High Altitude Adaptability and Meat Quality in Tibetan Pigs: A Reference for Local Pork Processing and Genetic Improvement

 , , ,
, , ,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals and Treatments
2.2. Measurement of Carcass Characteristics
2.3. Measurement of Physical Correlates of Meat Quality
2.4. Muscle Chemical Composition
2.5. Quantitative Real-Time PCR
2.6. Analysis of Free Amino Acids (FAA) and Fatty Acid
2.7. Statistical Analysis
3. Results
3.1. Carcass Traits
3.2. Meat Quality and Muscle Chemical Composition
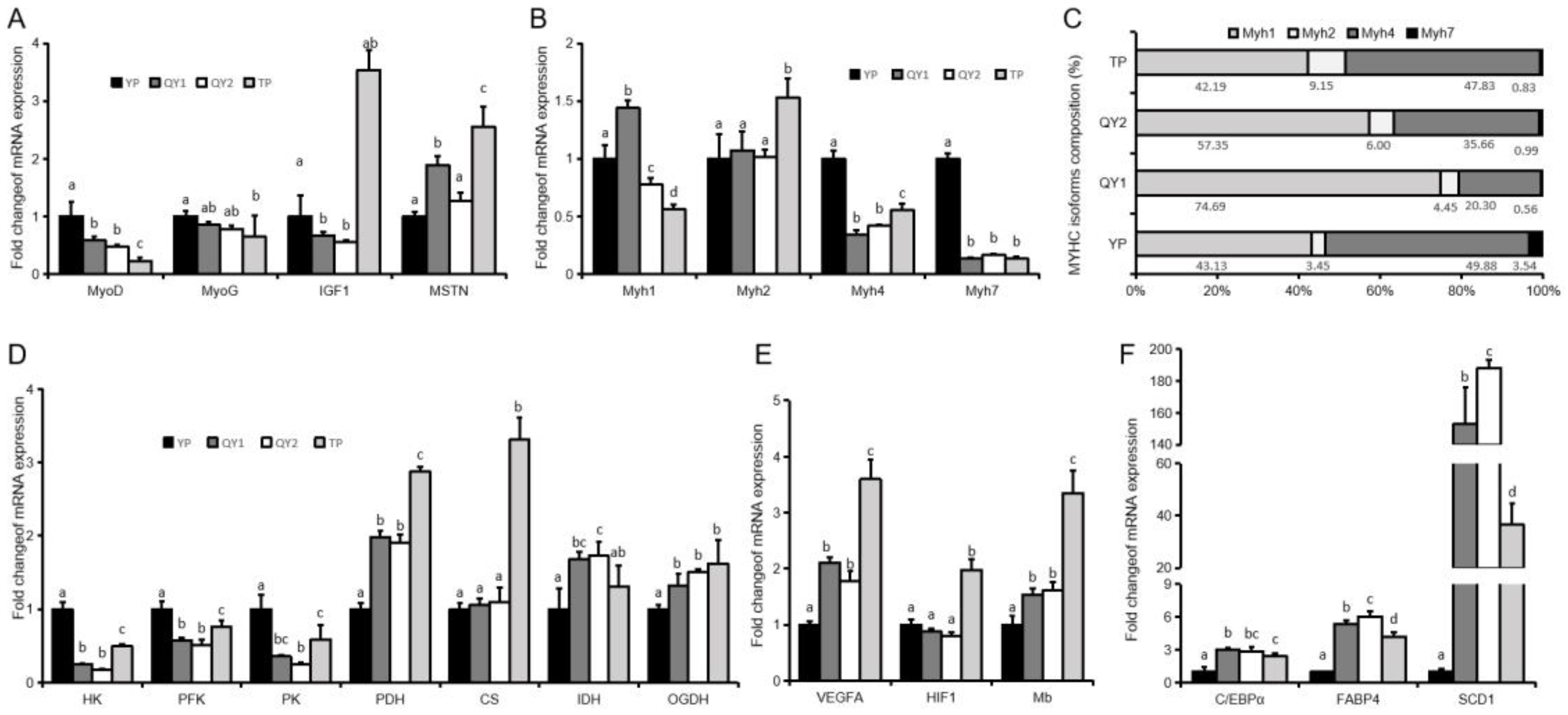
3.3. Expression of Genes in Longissimus Dorsi Muscle
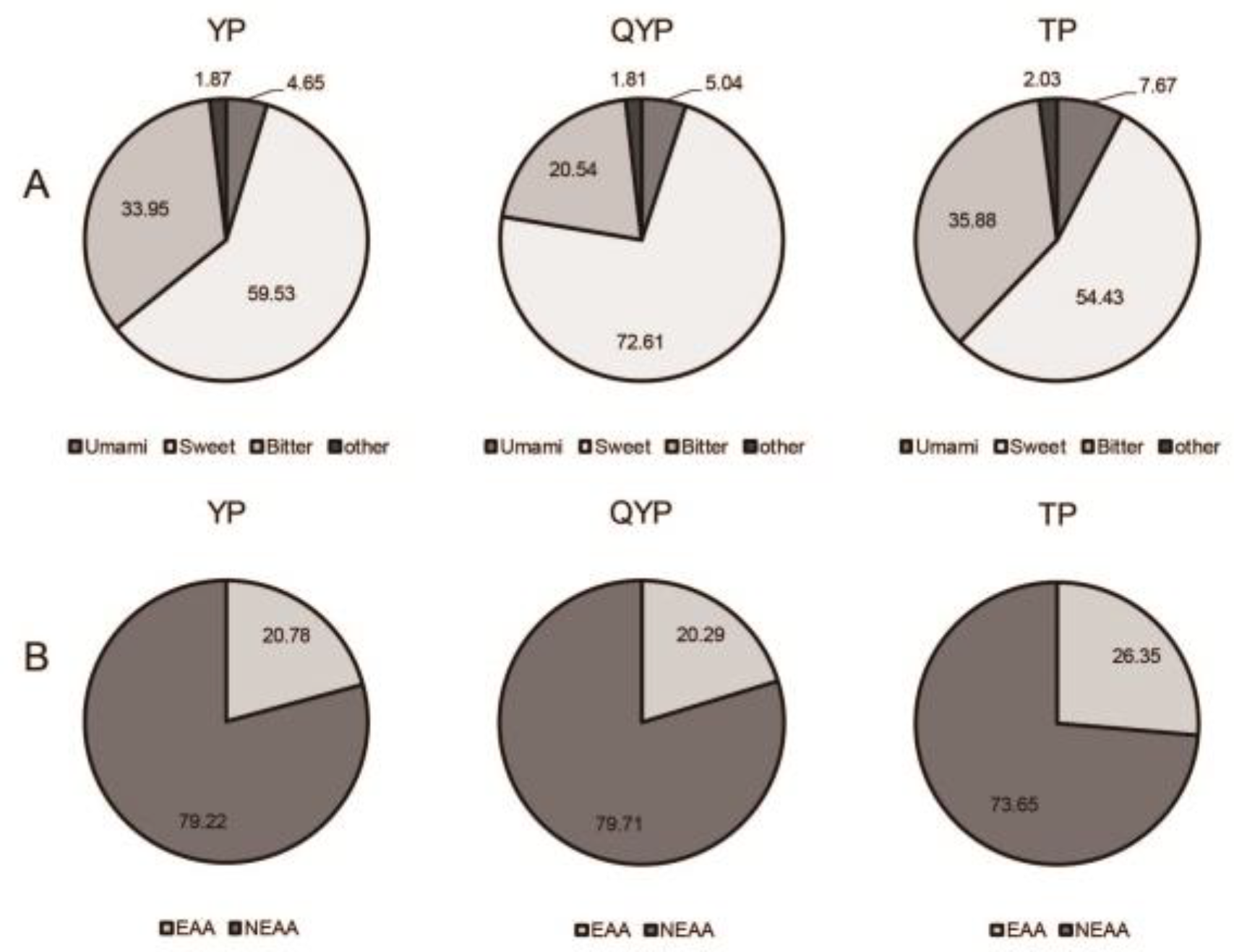
3.4. Free Amino Acid Content
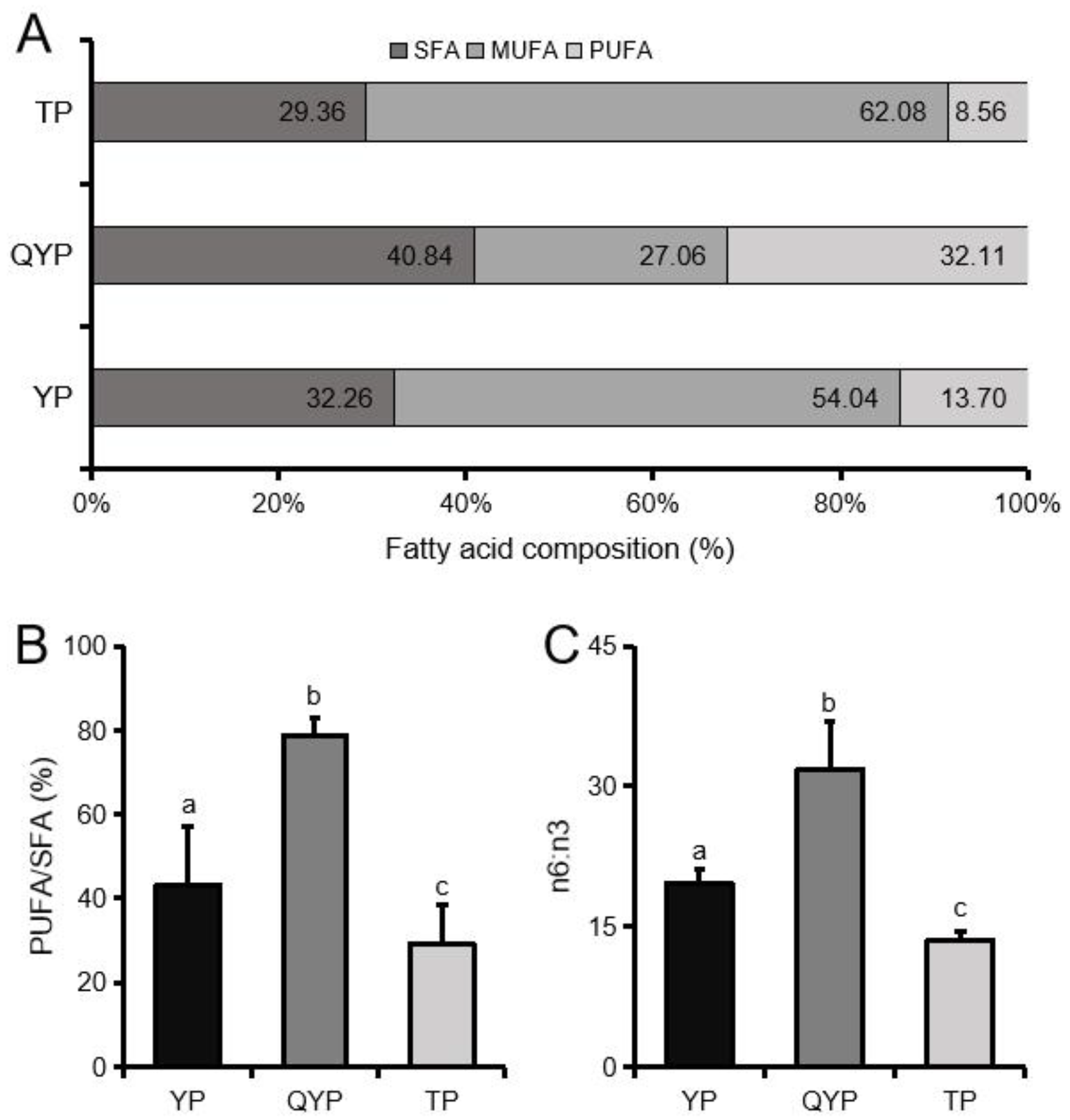
3.5. Fatty Acid Levels
4. Discussion
4.1. Plateau Adaptability of Tibetan Pigs
4.2. Carcass Traits and Meat Quality Differences between Yorkshire, Penzhou Mountain, and Tibetan Pigs
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Keenan, D.F. Pork meat quality, production and processing on. Encycl. Food Health 2016, 419–431. [Google Scholar] [CrossRef]
- Chen, G.; Cheng, X.; Shi, G.; Zou, C.; Chen, L.; Li, J.; Li, M.; Fang, C.; Li, C. Transcriptome analysis reveals the effect of long intergenic noncoding rnas on pig muscle growth and fat deposition. Biomed. Res. Int. 2019, 2019, 2951427. [Google Scholar] [CrossRef] [Green Version]
- Lebret, B.; Ecolan, P.; Bonhomme, N.; Méteau, K.; Prunier, A. Influence of production system in local and conventional pig breeds on stress indicators at slaughter, muscle and meat traits and pork eating quality. Animal 2015, 9, 1404–1413. [Google Scholar] [CrossRef] [Green Version]
- Luo, J.; Shen, L.; Tan, Z.; Cheng, X.; Yang, D.; Fan, Y.; Yang, Q.; Ma, J.; Tang, Q.; Jiang, A.A.; et al. Comparison reproductive, growth performance, carcass and meat quality of liangshan pig crossbred with duroc and berkshire genotypes and heterosis prediction. Livest. Sci. 2018, 212, 61–68. [Google Scholar] [CrossRef]
- Megens, H.J.; Crooijmans, R.P.M.A.; Cristobal, M.S.; Hui, X.; Li, N.; Groenen, M.A.M. Biodiversity of pig breeds from china and europe estimated from pooled DNA samples: Differences in microsatellite variation between two areas of domestication. Genet. Sel. Evol. 2008, 40, 103–128. [Google Scholar]
- Li, M.; Tian, S.; Jin, L.; Zhou, G.; Li, Y.; Zhang, Y.; Wang, T.; Yeung, C.K.; Chen, L.; Ma, J.; et al. Genomic analyses identify distinct patterns of selection in domesticated pigs and tibetan wild boars. Nat. Genet. 2013, 45, 1431–1438. [Google Scholar]
- Kong, X.; Dong, X.; Yang, S.; Qian, J.; Yang, J.; Jiang, Q.; Li, X.; Wang, B.; Yan, D.; Lu, S.; et al. Natural selection on tmprss6 associated with the blunted erythropoiesis and improved blood viscosity in tibetan pigs. Comp. Biochem. Physiol. Part B Biochem. Mol. Boil. 2019, 233, 11–22. [Google Scholar] [CrossRef]
- Peng, J.H.; Sun, D.; Nevo, E. Domestication evolution, genetics and genomics in wheat. Mol. Breed. 2011, 28, 281–301. [Google Scholar] [CrossRef]
- Luo, J.; Shen, L.Y.; Lei, H.G.; Zhu, K.P.; Jiang, Y.Z.; Bai, L.; Li, M.Z.; Tang, G.Q.; Li, X.W.; Zhang, S.H. Correlation between three glycometabolic-related hormones and muscle glycolysis as well as meat quality in three pig breeds. J. Sci. Food Agric. 2016, 97, 2706–2713. [Google Scholar] [CrossRef]
- Cheng, C.; Liu, Z.; Zhou, Y.; Wei, H.; Zhang, X.; Xia, M.; Deng, Z.; Zou, Y.; Jiang, S.; Peng, J. Effect of oregano essential oil supplementation to a reduced-protein, amino acid-supplemented diet on meat quality, fatty acid composition, and oxidative stability of longissimus thoracis muscle in growing-finishing pigs. Meat Sci. 2017, 133, 103–109. [Google Scholar] [CrossRef]
- Shen, L.; Lei, H.; Zhang, S.; Li, X.; Li, M.; Jiang, X.; Zhu, K.; Zhu, L. The comparison of energy metabolism and meat quality among three pig breeds. Anim. Sci. J. 2014, 85, 770–779. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative pcr and the 2(-delta delta c(t)) method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Wu, Q.; Hao, Y.; Fang, X.; Cheng, Y.; Dong, L.; Wei, W.; Gang, W.; Fu, H.; Liu, S.; Hao, L. The association of haplotypes in igfbp-3 gene promoter region and tissue expressions in three pig breeds. Anim. Cells. Syst. 2016, 20, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Yang, W.; Meng, F.; Peng, J.; Han, P.; Fang, F.; Ma, L.; Cao, B. Isolation and identification of a cellulolytic bacterium from the tibetan pig’s intestine and investigation of its cellulase production. Electron. J. Biotechnol. 2014, 17, 262–267. [Google Scholar] [CrossRef] [Green Version]
- Cai, X.H.; Huang, Y.T.; Zhang, Z.P.; Wang, Y.L. Hypoxia inducible factor-1 (hif-1) and its research advance in aquatic animals. J. Agric. Biotechnol. 2014, 22, 119–132. [Google Scholar]
- Hyun Ah, K.; Soyeon, L.; Hyung-Ho, M.; Sung Wan, K.; Ki-Chul, H.; Minhyung, L.; Hwa, K.S.; Donghoon, C. Hypoxia-inducible vascular endothelial growth factor gene therapy using the oxygen-dependent degradation domain in myocardial ischemia. Pharm. Res. 2010, 27, 2075–2084. [Google Scholar]
- Zhang, B.; Qiangba, Y.; Shang, P.; Lu, Y.; Yang, Y.; Wang, Z.; Zhang, H. Gene expression of vascular endothelial growth factor a and hypoxic adaptation in tibetan pig. J. Anim. Sci. Biotechnol. 2016, 4, 474–481. [Google Scholar] [CrossRef] [Green Version]
- Mancini, R.A.; Hunt, M.C. Current research in meat color. Meat Sci. 2005, 71, 100–121. [Google Scholar] [CrossRef]
- Schiaffino, S.; Reggiani, C. Fiber types in mammalian skeletal muscles. Physiol. Rev. 2011, 91, 1447–1531. [Google Scholar] [CrossRef] [Green Version]
- Avivi, A.; Band, M.; Joel, A.; Shenzer, P.; Coleman, R. Adaptive features of skeletal muscles of mole rats (spalax ehrenbergi) to intensive activity under subterranean hypoxic conditions. Acta Histochem. 2009, 111, 415–419. [Google Scholar] [CrossRef]
- Shen, Q.W.; Means, W.J.; Thompson, S.A.; Underwood, K.R.; Zhu, M.J.; Mccormick, R.J.; Ford, S.P.; Du, M. Pre-slaughter transport, amp-activated protein kinase, glycolysis, and quality of pork loin. Meat Sci. 2006, 74, 388–395. [Google Scholar] [CrossRef]
- Wang, F.-H.; Liu, J.; Deng, Q.-J.; Qi, Y.; Wang, M.; Wang, Y.; Zhang, X.-G.; Zhao, D. Association between plasma essential amino acids and atherogenic lipid profile in a chinese population: A cross-sectional study. Atherosclerosis 2019, 286, 7–13. [Google Scholar] [CrossRef]
- Brestenský, M.; Nitrayová, S.; Patráš, P.; Heger, J.; Nitray, J. Branched chain amino acids and their importance in nutrition. JMBFS 2015, 5, 197–202. [Google Scholar] [CrossRef]
- Gannon, N.P.; Schnuck, J.K.; Vaughan, R.A. Bcaa metabolism and insulin sensitivity—Dysregulated by metabolic status? Mol. Nutr. Food Res. 2018, 62, e1700756. [Google Scholar] [CrossRef] [PubMed]
- Guido, F.; Rita, R.; Michela, T.; Gianluca, D.; Umberto, B.; Giovanni Battista, F. Tryptophan-derived catabolites are responsible for inhibition of t and natural killer cell proliferation induced by indoleamine 2,3-dioxygenase. J. Exp. Med. 2002, 196, 459–468. [Google Scholar]
- Fallarino, F.; Grohmann, U.; Vacca, C.; Bianchi, R.; Orabona, C.; Spreca, A.; Fioretti, M.C.; Puccetti, P. T cell apoptosis by tryptophan catabolism. Cell Death Differ. 2002, 9, 1069–1077. [Google Scholar] [CrossRef]
- Rey, B.; Dégletagne, C.; Bodennec, J.; Monternier, P.-A.; Mortz, M.; Roussel, D.; Romestaing, C.; Rouanet, J.-L.; Tornos, J.; Duchamp, C. Hormetic response triggers multifaceted anti-oxidant strategies in immature king penguins (aptenodytes patagonicus). Free Radic. Biol. Med. 2016, 97, 577–587. [Google Scholar] [CrossRef]
- Sakamoto, A.; Sulpice, R.; Hou, C.X.; Kinoshita, M.; Higashi, S.I.; Kanaseki, T.; Nonaka, H.; Moon, B.Y.; Murata, N. Genetic modification of the fatty acid unsaturation of phosphatidylglycerol in chloroplasts alters the sensitivity of tobacco plants to cold stress. Plant Cell Environ. 2010, 27, 99–105. [Google Scholar] [CrossRef]
- Králová, S. Role of fatty acids in cold adaptation of antarctic psychrophilic flavobacterium spp. Syst. Appl. Microbiol. 2017, 40, 329–333. [Google Scholar] [CrossRef]
- Mao, Z.-X.; Fu, H.; Nan, Z.-B.; Wang, J.; Wan, C.-G. Fatty acid content of common vetch (vicia sativa l.) in different regions of northwest china. Biochem. Syst. Ecol. 2012, 44, 347–351. [Google Scholar] [CrossRef]
- Sobczyńska, M.; Blicharski, T.; Tyra, M. Relationships between longevity, lifetime productivity, carcass traits and conformation in polish maternal pig breeds. J. Anim. Breed. Genet. 2014, 130, 361–371. [Google Scholar]
- Qiao, M.; Huang, J.; Wu, H.; Wu, J.; Peng, X.; Mei, S. Molecular characterization, transcriptional regulation and association analysis with carcass traits of porcine tcap gene. Gene 2014, 538, 273–279. [Google Scholar] [CrossRef] [PubMed]
- ÇInar, M.U.; Fan, H.T. The mrna expression pattern of skeletal muscle regulatory factors in divergent phenotype swine breeds. Kafkas Univ. Vet. Fak. 2012, 18, 685–690. [Google Scholar]
- Buys, N. Characterization of the complete porcine mstn gene and expression levels in pig breeds differing in muscularity. Anim. Genet. 2010, 39, 586–596. [Google Scholar]
- Rahelic, S.; Puac, S. Fibre types in longissimus dorsi from wild and highly selected pig breeds. Meat Sci. 1981, 5, 439–450. [Google Scholar] [CrossRef]
- Cho, E.S.; Lee, K.T.; Kim, J.M.; Lee, S.W.; Jeon, H.J.; Lee, S.H.; Hong, K.C.; Kim, T.H. Association of a single nucleotide polymorphism in the 5’ upstream region of the porcinemyosin heavy chain 4gene with meat quality traits in pigs. Anim. Sci. J. 2016, 87, 330–335. [Google Scholar] [CrossRef]
- Larzul, C. Pig genetics: Insight in minipigs. In Proceedings of the Bilateral Symposium on Miniature Pigs for Biomedical Research in Taiwan and France, Hsinhua, Taiwan, 22–23 October 2013; pp. 1–6. [Google Scholar]
- Li, X.; Xia, A.-Q.; Chen, L.-J.; Du, M.-T.; Chen, L.; Kang, N.; Zhang, D.-Q. Effects of lairage after transport on post mortem muscle glycolysis, protein phosphorylation and lamb meat quality. J. Integr. Agric. 2018, 17, 2336–2344. [Google Scholar] [CrossRef] [Green Version]
- Legeard, D.; Marty-Mahe, P.; Camillerapp, J.; Marchal, P.; Leredde, C. Real-time quality evaluation of pork hams by color machine vision. Proc. SPIE Int. Soc. Opt. Eng. 1999, 3652, 138–149. [Google Scholar]
- Gajewczyk, P.; Gajewczyk, B.; Akin´Cza, J.; Szman´Ko, T. Influence of crossing polish and foreign pig breeds on physicochemical traits of longissimus lumborum muscle. Turk. J. Vet. Anim. Sci. 2014, 38, 183–188. [Google Scholar] [CrossRef]
- Zhou, W.; Sun-Waterhouse, D.; Xiong, J.; Cui, C.; Wang, W.; Dong, K. Desired soy sauce characteristics and autolysis of aspergillus oryzae induced by low temperature conditions during initial moromi fermentation. J. Food Sci. Technol. 2019, 56, 2888–2898. [Google Scholar] [CrossRef]
- Gao, X.; Zhang, J.; Regenstein, J.M.; Yin, Y.; Zhou, C. Characterization of taste and aroma compounds in tianyou, a traditional fermented wheat flour condiment. Food Res. Int. 2018, 106, 156–163. [Google Scholar] [CrossRef]
- Pallottini, A.C.; Sales, C.H.; Dads, V.; Marchioni, D.M.; Fisberg, R.M. Dietary bcaa intake is associated with demographic, socioeconomic and lifestyle factors in residents of são paulo, brazil. Nutrients 2017, 9, 449. [Google Scholar] [CrossRef] [Green Version]
- Elroy, N.N.; Rogers, J.; Mafi, G.G.; VanOverbeke, D.L.; Hartson, S.D.; Ramanathan, R. Species-specific effects on non-enzymatic metmyoglobin reduction in vitro. Meat Sci. 2015, 105, 108–113. [Google Scholar] [CrossRef] [PubMed]
- Mensink, R.P. Fatty acids: Health effects of saturated fatty acids. In Encyclopedia of Human Nutrition, 3rd ed.; Academic Press: London, UK, 2013; pp. 215–219. [Google Scholar]
- Doychev, V.; Mihaylova, G. Effect of flaxsee d and alpha tocopherol supplementation of pig diets on fatty acid content and lipid oxidation stability of m. Longissimus. Bulg. J. Agric. Sci. 2013, 19, 1416–1424. [Google Scholar]
- Tapiero, H.; Ba, G.N.; Couvreur, P.; Tew, K.D. Polyunsaturated fatty acids (pufa) and eicosanoids in human health and pathologies. Biomed. Pharmacother. 2002, 56, 215–222. [Google Scholar] [CrossRef]
- Starkey, C.P.; Geesink, G.H.; Collins, D.; Hutton Oddy, V.; Hopkins, D.L. Do sarcomere length, collagen content, ph, intramuscular fat and desmin degradation explain variation in the tenderness of three ovine muscles? Meat Sci. 2016, 113, 51–58. [Google Scholar] [CrossRef]
- Liu, L.; Ngadi, M.O. Predicting intramuscular fat content of pork using hyperspectral imaging. J. Food Eng. 2014, 134, 16–23. [Google Scholar] [CrossRef]
- Li, W.Z.; Zhao, S.M.; Huang, Y.; Yang, M.H.; Pan, H.B.; Zhang, X.; Ge, C.R.; Gao, S.Z. Expression of lipogenic genes during porcine intramuscular preadipocyte differentiation. Res. Vet. Sci. 2012, 93, 1190–1194. [Google Scholar] [CrossRef] [PubMed]
- Chen, Q.M.; Wang, H.; Zeng, Y.Q.; Chen, W. Developmental changes and effect on intramuscular fat content of h-fabp and a-fabp mrna expression in pigs. J. Appl. Genet. 2013, 54, 119–123. [Google Scholar] [CrossRef]
- Jiang, Z.; Michal, J.J.; Tobey, D.J.; Daniels, T.F.; Rule, D.C.; Macneil, M.D. Significant associations of stearoyl-coa desaturase (scd1) gene with fat deposition and composition in skeletal muscle. Int. J. Biol. Sci. 2008, 4, 345–351. [Google Scholar]
- Rogowski, M.P.; Flowers, M.T.; Stamatikos, A.D.; Ntambi, J.M.; Paton, C.M. Scd1 activity in muscle increases triglyceride pufa content, exercise capacity, and pparδ expression in mice. J. Lipid Res. 2013, 54, 2636–2646. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Carcass traits | YP (n = 39) | QYP1 (n = 5) | QYP2 (n = 12) | TP (n = 24) | Significance |
|---|---|---|---|---|---|
| Carcass weight, kg | 84.60 ± 9.39 a | 44.64 ± 3.99 b | 70.32 ± 16.96 a | 28.88 ± 4.04 c | <0.001 |
| Carcass length, cm | 81.09 ± 2.27 a | 64.4 ± 2.97 b | 73.83 ± 11.74 a | 57.4 ± 2.71 c | <0.001 |
| Dressing, % | 74.87 ± 3.05 a | 66.85 ± 5.03 b | 71.61 ± 7.8 ab | 69.69 ± 2.23 b | 0.002 |
| Bone rate, % | 13.1 ± 2.07 a | 9.26 ± 1.78 b | 11.41 ± 3.57 ab | 10.84 ± 1.62 b | 0.037 |
| Carcass lean, % | 64.11 ± 5.59 a | 43.32 ± 2.94 bc | 40.68 ± 5.12 b | 47.87 ± 3.63 c | <0.001 |
| Carcass fat, % | 22.79 ± 4.80 a | 47.34 ± 3.96 b | 47.79 ± 8.41 b | 41.16 ± 3.64 c | <0.001 |
| Back fat thickness, cm | 1.89 ± 0.48 a | 3.33 ± 0.54 b | 4.14 ± 1.14 c | 2.59 ± 0.5d | <0.001 |
| Loin eye area, cm2 | 49.39 ± 8.27 a | 17.84 ± 3.67 bc | 20.85 ± 3.42 c | 14.08 ± 3.48 b | <0.001 |
| Marbling scores | 1.05 ± 0.5 a | 2.63 ± 0.48 b | 3.13 ± 0.53 bc | 2.46 ± 0.45 b | <0.001 |
| Meat quality | YP (n = 32) | QYP1 (n = 5) | QYP2 (n = 12) | TP (n = 12) | Significance |
|---|---|---|---|---|---|
| PH45min | 6.28 ± 0.17 a | 6.8 ± 0.31 b | 6.6 ± 0.3 bc | 6.51 ± 0.15 c | <0.001 |
| PH24h | 5.7 ± 0.29 a | 6.11 ± 0.17 b | 5.92 ± 0.33 b | 6 ± 0.11 b | 0.002 |
| L*45min | 46.99 ± 1.98 a | 43.65 ± 3.25 b | 41.03 ± 2.76 b | 38.84 ± 2.7 c | <0.001 |
| a*45min | 5.93 ± 1.15 a | 8.42 ± 2.08 b | 10.59 ± 2.99 b | 12.57 ± 2.27 c | <0.001 |
| b*45min | 6.75 ± 0.73 a | 0.6 ± 0.42 b | 1.7 ± 1.57 c | 3.14 ± 0.64d | <0.001 |
| L*24h | 52.63 ± 3.31 a | 47.71 ± 4.61 b | 45.77 ± 2.46 b | 44.89 ± 4 b | <0.001 |
| a*24 | 9.82 ± 1.94 a | 8.83 ± 1.97 a | 11.05 ± 2.96 ab | 12.14 ± 2.73 b | 0.026 |
| b*24h | 7.83 ± 0.91 a | 3.49 ± 1.21 b | 4.97 ± 2.03 b | 5.11 ± 2.57 b | <0.001 |
| Drip loss, % | 3.75 ± 1.81 a | 1.91 ± 0.48 b | 2.12 ± 0.59 b | 2.37 ± 0.35 b | 0.013 |
| Cooking loss, % | 34.40 ± 1.82 | 33.96 ± 2.29 | 36.94 ± 8.23 | 33.26 ± 4.08 | 0.341 |
| Shear force, kg | 9.9 ± 3.49 | 6.05 ± 2.68 | 6.44 ± 2 | 4.7 ± 1.86 | 0.694 |
| Crude protein, % | 21.58 ± 3.77 ab | 24.22 ± 0.52 a | 21.89 ± 0.96 ab | 20.27 ± 2.36 b | 0.077 |
| Intramuscular fat, % | 1.43 ± 0.55 a | 2.48 ± 0.3 b | 4.62 ± 1.85 c | 1.88 ± 0.33 ab | <0.001 |
| Ash, % | 2.19 ± 0.5 a | 1.35 ± 0.07 b | 1.15 ± 0.06 b | 2.73 ± 1.47 a | <0.001 |
| AA | YP (n = 6) | QYP (n = 6) | TP (n = 6) | p Value |
|---|---|---|---|---|
| Ile | 2.09 ± 0.33 a | 2.16 ± 0.29 a | 3.69 ± 0.31 b | <0.001 |
| Leu | 2.76 ± 0.23 a | 3.47 ± 0.48 b | 4.01 ± 0.52 c | 0.001 |
| Lys | 5.95 ± 0.5 a | 3.6 ± 0.53 b | 3.8 ± 0.48 b | <0.001 |
| Thr | 2.93 ± 0.26 a | 3.53 ± 0.4 b | 3.76 ± 0.32 b | 0.002 |
| Val | 2.62 ± 0.32 a | 3.49 ± 0.36 b | 4.18 ± 0.39 c | <0.001 |
| Trp | 0.93 ± 0.09 a | 0.25 ± 0.04 b | 1.05 ± 0.10 c | <0.001 |
| Met | 1.33 ± 0.11 a | 0.97 ± 0.38 b | 3 ± 0.28 c | <0.001 |
| Phe | 2.18 ± 0.44 a | 2.61 ± 0.52 ab | 2.77 ± 0.21 b | 0.063 |
| Arg | 4.02 ± 0.55 a | 2.92 ± 0.49 b | 3.89 ± 0.57 a | 0.005 |
| His | 15.31 ± 1.29 a | 2.69 ± 0.26 b | 8.36 ± 1.1 c | <0.001 |
| Asn | 1.86 ± 0.17 | 1.8 ± 0.35 | 2.03 ± 0.11 | 0.246 |
| Ser | 2.58 ± 0.2 a | 3.19 ± 0.54 b | 3.93 ± 0.46 c | <0.001 |
| Gly | 7.36 ± 1.53 | 7.61 ± 1.09 | 8.01 ± 1.56 | 0.727 |
| Ala | 20.22 ± 2.25 ab | 21.73 ± 2.11 a | 17.82 ± 2.5 b | 0.031 |
| Tyr | 2.73 ± 0.11 a | 1.79 ± 0.26 b | 4.83 ± 0.34 c | <0.001 |
| Gln | 17.52 ± 2.61 a | 30.9 ± 4.33 b | 14.12 ± 0.97 a | <0.001 |
| Glu | 3.96 ± 1.44 a | 4.21 ± 0.39 a | 7.44 ± 0.86 b | <0.001 |
| Asp | 0.65 ± 0.15 ab | 0.86 ± 0.82 a | 0.19 ± 0.09 b | 0.079 |
| Pro | 3.00 ± 0.13 a | 2.21 ± 0.32 b | 3.13 ± 0.31 a | <0.001 |
| FAA | YP (n = 6) | QYP (n = 6) | TP (n = 6) | p Value |
|---|---|---|---|---|
| C10:0 | 0.07 ± 0.01 | 0.06 ± 0.05 | 0.06 ± 0.01 | 0.689 |
| C12:0 | 0.07 ± 0.01 a | 0.01 ± 0.02 b | 0.05 ± 0.01 a | <0.001 |
| C14:0 | 1.33 ± 0.13 | 1.31 ± 0.18 | 1.34 ± 0.14 | 0.964 |
| C15:0 | 0.03 ± 0.01 a | 0.10 ± 0.01 b | 0.03 ± 0.02 a | <0.001 |
| C16:0 | 13.95 ± 0.77 a | 22.90 ± 0.69 b | 15.60 ± 0.86 c | <0.001 |
| C16:1 | 3.13 ± 0.48 a | 3.40 ± 0.41 a | 5.60 ± 1.04 b | <0.001 |
| C17:0 | 0.17 ± 0.04 a | 0.35 ± 0.02 b | 0.15 ± 0.05 a | <0.001 |
| C17:1 | —— | 0.28 ± 0.01 a | 0.17 ± 0.04 b | <0.001 |
| C18:0 | 16.37 ± 1.72 a | 15.81 ± 0.47 a | 11.77 ± 0.53 b | <0.001 |
| C18:1n9 | 49.80 ± 3.02 a | 22.79 ± 1.8 b | 54.81 ± 3.34 c | <0.001 |
| C18:2n6 | 9.91 ± 2.53 a | 21.46 ± 2.04 b | 6.48 ± 1.65 c | <0.001 |
| C18:3n3 | 0.56 ± 0.14 a | 0.60 ± 0.09 a | 0.33 ± 0.07 b | <0.001 |
| C20:0 | 0.24 ± 0.09 a | 0.17 ± 0.02 a | 0.32 ± 0.05 b | 0.002 |
| C20:1 | 1.09 ± 0.25 a | 0.55 ± 0.1 b | 1.41 ± 0.12 c | <0.001 |
| C20:2 | 0.58 ± 0.12 a | 0.52 ± 0.08 a | 0.4 ± 0.07 b | 0.016 |
| C20:3n3 | 0.06 ± 0.01 a | 0.06 ± 0.01 a | 0.24 ± 0.12 b | 0.001 |
| C20:4n6 | 2.56 ± 1.21 a | 9.13 ± 1.03 b | 1.08 ± 0.61 c | <0.001 |
| C20:5n3 | 0.04 ± 0.02 a | 0.23 ± 0.04 b | 0.03 ± 0.02 a | <0.001 |
| C22:0 | 0.02 ± 0.01 a | 0.04 ± 0.00 b | 0.02 ± 0.00 a | <0.001 |
| C22:1n9 | 0.02 ± 0.01 a | 0.03 ± 0.00 a | 0.09 ± 0.02 b | <0.001 |
| C23:0 | —— | 0.04 ± 0.01 | —— | —— |
| C24:0 | 0.02 ± 0.01 a | 0.05 ± 0 b | 0.02 ± 0.01 a | <0.001 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gan, M.; Shen, L.; Fan, Y.; Guo, Z.; Liu, B.; Chen, L.; Tang, G.; Jiang, Y.; Li, X.; Zhang, S.; et al. High Altitude Adaptability and Meat Quality in Tibetan Pigs: A Reference for Local Pork Processing and Genetic Improvement. Animals 2019, 9, 1080. https://doi.org/10.3390/ani9121080
Gan M, Shen L, Fan Y, Guo Z, Liu B, Chen L, Tang G, Jiang Y, Li X, Zhang S, et al. High Altitude Adaptability and Meat Quality in Tibetan Pigs: A Reference for Local Pork Processing and Genetic Improvement. Animals. 2019; 9(12):1080. https://doi.org/10.3390/ani9121080
Chicago/Turabian StyleGan, Mailin, Linyuan Shen, Yuan Fan, Zhixian Guo, Bin Liu, Lei Chen, Guoqing Tang, Yanzhi Jiang, Xuewei Li, Shunhua Zhang, and et al. 2019. "High Altitude Adaptability and Meat Quality in Tibetan Pigs: A Reference for Local Pork Processing and Genetic Improvement" Animals 9, no. 12: 1080. https://doi.org/10.3390/ani9121080
APA StyleGan, M., Shen, L., Fan, Y., Guo, Z., Liu, B., Chen, L., Tang, G., Jiang, Y., Li, X., Zhang, S., Bai, L., & Zhu, L. (2019). High Altitude Adaptability and Meat Quality in Tibetan Pigs: A Reference for Local Pork Processing and Genetic Improvement. Animals, 9(12), 1080. https://doi.org/10.3390/ani9121080