Dietary Cadmium Chloride Supplementation Impairs Renal Function and Bone Metabolism of Laying Hens


Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animal Ethics
2.2. Birds, Diets, and Management
2.3. Collection of Samples and Measurement
2.4. Statistical Analysis
3. Results
3.1. Cd Residues and Ca, P Levels in the Serum and Kidney of Laying Hens
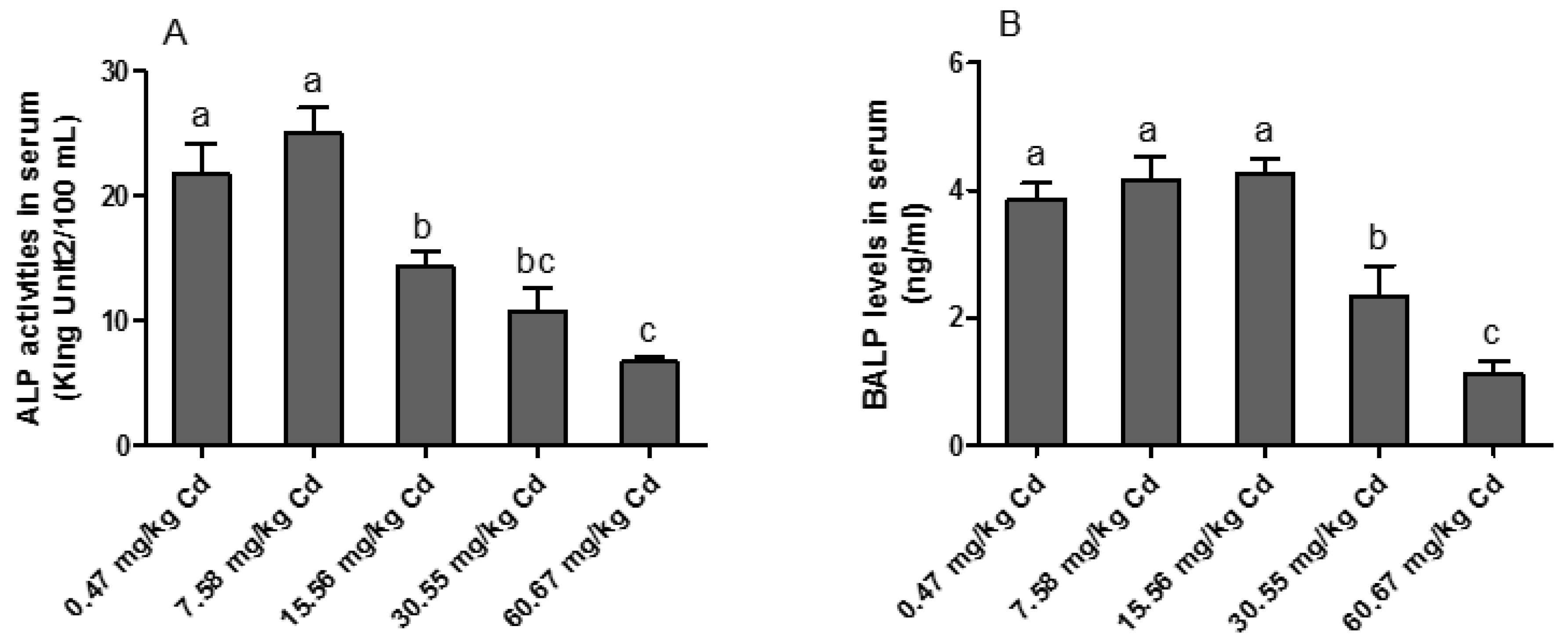
3.2. ALP Activity in the Serum of Laying Hens
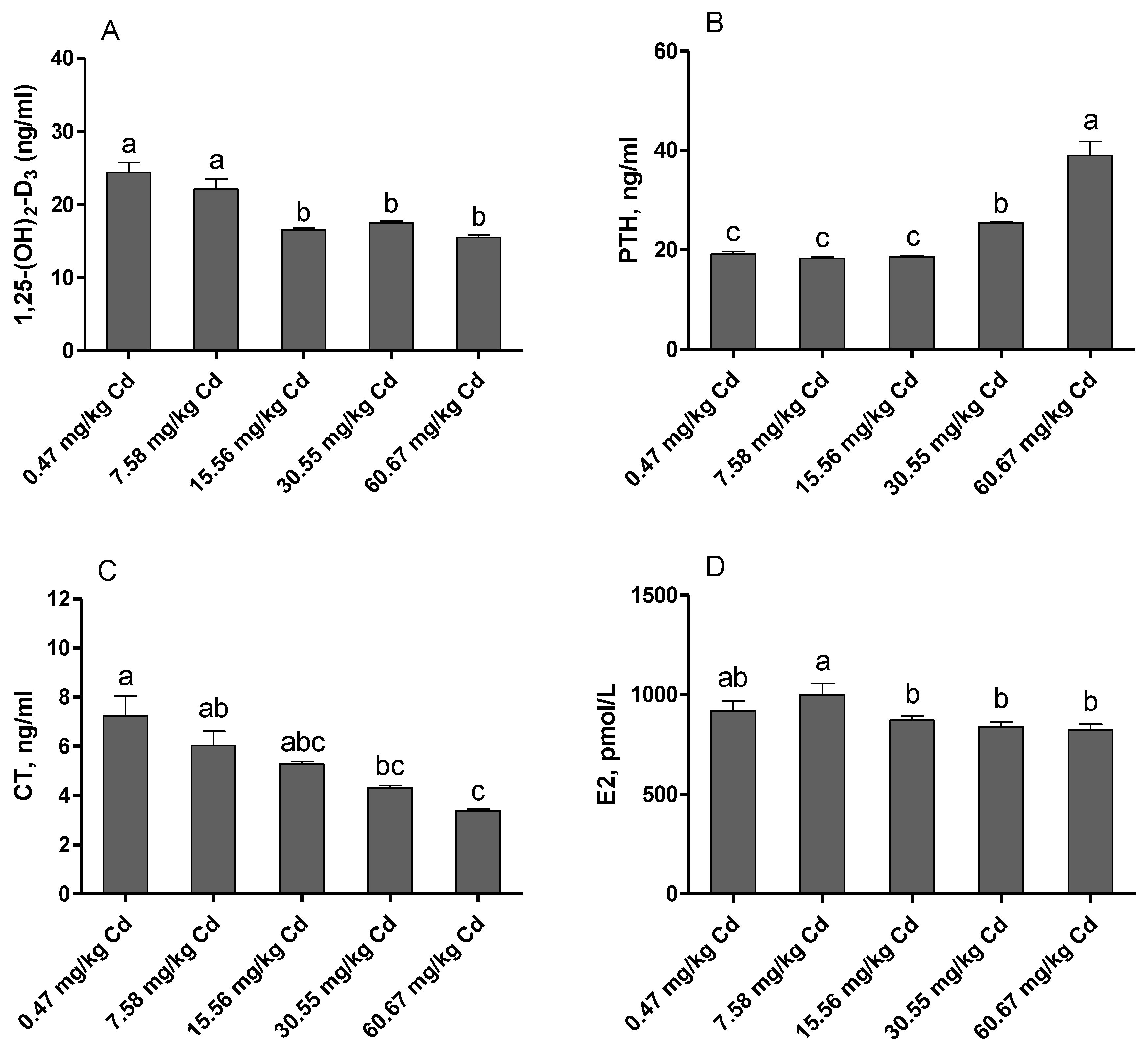
3.3. Hormone Levels in the Serum of Laying Hens
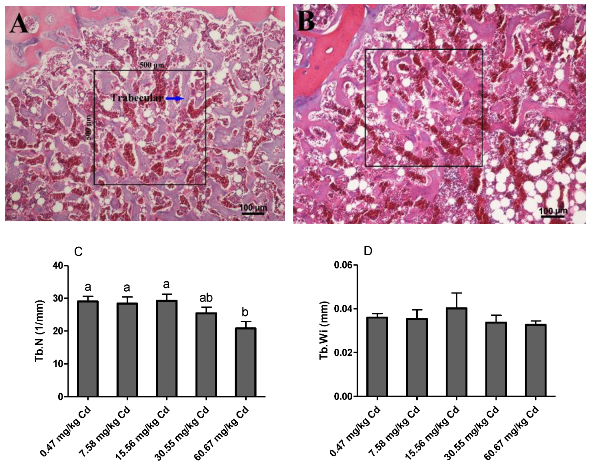
3.4. Bone (Tibia) Volume of Laying Hens Exposed to Cd
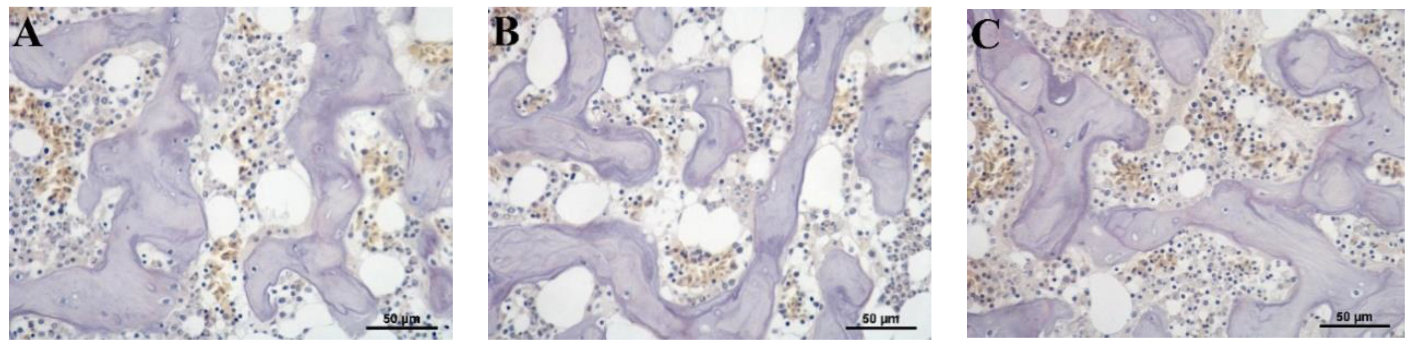
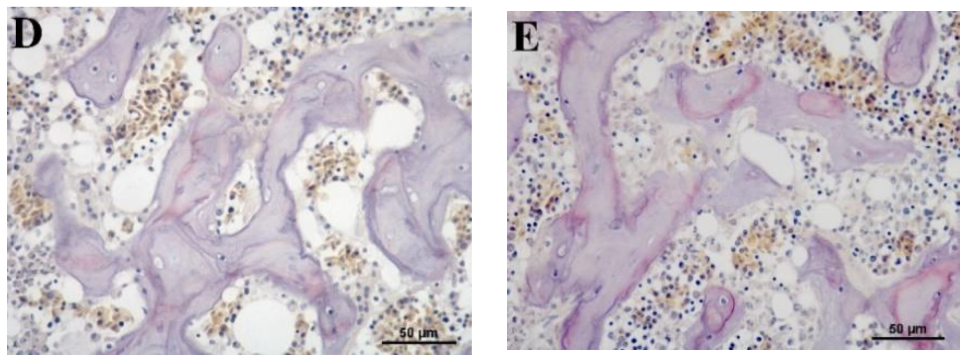
3.5. Histological Evaluation of Osteoclast
3.6. Renal Function of Laying Hens
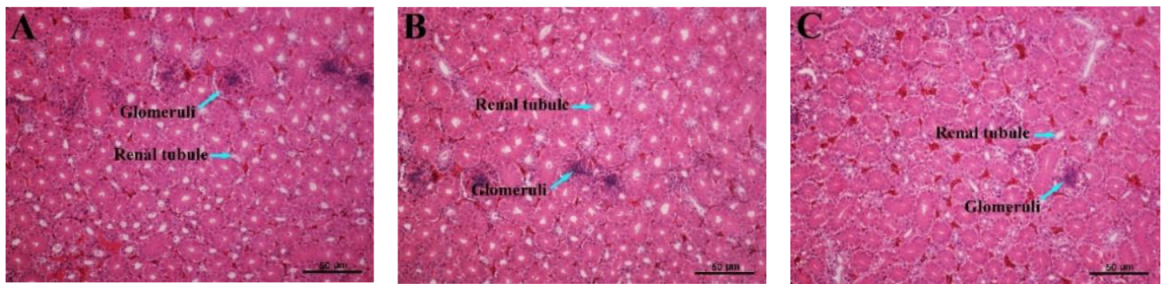
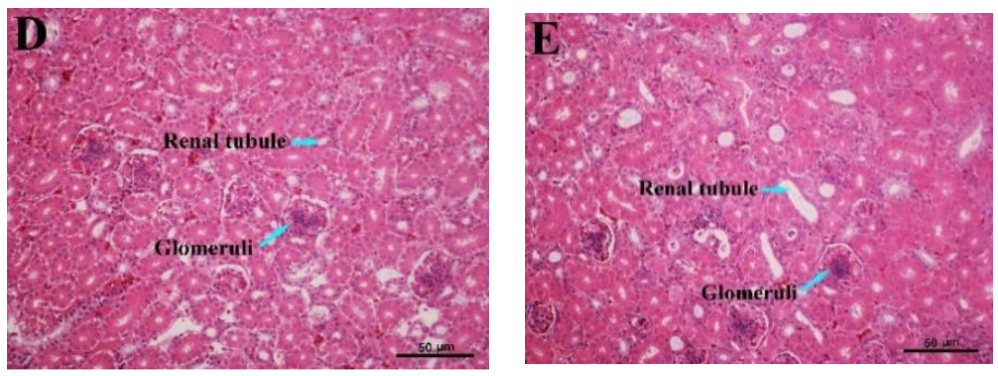
3.7. Histopathological Variations of the Kidney
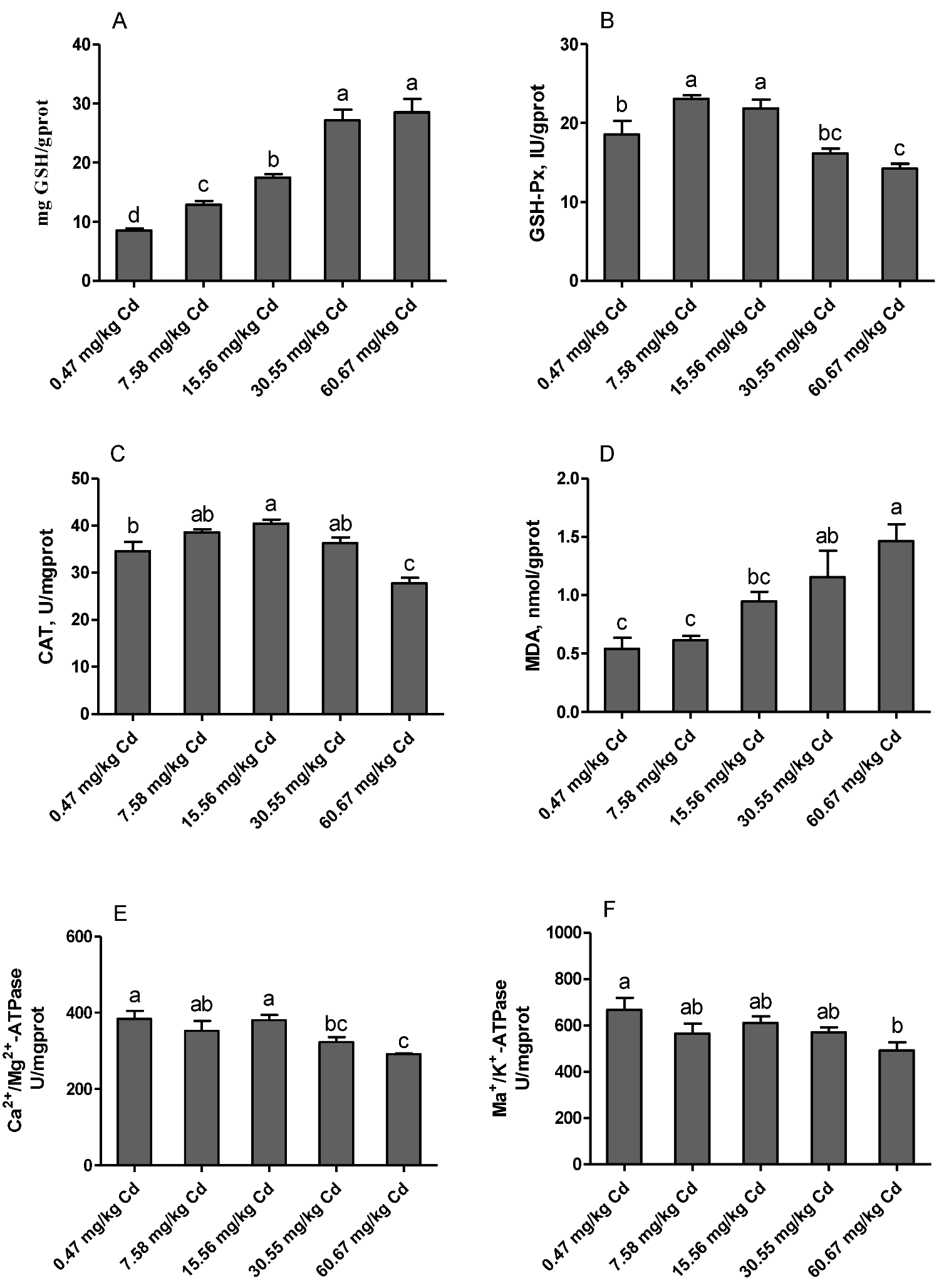
3.8. Renal Antioxidant Parameters and ATPase Activities
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- ATSDR. Toxicological Profile for Cadmium; Agency for Toxic Substances and Disease Registry (US): Atlanta, GA, USA, 1 September 2012. Available online: https://www.ncbi.nlm.nih.gov/books/NBK158840 (accessed on 4 November 2019).
- Wallin, M.; Sallsten, G.; Lundh, T.; Barregard, L. Low-level cadmium exposure and effects on kidney function. Occup. Environ. Med. 2014, 71, 848–854. [Google Scholar] [CrossRef] [PubMed]
- Lv, Y.J.; Wei, Q.Z.; Zhang, Y.C.; Huang, R.; Li, B.S.; Tan, J.B.; Wang, J.; Ling, H.T.; Wu, S.X.; Yang, X.F. Low-dose cadmium exposure acts on rat mesenchymal stem cells via RANKL/OPG and downregulate osteogenic differentiation genes. Environ. Pollut. 2019, 249, 620–628. [Google Scholar] [CrossRef] [PubMed]
- Satarug, S.; Vesey, D.A.; Gobe, G.C. Kidney cadmium toxicity, diabetes and high blood pressure: The perfect storm. Tohoku J. Exp. Med. 2017, 241, 65–87. [Google Scholar] [CrossRef] [PubMed]
- Jin, T.; Nordberg, G.; Ye, T.; Bo, M.; Wang, H.; Zhu, G.; Kong, Q.; Bernard, A. Osteoporosis and renal dysfunction in a general population exposed to cadmium in China. Environ. Res. 2004, 96, 353–359. [Google Scholar] [CrossRef]
- Chalkley, S.R.; Richmond, J.; Barltrop, D. Measurement of vitamin D3 metabolites in smelter workers exposed to lead and cadmium. Occup. Environ. Med. 1998, 55, 446–452. [Google Scholar] [CrossRef]
- Regunathan, A.; Glesne, D.A.; Wilson, A.K.; Song, J.; Nicolae, D.; Flores, T.; Bhattacharyya, M.H. Microarray analysis of changes in bone cell gene expression early after cadmium gavage in mice. Toxicol. Appl. Pharm. 2003, 191, 272–293. [Google Scholar] [CrossRef]
- Horiguchi, H.; Oguma, E.; Sasaki, S.; Miyamoto, K.; Ikeda, Y.; Machida, M.; Kayama, F. Environmental exposure to cadmium at a level insufficient to induce renal tubular dysfunction does not affect bone density among female Japanese farmers. Environ. Res. 2005, 97, 83–92. [Google Scholar] [CrossRef]
- Olgun, O.; Bahtiyarca, Y. Effects of dietary cadmium and boron supplementation on performance, eggshell quality and mineral concentrations of bone in laying hens. Biol. Trace Elem. Res. 2015, 167, 56–62. [Google Scholar] [CrossRef]
- Wallin, M.; Sallsten, G.; Fabricius-Lagging, E.; Öhrn, C.; Lundh, T.; Barregard, L. Kidney cadmium levels and associations with urinary calcium and bone mineral density: A cross-sectional study in Sweden. Environ. Health 2013, 12, 22. [Google Scholar] [CrossRef]
- Chen, W.; Zhao, F.; Tian, Z.M.; Zhang, H.X.; Ruan, D.; Li, Y.; Wang, S.; Zheng, C.T.; Lin, Y.C. Dietary calcium deficiency in laying ducks impairs eggshell quality by suppressing shell biomineralization. J. Exp. Biol. 2015, 218, 3336–3343. [Google Scholar] [CrossRef]
- Squire, M.E.; Veglia, M.K.; Drucker, K.A.; Brazeal, K.R.; Hahn, T.P.; Watts, H.E. Estrogen levels influence medullary bone quantity and density in female house finches and pine siskins. Gen. Comp. Endocrinol. 2017, 246, 249–257. [Google Scholar] [CrossRef] [PubMed]
- Kawashima, H.; Kurokawa, K. Unique hormonal regulation of vitamin D metabolism in the mammalian kidney. Min. Electrolyte Metab. 1983, 9, 227–235. [Google Scholar]
- Berglund, M.; Akesson, A.; Bjellerup, P.; Vahter, M. Metal–bone interactions. Toxicol. Lett. 2000, 112, 219–225. [Google Scholar] [CrossRef]
- Gobin, B.; Battaglia, S.; Lanel, R.; Chesneau, J.; Amiaud, J.; Rédini, F.; Ory, B.; Heymann, D. NVP-BEZ235, a dual PI3K/mTOR inhibitor, inhibits osteosarcoma cell proliferation and tumor development in vivo with an improved survival rate. Cancer Lett. 2014, 344, 291–298. [Google Scholar] [CrossRef]
- Reyer, H.; Oster, M.; Wittenburg, D.; Murani, E.; Ponsuksili, S.; Wimmers, K. Genetic Contribution to Variation in Blood Calcium, Phosphorus, and Alkaline Phosphatase Activity in Pigs. Front. Genet. 2019, 10, 590. [Google Scholar] [CrossRef]
- Rath, N.C.; Huff, G.R.; Huff, W.E.; Balog, J.M. Factors regulating bone maturity and strength in poultry. Poult. Sci. 2000, 79, 1024–1032. [Google Scholar] [CrossRef]
- Wong, C.K.C.; Wong, M.H. Morphological and biochemical changes in the gills of tilapia (Oreochromis mossambicus) to ambient cadmium exposure. Aquat. Toxicol. 2000, 48, 517–527. [Google Scholar] [CrossRef]
- Pratap, H.B.; Fu, H.; Lock, R.A.C.; Wendelaar Bonga, S.E. Effects of waterborne and dietary cadmium on plasma ions of the teleost Oreochromis mossambicus in relation to water calcium levels. Arch. Environ. Contam. Toxicol. 1989, 18, 568–575. [Google Scholar] [CrossRef]
- Liao, Y.; Cao, H.; Xia, B.; Xiao, Q.; Liu, P.; Hu, G.; Zhang, C. Changes in Trace Element Contents and Morphology in Bones of Duck Exposed to Molybdenum or/and Cadmium. Biol. Trace Elem. Res. 2017, 175, 449–457. [Google Scholar] [CrossRef]
- Zhou, X.; Hao, W.; Shi, H.; Hou, Y.; Xu, Q. Calcium Homeostasis Disruption—A Bridge Connecting Cadmium-Induced Apoptosis, Autophagy and Tumorigenesis. Oncol. Res. Treat. 2015, 38, 311–315. [Google Scholar] [CrossRef]
- Akerman, K.E.; Honkaniemi, J.; Scott, I.G.; Andersson, L.C. Interaction of Cd2+ with the calmodulin-activated (Ca2+ + Mg2+)-ATPase activity of human erythrocyte ghosts. Biochim. Biophys. Acta 1985, 845, 48–53. [Google Scholar] [CrossRef]
- Wu, X.; Jin, T.; Wang, Z.; Ye, T.; Kong, Q.; Nordberg, G. Urinary calcium as a biomarker of renal dysfunction in a general population exposed to cadmium. J. Occup. Environ. Med. 2001, 43, 898–904. [Google Scholar] [CrossRef] [PubMed]
- Kazantzis, G. Cadmium, osteoporosis and calcium metabolism. Biometals 2004, 17, 493–498. [Google Scholar] [CrossRef] [PubMed]
- Ali, N.N.; Rowe, J.; Teich, N.M. Constitutive expression of non-bone/liver/kidney alkaline phosphatase in human osteosarcoma cell lines. J. Bone Min. Res. 1996, 11, 512–520. [Google Scholar] [CrossRef]
- Magnusson, P.; Larsson, L.; Magnusson, M.; Davie, M.W.; Sharp, C.A. Isoforms of bone alkaline phosphatase: Characterization and origin in human trabecular and cortical bone. J. Bone Min. Res. 1999, 14, 1926–1933. [Google Scholar] [CrossRef]
- Treviño, S.; Andrade-García, A.; Herrera Camacho, I.; León-Chavez, B.A.; Aguilar-Alonso, P.; Flores, G.; Brambila, E. Chronic Cadmium Exposure Lead to Inhibition of Serum and Hepatic Alkaline Phosphatase Activity in Wistar Rats. J. Biochem. Mol. Toxicol. 2015, 29, 587–594. [Google Scholar] [CrossRef]
- Chen, X.; Zhu, G.; Gu, S.; Jin, T.; Shao, C. Effects of cadmium on osteoblasts and osteoclasts in vitro. Environ. Toxicol. Pharm. 2009, 28, 232–236. [Google Scholar] [CrossRef]
- Brzóska, M.M.; Moniuszko-Jakoniuk, J. Bone metabolism of male rats chronically exposed to cadmium. Toxicol. Appl. Pharm. 2005, 207, 195–211. [Google Scholar] [CrossRef]
- Morgan, S.L. Calcium and vitamin D in osteoporosis. Rheum. Dis. Clin. N. Am. 2001, 27, 101–130. [Google Scholar] [CrossRef]
- Gardella, T.J.; Kronenberg, H.M. Parathyroid Hormone (PTH). Encycl. Endocr. Dis. 2004, 513–520. [Google Scholar] [CrossRef]
- Naot, D.; Musson, D.S.; Cornish, J. The Activity of Peptides of the Calcitonin Family in Bone. Physiol. Rev. 2019, 99, 781–805. [Google Scholar] [CrossRef] [PubMed]
- Holick, M.F. Vitamin D for health and in chronic kidney disease. Semin. Dial. 2005, 18, 266–275. [Google Scholar] [CrossRef] [PubMed]
- Kjellström, T. Mechanism and epidemiology of bone effects of cadmium. IARC Sci. Publ. 1992, 118, 301–310. [Google Scholar]
- Wronski, T.J.; Dann, L.M.; Horner, S.L. Time course of vertebral osteopenia in ovariectomized rats. Bone 1989, 10, 295–301. [Google Scholar] [CrossRef]
- Parikka, V.; Lehenkari, P.; Sassi, M.L.; Halleen, J.; Risteli, J.; Härkönen, P.; Väänänen, H.K. Estrogen reduces the depth of resorption pits by disturbing the organic bone matrix degradation activity of mature osteoclasts. Endocrinology 2001, 142, 5371–5378. [Google Scholar] [CrossRef]
- Baksi, S.N.; Kenny, A.D. Vitamin D3 Metabolism in Immature Japanese Quail: Effects of Ovarian Hormones. Endocrinology 1977, 101, 1216–1220. [Google Scholar] [CrossRef]
- Rodríguez, J.; Mandalunis, P.M. Effect of cadmium on bone tissue in growing animals. Exp. Toxicol. Pathol. 2016, 68, 391–397. [Google Scholar] [CrossRef]
- Wilson, A.K.; Cerny, E.A.; Smith, B.D.; Wagh, A.; Bhattacharyya, M.H. Effects of cadmium on osteoclast formation and activity in vitro. Toxicol. Appl. Pharm. 1996, 140, 451–460. [Google Scholar] [CrossRef]
- Navaneethan, D.; Rasool, M. p-Coumaric acid, a common dietary polyphenol, protects cadmium chloride-induced nephrotoxicity in rats. Ren. Fail. 2014, 36, 244–251. [Google Scholar] [CrossRef]
- Brzóska, M.M.; Kamiński, M.; Dziki, M.; Moniuszko-Jakoniuk, J. Changes in the structure and function of the kidney of rats chronically exposed to cadmium. II. Histoenzymatic studies. Arch. Toxicol. 2004, 78, 226–231. [Google Scholar]
- Thévenod, F.; Friedmann, J.M. Cadmium-mediated oxidative stress in kidney proximal tubule cells induces degradation of Na+/K+-ATPase through proteasomal and endo-/lysosomal proteolytic pathways. FASEB J. 1999, 13, 1751–1761. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sabolic, I.; Ljubojevic, M.; Herak-Kramberger, C.M.; Brown, D. Cd-MT causes endocytosis of brush-border transporters in rat renal proximal tubules. Am. J. Physiol. Ren. Physiol. 2002, 283, 1389–1402. [Google Scholar] [CrossRef] [Green Version]
- Valko, M.M.H.C.M.; Morris, H.; Cronin, M.T.D. Metals, toxicity and oxidative stress. Curr. Med. Chem. 2005, 12, 1161–1208. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Uchida, M.; Teranishi, H.; Aoshima, K.; Katoh, T.; Kasuya, M.; Inadera, H. Reduction of erythrocyte catalase and superoxide dismutase activities in male inhabitants of a cadmium-polluted area in Jinzu river basin, Japan. Toxicol. Lett. 2004, 151, 451–457. [Google Scholar] [CrossRef] [PubMed]
- Cuypers, A.; Plusquin, M.; Remans, T.; Jozefczak, M.; Keunen, E.; Gielen, H.; Opdenakker, K.; Nair, A.R.; Munters, E.; Artois, T.J.; et al. Cadmium stress: An oxidative challenge. Biometals 2010, 23, 927–940. [Google Scholar] [CrossRef] [PubMed]
- Shaikh, A.Z.; Northup, J.B.; Vestergaard, P. Dependence of cadmium-metallothionein nephrotoxicity on glutathione. J. Toxicol. Environ. Health 1999, 57, 211–222. [Google Scholar]
- Reiter, R.J.; Melchiorri, D.; Sewerynek, E.; Poeggeler, B.; Barlow-Walden, L.; Chuang, J.; Ortiz, G.G.; Acuña-Castroviejo, D. A review of the evidence supporting melatonin’s role as an antioxidant. J. Pineal Res. 1995, 18, 1–11. [Google Scholar] [CrossRef]
- Nair, A.R.; Lee, W.K.; Smeets, K.; Swennen, Q.; Sanchez, A.; Thévenod, F.; Cuypers, A. Glutathione and mitochondria determine acute defense responses and adaptive processes in cadmium-induced oxidative stress and toxicity of the kidney. Arch. Toxicol. 2015, 89, 2273–2289. [Google Scholar] [CrossRef]
- Yuan, Y.; Jiang, C.Y.; Xu, H.; Sun, Y.; Hu, F.F.; Bian, J.C.; Liu, X.Z.; Gu, J.H.; Liu, Z.P. Cadmium-Induced apoptosis in primary rat cerebral cortical neurons culture Is mediated by a calcium signaling pathway. PLoS ONE 2013, 8, e64330. [Google Scholar] [CrossRef]
- Carafoli, E. Calcium pump of the plasma membrane. Physiol. Rev. 1991, 71, 129–153. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Basal Ingredients | Value | Nutrient Level 2 | Value |
|---|---|---|---|
| Corn, % | 65 | Metabolizable energy, MJ/kg | 2.65 |
| Soybean meal (42.0% crude protein), % | 21 | Crude protein, % | 15.73 |
| Feather meal, % | 1 | Ether extract, % | 6.32 |
| Fish meal, % | 1 | Lysine, % | 0.78 |
| Calcium carbonate, % | 7 | Methionine, % | 0.34 |
| Premix 1, % | 5 | Cysteine, % | 0.32 |
| Total phosphorus, % | 0.61 | ||
| Total, % | 100 | Calcium, % | 3.45 |
| Item | Dietary Cd Dosage, mg/kg | p-Value | SEM 3 | ||||
|---|---|---|---|---|---|---|---|
| 0.47 | 7.58 | 15.56 | 30.55 | 60.67 | |||
| Creatine, μmol/L | 10.91 b | 16.81 ab | 14.85 ab | 13.33 ab | 21.47 a | 0.025 | 2.670 |
| UA1, mg/L | 22.64 c | 25.66 c | 27.07 bc | 32.37 ab | 35.22 a | <0.001 | 1.794 |
| BUN2, mmol/L | 3.56 b | 3.52 b | 3.99 b | 4.67 ab | 5.82 a | 0.013 | 0.573 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhu, M.; Zhou, W.; Bai, L.; Li, H.; Wang, L.; Zou, X. Dietary Cadmium Chloride Supplementation Impairs Renal Function and Bone Metabolism of Laying Hens. Animals 2019, 9, 998. https://doi.org/10.3390/ani9110998
Zhu M, Zhou W, Bai L, Li H, Wang L, Zou X. Dietary Cadmium Chloride Supplementation Impairs Renal Function and Bone Metabolism of Laying Hens. Animals. 2019; 9(11):998. https://doi.org/10.3390/ani9110998
Chicago/Turabian StyleZhu, Mingkun, Wenting Zhou, Luhong Bai, Huaiyu Li, Liansong Wang, and Xiaoting Zou. 2019. "Dietary Cadmium Chloride Supplementation Impairs Renal Function and Bone Metabolism of Laying Hens" Animals 9, no. 11: 998. https://doi.org/10.3390/ani9110998
APA StyleZhu, M., Zhou, W., Bai, L., Li, H., Wang, L., & Zou, X. (2019). Dietary Cadmium Chloride Supplementation Impairs Renal Function and Bone Metabolism of Laying Hens. Animals, 9(11), 998. https://doi.org/10.3390/ani9110998