The Relationship between S. aureus and Branched-Chain Amino Acids Content in Composite Cow Milk

 , , , , , , and
, , , , , , and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Milk Sampling
2.2. Somatic Cells Count
2.3. S. Aureus Identification
2.4. Bacterial Strain and Artificial Specimen Preparation for Baseline Contamination
2.5. DNA Extraction
2.6. Oligonucleotide Primers and DNA Amplification
2.7. HPLC Measurements and Extraction of Free Amino Acids from the Raw Milk Sample
2.8. Statistical Methods
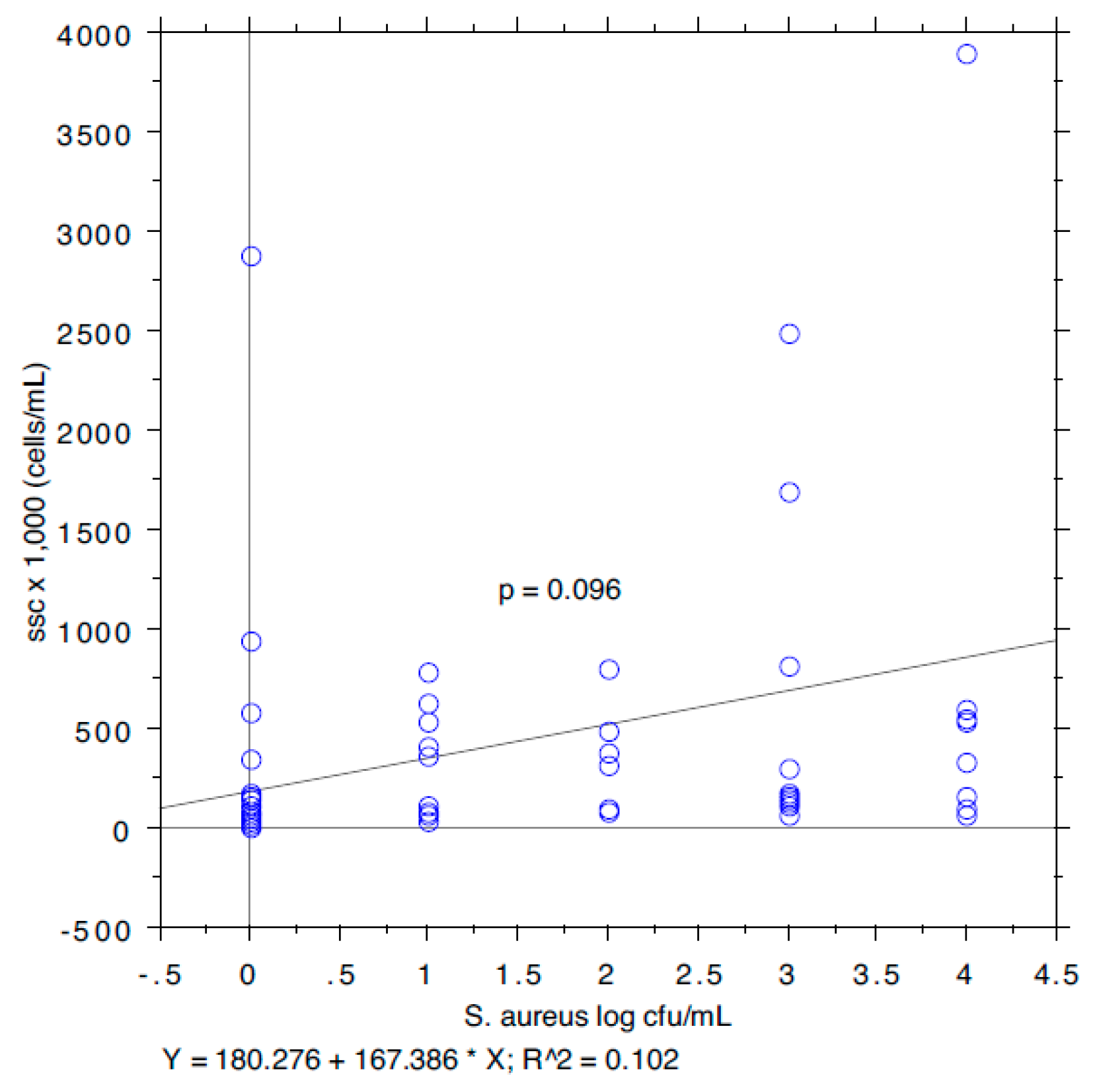
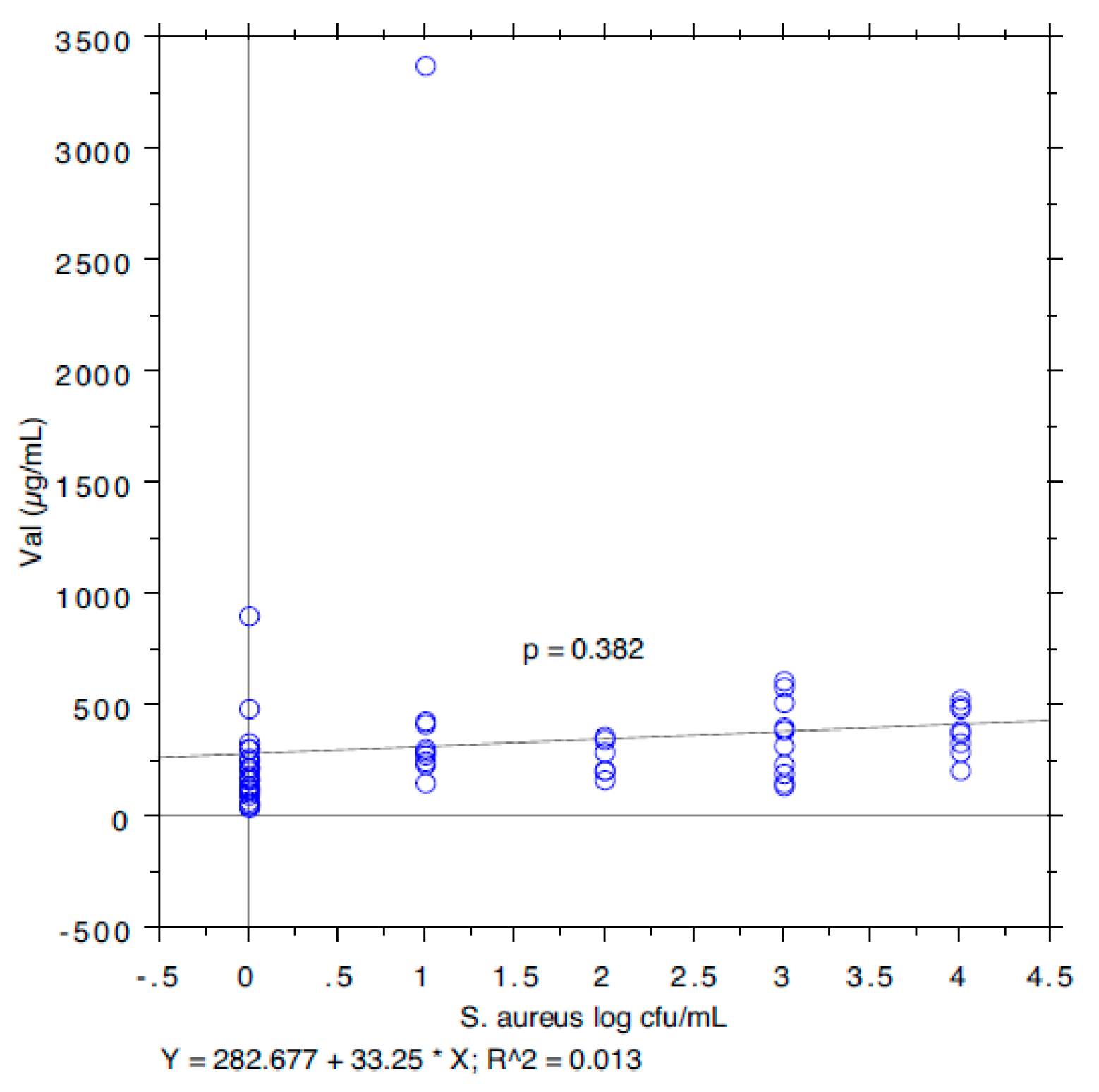
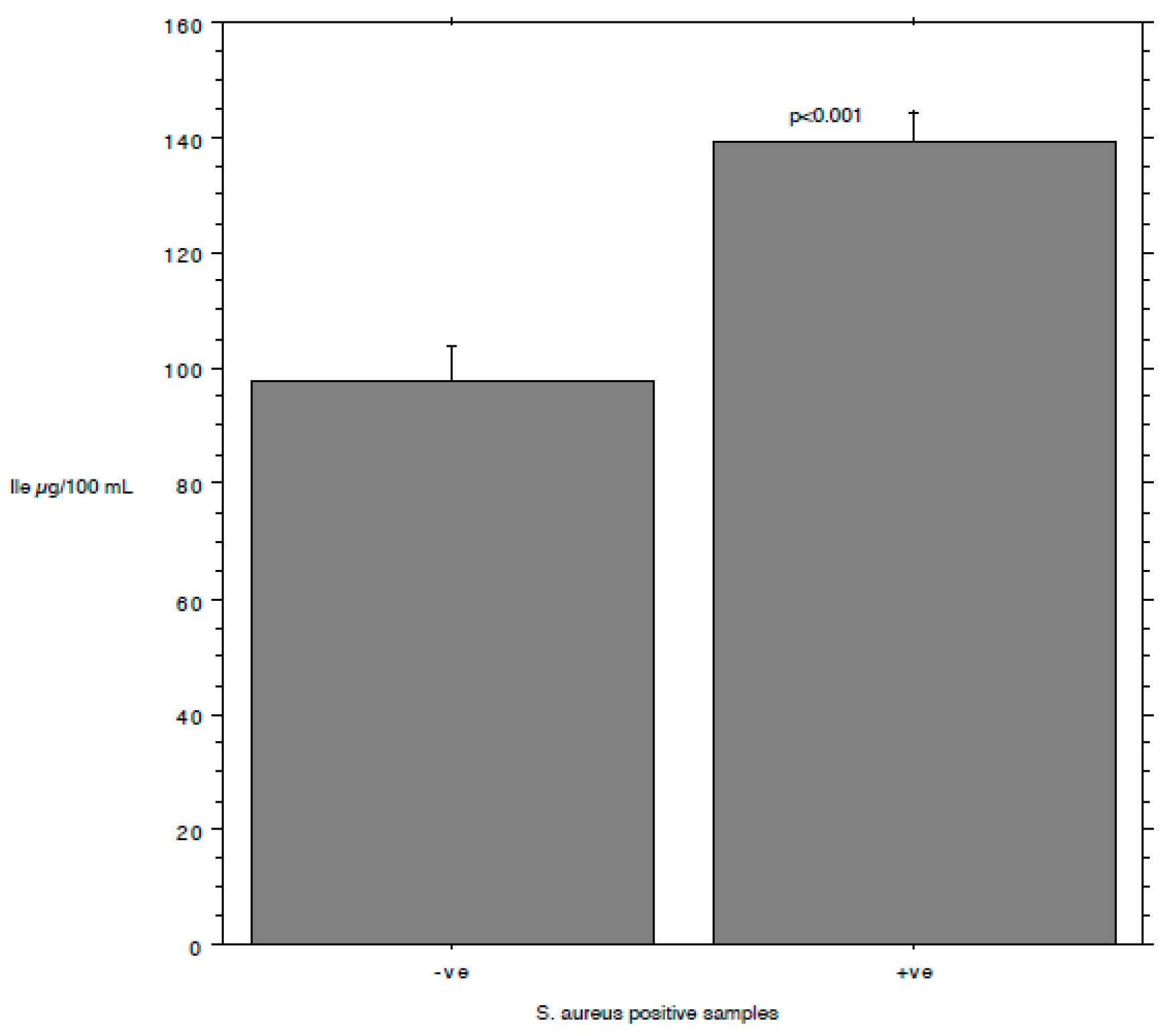
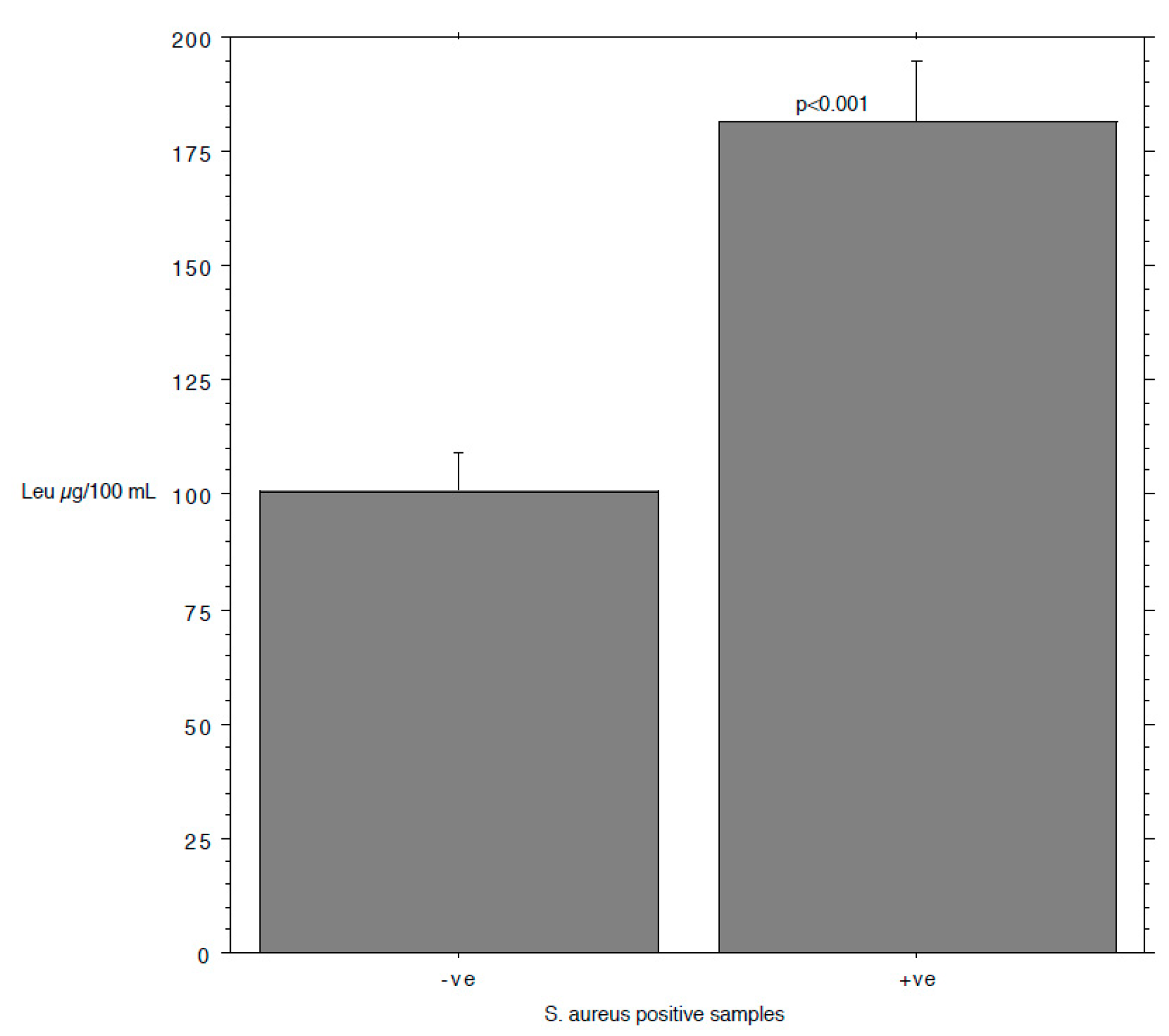
3. Results and Discussion
4. Conclusions
Author Contributions
Acknowledgments
Conflicts of Interest
References
- Grispoldi, L.; Massetti, L.; Sechi, P.; Iulietto, M.F.; Ceccarelli, M.; Karama, M.; Popescu, P.A.; Pandolfi, F.; Cenci-Goga, B.T. Short communication: Characterization of enterotoxin-producing Staphylococcus aureus isolated from mastitic cows. J. Dairy Sci. 2019, 102, 1059–1065. [Google Scholar] [CrossRef] [PubMed]
- Roberson, J.R.; Fox, L.K.; Hancock, D.D.; Gay, J.M.; Besser, T.E. Ecology of Staphylococcus aureus Isolated from Various Sites on Dairy Farms. J. Dairy Sci. 1994, 77, 3354–3364. [Google Scholar] [CrossRef]
- Fursova, K.K.; Shchannikova, M.P.; Loskutova, I.V.; Shepelyakovskaya, A.O.; Laman, A.G.; Boutanaev, A.M.; Sokolov, S.L.; Artem’eva, O.A.; Nikanova, D.A.; Zinovieva, N.A.; et al. Exotoxin diversity of Staphylococcus aureus isolated from milk of cows with subclinical mastitis in Central Russia. J. Dairy Sci. 2018, 101, 4325–4331. [Google Scholar] [CrossRef] [PubMed]
- Birhanu, M.; Leta, S.; Mamo, G.; Tesfaye, S. Prevalence of bovine subclinical mastitis and isolation of its major causes in Bishoftu Town, Ethiopia. BMC Res. Notes 2017, 10, 767. [Google Scholar] [CrossRef] [PubMed]
- Schukken, Y.H.; González, R.N.; Tikofsky, L.L.; Schulte, H.F.; Santisteban, C.G.; Welcome, F.L.; Bennett, G.J.; Zurakowski, M.J.; Zadoks, R.N. CNS mastitis: Nothing to worry about? Vet. Microbiol. 2009, 134, 9–14. [Google Scholar] [CrossRef] [PubMed]
- Tenhagen, B.A.; Hansen, I.; Reinecke, A.; Heuwieser, W. Prevalence of pathogens in milk samples of dairy cows with clinical mastitis and in heifers at first parturition. J. Dairy Res. 2009, 76, 179–187. [Google Scholar] [CrossRef] [PubMed]
- Boddie, R.L.; Nickerson, S.C.; Owens, W.E.; Watts, J.L. Udder microflora in nonlactating heifers. Agri Pract. 1987, 8, 22–25. [Google Scholar]
- Trinidad, P.; Nickerson, S.C.; Adkinson, R.W. Histopathology of Staphylococcal Mastitis in Unbred Dairy Heifers. J. Dairy Sci. 1990, 73, 639–647. [Google Scholar] [CrossRef]
- Trinidad, P.; Nickerson, S.C.; Alley, T.K. Prevalence of Intramammary Infection and Teat Canal Colonization In Unbred and Primigravid Dairy Heifers. J. Dairy Sci. 1990, 73, 107–114. [Google Scholar] [CrossRef]
- Jones, G.M.; Pearson, R.E.; Clabaugh, G.A.; Heald, C.W. Relationships Between Somatic Cell Counts and Milk Production. J. Dairy Sci. 1984, 67, 1823–1831. [Google Scholar] [CrossRef]
- Kehrli, M.E., Jr.; Shuster, D.E. Factors Affecting Milk Somatic Cells and Their Role in Health of the Bovine Mammary Gland. J. Dairy Sci. 1994, 77, 619–627. [Google Scholar] [CrossRef]
- Sharma, N.; Singh, N.K.; Bhadwal, M.S. Relationship of Somatic Cell Count and Mastitis: An Overview. Asian-australas. J. Anim. Sci. 2011, 24, 429–438. [Google Scholar] [CrossRef]
- Smith, K.L.; Hillerton, J.E.; Harmon, R.J. NMC Guidelines on Normal and Abnormal Raw Milk Based on SCC and Signs of Clinical Mastitis; NMC, Inc.: Madison, WI, USA, 2001. [Google Scholar]
- Petzer, I.-M.; Karzis, J.; Donkin, E.F.; Webb, E.C.; Etter, E.M.C. Validity of somatic cell count as indicator of pathogen-specific intramammary infections. J. S. Afr. Vet. Assoc. 2017, 88, e1–e10. [Google Scholar] [CrossRef] [PubMed]
- Baeker, R.; Haebel, S.; Schlatterer, K.; Schlatterer, B. Lipocalin-type prostaglandin D synthase in milk: A new biomarker for bovine mastitis. Prostaglandins Other Lipid Mediat. 2002, 67, 75–88. [Google Scholar] [CrossRef]
- Csapó, J.; Csapó-Kiss, Z.; Stefler, J.; Martin, T.G.; Némethy, S. Influence of Mastitis on D-Amino Acid Content of Milk. J. Dairy Sci. 1995, 78, 2375–2381. [Google Scholar] [CrossRef]
- Viguier, C.; Arora, S.; Gilmartin, N.; Welbeck, K.; O’Kennedy, R. Mastitis detection: Current trends and future perspectives. Trends Biotechnol. 2008, 27, 486–493. [Google Scholar] [CrossRef] [PubMed]
- Andrei, S.; Culea, M.; Matei, S.; Pintea, A.; Groza, I.S. Amino Acid Concentration in Normal and Subclinical Mastitis Milk. Bull. Univ. Agric. Sci. Vet. Med. Cluj Napoca 2011, 1, 15–19. [Google Scholar] [CrossRef]
- Brückner, H.; Jaek, P.; Langer, M.; Godel, H. Liquid chromatographic determination of D-amino acids in cheese and cow milk. Implication of starter cultures, amino acid racemases, and rumen microorganisms on formation, and nutritional considerations. Amino Acids 1992, 2, 271–284. [Google Scholar] [CrossRef]
- Ianni, F.; Sardella, R.; Lisanti, A.; Gioiello, A.; Cenci Goga, B.T.; Lindner, W.; Natalini, B. Achiral-chiral two-dimensional chromatography of free amino acids in milk: A promising tool for detecting different levels of mastitis in cows. J. Pharm. Biomed. Anal. 2015, 116, 40–46. [Google Scholar] [CrossRef]
- Ianni, F.; Sechi, P.; La Mantia, A.; Pucciarini, L.; Camaioni, E.; Cenci Goga, T.B.; Sardella, R.; Natalini, B. The Relationships between Somatic Cells and Isoleucine, Leucine and Tyrosine Content in Cow Milk. Appl. Sci. 2019, 9. [Google Scholar] [CrossRef]
- Csapo, J.; Csapo-Kiss, Z.; Fau-Stefler, J.; Stefler, J.; Fau-Martin, T.G.; Martin Tg Fau-Nemethy, S.; Nemethy, S. Influence of mastitis on D-amino acid content of milk. J. Dairy Sci. 1995, 78, 2375–2381. [Google Scholar] [PubMed]
- Kaiser, J.C.; King, A.N.; Grigg, J.C.; Sheldon, J.R.; Edgell, D.R.; Murphy, M.E.P.; Brinsmade, S.R.; Heinrichs, D.E. Repression of branched-chain amino acid synthesis in Staphylococcus aureus is mediated by isoleucine via CodY, and by a leucine-rich attenuator peptide. PLoS Genet. 2018, 14, e1007159. [Google Scholar] [CrossRef] [PubMed]
- Lincoln, R.A.; Leigh, J.A.; Jones, N.C. The amino acid requirements of Staphylococcus aureus isolated from cases of bovine mastitis. Vet. Microbiol. 1995, 45, 275–279. [Google Scholar] [CrossRef]
- Salasia, S.I.; Khusnan, Z.; Lammler, C.; Zschock, M. Comparative studies on pheno- and genotypic properties of Staphylococcus aureus isolated from bovine subclinical mastitis in central Java in Indonesia and Hesse in Germany. J. Vet. Sci. 2004, 5, 103–109. [Google Scholar] [CrossRef]
- Kaiser, J.C.; Omer, S.; Sheldon, J.R.; Welch, I.; Heinrichs, D.E. Role of BrnQ1 and BrnQ2 in branched-chain amino acid transport and virulence in Staphylococcus aureus. Infect. Immun. 2015, 83, 1019–1029. [Google Scholar] [CrossRef]
- Kaiser, J.C.; Sen, S.; Sinha, A.; Wilkinson, B.J.; Heinrichs, D.E. The role of two branched-chain amino acid transporters in Staphylococcus aureus growth, membrane fatty acid composition and virulence. Mol. Microbiol. 2016, 102, 850–864. [Google Scholar] [CrossRef]
- Onoue, Y.; Mori, M. Amino acid requirements for the growth and enterotoxin production by Staphylococcus aureus in chemically defined media. Int. J. Food Microbiol. 1997, 36, 77–82. [Google Scholar] [CrossRef]
- Mariano, V.; McCrindle, C.M.E.; Cenci-Goga, B.; Picard, J.A. Case-Control Study to Determine whether River Water Can Spread Tetracycline Resistance to Unexposed Impala (Aepyceros melampus) in Kruger National Park (South Africa). Appl. Environ. Microbiol. 2009, 75, 113–118. [Google Scholar] [CrossRef]
- Cenci-Goga, B.T.; Karama, M.; Rossitto, P.V.; Morgante, R.A.; Cullor, J.S. Enterotoxin Production by Staphylococcus aureus Isolated from Mastitic Cows. J. Food Prot. 2003, 66, 1693–1696. [Google Scholar] [CrossRef]
- Ibrahim, G.F. A simple sensitive method for determining staphylococcal thermonuclease in cheese. J. Appl. Bacteriol. 1981, 51, 307–312. [Google Scholar] [CrossRef]
- Cenci-Goga, B.T.; Crotti, S.; Costarelli, S.; Rondini, C.; Karama, M.; Bennett, P. Detection of tet(M) gene from raw milk by rapid DNA extraction followed by a two-step PCR with nested primers. J. Food Prot. 2004, 67, 2833–2838. [Google Scholar] [CrossRef] [PubMed]
- Ishizawa, M.; Kobayashi, Y.; Miyamura, T.; Matsuura, S. Simple procedure of DNA isolation from human serum. Nucleic Acid Res. 1991, 19, 5792. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chaimbault, P.; Petritis, K.; Elfakir, C.; Dreux, M. Determination of 20 underivatized proteinic amino acids by ion-pairing chromatography and pneumatically assisted electrospray mass spectrometry. J. Chromatogr. A 1999, 855, 191–202. [Google Scholar] [CrossRef]
- Chaimbault, P.; Petritis, K.; Elfakir, C.; Dreux, M. Ion-pair chromatography on a porous graphitic carbon stationary phase for the analysis of twenty underivatized protein amino acids. J. Chromatogr. A 2000, 870, 245–254. [Google Scholar] [CrossRef]
- Petritis, K.; Person, M.d.; Elfakir, C.; Dreux, M. Validation of an Ion-Interaction Chromatography Analysis of Underivatized Amino Acids in Commercial Preparation Using Evaporative Light Scattering Detection. Chromatographia 2004, 60, 293–298. [Google Scholar] [CrossRef]
- Petritis, K.N.; Chaimbault, P.; Elfakir, C.; Dreux, M. Ion-pair reversed-phase liquid chromatography for determination of polar underivatized amino acids using perfluorinated carboxylic acids as ion pairing agent. J. Chromatogr. A 1999, 833, 147–155. [Google Scholar] [CrossRef]
- Lei, T.; Yang, J.; Ji, Y. Determination of essentiality and regulatory function of staphylococcal YeaZ in branched-chain amino acid biosynthesis. Virulence 2015, 6, 75–84. [Google Scholar] [CrossRef] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| ssc Count (× 1000) | S. aureus + ve (n) | S. aureus Load (log cfu/mL) | Ile (µg/mL) | Leu (µg/mL) | Val (µg/mL) | Tyr (µg/mL) | |||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Scc Groups | Mean | sd | Mean | sd | Mean | sd | Mean | sd | Mean | sd | Mean | sd | |
| < 100 (n = 27) | 43.6 | 32.6 | 11 | 0.89 | 1.37 | 118 | 32 | 135 | 57 | 428 | 638 | 127 | 138 |
| 100–199 (n = 15) | 142.6 | 26.1 | 8 | 1.53 | 1,60 | 111 | 33 | 126 | 51 | 256 | 153 | 97 | 60 |
| 200–499 (n = 8) | 364.3 | 57.8 | 7 | 1.88 | 1,25 | 118 | 34 | 144 | 49 | 246 | 139 | 113 | 51 |
| 500–999 (n = 10) | 672.2 | 147.5 | 8 | 2.00 | 1,63 | 128 | 47 | 173 | 109 | 226 | 104 | 109 | 43 |
| > 1000 (n = 5) | 2967.2 | 941.3 | 4 | 2.80 | 1,64 | 168 | 54 | 250 | 143 | 335 | 203 | 165 | 58 |
| All (n = 65) | 427.5 | 808.9 | 38 | 1.48 | 1,54 | 122 | 38 | 149 | 79 | 330 | 433 | 118 | 97 |
| Threshold | se | 95% LCI and UCI | sp | 95% LCI and UCI |
|---|---|---|---|---|
| SCC > 400,000/mL | 0.37 | (0.14–0.60) | 0.89 | (0.80–0.98) |
| SCC > 200,000/mL | 0.50 | (0.30–0.70) | 0.85 | (0.74–0.96) |
| SCC > 150,000/mL | 0.58 | (0.41–0.75) | 0.67 | (0.51–0.83) |
| SCC > 100,000/mL | 0.71 | (0.57–0.85) | 0.59 | (0.19–0.41) |
| Ile > 100 µg/mL | 0.97 | (0.93–1.00) | 0.70 | (0.50–0.90) |
| Ile > 150 µg/mL | 0.32 | (0.07–0.56) | 0.93 | (0.85–1.00) |
| Leu > 100 µg/mL | 0.92 | (0.84–1.00) | 0.52 | (0.28–0.76) |
| Leu > 100 µg/mL | 0.55 | (0.37–0.74) | 0.78 | (0.65–0.91) |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Grispoldi, L.; Karama, M.; Ianni, F.; La Mantia, A.; Pucciarini, L.; Camaioni, E.; Sardella, R.; Sechi, P.; Natalini, B.; Cenci-Goga, B.T. The Relationship between S. aureus and Branched-Chain Amino Acids Content in Composite Cow Milk. Animals 2019, 9, 981. https://doi.org/10.3390/ani9110981
Grispoldi L, Karama M, Ianni F, La Mantia A, Pucciarini L, Camaioni E, Sardella R, Sechi P, Natalini B, Cenci-Goga BT. The Relationship between S. aureus and Branched-Chain Amino Acids Content in Composite Cow Milk. Animals. 2019; 9(11):981. https://doi.org/10.3390/ani9110981
Chicago/Turabian StyleGrispoldi, L., M. Karama, F. Ianni, A. La Mantia, L. Pucciarini, E. Camaioni, R. Sardella, P. Sechi, B. Natalini, and B. T. Cenci-Goga. 2019. "The Relationship between S. aureus and Branched-Chain Amino Acids Content in Composite Cow Milk" Animals 9, no. 11: 981. https://doi.org/10.3390/ani9110981
APA StyleGrispoldi, L., Karama, M., Ianni, F., La Mantia, A., Pucciarini, L., Camaioni, E., Sardella, R., Sechi, P., Natalini, B., & Cenci-Goga, B. T. (2019). The Relationship between S. aureus and Branched-Chain Amino Acids Content in Composite Cow Milk. Animals, 9(11), 981. https://doi.org/10.3390/ani9110981