Dietary Inositol Reduces Fearfulness and Avoidance in Laying Hens

 , and
, and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Experimental Treatments
2.2. Birds and Bird Housing
2.3. Data Collection
2.4. Chemical Analysis
2.5. Statistical Analyses
3. Results and Discussion
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Leske, K.; Coon, C. A bioassay to determine the effect of phytase on phytate phosphorus hydrolysis and total phosphorus retention of feed ingredients as determined with broilers and laying hens. Poult. Sci. 1999, 78, 1151–1157. [Google Scholar] [CrossRef] [PubMed]
- Nelson, T.S. The utilization of phytate phosphorus by poultry—A review. Poult. Sci. 1967, 46, 862–871. [Google Scholar] [CrossRef] [PubMed]
- Dersjant-Li, Y.; Awati, A.; Schulze, H.; Partridge, G. Phytase in non-ruminant animal nutrition: A critical review on phytase activities in the gastrointestinal tract and influencing factors. J. Sci. Food Agric. 2015, 95, 878–896. [Google Scholar] [CrossRef]
- Walk, C.L.; Bedford, M.R.; Santos, T.S.; Paiva, D.; Bradley, J.R.; Wladecki, H.; Honaker, C.; McElroy, A.P. Extra-phosphoric effects of superdoses of a novel microbial phytase. Poult. Sci. 2013, 92, 719–725. [Google Scholar] [CrossRef] [PubMed]
- Walk, C.L.; Santos, T.T.; Bedford, M.R. Influence of superdoses of a novel microbial phytase on growth performance, tibia ash, and gizzard phytate and inositol in young broilers. Poult. Sci. 2014, 93, 1172–1177. [Google Scholar] [CrossRef] [PubMed]
- Walk, C.L.; Bedford, M.R.; Olukosi, O.A. Effect of phytase on growth performance, phytate degradation and gene expression of myo-inositol transporters in the small intestine, liver and kidney of 21 day old broilers. Poult. Sci. 2018, 97, 1155–1162. [Google Scholar] [CrossRef]
- Lee, S.A.; Bedford, M.R. Inositol—An effective growth promotor? Worlds. Poult. Sci. J. 2016, 72, 743–760. [Google Scholar] [CrossRef]
- Leybaert, L.; Paemeleire, K.; Strahonja, A.; Sanderson, M.J. Inositol-trisphosphate-dependent intercellular calcium signaling in and between astrocytes and endothelial cells. Glia 1998, 24, 398–407. [Google Scholar] [CrossRef]
- Croze, M.L.; Soulage, C.O. Potential role and therapeutic interests of myo-inositol in metabolic diseases. Biochimie 2013, 95, 1811–1827. [Google Scholar] [CrossRef]
- Wei, H.; Landgraf, D.; Wang, G.; McCarthy, M.J. Inositol polyphosphates contribute to cellular circadian rhythms: Implications for understanding lithium’s molecular mechanism. Cell. Signal. 2018, 44, 82–91. [Google Scholar] [CrossRef]
- Schneider, S. Inositol transport proteins. FEBS Lett. 2015, 589, 1049–1058. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Priya, S.S.; Bhyvaneswari, K.; Patwari, P. A study on the effects of scyllo-inositol on learning and memory in senile swiss albino mice. Int. J. Pharm. Biol. Arch. 2013, 4, 868–872. [Google Scholar]
- Einat, H.; Belmaker, R.H. The effects of inositol treatment in animal models of psychiatric disorders. J. Affect. Disord. 2001, 62, 113–121. [Google Scholar] [CrossRef]
- Fisher, S.K.; Novak, J.E.; Agranoff, B.W. Inositol and higher inositol phosphates in neural tissues: Homeostasis, metabolism and functional significance. J. Neurochem. 2002, 82, 736–754. [Google Scholar] [CrossRef] [PubMed]
- Shirayama, Y.; Takahashi, M.; Osone, F.; Hara, A.; Okubo, T. Myo-inositol, glutamate, and glutamine in the prefrontal cortex, hippocampus, and amygdala in major depression. Biol. Psychiatry Cogn. Neurosci. Neuroimaging 2017, 2, 196–204. [Google Scholar] [CrossRef] [PubMed]
- Tumati, S.; Opmeer, E.M.; Marsman, J.B.C.; Martens, S.; Reesink, F.E.; De Deyn, P.P.; Aleman, A. Lower choline and myo-inositol in temporo-parietal cortex is associated with apathy in amnestic MCI. Front. Aging Neurosci. 2018, 10, 1–9. [Google Scholar] [CrossRef]
- Huber, K.; Kenez, A.; Rodehutscord, M. Dietary myo-inositol enhances serotonin and dopamine concentrations in plasma of 21-day-old broilers. In Proceedings of the Society of Nutrition Physiology; DLG-Verlag: Frankfurt am Main, Germany, 2017; Volume 26, p. 100. [Google Scholar]
- Narvaes, R.; Martins de Almeida, R.M. Aggressive behavior and three neurotransmitters: Dopamine, GABA, and serotonin—A review of the last 10 years. Psychol. Neurosci. 2014, 7, 601–607. [Google Scholar] [CrossRef]
- van Hierden, Y.M.; Korte, S.M.; Ruesink, E.W.; van Reenen, C.G.; Engel, B.; Korte-Bouws, G.A.H.; Koolhaas, J.M.; Blokhuis, H.J. Adrenocortical reactivity and central serotonin and dopamine turnover in young chicks from a high and low feather-pecking line of laying hens. Physiol. Behav. 2002, 75, 653–659. [Google Scholar] [CrossRef] [Green Version]
- van Hierden, Y.M.; de Boer, S.F.; Koolhaas, J.M.; Korte, S.M. The control of feather pecking by serotonin. Behav. Neurosci. 2004, 118, 575–583. [Google Scholar] [CrossRef]
- Dennis, R.L.; Fahey, A.G.; Cheng, H.W. Alterations to Embryonic Serotonin Change Aggression and Fearfulness. Aggress. Behav. 2013, 39, 91–98. [Google Scholar] [CrossRef]
- Dennis, R.L.; Muir, W.M.; Cheng, H.W. Effects of raclopride on aggression and stress in diversely selected chicken lines. Behav. Brain Res. 2006, 175, 104–111. [Google Scholar] [CrossRef] [PubMed]
- Bubier, N.E. The behavioural priorities of laying hens: The effects of two methods of environment enrichment on time budgets. Behav. Processes 1996, 37, 239–249. [Google Scholar] [CrossRef]
- van Krimpen, M.M.; Kwakkel, R.P.; Reuvekamp, B.F.J.; van der Peet-Schwering, C.M.C.; Den Hartog, L.A.; Verstegen, M.W.A. Impact of feeding management on feather pecking in laying hens. Worlds Poult. Sci. J. 2005, 61, 663–685. [Google Scholar] [CrossRef]
- Kumar, D.; Raginski, C.; Schwean-Lardner, K.V.; Classen, H.L. Assessing the performance response of laying hens to intake levels of digestible balanced protein from 27 to 66 weeks of age. Can. J. Anim. Sci. 2018, 98, 801–808. [Google Scholar] [CrossRef]
- Canadian Council on Animal Care. Guide to the Care and Use of Experimental Animals; Olfert, E.D., Cross, B.M., McWilliams, A.A., Eds.; Canadian Council on Animal Care: Ottawa, ON, Canada, 1993; Volume 1. [Google Scholar]
- Canadian Council on Animal Care CCAC guidelines on: The care and use of farm animals in research, teaching and testing. Available online: http//www.ccac.ca (accessed on 26 September 2011).
- Aviagen Broiler Ross 308 Performance Objectives. Available online: http//eu.aviagen.com/assets/Tech_Center/Ross_Broiler/Ross-308-Broiler-PO-2014-EN.pdf (accessed on 20 October 2014).
- Herwig, E.; Schwean-Lardner, K.V.; Walk, C.; Van Kessel, A.G.; Bedford, M.R.; Classen, H.L. Effect of phytase and myo-inositol supplementation on the expression of inositol transporters in the small intestine of laying hens, and blood and yolk inositol concentrations. In Proceedings of the Poultry Science Association 108th Annual Meeting, Montreal, QC, Canada, 15–18 July 2019; U.S. Animal Health Association: Richmond, VA, USA, 2019; p. 194. [Google Scholar]
- van Liere, D.W.; Aggrey, S.E.; Brouns, F.M.R.; Wiepkema, P.R. Oiling behaviour and the effect of lipids on dustbathing behaviour in laying hens Gallus gallus domesticus. Behav. Processes 1991, 24, 71–81. [Google Scholar] [CrossRef]
- Savory, C.J. Feather pecking and cannibalism. Worlds. Poult. Sci. J. 1995, 51, 215–219. [Google Scholar] [CrossRef]
- Gabrush, T. Effects of the degree of beak trimming on the performance of White Leghorns. MSc Diss., University of Saskatchewan, Saskatoon, SK, Canada, 2011. [Google Scholar]
- Hunniford, M.E.; Widowski, T.M. Nest alternatives: Adding a wire partition to the scratch area affects nest use and nesting behaviour of laying hens in furnished cages. Appl. Anim. Behav. Sci. 2017, 186, 29–34. [Google Scholar] [CrossRef]
- Davami, A.; Wineland, M.J.; Jones, W.T.; Ilardi, R.L.; Peterson, R.A. Effects of population size, floor space, and feeder space upon productive performance, external appearance, and plasma corticosterone concentration of laying hens. Poult. Sci. 1987, 66, 251–257. [Google Scholar] [CrossRef]
- Ali, A.; Cheng, K.M. Early egg production in genetically blind (rc/rc) chickens in comparison with sighted (Rc+/rc) controls. Poult. Sci. 1985, 64, 789–794. [Google Scholar] [CrossRef]
- Welfare Quality Consortium. Welfare Quality Assessment Protocol for Poultry (broilers, laying hens); Welfare Quality Consortium: Lelustad, The Netherlands, 2009. [Google Scholar]
- AOAC. Official Methods of Analysis, 18th ed.; Association of Official Analytical Chemists: Arlington, VA, USA, 2006. [Google Scholar]
- Laird, S.; Kühn, I.; Wilcock, P.; Miller, H.M. The effects of phytase on grower pig growth performance and ileal inositol phosphate degradation. J. Anim. Sci. 2016, 94, 142–145. [Google Scholar] [CrossRef]
- Pagel, M.; Dawkins, M.S. Peck orders and group size in laying hens: “Futures contracts” for non-aggression. Behav. Processes 1997, 40, 13–25. [Google Scholar] [CrossRef]
- D’Eath, R.B.; Keeling, L.J. Social discrimination and aggression by laying hens in large groups: From peck orders to social tolerance. Appl. Anim. Behav. Sci. 2003, 84, 197–212. [Google Scholar] [CrossRef]
- Anderson, K.E.; Davis, G.S.; Jenkins, P.K.; Carroll, A.S. Effects of bird age, density, and molt on behavioral profiles of two commercial layer strains in cages. Poult. Sci. 2003, 83, 15–23. [Google Scholar] [CrossRef] [PubMed]
- Lohmann Tierzucht Layers Lohmann LSL-Lite Management Guide. Available online: http://www.hylinena.com/UserDocs/products/Lohmann (accessed on 6 November 2018).
- Nicol, C. Behaviour requirements within a cage environment. Worlds. Poult. Sci. J. 1990, 46, 31–33. [Google Scholar] [CrossRef]
- Albentosa, M.J.; Cooper, J.J.; Luddem, T.; Redgate, S.E.; Elson, H.A.; Walker, A.W. Evaluation of the effects of cage height and stocking density on the behaviour of laying hens in furnished cages. Br. Poult. Sci. 2007, 48, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Duncan, I.J.H. Behavior and behavioral needs. Poult. Sci. 1998, 77, 1766–1772. [Google Scholar] [CrossRef] [PubMed]
- Kofman, O.; Agam, G.; Shapiro, J.; Spencer, A. Chronic dietary inositol enhances locomotor activity and brain inositol levels in rats. Psychopharmacology (Berl.) 1998, 139, 239–242. [Google Scholar] [CrossRef]
- Einat, H.; Karbovski, H.; Korik, J.; Tsalah, D.; Belmaker, R.H. Inositol reduces depressive-like behaviors in two different animal models of depression. Psychopharmacology (Berl.) 1999, 144, 158–162. [Google Scholar] [CrossRef]
- Einat, H.; Levine, J.; Grauer, E.; Belmaker, R. Effect of inositol treatment on the behavior of macaque monkeys. Prog. Neuro-Psychopharmacol. Biol. Psychiat. 1998, 22, 999–1005. [Google Scholar] [CrossRef]
- Harvey, B.H.; Scheepers, A.; Brand, L.; Stein, D.J. Chronic inositol increases striatal D2 receptors but does not modify dexamphetamine-induced motor behavior. Pharmacol. Biochem. Behav. 2001, 68, 245–253. [Google Scholar] [CrossRef]
- Martinsen, E.W. Physical activity and depression: Clinical experience. Acta Psychiatr. Scand. 1994, 377, 23–27. [Google Scholar] [CrossRef] [PubMed]
- Etches, R.J.; Williams, J.B.; Rzasa, J. Effects of corticosterone and dietary changes in the hen on ovarian function, plasma LH and steroids and the response to exogenous LH-RH. Reproduction 1984, 70, 121–130. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Astheimer, L.B.; Buttemer, W.A.; Wingfield, J.C.; Astheimer, L.B.; Buttemer, W.A.; Wingfield, J.C. Interactions of corticosterone with feeding, activity and metabolism in passerine birds. Ornis Scand. 1992, 23, 355–365. [Google Scholar] [CrossRef]
- Webster, A.B. Physiology and behavior of the hen during induced molt. Poult. Sci. 2003, 82, 992–1002. [Google Scholar] [CrossRef]
- Park, J.; Longo, F.; Park, S.J.; Lee, S.; Bae, M.; Tyagi, R.; Han, J.-H.; Kim, S.; Santini, E.; Klann, E.; et al. Inositol polyphosphate multikinase mediates extinction of fear memory. Proc. Natl. Acad. Sci. USA 2019, 116, 2707–2712. [Google Scholar] [CrossRef] [Green Version]
- Gross, W.B.; Siegel, H.S. Evaluation of the heterophil/lymphocyte ratio as a measure of stress in chickens. Avian Dis. 1983, 27, 972–979. [Google Scholar] [CrossRef]
- Maxwell, M.H.; Robertson, G.W. The avian heterophil leucocyte: A review. Worlds. Poult. Sci. J. 1998, 54, 155–178. [Google Scholar] [CrossRef]
- Savory, C.J.; Mann, J.S. Behavioural development in groups of pen-housed pullets in relation to genetic strain, age and food form. Br. Poult. Sci. 1997, 38, 38–47. [Google Scholar] [CrossRef]
- Jones, R.B.; Hocking, P.M. Genetic selection for poultry behaviour: Big bad wolf or friend in need? Anim. Welf. 1999, 8, 343–359. [Google Scholar]
- Guesdon, V.; Ahmed, A.M.H.; Mallet, S.; Faure, J.M.; Nys, Y. Effects of beak trimming and cage design on laying hen performance and egg quality. Br. Poult. Sci. 2006, 47, 1–12. [Google Scholar] [CrossRef]
- Bach, D.R.; Dayan, P. Algorithms for survival: A comparative perspective on emotions. Nat. Rev. Neurosci. 2017, 18, 311–319. [Google Scholar] [CrossRef] [PubMed]
- Sajja, V.S.S.S.; Perrine, S.A.; Ghoddoussi, F.; Hall, C.S.; Galloway, M.P.; VandeVord, P.J. Blast neurotrauma impairs working memory and disrupts prefrontal myo-inositol levels in rats. Mol. Cell. Neurosci. 2014, 59, 119–126. [Google Scholar] [CrossRef] [PubMed]
- Sheth, C.; Prescot, A.; Bueler, E.; DiMuzio, J.; Legarreta, M.; Renshaw, P.F.; Yurgelun-Todd, D.; McGlade, E. Alterations in anterior cingulate cortex myoinositol and aggression in veterans with suicidal behavior: A proton magnetic resonance spectroscopy study. Psychiatry Res. Neuroimaging 2018, 276, 24–32. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Ingredients (%) | 19–27 Weeks of Age | 28–59 Weeks of Age | ||
|---|---|---|---|---|
| HBP | RBP | HBP | RBP | |
| Wheat | 53.37 | 63.32 | 58.82 | 67.37 |
| Soybean meal | 18.81 | 9.88 | 14.39 | 6.72 |
| Canola meal | 10.00 | 10.00 | 10.00 | 10.00 |
| Wheat bran | 2.00 | 2.00 | 2.00 | 2.00 |
| Canola oil | 4.54 | 3.57 | 3.88 | 3.05 |
| Limestone | 9.27 | 9.31 | 9.22 | 9.26 |
| Mono-dicalcium phosphate | 0.77 | 0.77 | 0.52 | 0.52 |
| Sodium chloride | 0.33 | 0.32 | 0.30 | 0.30 |
| Vitamin-mineral premix 1 | 0.50 | 0.50 | 0.50 | 0.50 |
| Choline chloride | 0.10 | 0.10 | 0.10 | 0.10 |
| Rovimix Hy-D 62.5 2 | 0.04 | 0.04 | 0.04 | 0.04 |
| L-Lysine HCl | 0.01 | 0.05 | 0.03 | 0.06 |
| DL-Methionine | 0.20 | 0.10 | 0.15 | 0.06 |
| L-Threonine | 0.05 | 0.03 | 0.04 | 0.02 |
| Xylanase 3 | 0.01 | 0.01 | 0.01 | 0.01 |
| Calculated composition (as is) | ||||
| Apparent metabolizable energy (kcal/kg) | 2800 | 2800 | 2800 | 2800 |
| Crude Protein | 19.92 | 16.98 | 18.54 | 16.00 |
| Digestible lysine | 0.84 | 0.67 | 0.76 | 0.61 |
| Digestible methionine | 0.48 | 0.34 | 0.41 | 0.29 |
| Digestible methionine and cysteine | 0.79 | 0.63 | 0.71 | 0.57 |
| Digestible threonine | 0.77 | 0.52 | 0.59 | 0.47 |
| Calcium | 3.70 | 3.70 | 3.63 | 3.63 |
| Phytate | 1.02 | 0.97 | 1.00 | 0.96 |
| Non-phytate phosphorus | 0.35 | 0.35 | 0.30 | 0.30 |
| Category | Behaviour | Description 1 |
|---|---|---|
| Aggression | Aggressive pecking | Forceful pecking directed to another bird’s head, legs or vent areas. |
| Severe feather pecking | Pecking at the plumage of a cage-mate with the intention of removing feathers. Recipient hen can squawk and withdraw. | |
| Fighting | More than two aggressive pecks between two or more hens. | |
| Comfort | Wing or leg stretching | Extension of wing or leg on one side of the body. |
| Other | Flapping, tail wagging, etc. | |
| Displacement | Displacement | Hen stops another hen’s behaviour by taking her location in the cage. |
| Investigative | Gentle feather pecking | Pecking at the plumage of a cage-mate with little or no damage to it. Feathers are not removed and it is usually ignored by the recipient. |
| Object pecking | Pecking at anything other than feed, water, or cage-mate. | |
| Maintenance | Preening | Self-manipulation of feathers on the body using the beak or self-scratching. |
| Sham-dustbathing | Side or head-rubbing, vertical wing shaking and side laying with scratching. | |
| Ruffling | Fluffing up the feathers. | |
| Nutritive | Feeding | Head extended into the feeder, manipulating or ingesting feed. |
| Drinking | Head extended to the water line, manipulating the water nipple. | |
| Sitting | Sitting/Sleeping/Laying | A bird sitting with breast on the floor. |
| Standing | Standing | Hen standing on both legs without doing any other particular action. |
| Stereotypy | Pacing | Hen restless and walking back and forward without another purpose. |
| Walking | Walking | Any hen taking more than two steps in a direction and not pacing. |
| Behaviour | Diets | SEM | ANOVA p Values | Contrasts p Values | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| HBP | HBP-I | RBP | RBP-I | RBP-P | Diet (D) | Time (T) | D × T | Protein | HBP vs. HBP-I | Inositol | RBP-I vs. RBP-P | ||
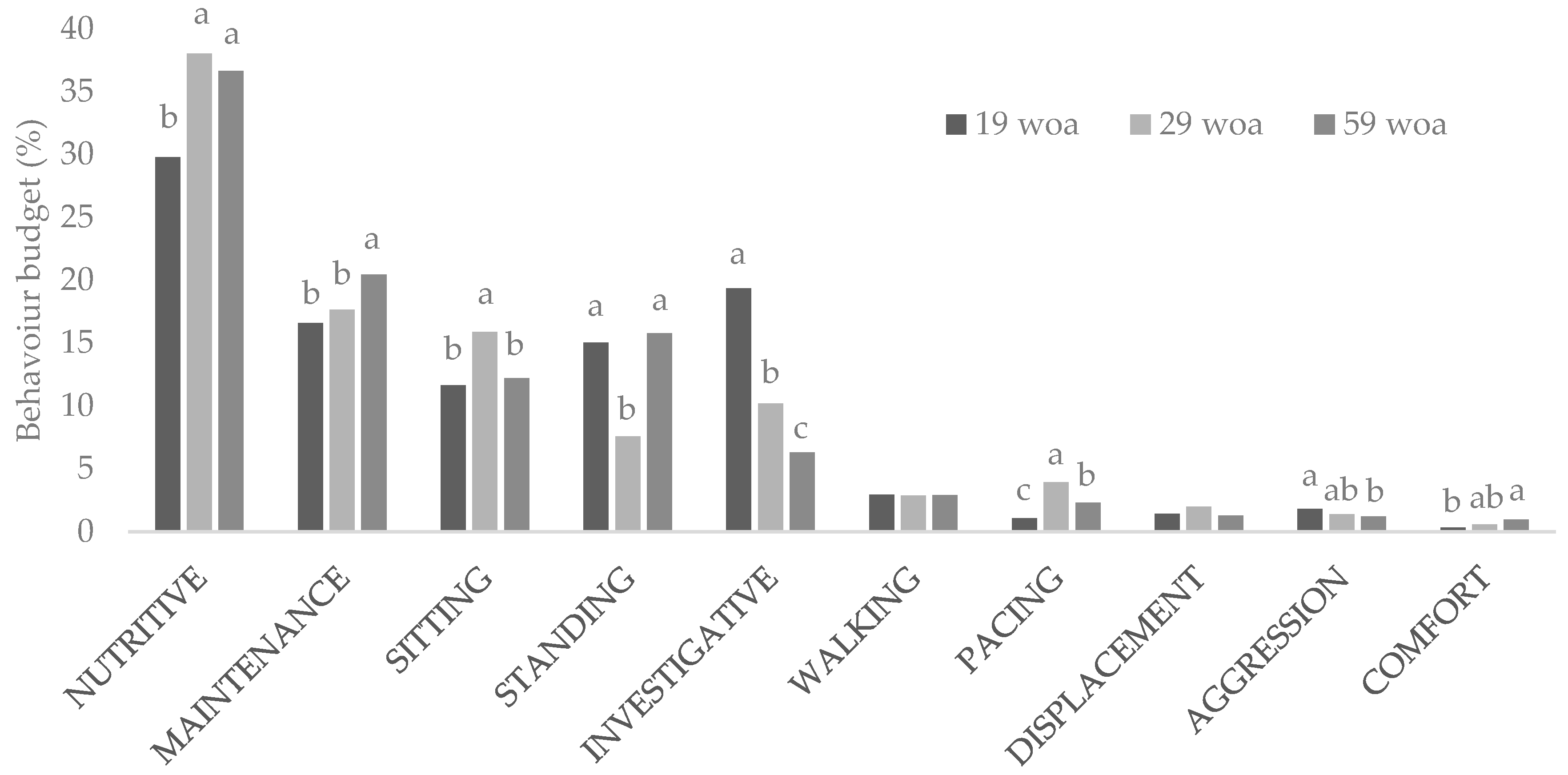
| Nutritive | 34.45 | 36.10 | 32.96 | 34.42 | 36.16 | 0.611 | 0.201 | <0.001 | 0.685 | 0.147 | 0.284 | 0.154 | 0.260 |
| Maintenance | 17.96 | 17.45 | 19.09 | 19.34 | 17.28 | 0.441 | 0.417 | 0.001 | 0.939 | 0.115 | 0.703 | 0.889 | 0.128 |
| Sitting | 12.08 | 12.01 | 14.37 | 14.65 | 13.13 | 0.413 | 0.062 | <0.001 | 0.786 | 0.003 | 0.950 | 0.899 | 0.189 |
| Standing | 13.93 | 11.67 | 12.73 | 12.58 | 13.05 | 0.537 | 0.452 | <0.001 | 0.881 | 0.864 | 0.063 | 0.159 | 0.697 |
| Investigative | 12.97 | 11.96 | 11.99 | 11.70 | 11.06 | 0.650 | 0.464 | <0.001 | 0.294 | 0.440 | 0.138 | 0.165 | 0.948 |
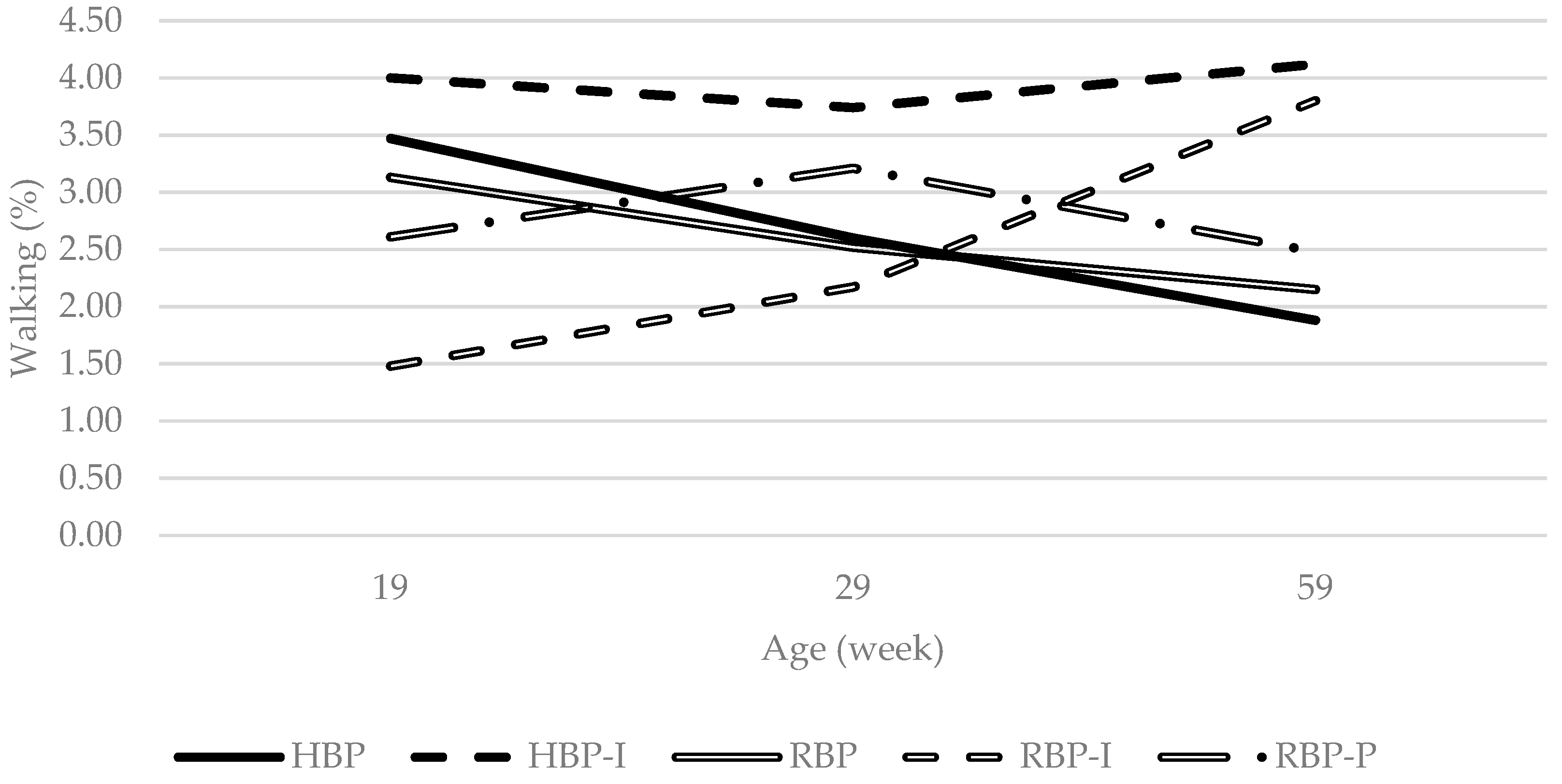
| Walking | 2.65 b | 3.95 a | 2.60 b | 2.48 b | 2.77 a,b | 0.155 | 0.010 | 0.969 | 0.050 | 0.019 | 0.005 | 0.066 | 0.528 |
| Pacing | 2.46 a,b | 2.96 a | 2.47 a,b | 1.67 b | 2.56 a,b | 0.184 | 0.033 | <0.001 | 0.782 | 0.021 | 0.384 | 0.389 | 0.022 |
| Displacement | 1.37 | 1.84 | 1.51 | 1.39 | 1.72 | 0.118 | 0.592 | 0.080 | 0.640 | 0.420 | 0.148 | 0.356 | 0.703 |
| Aggression | 1.38 | 1.54 | 1.60 | 1.29 | 1.54 | 0.090 | 0.685 | 0.010 | 0.205 | 0.902 | 0.646 | 0.514 | 0.311 |
| Comfort | 0.74 | 0.52 | 0.67 | 0.48 | 0.74 | 0.069 | 0.487 | <0.001 | 0.753 | 0.739 | 0.234 | 0.097 | 0.202 |
| Feather Coverage | Diets | SEM | ANOVA p Value | Contrasts p Values | |||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| HBP | HBP-I | RBP | RBP-I | RBP-P | Protein | HBP vs. HBP-I | Inositol | RBP-P vs. RBP-I | |||
| Neck | 2.48 | 2.44 | 2.31 | 2.12 | 2.33 | 0.072 | 0.595 | 0.151 | 0.893 | 0.521 | 0.380 |
| Wings | 3.43 | 3.43 | 3.33 | 3.45 | 3.47 | 0.064 | 0.972 | 0.761 | 1.000 | 0.704 | 0.921 |
| Back | 3.36 | 3.29 | 3.14 | 3.25 | 3.27 | 0.087 | 0.961 | 0.535 | 0.818 | 0.915 | 0.955 |
| Vent | 2.52 | 2.01 | 2.55 | 2.12 | 2.29 | 0.097 | 0.323 | 0.740 | 0.108 | 0.038 | 0.575 |
| Breast | 2.70 | 2.57 | 2.76 | 2.54 | 2.56 | 0.074 | 0.875 | 0.914 | 0.595 | 0.334 | 0.952 |
| Total | 14.48 | 13.74 | 14.08 | 13.49 | 14.97 | 0.338 | 0.580 | 0.624 | 0.438 | 0.323 | 0.143 |
| Lesions | |||||||||||
| Skin | 0.12 | 0.13 | 0.07 | 0.15 | 0.12 | 0.024 | 0.895 | 0.873 | 0.902 | 0.427 | 0.666 |
| Comb | 2.04 | 2.00 | 2.22 | 1.88 | 2.10 | 0.056 | 0.451 | 0.808 | 0.846 | 0.155 | 0.241 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Herwig, E.; Classen, H.L.; Walk, C.L.; Bedford, M.; Schwean-Lardner, K. Dietary Inositol Reduces Fearfulness and Avoidance in Laying Hens. Animals 2019, 9, 938. https://doi.org/10.3390/ani9110938
Herwig E, Classen HL, Walk CL, Bedford M, Schwean-Lardner K. Dietary Inositol Reduces Fearfulness and Avoidance in Laying Hens. Animals. 2019; 9(11):938. https://doi.org/10.3390/ani9110938
Chicago/Turabian StyleHerwig, Eugenia, Henry L. Classen, Carrie L. Walk, Mike Bedford, and Karen Schwean-Lardner. 2019. "Dietary Inositol Reduces Fearfulness and Avoidance in Laying Hens" Animals 9, no. 11: 938. https://doi.org/10.3390/ani9110938
APA StyleHerwig, E., Classen, H. L., Walk, C. L., Bedford, M., & Schwean-Lardner, K. (2019). Dietary Inositol Reduces Fearfulness and Avoidance in Laying Hens. Animals, 9(11), 938. https://doi.org/10.3390/ani9110938