The Impact of Heat Load on Cattle

 ,
,  ,
, 
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Climate Change
3. Heat Wave Events
- Hahn [20] reported the loss of 100 feedlot cattle in central Nebraska over a heat wave that had three spikes in thermal loads. Deaths occurred during the third spike where it was hypothesized that ad libitum feed intake resulted in large metabolic heat load and in conjunction with environmental heat load, surpassed the animals’ ability to maintain thermal balance [20]
- February 2000–1255 cattle died in southwestern New South Wales with deaths occurring after a rainfall event where climatic conditions presented high relative humidity and high overnight ambient temperature [22]
- June 2017–4000 to 6000 dairy cows died in Fresno, Kings and Tulare counties USA [27] during a heat wave
4. Defining Heat Load
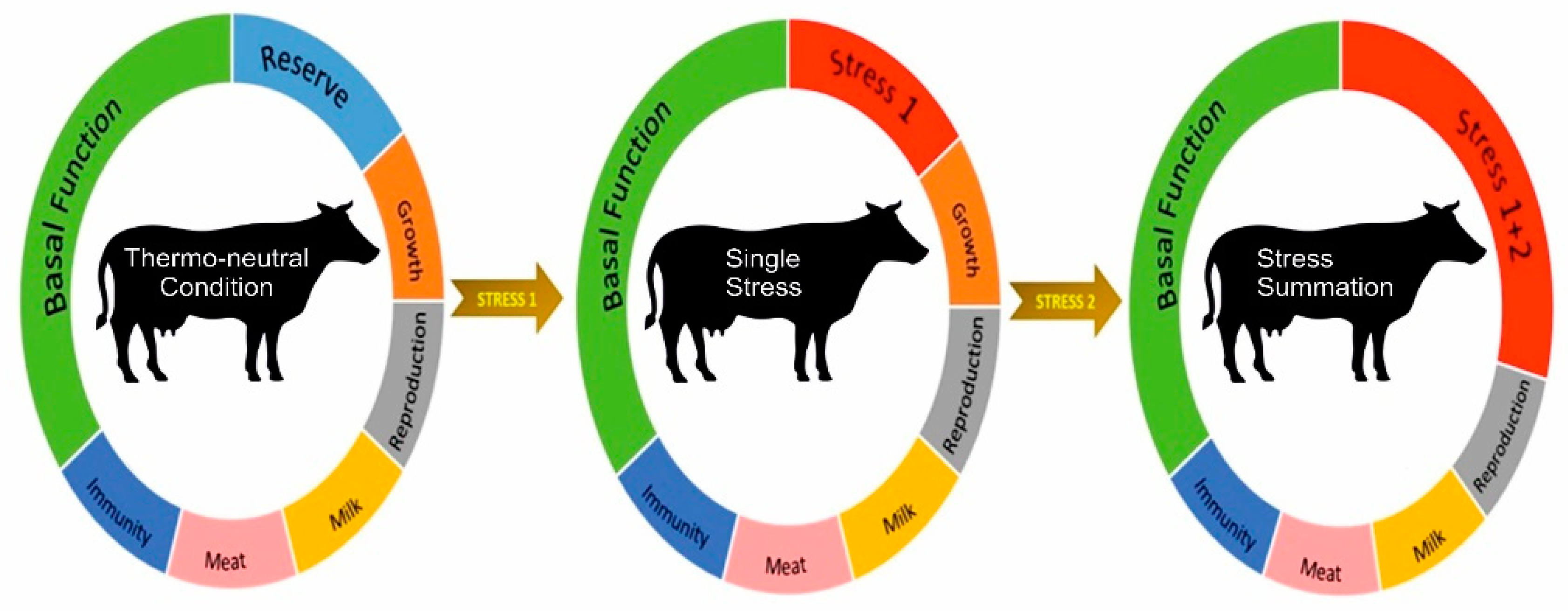
Multiple Stressors
5. Implications of Hot Environmental Conditions
5.1. Nutrition and Eating Behavior
5.2. Water Intake
5.3. Metabolic Dysfunction
5.4. Body Temperature
5.5. Reproduction
5.5.1. Impact on Males
5.5.2. Impact on Females
5.6. Health
5.7. Productivity
5.7.1. Growth
5.7.2. Milk Production and Composition
5.7.3. Dark Cutting Beef
6. Mitigation Opportunities
6.1. Shade Structures
6.2. Nutrition
6.3. Genetics
6.4. Genomic Selection for Heat Tolerance
7. Adaptation and Acclimation
7.1. Acclimation
7.2. Adaptation
8. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Ames, D. Thermal Environment Affects Production Efficiency of Livestock. BioScience 1980, 30, 457–460. [Google Scholar] [CrossRef]
- Mader, T.L.; Griffin, D. Management of Cattle Exposed to Adverse Environmental Conditions. Vet. Clin. N. Am. Food Anim. Pract. 2015, 31, 247–258. [Google Scholar] [CrossRef] [PubMed]
- Belasco, E.J.; Cheng, Y.; Schroeder, T.C. The impact of extreme weather on cattle feeding profits. J. Agric. Resour. Econ. 2015, 40, 285–305. [Google Scholar]
- St-Pierre, N.R.; Cobanov, B.; Schnitkey, G. Economic Losses from Heat Stress by US Livestock Industries. J. Dairy Sci. 2003, 86, E52–E77. [Google Scholar] [CrossRef] [Green Version]
- Sackett, D.; Holmes, P.; Abbot, K.; Jephcott, S.; Barber, M. Assessing the Economic Cost of Endemic Disease on the Profitability of Australian Beef Cattle and Sheep Producers; MLA Final Report AHW.087; Meat and Livestock Australia: Sydney, Australia, 2006. [Google Scholar]
- Sejian, V.; Bhatta, R.; Soren, N.M.; Malik, P.K.; Ravindra, J.P.; Prasad, C.; Lal, R. Introduction to Concepts of Climate Change Impact on Livestock and Its Adaptation and Mitigation. In Climate Change Impact on Livestock: Adaptation and Mitigation; Sejian, V., Gaughan, J., Baumgard, L., Prasad, C., Eds.; Springer: New Delhi, India, 2015; pp. 1–23. [Google Scholar] [CrossRef]
- Angilletta, M.J., Jr. Thermal Acclimation. In Thermal Adaptation: A Theoretical and Empirical Synthesis; Oxford University Press Inc.: New York, NY, USA, 2009; pp. 127–156. [Google Scholar]
- Gaughan, J.; Cawdell-Smith, A.J. Impact of Climate Change on Livestock Production and Reproduction. In Climate Change Impact on Livestock: Adaptation and Mitigation; Sejian, V., Gaughan, J., Baumgard, L., Prasad, C., Eds.; Springer: New Delhi, India, 2015; Volume 4, pp. 51–60. [Google Scholar] [CrossRef]
- Nardone, A.; Ronchi, B.; Lacetera, N.; Ranieri, M.S.; Bernabucci, U. Effects of climate changes on animal production and sustainability of livestock systems. Livest. Sci. 2010, 130, 57–69. [Google Scholar] [CrossRef]
- Nidumolu, U.; Crimp, S.; Gobbett, D.; Laing, A.; Howden, M.; Little, S. Spatio-temporal modelling of heat stress and climate change implications for the Murray dairy region, Australia. Int. J. Biometeorol. 2014, 58, 1095–1108. [Google Scholar] [CrossRef] [PubMed]
- Hennessey, K.; Fitzharris, B.; Bates, B.C.; Harvey, N.; Howden, S.M.; Hughes, L.; Salinger, J.; Warrick, R. Australia and New Zealand. In Climate Change 2007: Impacts, Adaptation, Vulnerability. Contribution of Working Group II to the Fourth Assessment Report of the Intergovernmental Panel on Climate Change; Parry, M.L., Canziani, O.F., Palutikof, J.P., van der Linden, P.J., Hanson, C.E., Eds.; Cambride University Press: Cambridge, UK, 2007; pp. 507–540. [Google Scholar]
- Henry, B.; Charmley, E.; Eckard, R.; Gaughan, J.B.; Hegarty, R. Livestock production in a changing climate: Adaptation and mitigation research in Australia. Crop Pasture Sci. 2012, 63, 191–202. [Google Scholar] [CrossRef]
- Nienaber, J.A.; Hahn, G.L.; Brown-Brandl, T.M.; Eigenberg, R.A. Summer Heat Waves—Extreme Years. In Proceedings of the ASABE Annual International Meeting, Minneapolis, Minnesota, 17–20 June 2007. [Google Scholar]
- Mader, T.L.; Gaughan, J.B.; Johnson, L.J.; Hahn, G.L. Tympanic temperature in confined beef cattle exposed to excessive heat load. Int. J. Biometeorol. 2010, 54, 629–635. [Google Scholar] [CrossRef]
- Solomon, S.; Qin, D.; Manning, M.; Marquis, M.; Averyt, K.; Tignore, M.M.B. (Eds.) Climate Change 2007: The Physical Science Basis; Cambridge University Press: New York, NY, USA, 2007. [Google Scholar]
- Westcott, N.E. The Prolonged 1954 Midwestern, U.S. Heat Wave: Impacts and Responses. Weather. Clim. Soc. 2011, 3, 165–176. [Google Scholar] [CrossRef]
- Robinson, P.J. On the Definition of a Heat Wave. J. Appl. Meteorol. 2001, 40, 762–775. [Google Scholar] [CrossRef]
- Mader, T.L. Environmental stress in confined beef cattle. J. Anim. Sci. 2003, 81, E110–E119. [Google Scholar]
- Blackshaw, J.; Blackshaw, A. Heat stress in cattle and the effect of shade on production and behaviour: A review. Aust. J. Exp. Agric. 1994, 34, 285–295. [Google Scholar] [CrossRef]
- Hahn, G.L. Dynamic Responses of Cattle to Thermal Heat Loads. J. Anim. Sci. 1999, 77 (Suppl. 2), 10–20. [Google Scholar] [CrossRef]
- Gaughan, J.B. Respiration Rate and Rectal Temperature Responses of Feedlot Cattle in Dynamic, Thermally Challenging Environments; The University of Queensland Gatton: Queensland, Australia, 2002. [Google Scholar]
- Entwistle, K.; Rose, M.; McKiernan, B. Mortalities in Feedlot Cattle at Prime City Feedlot, Tabbita, NSW, February 2000; NSW Agriculture Sydney: Sydney, Australia, 2000. [Google Scholar]
- Bushby, D.; Loy, D. Heat Stress in Feedlot Cattle: Producer Survey Results. Beef Research Report, 1996. Paper 26. 1997. Available online: http://lib.dr.iastate.edu/beefreports_1996/1926 (accessed on 6 March 2014).
- Hahn, G.L.; Mader, T.L. Heat Waves in Relation to Thermoregulation, Feeding Behaviour and Mortality of Feedlot Cattle. In Livestock Environment V, Proceedings of the Fifth International Symposium; Bottcher, R.W., Hoff, S.J., Eds.; American Society of Agricultural Engineers: St. Joseph, MI, USA, 1997; Volume I, pp. 563–571. [Google Scholar]
- Brown-Brandl, T.M.; Nienaber, J.A.; Eigenberg, R.A.; Mader, T.L.; Morrow, J.L.; Dailey, J.W. Comparison of heat tolerance of feedlot heifers of different breeds. Livest. Sci. 2006, 105, 19–26. [Google Scholar] [CrossRef] [Green Version]
- Brown-Brandl, T.M.; Eigenberg, R.A.; Nienaber, J.A. Heat stress risk factors of feedlot heifers. Livest. Sci. 2006, 105, 57–68. [Google Scholar] [CrossRef] [Green Version]
- Associated Press. Thousands of Cows Die in California Heat Wave; Disposing Them Becomes a Problem. Available online: https://www.latimes.com/local/lanow/la-me-cattle-deaths-20170708-story.html (accessed on 12 April 2019).
- Buffington, D.; Collazo-Arocho, A.; Canton, G.; Pitt, D.; Thatcher, W.; Collier, R. Black Globe-Humidity Index (BGHI) as a Comfort Equation for Dairy Cows. Trans. Am. Soc. Agric. Eng. 1981, 27, 711–714. [Google Scholar] [CrossRef]
- Gaughan, J.B.; Mader, T.L.; Holt, S.M.; Sullivan, M.L.; Hahn, G.L. Assessing the heat tolerance of 17 beef cattle genotypes. Int. J. Biometeorol. 2010, 54, 617–627. [Google Scholar] [CrossRef]
- Sejian, V.; Bhatta, R.; Gaughan, J.B.; Dunshea, F.R.; Lacetera, N. Review: Adaptation of animals to heat stress. Animal 2018, 12, s431–s444. [Google Scholar] [CrossRef] [Green Version]
- Gaughan, J.; Kreikemeier, W.; Mader, T. Hormonal growth-promotant effects on grain-fed cattle maintained under different environments. Int. J. Biometeorol. 2005, 49, 396–402. [Google Scholar] [CrossRef]
- Sejian, V.; Kumar, D.; Gaughan, J.B.; Naqvi, S.M.K. Effect of multiple environmental stressors on the adaptive capability of Malpura rams based on physiological responses in a semi-arid tropical environment. J. Vet. Behav. Clin. Appl. Res. 2017, 17, 6–13. [Google Scholar] [CrossRef]
- Sejian, V.; Maurya, V.P.; Naqvi, S.M. Effect of walking stress on growth, physiological adaptability and endocrine responses in Malpura ewes in a semi-arid tropical environment. Int. J. Biometeorol. 2012, 56, 243–252. [Google Scholar] [CrossRef]
- Sejian, V.; Maurya, V.P.; Naqvi, S.M.K. Effect of thermal stress, restricted feeding and combined stresses (thermal stress and restricted feeding) on growth and plasma reproductive hormone levels of Malpura ewes under semi-arid tropical environment. J. Anim. Physiol. Anim. Nutr. 2011, 95, 252–258. [Google Scholar] [CrossRef]
- Shilja, S.; Sejian, V.; Bagath, M.; Mech, A.; David, C.; Kurien, E.; Varma, G.; Bhatta, R. Adaptive capability as indicated by behavioral and physiological responses, plasma HSP70 level, and PBMC HSP70 mRNA expression in Osmanabadi goats subjected to combined (heat and nutritional) stressors. Int. J. Biometeorol. 2016, 60, 1311–1323. [Google Scholar] [CrossRef]
- Kumar, D.; Sejian, V.; Gaughan, J.B.; Naqvi, S.M.K. Biological functions as affected by summer season-related multiple environmental stressors (heat, nutritional and walking stress) in Malpura rams under semi-arid tropical environment. Biol. Rhythm Res. 2017, 48, 593–606. [Google Scholar] [CrossRef]
- Abdul Niyas, P.; Sejian, V.; Bagath, M.; Parthipan, S.; Selvaraju, S.; Manjunathareddy, G.; Kurien, E.; Varma, G.; Bhatta, R. Effect of heat and nutritional stress on growth and testicular HSP70 expression in goats. J. Agrometeorol. 2017, 19, 189–194. [Google Scholar]
- Sejian, V.; Maurya, V.P.; Kumar, K.; Naqvi, S.M.K. Effect of multiple stresses on growth and adaptive capability of Malpura ewes under semi-arid tropical environment. Trop. Anim. Health Prod. 2012, 45, 107–116. [Google Scholar] [CrossRef]
- Eigenberg, R.A.; Brown-Brandl, T.M.; Nienaber, J.A.; Hahn, G.L. Dynamic Response Indicators of Heat Stress in Shaded and Non-shaded Feedlot Cattle, Part 2: Predictive Relationships. Biosyst. Eng. 2005, 91, 111–118. [Google Scholar] [CrossRef] [Green Version]
- Brown-Brandl, T.M.; Eigenberg, R.A.; Nienaber, J.A.; Hahn, G.L. Dynamic Response Indicators of Heat Stress in Shaded and Non-shaded Feedlot Cattle, Part 1: Analyses of Indicators. Biosyst. Eng. 2005, 90, 451–462. [Google Scholar] [CrossRef] [Green Version]
- Jordan, E.R. Effects of Heat Stress on Reproduction. J. Dairy Sci. 2003, 86, E104–E114. [Google Scholar] [CrossRef]
- West, J.W. Effects of Heat-Stress on Production in Dairy Cattle. J. Dairy Sci. 2003, 86, 2131–2144. [Google Scholar] [CrossRef]
- Rhoads, M.L.; Rhoads, R.P.; VanBaale, M.J.; Collier, R.J.; Sanders, S.R.; Weber, W.J.; Crooker, B.A.; Baumgard, L.H. Effects of heat stress and plane of nutrition on lactating Holstein cows: I. Production, metabolism, and aspects of circulating somatotropin. J. Dairy Sci. 2009, 92, 1986–1997. [Google Scholar] [CrossRef] [Green Version]
- Gaughan, J.B.; Holt, S.M.; Hahn, G.L.; Mader, T.L.; Eigenberg, R.A. Respiration Rate—Is It a Good Measure of Heat Stress in Cattle. Asian-Australas J. Anim. Sci. 2000, 13, 329–332. [Google Scholar]
- Mader, T.L.; Davis, M.S.; Brown-Brandl, T.M. Environmental factors influencing heat stress in feedlot cattle. J. Anim. Sci. 2006, 84, 712–719. [Google Scholar] [CrossRef]
- Robertshaw, D. Heat Loss of Cattle. In Stress Physiology in Livestock; Yousef, M.K., Ed.; CRC Press Inc.: Baco Raton, FL, USA, 1985; Volume I, pp. 55–66. [Google Scholar]
- Young, B.A.; Hall, A.B. Heat load in cattle in the Australian Environment. In Australian Beef; Coombes, R., Ed.; Morescope Publishing: Melbourne, Australia, 1993. [Google Scholar]
- Collier, R.J.; Collier, J.L.; Rhoads, R.P.; Baumgard, L.H. Invited Review: Genes Involved in the Bovine Heat Stress Response. J. Dairy Sci. 2008, 91, 445–454. [Google Scholar] [CrossRef]
- Nienaber, J.A.; Hahn, G.L.; Brown-Brandl, T.M.; Eigenberg, R.A. Heat stress climatic conditions and the physiological responses of cattle. In Proceedings of the Fifth International Dairy Housing, Fort Worth, TX, USA, 29–31 January 2003; pp. 255–262. [Google Scholar]
- Brown-Brandl, T.M.; Nienaber, J.A.; Eigenberg, R.A.; Hahn, G.L.; Freetly, H. Thermoregulatory responses of feeder cattle. J. Therm. Biol. 2003, 28, 149–157. [Google Scholar] [CrossRef]
- Beatty, D.T.; Barnes, A.; Taylor, E.; Maloney, S.K. Do changes in feed intake or ambient temperature cause changes in cattle rumen temperature relative to core temperature? J. Therm. Biol. 2008, 33, 12–19. [Google Scholar] [CrossRef]
- Czerkawski, J.W. A novel estimate of the magnitude of heat produced in the rumen. Br. J. Nutr. 1980, 42, 239–243. [Google Scholar] [CrossRef]
- Ray, D.E.; Roubicek, C.B. Behavior of feedlot cattle during two seasons. J. Anim. Sci. 1971, 33, 72–76. [Google Scholar] [CrossRef]
- Hicks, R.; Owens, F.; Gill, D. Behavioral Patterns of Feedlot Steers; Oklahoma State University Animal Science Research Report, MP-127; Oklahoma State University: Stillwater, MN, USA, 1989; pp. 94–105. [Google Scholar]
- Beede, D.K.; Collier, R.J. Potential Nutritional Strategies for Intensively Managed Cattle during Thermal Stress. J. Anim. Sci. 1986, 62, 543–554. [Google Scholar] [CrossRef]
- NRC. Effect of Environment on Nutrient Requirements of Domestic Animals; National Academy Press: Washington, DC, USA, 1981. [Google Scholar]
- Arias, R.A.; Mader, T.L. Environmental factors affecting daily water intake on cattle finished in feedlots. J. Anim. Sci. 2011, 89, 245–251. [Google Scholar] [CrossRef] [Green Version]
- NRC. Nutrient Requirements of Beef Cattle; National Research Council: Washington, DC, USA, 2000. [Google Scholar]
- McDowell, R.E.; Weldy, J.R. Water Exhcange of cattle under heat stress. In Proceedings of the 3rd International Biometeorological Congress, London, UK, 1967; pp. 414–424. [Google Scholar]
- Black, A.L.; Baker, N.F.; Bartley, J.C.; Chapman, T.E.; Phillips, R.W. Water Turnover in Cattle. Science 1964, 144, 876–878. [Google Scholar] [CrossRef]
- Silanikove, N. Effects of water scarcity and hot environment on appetite and digestion in ruminants: A review. Livest. Prod. Sci. 1992, 30, 175–194. [Google Scholar] [CrossRef]
- Baumgard, L.H.; Rhoads, R.P. RUMINANT NUTRITION SYMPOSIUM: Ruminant Production and Metabolic Responses to Heat Stress. J. Anim. Sci. 2012, 90, 1855–1865. [Google Scholar] [CrossRef]
- Baumgard, L.H.; Rhoads, R.P. The Effects of Hyperthermia on Nutrient Paritioning. Available online: https://www.sid.ir/En/Journal/ViewPaper.aspx?ID=352520 (accessed on 1 October 2012).
- Engelhardt, W.V.; Hales, J.R.S. Partition of capillary blood flow in rumen, reticulum, and omasum of sheep. Am. J. Physiol. 1977, 232, E53–E56. [Google Scholar] [CrossRef]
- NRC. Nutrient Requirements of Beef Cattle; National Research Council: Washington, DC, USA, 2001. [Google Scholar]
- Carroll, J.A.; Burdick Sanchez, N.C.; Bill, E. Kunkle Interdisciplinary Beef Symposium: Overlapping physiological responses and endocrine biomarkers that are indicative of stress responsiveness and immune function in beef cattle. J. Anim. Sci. 2014, 92, 5311–5318. [Google Scholar] [CrossRef]
- Baumgard, L.H.; Rhoads, R.P. Effects of Heat Stress on Postabsorptive Metabolism and Energetics. Annu. Rev. Anim. Biosci. 2013, 1, 311–337. [Google Scholar] [CrossRef] [Green Version]
- Wheelock, J.B.; Rhoads, R.P.; VanBaale, M.J.; Sanders, S.R.; Baumgard, L.H. Effects of heat stress on energetic metabolism in lactating Holstein cows. J. Dairy Sci. 2010, 93, 644–655. [Google Scholar] [CrossRef]
- Lees, A.M.; Sejian, V.; Lees, J.C.; Sullivan, M.L.; Lisle, A.T.; Gaughan, J.B. Evaluating rumen temperature as an estimate of core body temperature in Angus feedlot cattle during summer. Int. J. Biometeorol. 2019. [Google Scholar] [CrossRef]
- Maurya, V.P.; Sejian, V.; Gupta, M.; Dangi, S.S.; Kushwaha, A.; Singh, G.; Sarkar, M. Adaptive Mechanisms of Livestock to Changing Climate. In Climate Change Impact on Livestock: Adaptation and Mitigation; Sejian, V., Gaughan, J., Baumgard, L., Prasad, C., Eds.; Springer: New Delhi, India, 2015; Volume 9, pp. 123–138. [Google Scholar] [CrossRef]
- Ravagnolo, O.; Misztal, I. Effect of Heat Stress on Nonreturn Rate in Holsteins: Fixed-Model Analyses. J. Dairy Sci. 2002, 85, 3101–3106. [Google Scholar] [CrossRef]
- Bitman, J.; Lefcourt, A.; Wood, D.L.; Stroud, B. Circadian and Ultradian Temperature Rhythms of Lactating Dairy Cows. J. Dairy Sci. 1984, 67, 1014–1023. [Google Scholar] [CrossRef]
- Lefcourt, A.M.; Huntington, J.B.; Akers, R.M.; Wood, D.L.; Bitman, J. Circadian and ultradian rhythms of body temperature and peripheral concentrations of insulin and nitrogen in lactating dairy cows. Domest. Anim. Endocrinol. 1999, 16, 41–55. [Google Scholar] [CrossRef]
- Sjaastad, O.V.; Hove, K.; Sand, O. Physiology of Domestic Animals; Scandinavian Veterinary Press: Olso, Norway, 2003. [Google Scholar]
- Findlay, J.D. Physiological Reactions of Cattle to Climatic Stress. Proc. Nutr. Soc. 1958, 17, 186–190. [Google Scholar] [CrossRef]
- Verwoerd, W.; Wellby, M.; Barrell, G. Absence of a causal relationship between environmental and body temperature in dairy cows (Bos taurus) under moderate climatic conditions. J. Therm. Biol. 2006, 31, 533–540. [Google Scholar] [CrossRef]
- Spiers, D.E.; Spain, J.N.; Sampson, J.D.; Rhoads, R.P. Use of physiological parameters to predict milk yield and feed intake in heat-stressed dairy cows. J. Therm. Biol. 2004, 29, 759–764. [Google Scholar] [CrossRef]
- Mehla, K.; Magotra, A.; Choudhary, J.; Singh, A.K.; Mohanty, A.K.; Upadhyay, R.C.; Srinivasan, S.; Gupta, P.; Choudhary, N.; Antony, B.; et al. Genome-wide analysis of the heat stress response in Zebu (Sahiwal) cattle. Gene 2014, 533, 500–507. [Google Scholar] [CrossRef]
- Casady, R.B.; Myers, R.M.; Legates, J.E. The Effect of Exposure to High Ambient Temperature on Spermatogenesis in the Dairy Bull. J. Dairy Sci. 1953, 36, 14–23. [Google Scholar] [CrossRef]
- Johnston, J.E.; Naelapaa, H.; Frye, J.B., Jr. Physiological Responses of Holstein, Brown Swiss and Red Sindhi Crossbred Bulls Exposed to High Temperatures and Humidities. J. Anim. Sci. 1963, 22, 432–436. [Google Scholar] [CrossRef]
- Kastelic, J.; Cook, R.B.; Coulter, G.H. Contribution of the scrotum and testes to scrotal and testicular thermoregulation in bulls and rams. Reproduction 1996, 108, 81–85. [Google Scholar] [CrossRef] [Green Version]
- Kastelic, J.P.; Cook, R.B.; Coulter, G.H.; Saacke, R.G. Insulating the scrotal neck affects semen quality and scrotal/testicular temperatures in the bull. Theriogenology 1996, 45, 935–942. [Google Scholar] [CrossRef]
- Meyerhoeffer, D.C.; Wells, M.E.; Wettemann, R.P.; Coleman, S.W. Reproductive Criteria of Beef Bulls during and after Exposure to Increased Ambient Temperature. J. Anim. Sci. 1985, 60, 352–357. [Google Scholar] [CrossRef]
- Meyerhoeffer, D.C.; Turman, E.J.; Minton, J.E.; Hintz, R.L.; Wettemann, R.P. Serum Luteinizing Hormone and Testosterone in Bulls during Exposure to Elevated Ambient Temperature. J. Anim. Sci. 1981, 53, 1551–1558. [Google Scholar] [CrossRef]
- Skinner, J.D.; Louw, G.N. Heat stress and spermatogenesis in Bos indicus and Bos taurus cattle. J. Appl. Physiol. 1966, 21, 1784–1790. [Google Scholar] [CrossRef]
- Vogler, C.J.; Bame, J.H.; DeJarnette, J.M.; McGilliard, M.L.; Saacke, R.G. Effects of elevated testicular temperature on morphology characteristics of ejaculated spermatozoa in the bovine. Theriogenology 1993, 40, 1207–1219. [Google Scholar] [CrossRef]
- Cruz Júnior, C.A.; Lucci, C.M.; Peripolli, V.; Silva, A.F.; Menezes, A.M.; Morais, S.R.L.; Araújo, M.S.; Ribeiro, L.M.C.S.; Mattos, R.C.; McManus, C. Effects of testicle insulation on seminal traits in rams: Preliminary study. Small Rumin. Res. 2015, 130, 157–165. [Google Scholar] [CrossRef]
- Wallage, A.L.; Gaughan, J.B.; Lisle, A.T.; Beard, L.; Collins, C.W.; Johnston, S.D. Measurement of bovine body and scrotal temperature using implanted temperature sensitive radio transmitters, data loggers and infrared thermography. Int. J. Biometeorol. 2017, 61, 1309–1321. [Google Scholar] [CrossRef]
- Wallage, A.L.; Johnston, S.D.; Lisle, A.T.; Beard, L.; Lees, A.M.; Collins, C.W.; Gaughan, J.B. Thermoregulation of the bovine scrotum 1: Measurements of free-range animals in a paddock and pen. Int. J. Biometeorol. 2017, 61, 1381–1387. [Google Scholar] [CrossRef]
- Wilson, S.J.; Kirby, C.J.; Koenigsfeld, A.T.; Keisler, D.H.; Lucy, M.C. Effects of Controlled Heat Stress on Ovarian Function of Dairy Cattle. 2. Heifers. J. Dairy Sci. 1998, 81, 2132–2138. [Google Scholar] [CrossRef]
- Schüller, L.K.; Michaelis, I.; Heuwieser, W. Impact of heat stress on estrus expression and follicle size in estrus under field conditions in dairy cows. Theriogenology 2017, 102, 48–53. [Google Scholar] [CrossRef]
- Wolfenson, D.; Thatcher, W.W.; Badinga, L.; Savio, J.D.; Meidan, R.; Lew, B.J.; Braw-tal, R.; Berman, A. Effect of Heat Stress on Follicular Development during the Estrous Cycle in Lactating Dairy Cattle. Biol. Reprod. 1995, 52, 1106–1113. [Google Scholar] [CrossRef]
- Jonsson, N.N.; McGowan, M.R.; McGuigan, K.; Davison, T.M.; Hussain, A.M.; Kafi, M.; Matschoss, A. Relationships among calving season, heat load, energy balance and postpartum ovulation of dairy cows in a subtropical environment. Anim. Reprod. Sci. 1997, 47, 315–326. [Google Scholar] [CrossRef]
- García-Ispierto, I.; López-Gatius, F.; Santolaria, P.; Yániz, J.L.; Nogareda, C.; López-Béjar, M.; De Rensis, F. Relationship between heat stress during the peri-implantation period and early fetal loss in dairy cattle. Theriogenology 2006, 65, 799–807. [Google Scholar] [CrossRef]
- Torres-Júnior, J.R.D.S.; Pires, M.D.F.A.; de Sá, W.F.; Ferreira, A.D.M.; Viana, J.H.M.; Camargo, L.S.A.; Ramos, A.A.; Folhadella, I.M.; Polisseni, J.; de Freitas, C.; et al. Effect of maternal heat-stress on follicular growth and oocyte competence in Bos indicus cattle. Theriogenology 2008, 69, 155–166. [Google Scholar] [CrossRef]
- Al-Katanani, Y.M.; Paula-Lopes, F.F.; Hansen, P.J. Effect of Season and Exposure to Heat Stress on Oocyte Competence in Holstein Cows. J. Dairy Sci. 2002, 85, 390–396. [Google Scholar] [CrossRef]
- Ealy, A.D.; Aréchiga, C.F.; Howell, J.L.; Hansen, P.J.; Monterroso, V.H. Developmental changes in sensitivity of bovine embryos to heat shock and use of antioxidants as thermoprotectants2. J. Anim. Sci. 1995, 73, 1401–1407. [Google Scholar] [CrossRef]
- Gendelman, M.; Aroyo, A.; Yavin, S.; Roth, Z. Seasonal effects on gene expression, cleavage timing, and developmental competence of bovine preimplantation embryos. Reproduction 2010, 140, 73–82. [Google Scholar] [CrossRef] [Green Version]
- Biggers, B.G.; Buchanan, D.S.; Geisert, R.D.; Wetteman, R.P. Effect of Heat Stress on Early Embryonic Development in the Beef Cow. J. Anim. Sci. 1987, 64, 1512–1518. [Google Scholar] [CrossRef] [Green Version]
- Ryan, D.P.; Blakewood, E.G.; Munyakazi, L.; Godke, R.A.; Lynn, J.W. Effect of heat-stress on bovine embryo development in vitro. J. Anim. Sci. 1992, 70, 3490–3497. [Google Scholar] [CrossRef]
- Wolfenson, D.; Roth, Z.; Meidan, R. Impaired reproduction in heat-stressed cattle: Basic and applied aspects. Anim. Reprod. Sci. 2000, 60–61, 535–547. [Google Scholar] [CrossRef]
- Roman-Ponce, H.; Thatcher, W.W.; Caton, D.; Barron, D.H.; Wilcox, C.J. Thermal Stress Effects on Uterine Blood Flow in Dairy Cows. J. Anim. Sci. 1978, 46, 175–180. [Google Scholar] [CrossRef]
- Pennington, J.A.; Albright, J.L.; Diekman, M.A.; Callahan, C.J. Sexual Activity of Holstein Cows: Seasonal Effects. J. Dairy Sci. 1985, 68, 3023–3030. [Google Scholar] [CrossRef]
- Schüller, L.K.; Burfeind, O.; Heuwieser, W. Impact of heat stress on conception rate of dairy cows in the moderate climate considering different temperature–humidity index thresholds, periods relative to breeding, and heat load indices. Theriogenology 2014, 81, 1050–1057. [Google Scholar] [CrossRef]
- Morton, J.M.; Tranter, W.P.; Mayer, D.G.; Jonsson, N.N. Effects of Environmental Heat on Conception Rates in Lactating Dairy Cows: Critical Periods of Exposure. J. Dairy Sci. 2007, 90, 2271–2278. [Google Scholar] [CrossRef]
- Hansen, P.J.; Areéchiga, C.F. Strategies for managing reproduction in the heat-stressed dairy cow. J. Anim. Sci. 1999, 77, 36–50. [Google Scholar] [CrossRef]
- Collier, R.J.; Beede, D.K.; Thatcher, W.W.; Israel, L.A.; Wilcox, C.J. Influences of Environment and Its Modification on Dairy Animal Health and Production. J. Dairy Sci. 1982, 65, 2213–2227. [Google Scholar] [CrossRef]
- Kadzere, C.T.; Murphy, M.R.; Silanikove, N.; Maltz, E. Heat stress in lactating dairy cows: A review. Livest. Prod. Sci. 2002, 77, 59–91. [Google Scholar] [CrossRef]
- Morse, D.; DeLorenzo, M.A.; Wilcox, C.J.; Collier, R.J.; Natzke, R.P.; Bray, D.R. Climatic Effects on Occurrence of Clinical Mastitis. J. Dairy Sci. 1988, 71, 848–853. [Google Scholar] [CrossRef]
- Howell, D.; Wilson, C.D.; Vessey, M.P. A survey of the incidence of mastitis in dairy cows in the Reading area. Vet. Rec. 1964, 76, 1107. [Google Scholar]
- Gardner, B.A.; Dolezal, H.G.; Bryant, L.K.; Owens, F.N.; Smith, R.A. Health of finishing steers: Effects on performance, carcass traits, and meat tenderness. J. Anim. Sci. 1999, 77, 3168–3175. [Google Scholar] [CrossRef]
- Silanikove, N. Effects of heat stress on the welfare of extensively managed domestic ruminants. Livest. Prod. Sci. 2000, 67, 1–18. [Google Scholar] [CrossRef]
- DeShazer, J.A.; Hahn, G.L.; Xinm, H. Basic Principals of the Thermal Environment and Livestock Energetics. In Livestock Energetics and Thermal Environmental Management; DeShazer, J.A., Ed.; American Society of Agricultural and Biological Engineers: St. Joseph, MI, USA, 2009. [Google Scholar]
- Mitlöhner, F.M.; Galyean, M.L.; McGlone, J.J. Shade effects on performance, carcass traits, physiology, and behavior of heat-stressed feedlot heifers. J. Anim. Sci. 2002, 80, 2043–2050. [Google Scholar] [CrossRef]
- Gaughan, J.B.; Mader, T.L. Body temperature and respiratory dynamics in un-shaded beef cattle. Int. J. Biometeorol. 2014, 58, 1443–1450. [Google Scholar] [CrossRef]
- Gaughan, J.B.; Bonner, S.; Loxton, I.; Mader, T.L.; Lisle, A.; Lawrence, R. Effect of shade on body temperature and performance of feedlot steers. J. Anim. Sci. 2010, 88, 4056–4067. [Google Scholar] [CrossRef]
- Lees, A.M.; Lees, J.C.; Lisle, A.T.; Sullivan, M.L.; Gaughan, J.B. Effect of heat stress on rumen temperature of three breeds of cattle. Int. J. Biometeorol. 2018, 62, 207–215. [Google Scholar] [CrossRef]
- Sullivan, M.L.; Cawdell-Smith, A.J.; Mader, T.L.; Gaughan, J.B. Effect of shade area on performance and welfare of short-fed feedlot cattle. J. Anim. Sci. 2011, 89, 2911–2925. [Google Scholar] [CrossRef]
- Lees, A.M.; Lees, J.C.; Sejian, V.; Wallage, A.L.; Gaughan, J.B. Short communication: Using infrared thermography as an in situ measure of core body temperature in lot-fed Angus steers. Int. J. Biometeorol. 2018, 62, 3–8. [Google Scholar] [CrossRef]
- Clarke, M.; Kelly, A. Some effects of shade on Hereford steers in a feedlot. Proc. Aust. Soc. Anim. Prod. 1996, 21, 235–238. [Google Scholar]
- Gaughan, J.B.; Bonner, S.L.; Loxton, I.; Mader, T.L. Effects of chronic heat stress on plasma concentration of secreted heat shock protein 70 in growing feedlot cattle. J. Anim. Sci. 2013, 91, 120–129. [Google Scholar] [CrossRef]
- Avendaño-Reyes, L.; Álvarez-Valenzuela, F.D.; Correa-Calderón, A.; Algándar-Sandoval, A.; Rodríguez-González, E.; Pérez-Velázquez, R.; Macías-Cruz, U.; Díaz-Molina, R.; Robinson, P.H.; Fadel, J.G. Comparison of three cooling management systems to reduce heat stress in lactating Holstein cows during hot and dry ambient conditions. Livest. Sci. 2010, 132, 48–52. [Google Scholar] [CrossRef]
- McDowell, R.E.; Hooven, N.W.; Camoens, J.K. Effect of Climate on Performance of Holsteins in First Lactation. J. Dairy Sci. 1976, 59, 965–971. [Google Scholar] [CrossRef]
- TapkI, I.; Sahin, A. Comparison of the thermoregulatory behaviours of low and high producing dairy cows in a hot environment. Appl. Anim. Behav. Sci. 2006, 99, 1–11. [Google Scholar] [CrossRef]
- Bouraoui, R.; Lahmar, M.; Abdessalem, M.; Djemali, M.n.; Belyea, R. The relationship of temperature-humidity index with milk production of dairy cows in a Mediterranean climate. Anim. Res. 2002, 51, 479–491. [Google Scholar] [CrossRef] [Green Version]
- Staples, C.R.; Thatcher, W.W. Stress in Dairy Animals | Heat Stress: Effects on Milk Production and Composition. In Encyclopedia of Dairy Sciences, 2nd ed.; John, W., Ed.; Academic Press: San Diego, CA, USA, 2011; pp. 561–566. [Google Scholar] [CrossRef]
- Ravagnolo, O.; Misztal, I. Genetic Component of Heat Stress in Dairy Cattle, Parameter Estimation. J. Dairy Sci. 2000, 83, 2126–2130. [Google Scholar] [CrossRef]
- Purwanto, B.; Abo, Y.; Sakamoto, R.; Furumoto, F.; Yamamoto, S. Diurnal patterns of heat production and heart rate under thermoneutral conditions in Holstein Friesian cows differing in milk production. J. Agric. Sci. 1990, 114, 139–142. [Google Scholar] [CrossRef]
- Lambertz, C.; Sanker, C.; Gauly, M. Climatic effects on milk production traits and somatic cell score in lactating Holstein-Friesian cows in different housing systems. J. Dairy Sci. 2014, 97, 319–329. [Google Scholar] [CrossRef]
- Sharma, A.; Rodriguez, L.; Mekonnen, G.; Wilcox, C.; Bachman, K.; Collier, R. Climatological and Genetic Effects on Milk Composition and Yield. J. Dairy Sci. 1983, 66, 119–126. [Google Scholar] [CrossRef]
- Heck, J.M.L.; Schennink, A.; van Valenberg, H.J.F.; Bovenhuis, H.; Visker, M.H.P.W.; van Arendonk, J.A.M.; van Hooijdonk, A.C.M. Effects of milk protein variants on the protein composition of bovine milk. J. Dairy Sci. 2009, 92, 1192–1202. [Google Scholar] [CrossRef]
- Pollott, G.E. Deconstructing Milk Yield and Composition During Lactation Using Biologically Based Lactation Models. J. Dairy Sci. 2004, 87, 2375–2387. [Google Scholar] [CrossRef]
- Rodriquez, L.A.; Mekonnen, G.; Wilcox, C.J.; Martin, F.G.; Krienke, W.A. Effects of Relative Humidity, Maximum and Minimum Temperature, Pregnancy, and Stage of Lactation on Milk Composition and Yield. J. Dairy Sci. 1985, 68, 973–978. [Google Scholar] [CrossRef]
- Bernabucci, U.; Lacetera, N.; Baumgard, L.H.; Rhoads, R.P.; Ronchi, B.; Nardone, A. Metabolic and hormonal acclimation to heat stress in domesticated ruminants. Animal 2010, 4, 1167–1183. [Google Scholar] [CrossRef] [Green Version]
- Quist, M.A.; LeBlanc, S.J.; Hand, K.J.; Lazenby, D.; Miglior, F.; Kelton, D.F. Milking-to-Milking Variability for Milk Yield, Fat and Protein Percentage, and Somatic Cell Count. J. Dairy Sci. 2008, 91, 3412–3423. [Google Scholar] [CrossRef]
- Bernabucci, U.; Basiricò, L.; Morera, P.; Dipasquale, D.; Vitali, A.; Piccioli Cappelli, F.; Calamari, L. Effect of summer season on milk protein fractions in Holstein cows. J. Dairy Sci. 2015, 98, 1815–1827. [Google Scholar] [CrossRef] [Green Version]
- Sharma, A.K.; Rodriguez, L.A.; Wilcox, C.J.; Collier, R.J.; Bachman, K.C.; Martin, F.G. Interactions of Climatic Factors Affecting Milk Yield and Composition. J. Dairy Sci. 1988, 71, 819–825. [Google Scholar] [CrossRef]
- Hill, D.L.; Wall, E. Dairy cattle in a temperate climate: The effects of weather on milk yield and composition depend on management. Animal 2014, 9, 138–149. [Google Scholar] [CrossRef]
- Garner, J.B.; Douglas, M.; Williams, S.R.O.; Wales, W.J.; Marett, L.C.; DiGiacomo, K.; Leury, B.J.; Hayes, B.J. Responses of dairy cows to short-term heat stress in controlled-climate chambers. Anim. Prod. Sci. 2017, 57, 1233–1241. [Google Scholar] [CrossRef]
- Bernabucci, U.; Lacetera, N.; Ronchi, B.; Nardone, A. Effects of the hot season on milk protein fractions in Holstein cows. Anim. Res. 2002, 51, 25–33. [Google Scholar] [CrossRef] [Green Version]
- Ferris, T.A.; Vasavada, P.C. Altering Milk Composition—An Introduction. J. Dairy Sci. 1989, 72, 2788–2789. [Google Scholar] [CrossRef]
- Laben, R.C. Factors Responsible for Variation in Milk Composition. J. Dairy Sci. 1963, 46, 1293–1301. [Google Scholar] [CrossRef]
- Loudon, K.M.W.; Lean, I.J.; Pethick, D.W.; Gardner, G.E.; Grubb, L.J.; Evans, A.C.; McGilchrist, P. On farm factors increasing dark cutting in pasture finished beef cattle. Meat Sci. 2018, 144, 110–117. [Google Scholar] [CrossRef]
- McGilchrist, P.; Alston, C.L.; Gardner, G.E.; Thomson, K.L.; Pethick, D.W. Beef carcasses with larger eye muscle areas, lower ossification scores and improved nutrition have a lower incidence of dark cutting. Meat Sci. 2012, 92, 474–480. [Google Scholar] [CrossRef] [Green Version]
- Voisinet, B.D.; Grandin, T.; O’Connor, S.F.; Tatum, J.D.; Deesing, M.J. Bos indicus-cross feedlot cattle with excitable temperaments have tougher meat and a higher incidence of borderline dark cutters. Meat Sci. 1997, 46, 367–377. [Google Scholar] [CrossRef]
- Voisinet, B.D.; Grandin, T.; Tatum, J.D.; O’Connor, S.F.; Struthers, J.J. Feedlot cattle with calm temperaments have higher average daily gains than cattle with excitable temperaments. J. Anim. Sci. 1997, 75, 892–896. [Google Scholar] [CrossRef] [PubMed]
- McGilchrist, P.; Perovic, J.L.; Gardner, G.E.; Pethick, D.W.; Jose, C.G. The incidence of dark cutting in southern Australian beef production systems fluctuates between months. Anim. Prod. Sci. 2014, 54, 1765–1769. [Google Scholar] [CrossRef] [Green Version]
- Scanga, J.A.; Belk, K.E.; Tatum, J.D.; Grandin, T.; Smith, G.C. Factors contributing to the incidence of dark cutting beef. J. Anim. Sci. 1998, 76, 2040–2047. [Google Scholar] [CrossRef] [PubMed]
- Hahn, G.L. Management and Housing of Farm Animals in Hot Environments. In Stress Physiology in Livestock; Yousef, M.K., Ed.; CRC Press Inc.: Boca Raton, FL, USA, 1985; Volume II, pp. 151–174. [Google Scholar]
- Sanchez, W.K.; McGuire, M.A.; Beede, D.K. Macromineral Nutrition by Heat Stress Interactions in Dairy Cattle: Review and Original Research. J. Dairy Sci. 1994, 77, 2051–2079. [Google Scholar] [CrossRef]
- Gaughan, J.B.; Davis, M.S.; Mader, T.L. Wetting and the physiological responses of grain-fed cattle in a heated environment. Aust. J. Agric. Res. 2004, 55, 253–260. [Google Scholar] [CrossRef]
- Gaughan, J.B.; Mader, T.L.; Holt, S.M. Cooling and feeding strategies to reduce heat load of grain-fed beef cattle in intensive housing. Livest. Sci. 2008, 113, 226–233. [Google Scholar] [CrossRef]
- Schütz, K.E.; Cox, N.R.; Matthews, L.R. How important is shade to dairy cattle? Choice between shade or lying following different levels of lying deprivation. Appl. Anim. Behav. Sci. 2008, 114, 307–318. [Google Scholar] [CrossRef]
- Schütz, K.E.; Cox, N.R.; Tucker, C.B. A field study of the behavioral and physiological effects of varying amounts of shade for lactating cows at pasture. J. Dairy Sci. 2014, 97, 3599–3605. [Google Scholar] [CrossRef]
- Schütz, K.E.; Rogers, A.R.; Cox, N.R.; Tucker, C.B. Dairy cows prefer shade that offers greater protection against solar radiation in summer: Shade use, behaviour, and body temperature. Appl. Anim. Behav. Sci. 2009, 116, 28–34. [Google Scholar] [CrossRef]
- Schütz, K.E.; Rogers, A.R.; Poulouin, Y.A.; Cox, N.R.; Tucker, C.B. The amount of shade influences the behavior and physiology of dairy cattle. J. Dairy Sci. 2010, 93, 125–133. [Google Scholar] [CrossRef]
- Tucker, C.B.; Rogers, A.R.; Schütz, K.E. Effect of solar radiation on dairy cattle behaviour, use of shade and body temperature in a pasture-based system. Appl. Anim. Behav. Sci. 2008, 109, 141–154. [Google Scholar] [CrossRef]
- Gaughan, J.B.; Goodwin, P.J.; Schoorl, T.A.; Young, B.A.; Imbeah, M.; Mader, T.L.; Hall, A. Shade preferences of lactating Holstein Friesian cows. Aust. J. Exp. Agric. 1998, 38, 17–21. [Google Scholar] [CrossRef]
- Buffington, D.; Collier, R.; Canton, G. Shade management systems to reduce heat stress for dairy cows in hot, humid climates. Trans. Am. Soc. Agric. Eng. 1983, 26, 1798–1802. [Google Scholar] [CrossRef]
- Kendall, P.E.; Nielsen, P.P.; Webster, J.R.; Verkerk, G.A.; Littlejohn, R.P.; Matthews, L.R. The effects of providing shade to lactating dairy cows in a temperate climate. Livest. Sci. 2006, 103, 148–157. [Google Scholar] [CrossRef]
- Gaughan, J.B.; Tait, L.A.; Eigenberg, R.; Bryden, W.L. Effect of shade on respiration rate and rectal temperature of Angus heifers. Anim. Prod. Aust. 2004, 25, 69–72. [Google Scholar] [CrossRef]
- Bond, T.E.; Kelly, C.F.; Morrison, S.R.; Periera, N. Solar, Atmospheric, and Terrestrial Radiation Received by Shaded and Unshaded Animals. Trans. Am. Soc. Agric. Eng. 1967, 10, 622–627. [Google Scholar] [CrossRef]
- Roman-Ponce, H.; Thatcher, W.W.; Buffington, D.E.; Wilcox, C.J.; Van Horn, H.H. Physiological and Production Responses of Dairy Cattle to a Shade Structure in a Subtropical Environment. J. Dairy Sci. 1977, 60, 424–430. [Google Scholar] [CrossRef]
- Gaughan, J.B.; Mader, T.L. Effects of sodium chloride and fat supplementation on finishing steers exposed to hot and cold conditions. J. Anim. Sci. 2009, 87, 612–621. [Google Scholar] [CrossRef] [Green Version]
- Dunshea, F.R.; Oluboyede, K.; DiGiacomo, K.; Leury, B.J.; Cottrell, J.J. Betaine Improves Milk Yield in Grazing Dairy Cows Supplemented with Concentrates at High Temperatures. Animals 2019, 9, 57. [Google Scholar] [CrossRef]
- DiGiacomo, K.; Simpson, S.; Leury, B.J.; Dunshea, F.R. Dietary Betaine Impacts the Physiological Responses to Moderate Heat Conditions in a Dose Dependent Manner in Sheep. Animals 2016, 6, 51. [Google Scholar] [CrossRef]
- Cronje, P. Heat stress in livestock–the role of the gut in its aetiology and a potential role for betaine in its alleviation. Recent Adv. Anim. Nutr. Aust. 2005, 15, 107–122. [Google Scholar]
- Moallem, U.; Lehrer, H.; Livshitz, L.; Zachut, M.; Yakoby, S. The effects of live yeast supplementation to dairy cows during the hot season on production, feed efficiency, and digestibility. J. Dairy Sci. 2009, 92, 343–351. [Google Scholar] [CrossRef] [PubMed]
- DeVries, T.J.; Chevaux, E. Modification of the feeding behavior of dairy cows through live yeast supplementation. J. Dairy Sci. 2014, 97, 6499–6510. [Google Scholar] [CrossRef] [PubMed]
- Crossland, W.L.; Cagle, C.M.; Sawyer, J.E.; Callaway, T.R.; Tedeschi, L.O. Evaluation of active dried yeast in the diets of feedlot steers. II. Effects on rumen pH and liver health of feedlot steers. J. Anim. Sci. 2019, 97, 1347–1363. [Google Scholar] [CrossRef] [PubMed]
- Crossland, W.L.; Jobe, J.T.; Ribeiro, F.R.B.; Sawyer, J.E.; Callaway, T.R.; Tedeschi, L.O. Evaluation of active dried yeast in the diets of feedlot steers: I. Effects on feeding performance traits, the composition of growth, and carcass characteristics. J. Anim. Sci. 2019, 97, 1335–1346. [Google Scholar] [CrossRef] [PubMed]
- Calamari, L.; Petrera, F.; Abeni, F.; Bertin, G. Metabolic and hematological profiles in heat stressed lactating dairy cows fed diets supplemented with different selenium sources and doses. Livest. Sci. 2011, 142, 128–137. [Google Scholar] [CrossRef]
- Mader, T.L.; Gaughan, J.B.; Young, B.A. Feedlot Diet Roughage Level for Hereford Cattle Exposed to Excessive Heat Load. Prof. Anim. Sci. 1999, 15, 53–62. [Google Scholar] [CrossRef]
- Brosh, A.; Aharoni, Y.; Degen, A.A.; Wright, D.; Young, B.A. Effects of solar radiation, dietary energy, and time of feeding on thermoregulatory responses and energy balance in cattle in a hot environment. J. Anim. Sci. 1998, 76, 2671–2677. [Google Scholar] [CrossRef]
- Carvalho, F.A.; Lammoglia, M.A.; Simoes, M.J.; Randel, R.D. Breed affects thermoregulation and epithelial morphology in imported and native cattle subjected to heat stress. J. Anim. Sci. 1995, 73, 3570–3573. [Google Scholar] [CrossRef]
- Rhoads, R.P.; Baumgard, L.H.; Suagee, J.K. 2011 and 2012 Early Careers Achievement Awards: Metabolic priorities during heat stress with an emphasis on skeletal muscle. J. Anim. Sci. 2013, 91, 2492–2503. [Google Scholar] [CrossRef]
- Nguyen, T.T.T.; Bowman, P.J.; Haile-Mariam, M.; Nieuwhof, G.J.; Hayes, B.J.; Pryce, J.E. Short communication: Implementation of a breeding value for heat tolerance in Australian dairy cattle. J. Dairy Sci. 2017, 100, 7362–7367. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nguyen, T.T.T.; Bowman, P.J.; Haile-Mariam, M.; Pryce, J.E.; Hayes, B.J. Genomic selection for tolerance to heat stress in Australian dairy cattle. J. Dairy Sci. 2016, 99, 2849–2862. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nguyen, T.T.T.; Hayes, B.J.; Pryce, J.E. A practical future-scenarios selection tool to breed for heat tolerance in Australian dairy cattle. Anim. Prod. Sci. 2017, 57, 1488–1493. [Google Scholar] [CrossRef]
- Garner, J.B.; Douglas, M.L.; Williams, S.R.O.; Wales, W.J.; Marett, L.C.; Nguyen, T.T.T.; Reich, C.M.; Hayes, B.J. Genomic Selection Improves Heat Tolerance in Dairy Cattle. Sci. Rep. 2016, 6, 34114. [Google Scholar] [CrossRef] [PubMed]
- Sánchez, J.P.; Misztal, I.; Aguilar, I.; Zumbach, B.; Rekaya, R. Genetic determination of the onset of heat stress on daily milk production in the US Holstein cattle. J. Dairy Sci. 2009, 92, 4035–4045. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lees, J.C. A Heat Load Index for Dairy Cattle. Ph.D. Thesis, The University of Queensland, School of Agriculture and Food Sciences, Queensland, Australia, 2018. [Google Scholar]
- Singh, K.; Erdman, R.A.; Swanson, K.M.; Molenaar, A.J.; Maqbool, N.J.; Wheeler, T.T.; Arias, J.A.; Quinn-Walsh, E.C.; Stelwagen, K. Epigenetic Regulation of Milk Production in Dairy Cows. J. Mammary Gland Biol. Neoplasia 2010, 15, 101–112. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, B.M.S.; Younas, U.; Asar, T.O.; Dikmen, S.; Hansen, P.J.; Dahl, G.E. Cows exposed to heat stress during fetal life exhibit improved thermal tolerance. J. Anim. Sci. 2017, 95, 3497–3503. [Google Scholar] [CrossRef] [PubMed]
- Tao, S.; Dahl, G.E.; Laporta, J.; Bernard, J.K.; Orellana Rivas, R.M.; Marins, T.N. Effects of heat stress during late gestation on the dam and its calf. J. Anim. Sci. 2019, 96, 351–352. [Google Scholar] [CrossRef]
- Skibiel, A.L.; Dado-Senn, B.; Fabris, T.F.; Dahl, G.E.; Laporta, J. In utero exposure to thermal stress has long-term effects on mammary gland microstructure and function in dairy cattle. PLoS ONE 2018, 13, e0206046. [Google Scholar] [CrossRef]
- Skibiel, A.L.; Peñagaricano, F.; Amorín, R.; Ahmed, B.M.; Dahl, G.E.; Laporta, J. In Utero Heat Stress Alters the Offspring Epigenome. Sci. Rep. 2018, 8, 14609. [Google Scholar] [CrossRef] [Green Version]
- Dahl, G.E.; Tao, S.; Laporta, J. TRIENNIAL LACTATION SYMPOSIUM/BOLFA: Late gestation heat stress of dairy cattle programs dam and daughter milk production. J. Anim. Sci. 2017, 95, 5701–5710. [Google Scholar] [CrossRef] [PubMed]
- Horowitz, M. Heat acclimation: Phenotypic plasticity and cues to the underlying molecular mechanisms. J. Therm. Biol. 2001, 26, 357–363. [Google Scholar] [CrossRef]
- Roy, K.S.; Collier, R.J. Regulation of Acclimation to Environmental Stress. In Environmental Physiology of Livestock; Collier, R.J., Collier, J.L., Eds.; Wiley Blackwell: West Sussex, UK, 2012. [Google Scholar]
- Hansen, P.J. Physiological and cellular adaptations of zebu cattle to thermal stress. Anim. Reprod. Sci. 2004, 82–83, 349–360. [Google Scholar] [CrossRef] [PubMed]

© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lees, A.M.; Sejian, V.; Wallage, A.L.; Steel, C.C.; Mader, T.L.; Lees, J.C.; Gaughan, J.B. The Impact of Heat Load on Cattle. Animals 2019, 9, 322. https://doi.org/10.3390/ani9060322
Lees AM, Sejian V, Wallage AL, Steel CC, Mader TL, Lees JC, Gaughan JB. The Impact of Heat Load on Cattle. Animals. 2019; 9(6):322. https://doi.org/10.3390/ani9060322
Chicago/Turabian StyleLees, Angela M., Veerasamy Sejian, Andrea L. Wallage, Cameron C. Steel, Terry L. Mader, Jarrod C. Lees, and John B. Gaughan. 2019. "The Impact of Heat Load on Cattle" Animals 9, no. 6: 322. https://doi.org/10.3390/ani9060322
APA StyleLees, A. M., Sejian, V., Wallage, A. L., Steel, C. C., Mader, T. L., Lees, J. C., & Gaughan, J. B. (2019). The Impact of Heat Load on Cattle. Animals, 9(6), 322. https://doi.org/10.3390/ani9060322