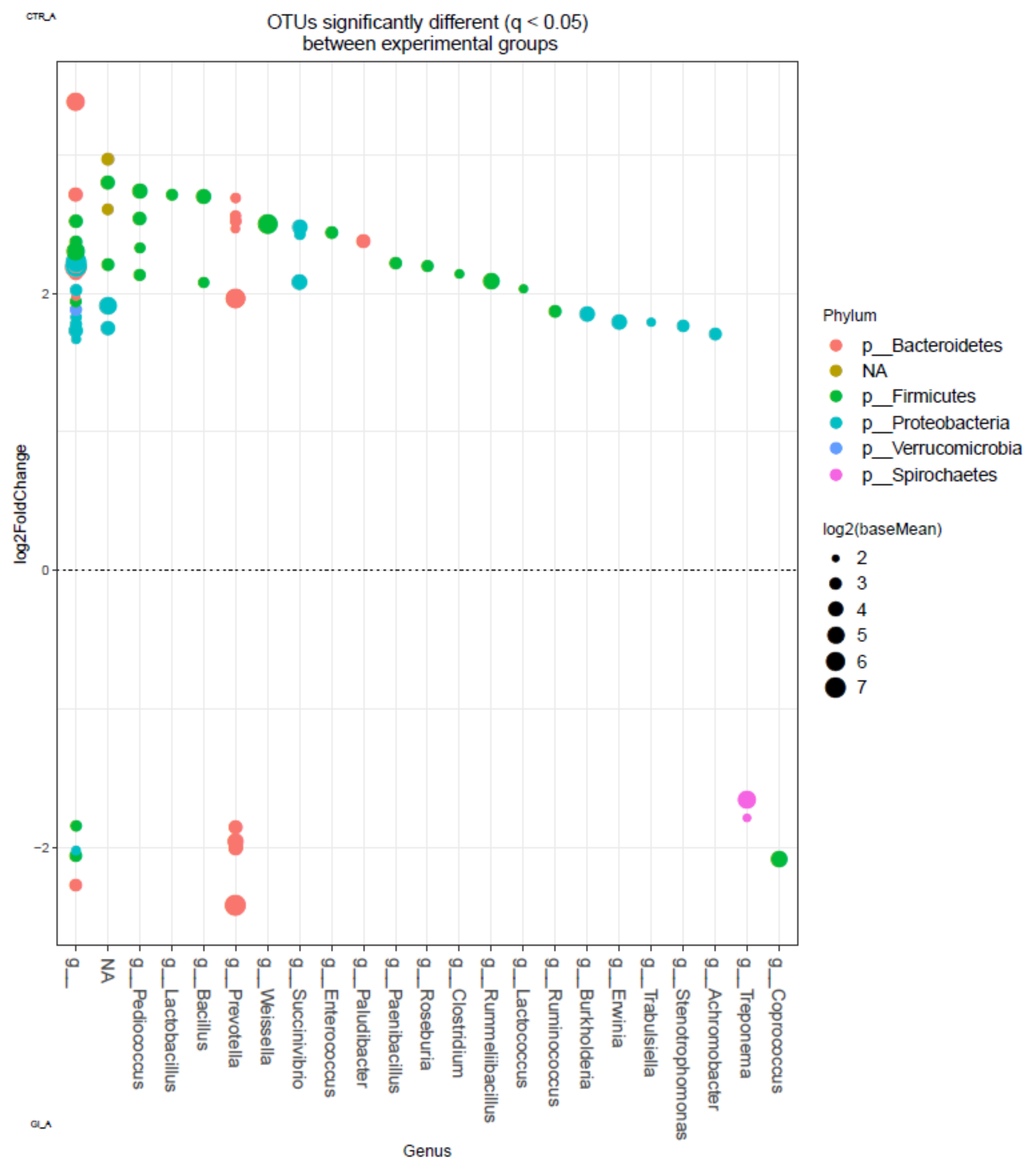
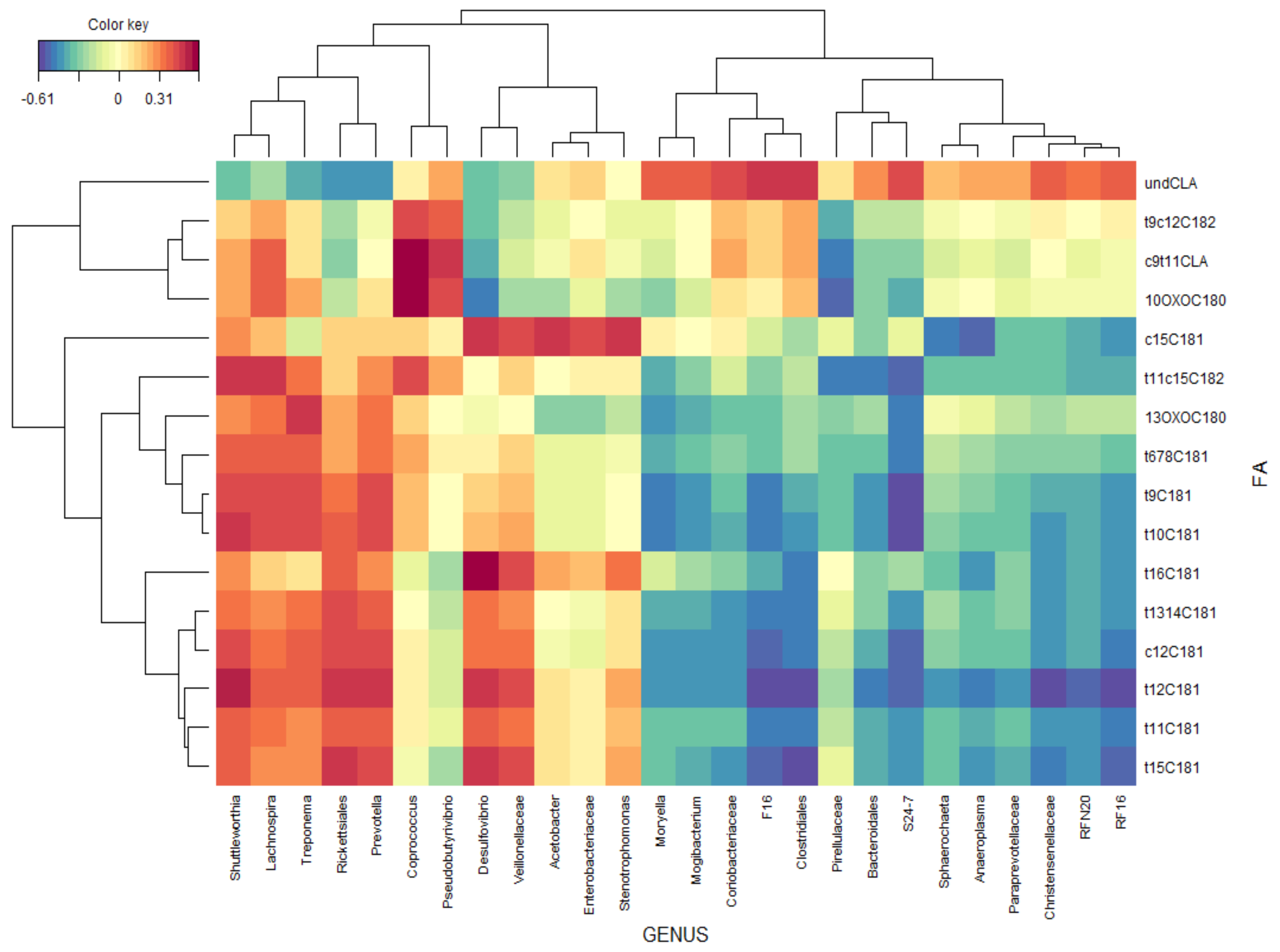
3.3. Ruminal Bacterial Community
4. Discussion
4.1. Ruminal Biohydrogenation
4.2. Rumen Fermentation and Microbial Protein Synthesis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Shingfield, K.J.; Bonnet, M.; Scollan, N. Recent developments in altering the fatty acid composition of ruminant-derived foods. Animal 2013, 7, 132–162. [Google Scholar] [CrossRef] [PubMed]
- Lock, A.L.; Bauman, D.E. Modifying milk fat composition of dairy cows to enhance fatty acids beneficial to human health. Lipids 2004, 39, 1197–1206. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Toral, P.; Monahan, F.; Hervás, G.; Frutos, P.; Moloney, A. Review: Modulating ruminal lipid metabolism to improve the fatty acid composition of meat and milk. Challenges and opportunities. Animal 2018, 12, S272–S281. [Google Scholar] [CrossRef] [PubMed]
- Doreau, M.; Bauchart, D.; Chilliard, Y. Enhancing fatty acid composition of milk and meat through animal feeding. Anim. Prod. Sci. 2010, 51, 19–29. [Google Scholar] [CrossRef]
- Ventto, L.; Leskinen, H.; Kairenius, P.; Stefański, T.; Bayat, A.; Vilkki, J.; Shingfield, K. Diet-induced milk fat depression is associated with alterations in ruminal biohydrogenation pathways and formation of novel fatty acid intermediates in lactating cows. Br. J. Nutr. 2017, 117, 364–376. [Google Scholar] [CrossRef] [Green Version]
- Rego, O.A.; Rosa, H.J.D.; Portugal, P.V.; Franco, T.; Vouzela, C.M.; Borba, A.E.S.; Bessa, R.J.B. The effects of supplementation with sunflower and soybean oils on the fatty acid profile of milk fat from grazing dairy cows. Anim. Res. 2005, 54, 17–24. [Google Scholar] [CrossRef] [Green Version]
- Shingfield, K.J.; Ahvenjärvi, S.; Toivonen, V.; Vanhatalo, A.; Huhtanen, P.; Griinari, J.M. Effect of incremental levels of sunflower-seed oil in the diet on ruminal lipid metabolism in lactating cows. Br. J. Nutr. 2008, 99, 971–983. [Google Scholar] [CrossRef]
- Rego, O.A.; Alves, S.P.; Antunes, L.M.; Rosa, H.J.; Alfaia, C.F.; Prates, J.A.; Cabrita, A.R.; Fonseca, A.J.; Bessa, R.J. Rumen biohydrogenation-derived fatty acids in milk fat from grazing dairy cows supplemented with rapeseed, sunflower, or linseed oils. J. Dairy Sci. 2009, 92, 4530–4540. [Google Scholar] [CrossRef] [Green Version]
- Palmquist, D.L.; Lock, A.L.; Shingfield, K.J.; Bauman, D.E. Biosynthesis of conjugated linoleic acid in ruminants and humans. Adv. Food Nutr. Res. 2005, 50, 179–217. [Google Scholar] [CrossRef]
- Benhissi, H.; Beltrán de Heredia, I.; Garcia-Rodriguez, A. Effect of replacing palm fat with high-linoleic cold-pressed rapeseed or sunflower cakes on fatty acid biohydrogenation in an artificial rumen (Rusitec). Anim. Prod. Sci. 2016, 58, 499–506. [Google Scholar] [CrossRef]
- Pascual, A.; Pineda-Quiroga, C.; Goiri, I.; Atxaerandio, R.; Ruiz, R.; García-Rodríguez, A. Effects of feeding UFA-rich cold-pressed oilseed cakes and sainfoin on dairy ewes’ milk fatty acid profile and curd sensory properties. Small Rumin. Res. 2019, 175, 96–103. [Google Scholar] [CrossRef]
- Benhissi, H.; García-Rodríguez, A.; Beltrán de Heredia, I. Effect of type and inclusion level of cold-pressed oilseed cakes on in vitro rumen fermentation. Anim. Prod. Sci. 2014, 54, 1709–1713. [Google Scholar] [CrossRef]
- Amores, G.; Virto, M.; Nájera, A.I.; Mandaluniz, N.; Arranz, J.; Bustamante, M.A.; Valdivielso, I.; Ruiz de Gordoa, J.C.; García-Rodríguez, A.; Barron, L.J.; et al. Rapeseed and sunflower oilcake as supplements for dairy sheep: Animal performance and milk fatty acid concentrations. J. Dairy Res. 2014, 81, 410–416. [Google Scholar] [CrossRef] [PubMed]
- Association of Official Analytical Chemists (AOAC). Official Methods of Analysis, 18th ed.; AOAC: Gaitherburg, MD, USA, 2007. [Google Scholar]
- Van Soest, P.J.; Robertson, J.B.; Lewis, B.A. Methods for dietary fiber, neutral detergent fiber, and nonstarch polysaccharides in relation to animal nutrition. J. Dairy Sci. 1991, 74, 3583–3597. [Google Scholar] [CrossRef]
- Robertson, J.B.; Van Soest, P.J. The detergent system of analysis. In The Analysis of Dietary Fibre in Food; James, W.P.T., Theander, O., Eds.; Marcel Dekker: New York, USA, 1981; pp. 123–158. [Google Scholar]
- MAPA. Métodos Oficiales del MAPA; BOE 52, 7167; MAPA: Madrid, Spain, 1995. [Google Scholar]
- Reynal y Broderick. Technical note: A new high-performance liquid chromatography purine assay for quantifying microbial flow. J. Dairy Sci. 2009, 92, 1177–1181. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Toral, P.G.; Bernard, L.; Belenguer, A.; Rouel, J.; Hervás, G.; Chilliard, Y.; Frutos, P. Comparison of ruminal lipid metabolism in dairy cows and goats fed diets supplemented with starch, plant oil, or fish oil. J. Dairy Sci. 2016, 99, 301–316. [Google Scholar] [CrossRef] [PubMed]
- Shingfield, K.J.; Ahvenjärvi, S.; Toivonen, V.; Ärölä, A.; Nurmela, K.; Huhtanen, P.; Griinari, J. Effect of dietary fish oil on biohydrogenation of fatty acids and milk fatty acid content in cows. Anim. Sci. 2003, 77, 165–179. [Google Scholar] [CrossRef]
- Toral, P.G.; Shingfield, K.J.; Hervás, G.; Toivonen, V.; Frutos, P. Effect of fish oil and sunflower oil on rumen fermentation characteristics and fatty acid composition of digesta in ewes fed a high concentrate diet. J. Dairy Sci. 2010, 93, 4804–4817. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Caporaso, J.G.; Lauber, C.L.; Walters, W.A.; Berg-Lyons, D.; Lozupone, C.A.; Turnbaugh, P.J.; Fierer, N.; Knight, R. Global patterns of 16S rRNA diversity at a depth of millions of sequences per sample. Proc. Natl. Acad. Sci. USA 2011, 108, 4516–4522. [Google Scholar] [CrossRef]
- Magoč, T.; Salzberg, S.L. FLASH: Fast length adjustment of short reads to improve genome assemblies. Bioinformatics 2011, 27, 2957–2963. [Google Scholar] [CrossRef]
- Caporaso, J.G.; Kuczynski, J.; Stombaugh, J.; Bittinger, K.; Bushman, F.D.; Costello, E.K.; Fierer, N.; Peña, A.G.; Goodrich, J.K.; Gordon, J.I.; et al. QIIME allows analysis of high-throughput community sequencing data. Nat. Methods 2010, 7, 335–336. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Edgar, R.C. Search and clustering orders of magnitude faster than BLAST. Bioinformatics 2010, 26, 2460–2461. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McDonald, D.; Price, M.N.; Goodrich, J.; Nawrocki, E.P.; DeSantis, T.Z.; Probst, A.; Andersen, G.L.; Knight, R.; Hugenholtz, P. An improved Greengenes taxonomy with explicit ranks for ecological and evolutionary analyses of bacteria and archaea. ISME J. 2012, 6, 610–618. [Google Scholar] [CrossRef] [PubMed]
- Broderick, G.A.; Merchen, N.R. Markers for quantifying microbial protein synthesis in the rumen. J. Dairy Sci. 1992, 75, 2618–2632. [Google Scholar] [CrossRef]
- Valadares, R.F.D.; Broderick, G.A.; Valadares Filho, S.C.; Clayton, M.K. Effect of replacing alfalfa silage with high moisture corn on ruminal protein synthesis estimated from excretion of total purine derivatives. J. Dairy Sci. 1999, 82, 2686–2696. [Google Scholar] [CrossRef]
- SAS. Enterprise’s Guide; SAS: Cary, NC, USA, 2017. [Google Scholar]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef] [Green Version]
- Le Cao, K.-A.; Rohart, F.; Gonzalez, I.; Dejean, S.; Gautier, B.; Bartolo, F.; Monget, P.; Coquery, J.; Yao, F.Z.; Liquet, B. Mixomics: Omics Data Integration Project. R Package Version 6.1.1. 2016. Available online: https://CRAN.R-project.org/package=mixOmics (accessed on 7 August 2019).
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2018; Available online: http://www.R-project.org/ (accessed on 7 August 2019).
- McKain, N.; Shingfield, K.J.; Wallace, R. Metabolism of conjugated linoleic acids and 18: 1 fatty acids by ruminal bacteria: Products and mechanisms. Microbiology 2010, 156, 579–588. [Google Scholar] [CrossRef] [PubMed]
- Zened, A.; Enjalbert, F.; Nicot, M.C.; Troegeler-Meynadier, A. Starch plus sunflower oil addition to the diet of dry dairy cows results in a trans-11 to trans-10 shift of biohydrogenation. J. Dairy Sci. 2013, 96, 451–459. [Google Scholar] [CrossRef] [Green Version]
- Moon, C.; Pacheco, D.; Kelly, W.; Leahy, S.; Li, D.; Kopečný, J.; Attwood, G. Reclassification of Clostridium proteoclasticum as Butyrivibrio proteoclasticus comb. nov., a butyrate-producing ruminal bacterium. Int. J. Syst. Evol. Microbiol. 2008, 58, 2041–2045. [Google Scholar] [CrossRef] [Green Version]
- Yokoyama, M.T.; Davis, C.L. Hydrogenation of unsaturated fatty acids by Treponema (Borrelia) strain B 2 5, a rumen spirochete. J. Bact. 1971, 107, 519–527. [Google Scholar]
- Dewanckele, L.; Vlaeminck, B.; Hernandez-Sanabria, E.; Ruiz-González, A.; Debruyne, S.; Jeyanathan, J.; Fievez, V. Rumen biohydrogenation and microbial community changes upon early life supplementation of 22:6n-3 enriched microalgae to goats. Front. Microbiol. 2018, 9, 573. [Google Scholar] [CrossRef] [PubMed]
- Bauman, D.E.; Perfield, J.W.; de Veth, M.J.; Lock, A.L. New perspectives on lipid digestion and metabolism in ruminants. In Proceedings of the Cornell Nutrition Conference, The Cornell University, New York, NY, USA, 21–23 October 2013; pp. 175–189. [Google Scholar]
- Lee, Y.J.; Jenkins, T.C. Biohydrogenation of linolenic acid to stearic acid by the rumen microbial population yields multiple intermediate conjugated diene isomers. J. Nutr. 2011, 141, 1445–1450. [Google Scholar] [CrossRef] [PubMed]
- Palmquist, D.L.; Jenkins, T.C. A 100-Year Review: Fat feeding of dairy cows. J. Dairy Sci. 2017, 100, 10061–10077. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Petri, R.M.; Vahmani, P.; Yang, H.E.; Dugan, M.E.R.; McAllister, T.A. Changes in rumen microbial profiles and subcutaneous fat composition when feeding extruded flaxseed mixed with or before hay. Front. Microbiol. 2018, 9, 1055. [Google Scholar] [CrossRef]
- Chow, T.T.; Fievez, V.; Moloney, A.P.; Raes, K.; Demeyer, D.; De Smet, S. Effect of fish oil on in vitro rumen lipolysis, apparent biohydrogenation of linoleic acid and accumulation of biohydrogenation intermediates. Anim. Feed Sci. Technol. 2004, 117, 1–12. [Google Scholar] [CrossRef]
- Jenkins, T.C. Lipid metabolism in the rumen. J. Dairy Sci. 1993, 76, 3851–3863. [Google Scholar] [CrossRef]
- Bayat, R.; Ventto, L.; Kairenius, P.; Stefański, T.; Leskinen, H.; Tapio, I.; Negussie, E.; Vilkki, J.; Shingfield, K.J. Dietary forage to concentrate ratio and sunflower oil supplement alter rumen fermentation, ruminal methane emissions, and nutrient utilization in lactating cows. Transl. Anim. Sci. 2017, 1, 277–286. [Google Scholar] [CrossRef]
- Beauchemin, K.A.; McGinn, S.M.; Benchaar, C.; Holtshausen, L. Crushed sunflower, flax, or canola seeds in lactating dairy cow diets: Effects on methane production, rumen fermentation, and milk production. J. Dairy Sci. 2009, 92, 2118–2127. [Google Scholar] [CrossRef]
- Maia, M.R.G.; Chaudhary, L.C.; Figueres, L.; Wallace, R.J. Metabolism of polyunsaturated fatty acids and their toxicity to the microflora of the rumen. Antonie Leeuwenhoek 2007, 91, 303–314. [Google Scholar] [CrossRef]
- Jenkins, T.C.; Wallace, R.J.; Moate, P.J.; Mosley, E.E. Board-invited review: Recent advances in biohydrogenation of unsaturated fatty acids within the rumen microbial ecosystem. J. Anim. Sci. 2008, 86, 397–412. [Google Scholar] [CrossRef]
- Maia, M.R.; Chaudhary, L.C.; Bestwick, C.S.; Richardson, A.J.; McKain, N.; Larson, T.R.; Graham, I.A.; Wallace, R.J. Toxicity of unsaturated fatty acids to the biohydrogenating ruminal bacterium, Butyrivibrio fibrisolvens. BMC Microbiol. 2010, 10, 52. [Google Scholar] [CrossRef] [PubMed]
- Mutsvangwa, T.; Hobin, M.R.; Gozho, G.N. Effects of method of barley grain processing and source of supplemental dietary fat on duodenal nutrient flows, milk fatty acid profiles, and microbial protein synthesis in dairy cows. J. Dairy Sci. 2012, 95, 5961–5977. [Google Scholar] [CrossRef] [PubMed]
- Oldick, B.S.; Firkins, J.L. Effects of degree of fat saturation on fiber digestion and microbial protein synthesis when diets are fed twelve times daily. J. Anim. Sci. 2000, 78, 2412–2420. [Google Scholar] [CrossRef] [PubMed]
- Ivan, M.; Mir, P.S.; Mir, Z.; Entz, T.; He, M.L.; McAllister, T.A. Effects of dietary sunflower seeds on rumen protozoa and growth of lambs. Br. J. Nutr. 2004, 92, 303–310. [Google Scholar] [CrossRef] [Green Version]
- Doranalli, K.; Mutsvangwa, T. Feeding sunflower oil to partially defaunate the rumen increases nitrogen retention, urea-nitrogen recycling to the gastrointestinal tract and the anabolic use of recycled urea-nitrogen in growing lambs. Br. J. Nutr. 2011, 105, 1453–1464. [Google Scholar] [CrossRef] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Ingredients (% on DM Basis) | CTR | CPSC | Basal Diet |
|---|---|---|---|
| Corn | 23.7 | 19.0 | |
| Soybean meal | 20.0 | 15.0 | |
| Cold-pressed sunflower cake | 0 | 23.0 | |
| Palm kernel meal | 15.0 | ||
| DDGs | 14.9 | 1.0 | |
| Barley | 10.8 | 15.7 | |
| Wheat | 6.0 | 15.0 | |
| Molasses | 2.0 | 2.0 | |
| Hydrogenated palm fat | 2.0 | 0 | |
| Alfalfa pellets | 2.0 | 5.5 | |
| Minerals and vitamins1 | 3.6 | 3.8 | |
| Maize silage | 29 | ||
| Grass silage | 61 | ||
| Barley straw | 9 | ||
| Chemical composition | |||
| UFL | 1.02 | 1.02 | |
| Fat (%) | 5.6 | 5.6 | 2.8 |
| Acid detergent fibre (%) | 9.7 | 9.8 | 33.6 |
| Neutral detergent fibre (%) | 22.5 | 19.5 | 41.1 |
| Acid detergent lignin (%) | 2.2 | 2.3 | 4.8 |
| Crude protein (%) | 19.0 | 19.0 | 10.7 |
| Starch (%) | 31.7 | 29.9 | 5.6 |
| Fatty acids (% FA) | |||
| C12:0 | 7.77 | 0.12 | |
| C13:0 | 0.03 | 0.02 | |
| C14:0 | 3.09 | 0.30 | |
| C150 | 0.04 | 0.04 | |
| C16:0 | 23.55 | 12.05 | |
| C17:0 | 0.08 | 0.08 | |
| C18:0 | 2.90 | 3.67 | |
| C20:0 | 0.33 | 0.34 | |
| C22:0 | 0.15 | 0.49 | |
| C23:0 | 0.06 | 0.08 | |
| C16:1 cis-9 | 0.12 | 0.16 | |
| C18:1 cis-9 | 25.39 | 20.56 | |
| C18:1 cis-11 | 1.13 | 1.59 | |
| C20:1 cis-11 | 0.22 | 0.22 | |
| C18:2n-6 | 33.04 | 57.82 | |
| C18:3n-3 | 1.67 | 1.53 |
| Item (g/100 g FA) | CTR | CPSC | SED | p-Value |
|---|---|---|---|---|
| C13:0 | 0.053 | 0.054 | 0.0056 | 0.875 |
| C13:0 iso | 0.056 | 0.056 | 0.0065 | 0.952 |
| C14:0 | 2.29 | 1.27 | 0.083 | <0.001 |
| C14:0 iso | 0.135 | 0.152 | 0.0272 | 0.558 |
| C15:0 | 0.793 | 0.870 | 0.060 | 0.241 |
| C15:0 iso | 0.245 | 0.252 | 0.0391 | 0.865 |
| C15:0 anteiso | 0.695 | 0.730 | 0.1078 | 0.755 |
| C16:0 | 21.9 | 15.7 | 0.53 | <0.001 |
| C16:0 iso | 0.244 | 0.303 | 0.0516 | 0.286 |
| C17:0 | 0.434 | 0.568 | 0.0174 | <0.001 |
| C17:0 iso | 0.213 | 0.220 | 0.0122 | 0.585 |
| C17:0 anteiso | 0.225 | 0.237 | 0.0307 | 0.704 |
| C18:0 | 44.0 | 51.3 | 0.73 | <0.001 |
| C18:0 iso | 0.057 | 0.070 | 0.0058 | 0.047 |
| 10-oxo-C18:0 | 0.510 | 0.571 | 0.0417 | 0.180 |
| 13-oxo-C18:0 | 0.233 | 0.386 | 0.0208 | <0.001 |
| C19:0 | 0.093 | 0.106 | 0.0037 | 0.007 |
| C20:0 | 0.750 | 0.803 | 0.0136 | 0.004 |
| C22:0 | 0.470 | 0.626 | 0.0326 | 0.001 |
| C23:0 | 0.162 | 0.181 | 0.0098 | 0.084 |
| C24:0 | 0.528 | 0.606 | 0.0135 | <0.001 |
| ∑ SFA | 76.9 | 76.2 | 0.83 | 0.421 |
| Item (g/100 g FA) | CTR | CPSC | SED | p-Value |
|---|---|---|---|---|
| C16:1 cis-9 | 0.068 | 0.074 | 0.0085 | 0.492 |
| C16:1 trans-9 | 0.012 | 0.016 | 0.0024 | 0.117 |
| C18:1 cis-9 | 4.25 | 3.99 | 0.370 | 0.512 |
| C18:1 cis-11 | 0.530 | 0.580 | 0.0455 | 0.297 |
| C18:1 cis-12 | 0.626 | 0.995 | 0.0514 | <0.001 |
| C18:1 cis-13 | 0.130 | 0.132 | 0.0134 | 0.886 |
| C18:1 cis-15 | 0.216 | 0.219 | 0.0099 | 0.774 |
| C18:1 cis-16 | 0.123 | 0.131 | 0.0057 | 0.327 |
| C18:1 trans-4 | 0.197 | 0.231 | 0.0154 | 0.054 |
| C18:1 trans-5 | 0.129 | 0.157 | 0.0123 | 0.052 |
| C18:1 trans-6-7-8 | 0.651 | 0.738 | 0.0363 | 0.043 |
| C18:1 trans-9 | 0.445 | 0.564 | 0.0234 | <0.001 |
| C18:1 trans-10 | 0.760 | 0.944 | 0.0377 | 0.001 |
| C18:1 trans-11 | 4.83 | 5.56 | 0.261 | 0.023 |
| C18:1 trans-12 | 0.910 | 1.120 | 0.020 | <0.001 |
| C18:1 trans-13-14 | 1.18 | 1.45 | 0.089 | 0.016 |
| C18:1 trans-15 | 0.836 | 0.963 | 0.0206 | <0.001 |
| C18:1 trans-16 | 0.761 | 0.849 | 0.0433 | 0.077 |
| C20:1 cis-11 | 0.153 | 0.097 | 0.0199 | 0.024 |
| C22:1 cis-13 | 0.039 | 0.031 | 0.0058 | 0.192 |
| C24:1 cis-15 | 0.098 | 0.091 | 0.0063 | 0.343 |
| C18:2 cis-9 cis-12 | 2.50 | 2.39 | 0.271 | 0.694 |
| C18:2 cis-9 trans-12 | 0.053 | 0.031 | 0.0045 | 0.001 |
| C18:2 trans-11 cis-15 | 0.423 | 0.627 | 0.0548 | 0.006 |
| C18:2 trans-11 trans-15 | 0.096 | 0.136 | 0.0052 | <0.001 |
| C18:2 cis-9 trans-11 CLA | 0.474 | 0.429 | 0.0802 | 0.589 |
| C18:2 trans-9 cis-11 CLA | 0.012 | 0.009 | 0.0046 | 0.536 |
| C18:2 trans-10 cis-12 CLA | 0.069 | 0.066 | 0.0145 | 0.858 |
| C18:2 trans-11 trans-13 CLA | 0.171 | 0.227 | 0.0164 | 0.009 |
| C18:3n-3 | 0.642 | 0.852 | 0.0965 | 0.062 |
| C18:3n-6 | 0.007 | 0.029 | 0.0015 | <0.001 |
| C20:2n-6 | 0.026 | 0.023 | 0.0041 | 0.473 |
| C20:3n-6 | 0.024 | 0.067 | 0.0149 | 0.021 |
| C20:4n-6 | 0.023 | 0.056 | 0.0059 | <0.001 |
| ∑ MUFA trans | 10.70 | 12.60 | 0.330 | <0.001 |
| ∑ MUFA cis | 6.20 | 6.30 | 0.420 | 0.794 |
| ∑ MUFA | 16.90 | 18.90 | 0.590 | 0.009 |
| ∑ PUFA | 4.77 | 5.09 | 0.400 | 0.446 |
| ∑CLA | 0.905 | 0.801 | 0.0955 | 0.306 |
| ∑ n-3 | 0.663 | 0.885 | 0.0967 | 0.050 |
| ∑ n-6 | 0.079 | 0.175 | 0.0179 | <0.001 |
| n-6:n-3 | 0.117 | 0.208 | 0.0243 | 0.006 |
| C18:1 trans-10:trans-11 | 0.161 | 0.172 | 0.0072 | 0.165 |
| Item | CTR | CPSC | SED | p-Value |
|---|---|---|---|---|
| SCFA (mmol/ 100 mmol) | ||||
| Acetate | 66.5 | 67.0 | 0.24 | 0.108 |
| Propionate | 16.9 | 16.9 | 0.21 | 0.940 |
| Isobutyrate | 0.90 | 0.88 | 0.034 | 0.696 |
| Butyrate | 13.2 | 12.7 | 0.11 | 0.005 |
| Isovalerate | 1.06 | 1.05 | 0.059 | 0.835 |
| Valerate | 1.41 | 1.43 | 0.034 | 0.546 |
| Acetate:propionate | 3.94 | 3.97 | 0.061 | 0.669 |
| Urinary purine derivatives excretion, mmol/d | ||||
| Allantoin | 318 | 335 | 8.9 | 0.093 |
| Uric acid | 27 | 29 | 2.1 | 0.439 |
| Creatinine | 41 | 39 | 2.0 | 0.434 |
| Total PD | 345 | 364 | 8.1 | 0.050 |
| Microbial N flow, g/d | 257 | 273 | 6.9 | 0.045 |
| Diversity Indices | CTR | CPSC | SED | p-Value |
|---|---|---|---|---|
| Number of observed OTUs | 13432 | 13703 | 480.35 | 0.5901 |
| Good’s coverage | 0.953 | 0.951 | 0.0027 | 0.4493 |
| Chao1 | 22766 | 23546 | 468.09 | 0.1396 |
| Shannon | 10.18 | 10.23 | 0.193 | 0.8061 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zubiria, I.; Garcia-Rodriguez, A.; Atxaerandio, R.; Ruiz, R.; Benhissi, H.; Mandaluniz, N.; Lavín, J.L.; Abecia, L.; Goiri, I. Effect of Feeding Cold-Pressed Sunflower Cake on Ruminal Fermentation, Lipid Metabolism and Bacterial Community in Dairy Cows. Animals 2019, 9, 755. https://doi.org/10.3390/ani9100755
Zubiria I, Garcia-Rodriguez A, Atxaerandio R, Ruiz R, Benhissi H, Mandaluniz N, Lavín JL, Abecia L, Goiri I. Effect of Feeding Cold-Pressed Sunflower Cake on Ruminal Fermentation, Lipid Metabolism and Bacterial Community in Dairy Cows. Animals. 2019; 9(10):755. https://doi.org/10.3390/ani9100755
Chicago/Turabian StyleZubiria, Izaro, Aser Garcia-Rodriguez, Raquel Atxaerandio, Roberto Ruiz, Hanen Benhissi, Nerea Mandaluniz, Jose Luis Lavín, Leticia Abecia, and Idoia Goiri. 2019. "Effect of Feeding Cold-Pressed Sunflower Cake on Ruminal Fermentation, Lipid Metabolism and Bacterial Community in Dairy Cows" Animals 9, no. 10: 755. https://doi.org/10.3390/ani9100755
APA StyleZubiria, I., Garcia-Rodriguez, A., Atxaerandio, R., Ruiz, R., Benhissi, H., Mandaluniz, N., Lavín, J. L., Abecia, L., & Goiri, I. (2019). Effect of Feeding Cold-Pressed Sunflower Cake on Ruminal Fermentation, Lipid Metabolism and Bacterial Community in Dairy Cows. Animals, 9(10), 755. https://doi.org/10.3390/ani9100755