The Anti-Oxidation and Mechanism of Essential Oil of Paederia scandens in the NAFLD Model of Chicken


Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Materials and Extraction
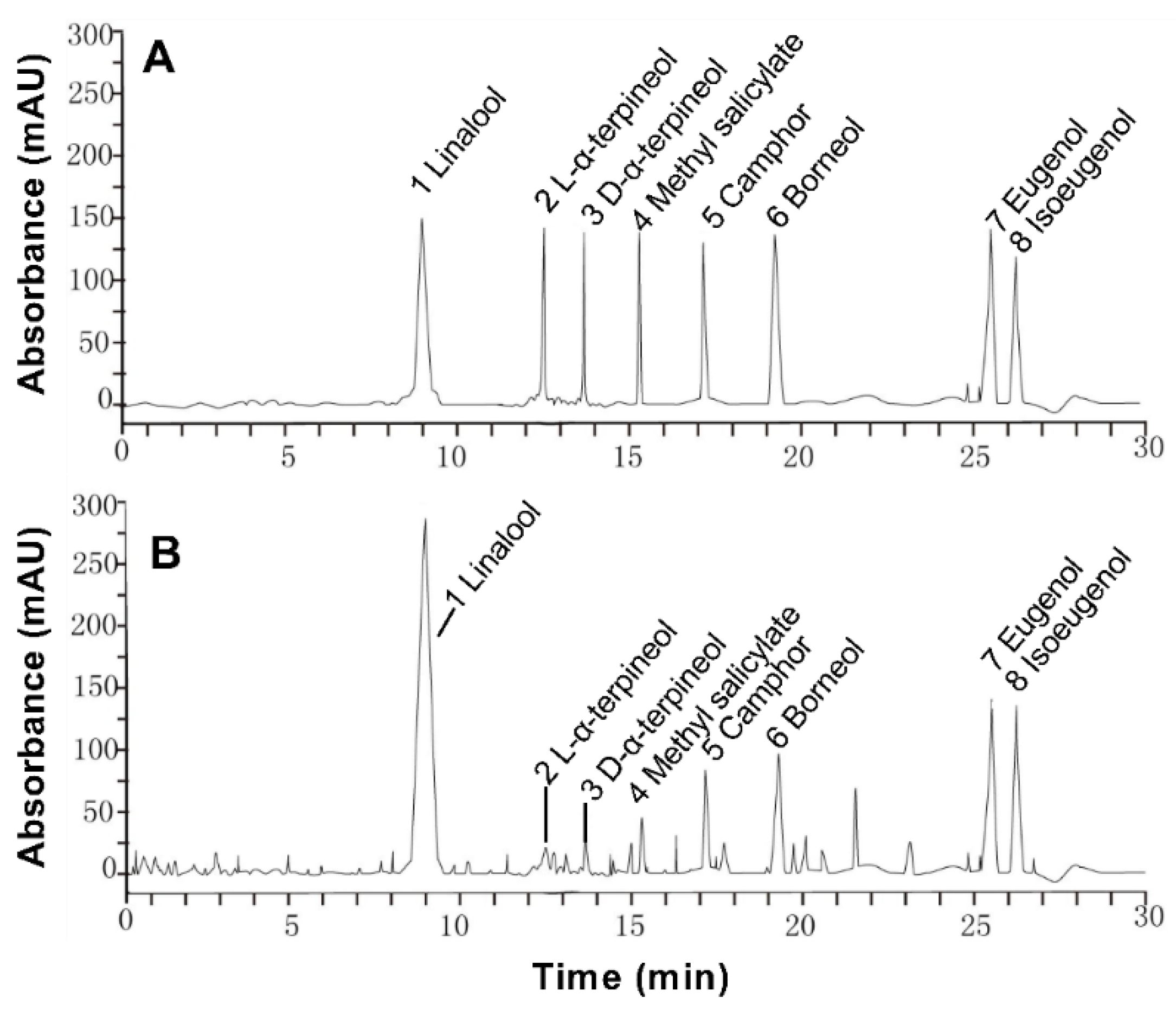
2.2. High Performance Liquid Chromatography (HPLC) Analysis
2.3. Animals and Experimental Procedure
2.4. Oxidative Stress Assays
2.5. Protein Separation and Identification
2.6. Statistical Analysis
3. Results
3.1. Paederia scandens Essential Oil Chemical composition
3.2. Clinical Symptoms and Relative Weight of the Liver
3.3. Hepatic ROS, MDA, and Superoxidase Dismutase (SOD) Levels
3.4. Two-Dimensional (2D) Gel Electrophoresis
3.5. Protein Separation and Identification
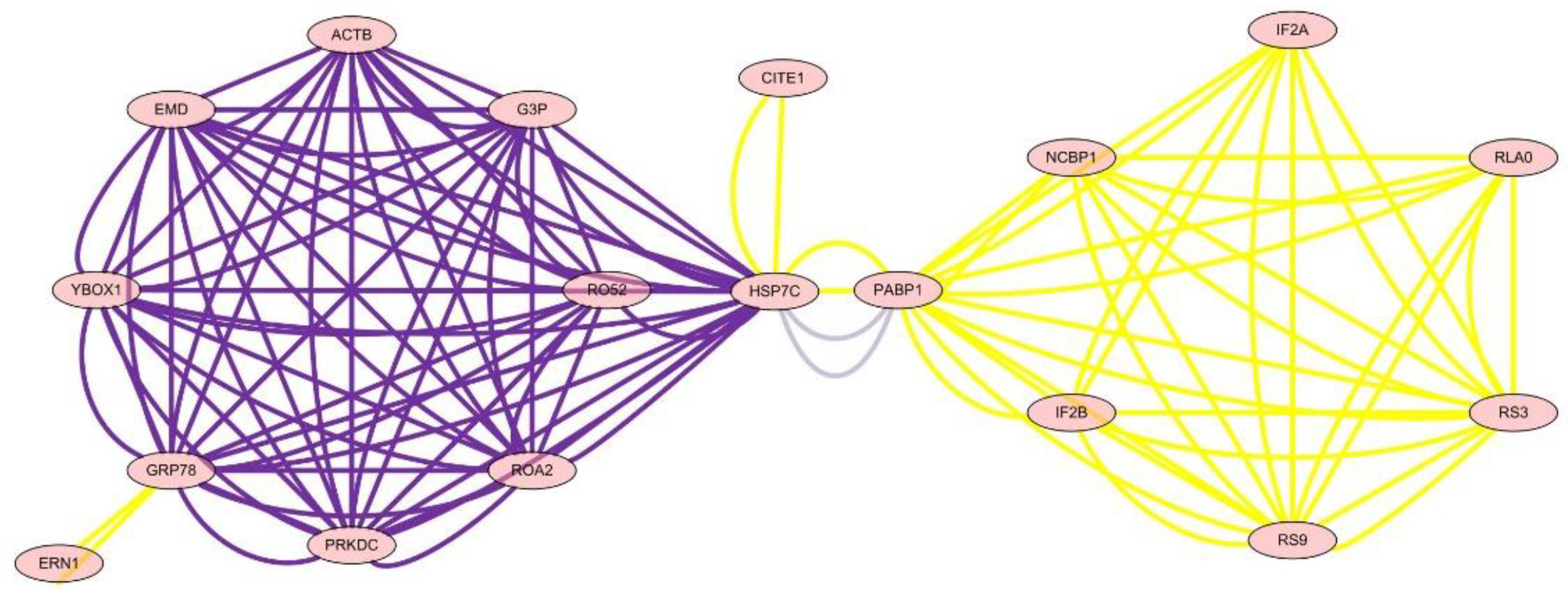
3.6. Analysis of the Network of Protein–Protein Interactions
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Than, N.N.; Newsome, P.N. A concise review of non-alcoholic fatty liver disease. Atherosclerosis 2015, 239, 192–202. [Google Scholar] [CrossRef] [Green Version]
- Zhou, Y.; Ding, Y.L.; Zhang, J.L.; Zhang, P.; Wang, J.Q.; Li, J.H. Alpinetin improved high fat diet-induced non-alcoholic fatty liver disease (NAFLD) through improving oxidative stress, inflammatory response and lipid metabolism. Biomed. Pharmacother. 2018, 97, 1397–1408. [Google Scholar] [CrossRef]
- Polin, D.; Wolford, J.H. Role of estrogen as a cause of fatty liver hemorrhagic syndrome. J. Nutr. 1977, 107, 873–886. [Google Scholar] [CrossRef]
- Miele, L.; Forgione, A.; Gasbarrini, G.; Grieco, A. Noninvasive assessment of fibrosis in non-alcoholic fatty liver disease (NAFLD) and non-alcoholic steatohepatitis (NASH). Transl. Res. 2007, 149, 114–125. [Google Scholar] [CrossRef] [PubMed]
- Bannister, D.W. The biochemistry of fatty liver and kidney syndrome. Biotin-mediated restoration of hepatic gluconeogenesis in vitro and its relationship to pyruvate carboxylase activity. Biochem. J. 1976, 156, 167–173. [Google Scholar] [CrossRef] [PubMed]
- Day, C.P.; James, O.F. Steatohepatitis: A tale of two “hits”? Gastroenterology 1998, 114, 842–845. [Google Scholar] [CrossRef]
- Byrne, C.D. Fatty liver: Role of inflammation and fatty acid nutrition. Prostaglandins Leukot. Essent. Fat. Acids 2010, 82, 265–271. [Google Scholar] [CrossRef]
- Gaté, L.; Paul, J.; Ba, G.N.; Tew, K.D.; Tapiero, H. Oxidative stress induced in pathologies: The role of antioxidants. Biomed. Pharm. 1999, 53, 169–180. [Google Scholar]
- Fromenty, B.; Robin, M.A.; Igoudjil, A.; Mansouri, A.; Pessayre, D. The ins and outs of mitochondrial dysfunction in NASH. Diabetes Metab. 2004, 30, 121–138. [Google Scholar] [CrossRef]
- Albano, E.; Mottaran, E.; Occhino, G.; Reale, E.; Vidali, M. Review article: Role of oxidative stress in the progression of non-alcoholic steatosis. Aliment. Pharm. Aliment. Pharm. 2005, 22, 71–73. [Google Scholar] [CrossRef]
- Andreadou, I.; Iliodromitis, E.K.; Mikros, E.; Bofilis, E.; Zoga, A.; Constantinou, M.; Tsantili-Kakoulidou, A.; Kremastinos, D.T. Melatonin does not prevent the protection of ischemic preconditioning in vivo despite its antioxidant effect against oxidative stress. Free Radic Biol. Med. 2004, 37, 500–510. [Google Scholar] [CrossRef] [PubMed]
- Wu, Q.; Yu, J.; Cui, H.; Li, Y. Role of essential oil oil of white Paederia scandens in endotoxin of Salmonella Enteritidis. Chin. Vet. Sci. 2012, 42, 758–764. [Google Scholar]
- Dhibi, M.; Brahmi, F.; Mnari, A.; Houas, Z.; Chargui, I.; Bchir, L.; Gazzah, N.; Alsaif, M.A.; Hammami, M. The intake of high fat diet with different trans fatty acid levels differentially induces oxidative stress and non alcoholic fatty liver disease (NAFLD) in rats. Nutr. Metab. 2011, 8, 65. [Google Scholar] [CrossRef] [PubMed]
- Das, S.K.; Vasudevan, D.M. Monitoring oxidative stress in patients with non-alcoholic and alcoholic liver diseases. Indian J. Clin. Biochem. 2005, 20, 24–28. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kusminski, C.M.; Shetty, S.; Orci, L.; Unger, R.H.; Scherer, P.E. Diabetes and apoptosis: Lipotoxicity. Apoptosis 2009, 14, 1484–1495. [Google Scholar] [CrossRef]
- Henson, P.M.; Bratton, D.L.; Fadok, V.A. The phosphatidylserine receptor: A crucial molecular switch? Nat. Rev. Mol. Cell Biol. 2001, 2, 627–633. [Google Scholar] [CrossRef]
- Landar, A.; Shiva, S.; Levonen, A.L.; Oh, J.Y.; Zaragoza, C.; Johnson, M.S.; Darley-Usmar, V.M. Induction of the permeability transition and cytochrome c release by 15-deoxy-Delta12, 14-prostaglandin J2 in mitochondria. Biochem. J. 2006, 394, 185–195. [Google Scholar] [CrossRef]
- Bailey, S.M.; Patel, V.B.; Young, T.A.; Asayama, K.; Cunningham, C.C. Chronic ethanol consumption alters the glutathione/glutathione peroxidase-1 system and protein oxidation status in rat liver. Alcohol. Clin. Exp. Res. 2001, 25, 726–733. [Google Scholar] [CrossRef]
- Polavarapu, R.; Spitz, D.R.; Sim, J.E.; Follansbee, M.H.; Oberley, L.W.; Rahemtulla, A.; Nanji, A.A. Increased lipid peroxidation and impaired antioxidant enzyme function is associated with pathological liver injury in experimental alcoholic liver disease in rats fed diets high in corn oil and fish oil. Hepatology 1998, 27, 1317–1323. [Google Scholar] [CrossRef]
- Chaudière, J.; Ferrari-Iliou, R. Intracellular antioxidants: From chemical to biochemical mechanisms. Food Chem. Toxicol. 1999, 37, 949–962. [Google Scholar] [CrossRef]
- García-Trevijano, E.R.; Iraburu, M.J.; Fontana, L.; Dominguez-Rosales, J.A.; Auster, A.S.; Covarrubias-Pinedo, A.; Rojkind, M. Transforming growth factor beta1 induces the expression of alpha1(I) procollagen mRNA by a hydrogen peroxide-C/EBPbeta-dependent mechanism in rat hepatic stellate cells. Hepatology 1999, 29, 960–970. [Google Scholar]
- Booth, L.; Roberts, J.L.; Dent, P. HSPA5/Dna K may be a useful target for human disease therapies. DNA Cell Biol. 2015, 34, 153–158. [Google Scholar] [CrossRef] [PubMed]
- Lee, W.S.; Sung, M.S.; Lee, E.G.; Yoo, H.G.; Cheon, Y.H.; Chae, H.J.; Yoo, W.H. A pathogenic role for ER stress-induced autophagy and ER chaperone GRP78/BiP in T lymphocyte systemic lupus erythematosus. J. Leukoc. Biol. 2015, 97, 425–433. [Google Scholar] [CrossRef] [PubMed]
- Attia, Y.A.; Bakhashwain, A.A.; Bertu, N.K. Thyme oil (Thyme vulgaris L.) as a natural growth promoter for broiler chickens reared under hot climate. Ital. J. Anim. Sci. 2017, 16, 275–282. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Ingredients (%) | High-Capacity Diets | Normal Diets |
|---|---|---|
| Corn | 53.00 | 62.48 |
| Soybean meal | - | - |
| Fish meal | 1.00 | 3.00 |
| Fried meal | 29.30 | 28.10 |
| Rapeseeds | - | 2.20 |
| Wheat bran | - | - |
| Cooked pigs’ oil | 12.95 | - |
| Additives | 0.75 | 0.75 |
| Lys | 0.12 | 0.09 |
| Met | 0.12 | 0.18 |
| CaHPO4 | 1.11 | 1.22 |
| Limestone meal | 1.52 | 1.82 |
| Salt | 0.13 | 0.16 |
| ME (MJ/kg) | 16.07 | 13.39 |
| CP (%) | 16.00 | 18.40 |
| Ca | 0.90 | 1.00 |
| AP | 0.35 | 0.45 |
| Lys | 1.00 | 1.10 |
| Met | 0.38 | 0.50 |
| No. | Ingredients | Raw Sample (mg/mL) |
|---|---|---|
| 1 | Linalool | 261.142 |
| 2 | L-α-terpineol | 10.126 |
| 3 | D-α-terpineol | 11.233 |
| 4 | Methyl salicylate | 78.902 |
| 5 | Camphor | 15.234 |
| 6 | Borneol | 118.784 |
| 7 | Eugenol | 24.634 |
| 8 | Isoeugenol | 21.346 |
| Time (Week) | Control Group | Model Group | Drug Group |
|---|---|---|---|
| 2 | 2.20 ± 0.15 | 3.02 ± 0.18 ** | 3.02 ± 0.18 ** |
| 3 | 2.22 ± 0.21 | 3.15 ± 0.11 ** | 2.83 ± 0.21 ** |
| 4 | 2.22 ± 0.22 | 3.05 ± 0.30 ** | 2.84 ± 0.17 |
| Item | Week | Control Group | Model Group | Drug Group |
|---|---|---|---|---|
| ROS (U/mg protein) | 1 | 88.56 ± 1.44 | 113.34 ± 4.06 ** | 97.64 ± 1.62 * |
| 2 | 88.56 ± 2.20 | 106.66 ± 4.48 ** | 98.33 ± 1.39 * | |
| 3 | 93.60 ± 3.55 | 107.49 ± 7.96 ** | 105.01 ± 6.20 * | |
| 4 | 100.69 ± 6.33 | 125.00 ± 5.86 ** | 109.08 ± 4.46 | |
| MDA (nmol/mg protein) | 1 | 6.57 ± 1.46 | 10.60 ± 1.34 ** | 9.04 ± 1.27 ** |
| 2 | 8.32 ± 1.63 | 10.51 ± 1.29 ** | 9.47 ± 1.56 ** | |
| 3 | 8.82 ± 1.26 | 13.47 ± 1.66 ** | 10.42 ± 1.79 ** | |
| 4 | 9.57 ± 1.18 | 13.46 ± 1.42 ** | 9.94 ± 1.03 | |
| SOD (U/mg protein) | 1 | 498.96 ± 35.83 | 461.32 ± 37.49 | 436.11 ± 43.69 * |
| 2 | 536.60 ± 31.67 | 501.19 ± 27.13 | 476.48 ± 26.99 ** | |
| 3 | 564.89 ± 26.22 | 492.36 ± 30.23 ** | 425.99 ± 26.58 ** | |
| 4 | 523.83 ± 29.05 | 443.22 ± 33.35 ** | 417.21 ± 30.79 ** |
| SSP | Protein | Gene | MW/PI | Trend |
|---|---|---|---|---|
| 1101 | Uncharacterized protein | - | - | - |
| 1203 | Regucalcin | Rgn | 33/5.3 | ↓ |
| 3704 | Heat shock 70 kDa protein 5 (glucose-regulated protein, 78 kDa) | Hspa5 | 72/5.1 | ↑ |
| 4103 | ATP synthase subunit d | Atp5h | 19/6.2 | ↓ |
| 4301 | Guanine nucleotide-binding protein G(I)/G(S)/G(T) subunit beta 1 | Gnb1 | 35/7.6 | ↓ |
| 4401 | S-adenosylmethionine synthase isoform type-1 | MAT1A | 44/5.6 | ↓ |
| 4402 | Fumarylacetoacetase | Fah | 46/6.7 | ↓ |
| 5201 | Carbonic anhydrase 3 | Car3 | 29/6.9 | ↓ |
| 5301 | 3-alpha-hydroxysteroid dehydrogenase | Akr1c2 | 37/6.7 | ↑ |
| 5603 | 3-hydroxy-3-methylglutaryl-Coenzyme A synthase 2 | Hmgcs2 | 57/8.9 | ↑ |
| 5702 | Catalase | Cat | 60/7.1 | ↑ |
| 6104 | Glutathione S-transferase Mu 1 | Gstm1 | 26/8.3 | ↓ |
| 6502 | Arginosuccinate synthase 1 | Ass1 | 46/7.6 | ↓ |
| 6503 | Long-chain specific acyl-CoA dehydrogenase | Acadl | 48/7.6 | ↑ |
| 6702 | Heat shock 70 kDa protein 8 | Hspa8 | 71/5.4 | ↑ |
| 6901 | Uncharacterized protein | - | - | - |
| 7003 | Beta-actin | actb | 15/5.7 | ↑ |
| 7303 | Uncharacterized protein | Ote | 39/8.9 | |
| 8301 | Electron transfer flavoprotein subunit alpha | Etfa | 35/8.6 | ↓ |
| 8302 | Glutathione S-transferase Mu 2 | Gstm2 | 28/8.2 | ↓ |
| 9301 | Glyceraldehyde-3-phosphate dehydrogenase-like | Gapdh | 36/8.4 | ↓ |
| 9501 | Betaine-homocysteine S-methyltransferase 1 | Bhmt | 45/8.0 | ↓ |
| 8603 | Uncharacterized protein | - | - | - |
| ID | Degree | Label | Gene | Name |
|---|---|---|---|---|
| 146490 | 22 | HSP7C | HSPA8 | Heat shock cognate 71 kDa protein |
| 145948 | 18 | GRP78 | HSPA5 | 78 kDa glucose-regulated protein |
| 93344 | 16 | PABP1 | PABPC1 | Emerin |
| 71781 | 16 | YBOX1 | YBX1 | Nuclease-sensitive element-binding protein 1 |
| 57072 | 16 | PRKDC | PRKDC | Heterogeneous nuclear ribonucleoproteins A2/B1 |
| 51704 | 16 | ACTB | ACTB | Polyadenylate-binding protein 1 |
| 50923 | 16 | RO52 | TRIM21 | E3 ubiquitin-protein ligase TRIM21 |
| 50095 | 16 | ROA2 | HNRNPA2B1 | DNA-dependent protein kinase catalytic subunit |
| 15490 | 16 | EMD | EMD | Actin, cytoplasmic 1 |
| 150257 | 16 | G3P | GAPDH | Glyceraldehyde-3-phosphate dehydrogenase |
| 89014 | 12 | RS3 | RPS3 | 40S ribosomal protein S9 |
| 88619 | 12 | RS9 | RPS9 | 40S ribosomal protein S3 |
| 5116 | 8 | IF2B | EIF2S2 | Eukaryotic translation initiation factor 2 subunit 2 |
| 50028 | 8 | RLA0 | RPLP0 | 60S acidic ribosomal protein P0 |
| 4817 | 8 | IF2A | EIF2S1 | Eukaryotic translation initiation factor 2 subunit 1 |
| 117387 | 8 | NCBP1 | NCBP1 | Nuclear cap-binding protein subunit 1 |
| 95707 | 2 | CITE1 | CITED1 | Cbp/p300-interacting trans-activator 1 |
| 15584 | 2 | ERN1 | ERN1 | Serine/threonine-protein kinase/endoribonuclease IRE1 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wu, Q.; Tang, H.; Wang, H. The Anti-Oxidation and Mechanism of Essential Oil of Paederia scandens in the NAFLD Model of Chicken. Animals 2019, 9, 850. https://doi.org/10.3390/ani9100850
Wu Q, Tang H, Wang H. The Anti-Oxidation and Mechanism of Essential Oil of Paederia scandens in the NAFLD Model of Chicken. Animals. 2019; 9(10):850. https://doi.org/10.3390/ani9100850
Chicago/Turabian StyleWu, Qiang, Huaqiao Tang, and Hongbin Wang. 2019. "The Anti-Oxidation and Mechanism of Essential Oil of Paederia scandens in the NAFLD Model of Chicken" Animals 9, no. 10: 850. https://doi.org/10.3390/ani9100850
APA StyleWu, Q., Tang, H., & Wang, H. (2019). The Anti-Oxidation and Mechanism of Essential Oil of Paederia scandens in the NAFLD Model of Chicken. Animals, 9(10), 850. https://doi.org/10.3390/ani9100850