Sexual Dimorphism and Foraging Trips of the Laysan Albatross (Phoebastria immutabilis) on Guadalupe Island

 , , ,
, , , 
Abstract
:Simple Summary
Abstract
1. Introduction
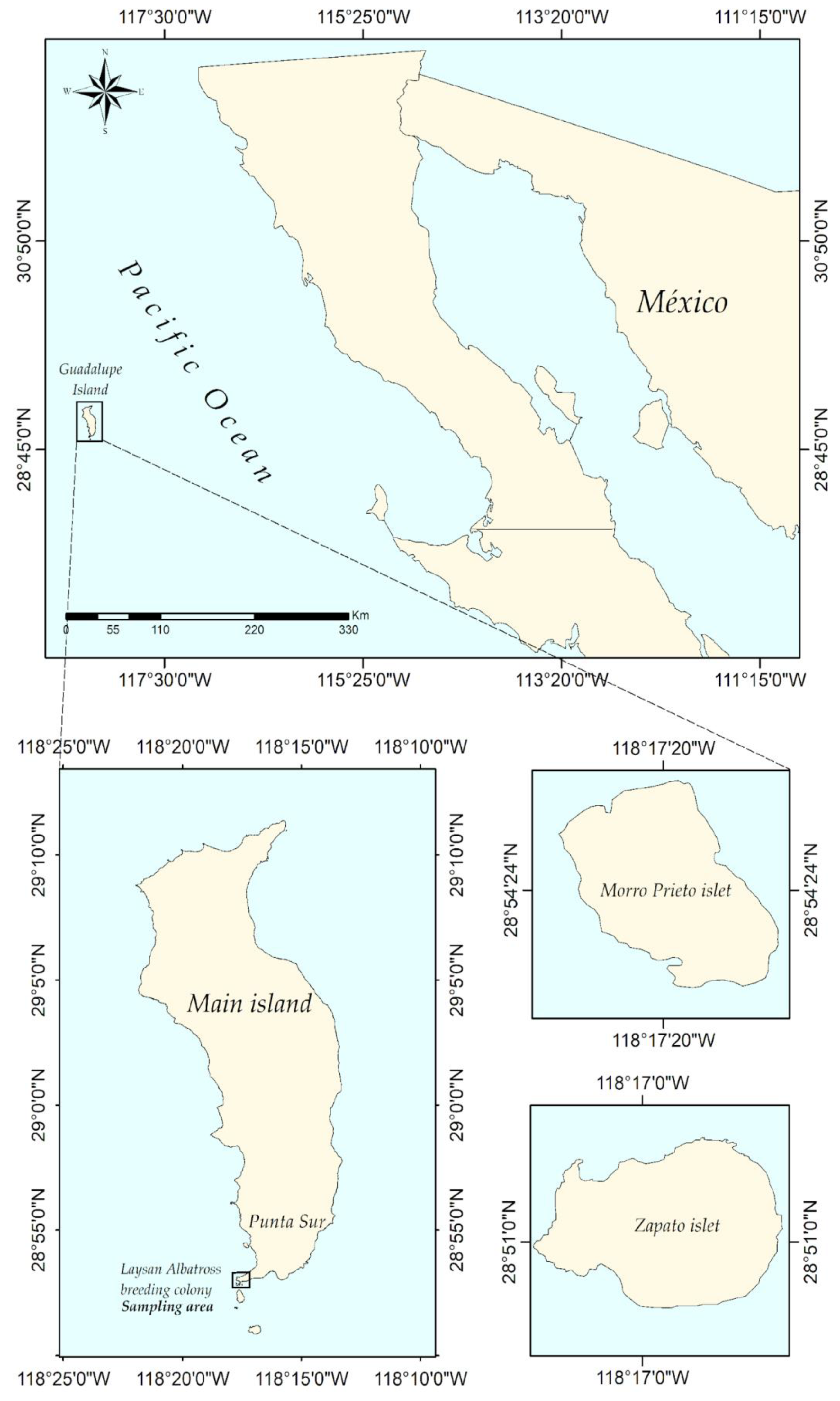
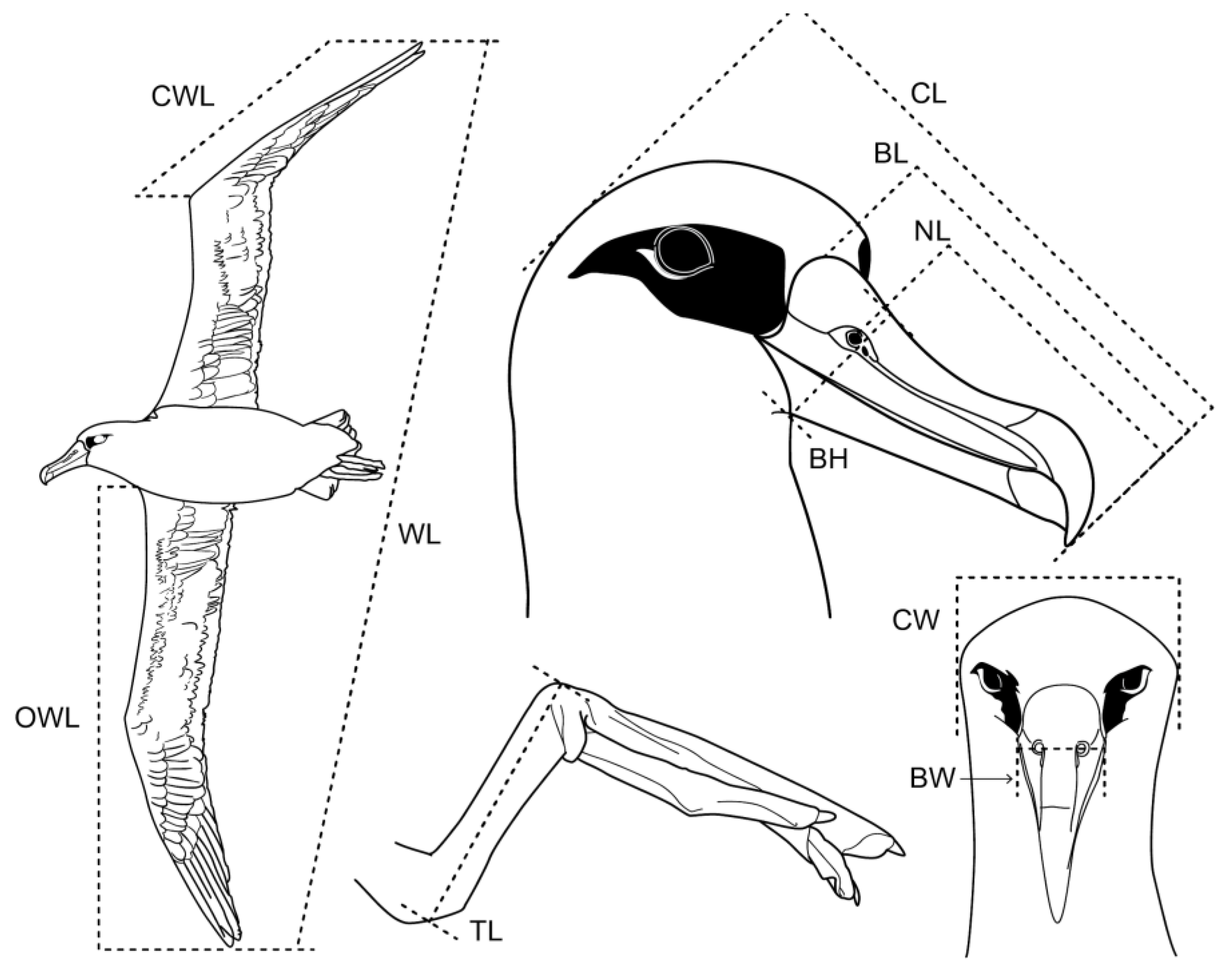
2. Materials and Methods
2.1. Data Collection
2.2. Tracking Data Processing
2.3. Statistical Analysis
3. Results
3.1. Genetic Sex Identification
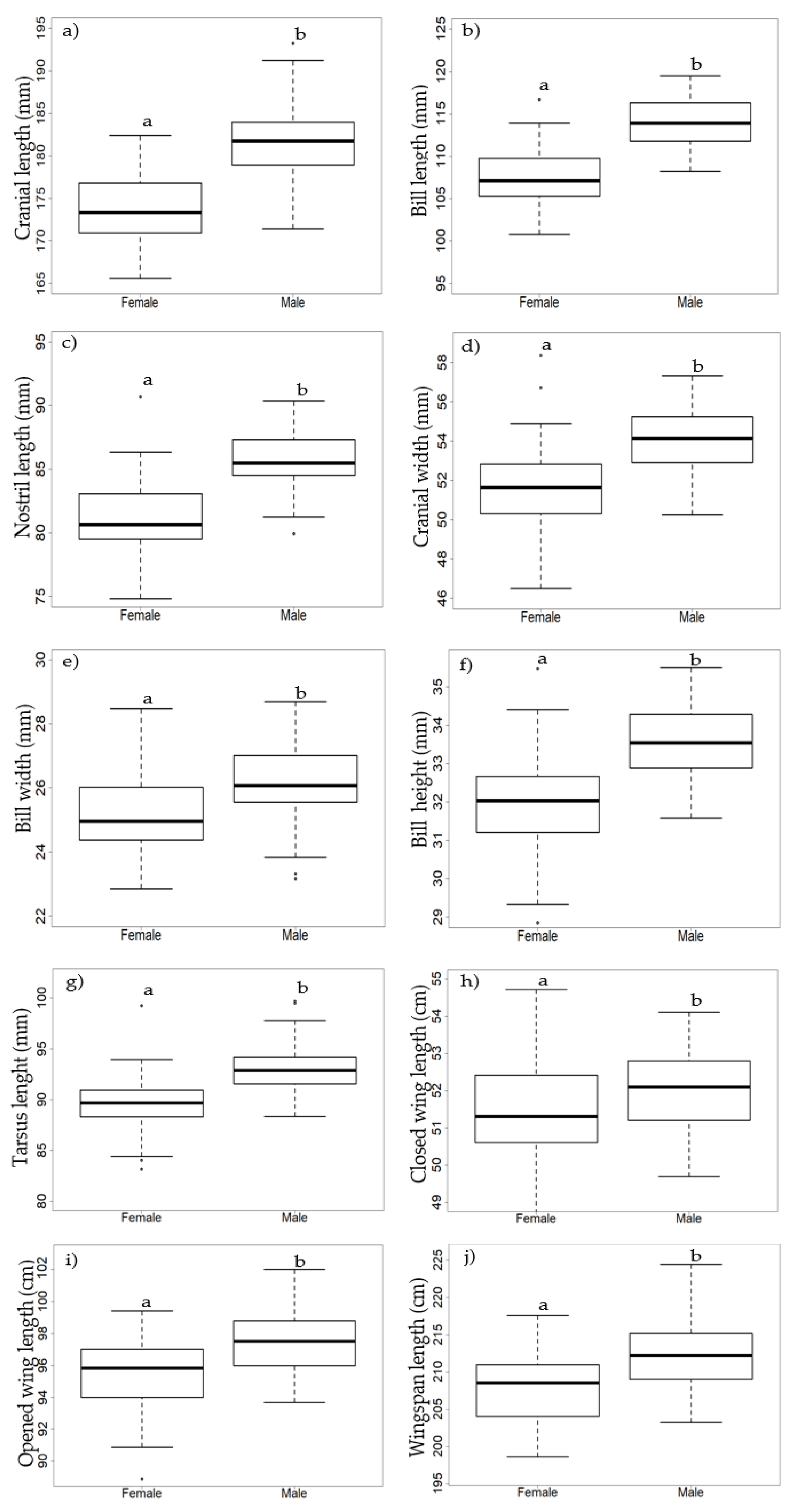
3.2. Morphological Dimorphism
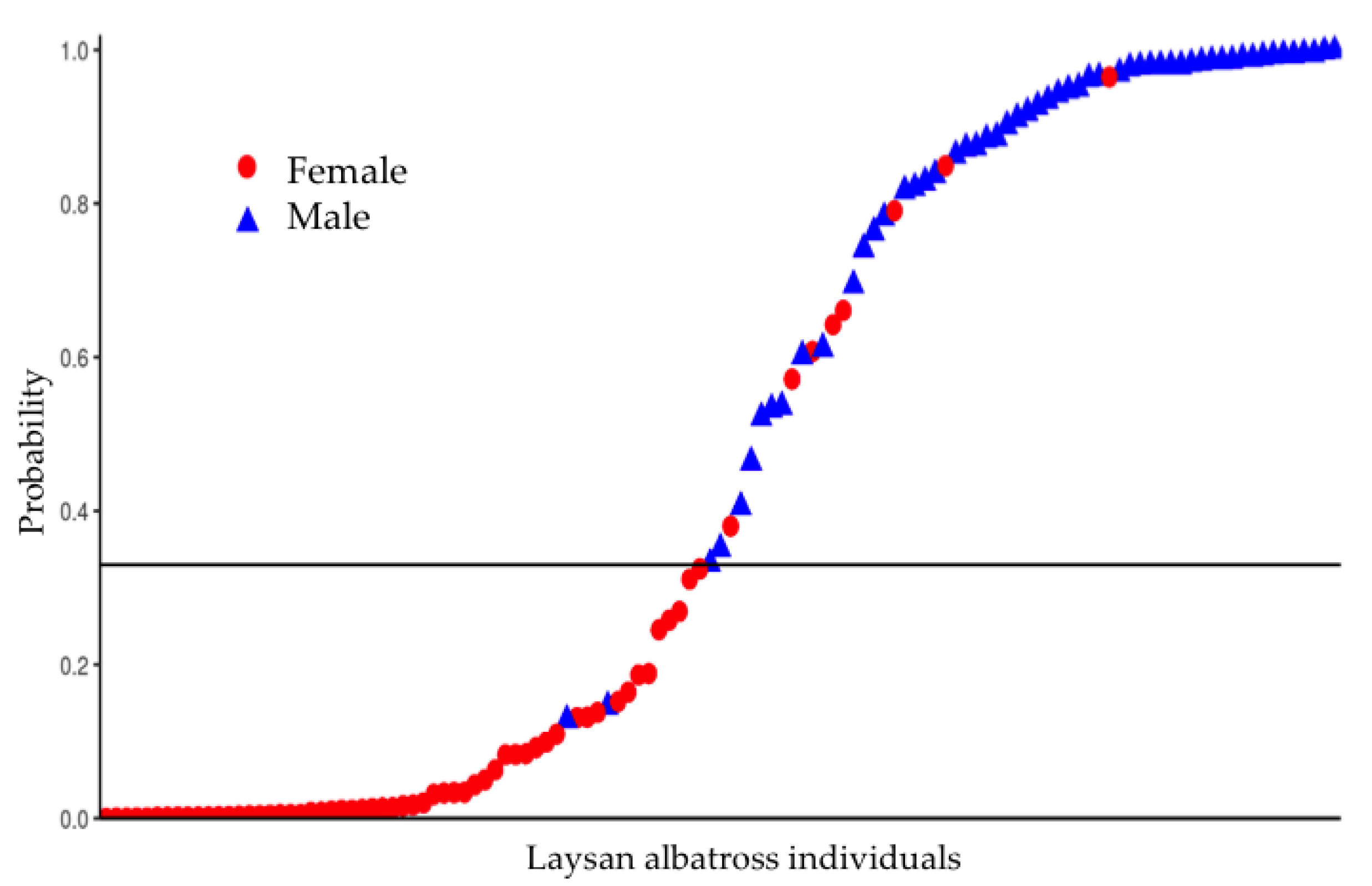
3.3. Sex Prediction Model
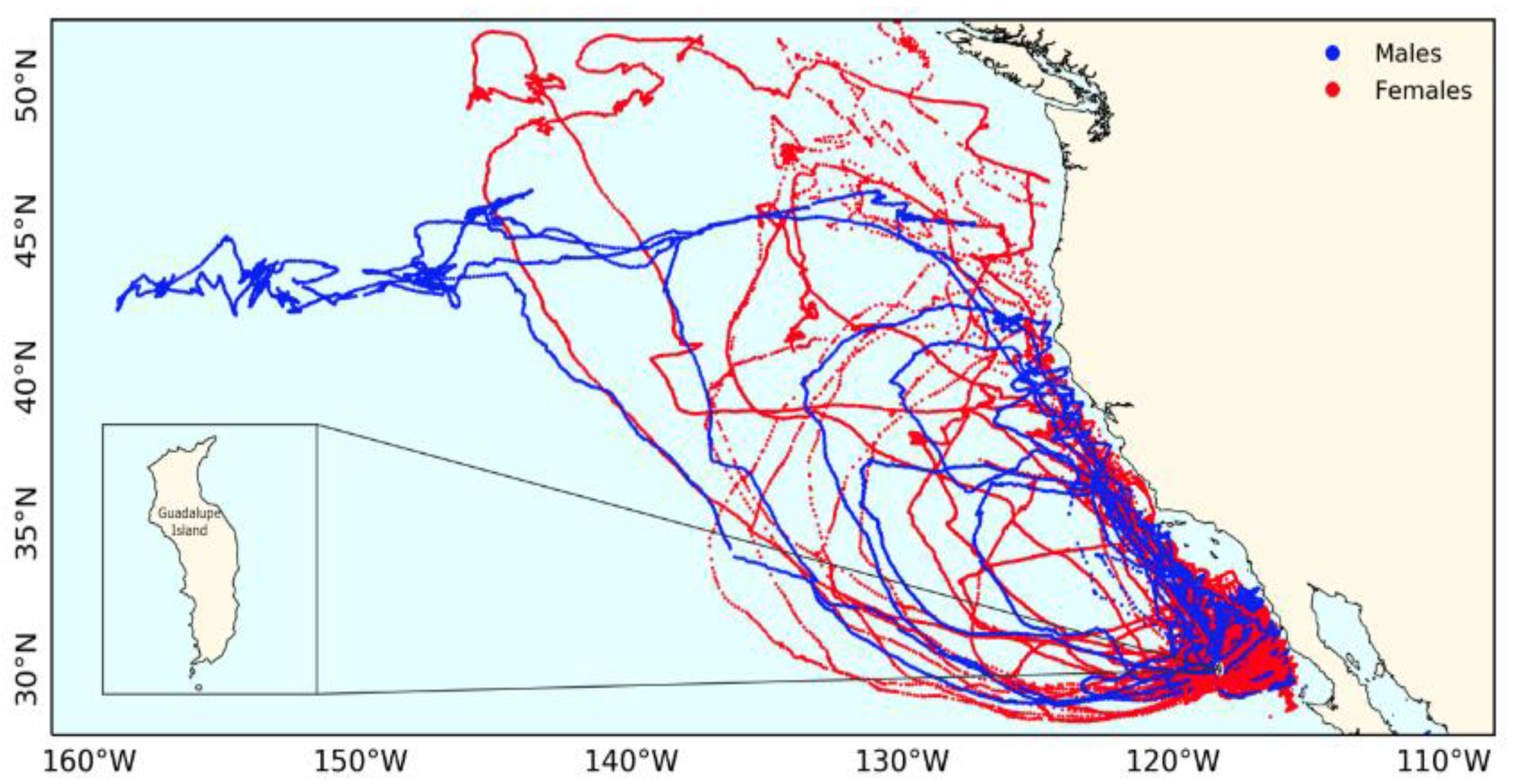
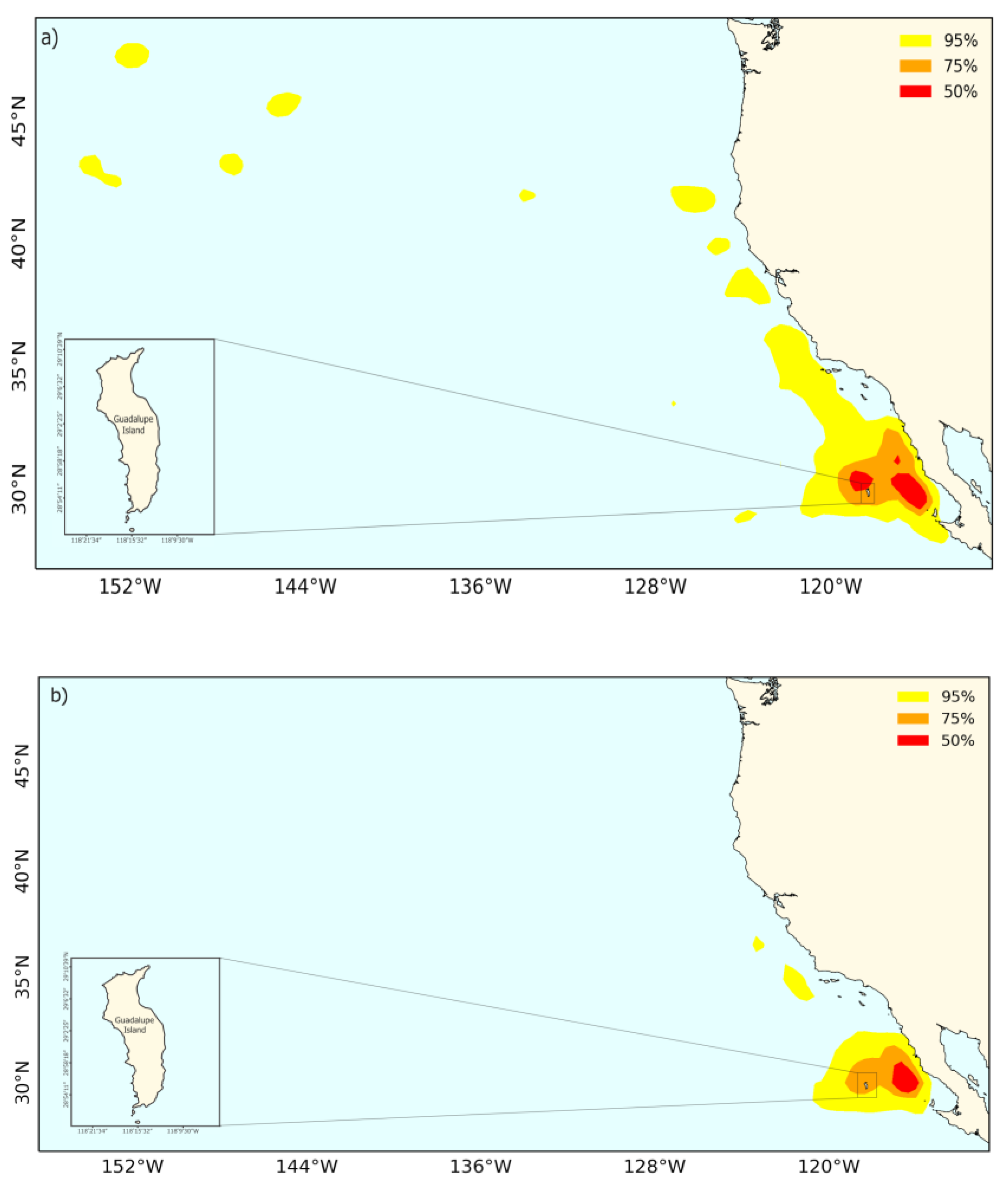
3.4. Laysan Albatross Foraging Trips
4. Discussion
4.1. Sexual Dimorphism
4.2. Sexual Segregation
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Fairbairn, D.J. Allometry for Sexual Size Dimorphism: Pattern and Process in the Coevolution of Body Size in Males and Females. Annu. Rev. Ecol. Syst. 1997, 28, 659–687. [Google Scholar] [CrossRef]
- Hedrick, A.V.; Temeles, E.J. The evolution of sexual dimorphism in animals: Hypotheses and tests. Trends Ecol. Evol. 1989, 4, 136–138. [Google Scholar] [CrossRef]
- Moore, A.J. The evolution of sexual dimorphism by sxual selection: The separate effects of intrasexual selection and intersexual selection. Evol. Int. J. Org. Evol. 1990, 44, 315–331. [Google Scholar] [CrossRef] [PubMed]
- Shine, R. Ecological causes for the evolution of sexual dimorphism: A review of the evidence. Q. Rev. Biol. 1989, 64, 419–461. [Google Scholar] [CrossRef]
- Amundsen, T.; Pärn, H. Female coloration: Review of functional and nonfunctional hypotheses. Bird Coloration 2006, 2, 280–345. [Google Scholar]
- Andersson, M.; Iwasa, Y. Sexual selection. Trends Ecol. Evol. 1996, 11, 53–58. [Google Scholar] [CrossRef]
- Irwin, R.E. The Evolution of Plumage Dichromatism in the New World Blackbirds: Social Selection on Female Brightness. Am. Nat. 1994, 144, 890–907. [Google Scholar] [CrossRef]
- Price, T. Sexual selection and natural selection in bird speciation. Philos. Trans. R. Soc. B 1998, 353, 251–260. [Google Scholar] [CrossRef] [Green Version]
- West-Eberhard, M.J. Sexual Selection, Social Competition, and Speciation. Q. Rev. Biol. 1983, 58, 155–183. [Google Scholar] [CrossRef] [Green Version]
- Owens, I.P.; Hartley, I.R. Sexual dimorphism in birds: Why are there so many different forms of dimorphism? Proc. R. Soc. Lond. Biol. Sci. 1998, 265, 397–407. [Google Scholar] [CrossRef]
- Selander, R.K. Sexual dimorphism and differential niche utilization in birds. Condor 1966, 68, 113–151. [Google Scholar] [CrossRef]
- Clutton-Brock, T.H. Reproductive Success: Studies of Individual Variation in Contrasting Breeding Systems; University of Chicago Press: Chicago, IL, USA, 1988. [Google Scholar]
- Clutton-Brock, T.H. The Evolution of Parental Care; Princeton University Press: Princeton, NJ, USA, 1991. [Google Scholar]
- Ketterson, E.D.; Nolan, V., Jr. Male parental behavior in birds. Annu. Rev. Ecol. Syst. 1994, 25, 601–628. [Google Scholar] [CrossRef]
- Orians, G.H. On the Evolution of Mating Systems in Birds and Mammals. Am. Nat. 1969, 103, 589–603. [Google Scholar] [CrossRef]
- Trivers, R. Parental Investment and Sexual Selection. In Sexual Selection and the Descent of Man; Campbell, B., Ed.; Aldine Press: Chicago, IL, USA, 1972. [Google Scholar]
- Rice, D.W.; Kenyon, K.W. Breeding distribution, history, and populations of North Pacific Albatrosses. Auk 1962, 79, 365–386. [Google Scholar] [CrossRef]
- Fisher, H.I. Some dynamics of a breeding colony of Laysan Albatrosses. Wilson Bull. 1976, 88, 121–142. [Google Scholar]
- Whittow, G.C. Laysan Albatross: Diomedea Immutabilis; American Ornithologists’ Union: Chicago, IL, USA, 1993. [Google Scholar]
- Hernández-Montoya, J.C.; Luna-Mendoza, L.; Aguirre-Muñoz, A.; Méndez-Sánchez, F.; Félix-Lizárraga, M.; Barredo-Barberena, J.M. Laysan Albatross on Guadalupe Island, México: Current status and conservation actions. Monogr. West. N. Am. Nat. 2014, 7, 543–554. [Google Scholar] [CrossRef]
- Howell, S.N.; Webb, S. The sea birds of Las Islas Revillagigedo, Mexico. Wilson Bull. 1990, 102, 140–146. [Google Scholar]
- Henry, R. Consequences of Range Expansion in Laysan Albatrosses. Ph.D. Thesis, University of California-Santa Cruz, Santa Cruz, CA, USA, 2011. [Google Scholar]
- Hyrenbach, K.D.; Fernández, P.; Anderson, D.J. Oceanographic habitats of two sympatric North Pacific albatrosses during the breeding season. Mar. Ecol. Prog. Ser. 2002, 233, 283–301. [Google Scholar] [CrossRef]
- Kappes, M.A.; Shaffer, S.A.; Tremblay, Y.; Foley, D.G.; Palacios, D.M.; Robinson, P.W.; Bograd, S.J.; Costa, D.P. Hawaiian albatrosses track interannual variability of marine habitats in the North Pacific. Prog. Oceanogr. 2010, 86, 246–260. [Google Scholar] [CrossRef]
- Phalan, B.; Phillips, R.A.; Silk, J.R.; Afanasyev, V.; Fukuda, A.; Fox, J.; Catry, P.; Higuchi, H.; Croxall, J.P. Foraging behaviour of four albatross species by night and day. Mar. Ecol. Prog. Ser. 2007, 340, 271–286. [Google Scholar] [CrossRef] [Green Version]
- Fairbairn, J.; Shine, R. Patterns of sexual size dimorphism in seabirds of the Southern Hemisphere. Oikos 1993, 68, 139–145. [Google Scholar] [CrossRef]
- Warham, J. The Petrels: Their Ecology and Breeding Systems; Academic Press: San Diego, CA, USA, 1990. [Google Scholar]
- Phillips, R.A.; Silk, J.R.; Phalan, B.; Catry, P.; Croxall, J.P. Seasonal sexual segregation in two Thalassarche albatross species: Competitive exclusion, reproductive role specialization or foraging niche divergence? Proc. Biol. Sci. 2004, 271, 1283–1291. [Google Scholar] [CrossRef] [PubMed]
- Shaffer, S.A.; Weimerskirch, H.; Costa, D.P. Functional significance of sexual dimorphism in Wandering Albatrosses, Diomedea exulans. Funct. Ecol. 2001, 15, 203–210. [Google Scholar] [CrossRef]
- Templeton, A.R. The meaning of species and speciation: A genetic perspective. In The Units of Evolution: Essays on the Nature of Species; MIT Press: Cambridge, UK, 1992; pp. 159–185. [Google Scholar]
- Wearmouth, V.J.; Sims, D.W. Sexual segregation in marine fish, reptiles, birds and mammals: Behaviour patterns, mechanisms and conservation implications. Adv. Mar. Biol. 2008, 54, 107–170. [Google Scholar] [PubMed]
- Gardali, T.; Seavy, N.E.; DiGaudio, R.T.; Comrack, L.A. A climate change vulnerability assessment of California’s at-risk birds. PLoS ONE 2012, 7, e29507. [Google Scholar] [CrossRef] [PubMed]
- Harwani, S.; Henry, R.W.; Rhee, A.; Kappes, M.A.; Croll, D.A.; Petreas, M.; Park, J.S. Legacy and contemporary persistent organic pollutants in North Pacific albatross. Environ. Toxicol. Chem. 2011, 30, 2562–2569. [Google Scholar] [CrossRef] [PubMed]
- Daunt, F.; Afanasyev, V.; Adam, A.; Croxall, J.; Wanless, S. From cradle to early grave: Juvenile mortality in European shags Phalacrocorax aristotelis results from inadequate development of foraging proficiency. Biol. Lett. 2007, 3, 371–374. [Google Scholar] [CrossRef] [PubMed]
- Frederiksen, M.; Daunt, F.; Harris, M.P.; Wanless, S. The demographic impact of extreme events: Stochastic weather drives survival and population dynamics in a long-lived seabird. J. Anim. Ecol. 2008, 77, 1020–1029. [Google Scholar] [CrossRef] [PubMed]
- Frederiksen, M.; Harris, M.P.; Daunt, F.; Rothery, P.; Wanless, S. Scale-dependent climate signals drive breeding phenology of three seabird species. Glob. Chang. Biol. 2004, 10, 1214–1221. [Google Scholar] [CrossRef]
- Furness, R.W.; Tasker, M.L. Seabird-fishery interactions: Quantifying the sensitivity of seabirds to reductions in sandeel abundance, and identification of key areas for sensitive seabirds in the North Sea. Mar. Ecol. Prog. Ser. 2000, 202, 253–264. [Google Scholar] [CrossRef]
- Kappes, M.A.; Shaffer, S.A.; Tremblay, Y.; Foley, D.G.; Palacios, D.M.; Bograd, S.J.; Costa, D.P. Reproductive constraints influence habitat accessibility, segregation, and preference of sympatric albatross species. Mov. Ecol. 2015, 3, 34. [Google Scholar] [CrossRef] [PubMed]
- Kuletz, K.J.; Renner, M.; Labunski, E.A.; Hunt, G.L., Jr. Changes in the distribution and abundance of albatrosses in the eastern Bering Sea: 1975–2010. Deep Sea Res. Part II Top. Stud. Oceanogr. 2014, 109, 282–292. [Google Scholar] [CrossRef]
- Pitman, R.; Walker, W.; Everet, W.; Gallo-Reynoso, J.P. Population status, foods and foraging of Laysan Albatross Phoebastria immutabilis nesting on Guadalupe Island, Mexico. Mar. Ornithol. 2004, 32, 159–165. [Google Scholar]
- Weimerskirch, H.; Bonadonna, F.; Bailleul, F.; Mabille, G.; Dell’Omo, G.; Lipp, H.-P. GPS tracking of foraging albatrosses. Science 2002, 295, 1259. [Google Scholar] [CrossRef] [PubMed]
- Conners, M.G.; Hazen, E.L.; Costa, D.P.; Shaffer, S.A. Shadowed by scale: Subtle behavioral niche partitioning in two sympatric, tropical breeding albatross species. Mov. Ecol. 2015, 3, 28. [Google Scholar] [CrossRef] [PubMed]
- Fernández, P.; Anderson, D.J.; Sievert, P.R.; Huyvaert, K.P. Foraging destinations of three low-latitude albatross (Phoebastria) species. J. Zool. 2001, 254, 391–404. [Google Scholar] [CrossRef]
- Alarcón, P.A.E.; Morales, J.M.; Donázar, J.A.; Sánchez-Zapata, J.A.; Hiraldo, F.; Lambertucci, S.A. Sexual-size dimorphism modulates the trade-off between exploiting food and wind resources in a large avian scavenger. Sci. Rep. 2017, 7, 11461. [Google Scholar] [CrossRef] [PubMed]
- Nebel, S.; Thompson, G.J. The evolution of sexual bill-size dimorphism in shorebirds: A morphometric test of the resource partitioning hypothesis. Evol. Ecol. Res. 2011, 13, 35–44. [Google Scholar]
- Gallo-Reynoso, J.P.; Figueroa-Carranza, A. The breeding colony of Laysan albatrosses on Isla Guadalupe, México. Western Birds 1996, 27, 70–76. [Google Scholar]
- Fisher, H.I. Mortality and survival in the Laysan Albatross, Diomedea immutabilis. Pac. Sci. 1975, 29, 279–300. [Google Scholar]
- Van Der Werf, E.A.; Young, L.C. Estimating survival and life-stage transitions in the Laysan albatross (Phoebastria Immutabilis) using multistate mark-recapture models. Auk 2011, 128, 726–736. [Google Scholar] [CrossRef]
- Henry, R.W., III; Shaffer, S.A.; Kappes, M.; Tremblay, Y.; Croll, D.A.; Costa, D.P. Movement of the high seas: Consequences of range expansion for a pelagic top consumer, the Laysan Albatross. In Proceedings of the ESA/SER Joint Meeting, Santa Jose, CA, USA, 10 August 2007; pp. 146–148. [Google Scholar]
- Hernández-Montoya, J.C.; Duarte-Canizales, A.; Méndez-Rosas, L.A.; Fernández-Robledo, A.; Luna-Mendoza, L.; Méndez-Sánchez, F.; Aguirre-Muñoz, A.; Iñigo-Elias, E.; Díaz-Castro, S.; Ortega-Rubio, A. Distribution, abundance, and reproductive success of Laysan albatross on Guadalupe Island, México. Manuscript in preparation.
- Lewis, S.; Benvenuti, S.; Dall-Antonia, L.; Griffiths, R.; Money, L.; Sherratt, T.N.; Wanless, S.; Hamer, K.C. Sex-specific foraging behavior in a monomorphic seabird. Proc. R. Soc. Lond. B Biol. Sci. 2002, 269, 1687–1693. [Google Scholar] [CrossRef] [PubMed]
- Young, L.C.; Zaun, B.J.; VanderWerf, E.A. Successful same-sex pairing in Laysan albatross. Biol. Lett. 2008, 4, 323–325. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aljanabi, S.M.; Martinez, I. Universal and rapid salt-extraction of high quality genomic DNA for PCR-based techniques. Nucleic Acids Res. 1997, 25, 4692–4693. [Google Scholar] [CrossRef] [PubMed]
- Fridolfsson, A.-K.; Ellegren, H. A simple and universal method for molecular sexing of non-ratite birds. J. Avian Biol. 1999, 30, 116–121. [Google Scholar] [CrossRef]
- Cuthbert, R.J.; Phillips, R.A.; Ryan, P.G. Separating the Tristan Albatross and the Wandering Albatross using morphometric measurements. Waterbirds 2003, 26, 338–345. [Google Scholar] [CrossRef]
- González-Solís, J. Sexual size dimorphism in northern giant petrels: Ecological correlates and scaling. Oikos 2004, 105, 247–254. [Google Scholar] [CrossRef]
- Phillips, R.A.; Xavier, J.C.; Croxall, J.P. Effects of satellite transmitters on albatrosses and petrels. Auk 2003, 120, 1082–1090. [Google Scholar] [CrossRef]
- Kotzerka, J.; Garthe, S.; Hatch, S.A. GPS tracking devices reveal foraging strategies of Black-legged Kittiwakes. J. Ornithol. 2010, 151, 459–467. [Google Scholar] [CrossRef]
- Wood, A.; Naef-Daenzer, B.; Prince, P.; Croxall, J. Quantifying habitat use in satellite-tracked pelagic seabirds: Application of kernel estimation to albatross locations. J. Avian Biol. 2000, 31, 278–286. [Google Scholar] [CrossRef]
- Worton, B.J. Kernel methods for estimating the utilization distribution in home-range studies. Ecology 1989, 70, 164–168. [Google Scholar] [CrossRef]
- The R Development Core Team. R: A Language and Environment for Statistical Computing. R Foundation for Statistical Computing, Vienna. Available online: www.cran.r-project.org/doc/ (accessed on 26 November 2018).
- Nau, R. Statistical Forecasting: Notes on Regression and Time Series Analysis. Stepwise and All Possible Regressions. Available online: https://people.duke.edu/~rnau/regstep.htm (accessed on 2 May 2019).
- Mayr, E. Populations, Species, and Evolution: An Abridgment of Animal Species and Evolution; Harvard University Press: Cambridge, MA, USA, 1970. [Google Scholar]
- Xavier, J.; Trathan, P.; Croxall, J.; Wood, A.; Podesta, G.; Rodhouse, P. Foraging ecology and interactions with fisheries of wandering albatrosses (Diomedea exulans) breeding at South Georgia. Fish. Oceanogr. 2004, 13, 324–344. [Google Scholar] [CrossRef]
- Tickell, W.L. The Biology of the Great Albatrosses, Diomedea Exulahs and Diomedea Epomophora. In Antarctic Bird Studies; Austin, O.L., Ed.; American Geophysical Union: Washington, DC, USA, 2013. [Google Scholar]
- Granadeiro, J. Variation in measurements of Cory’s Shearwater between populations and sexing by discriminant analysis. Ring. Migr. 1993, 14, 103–112. [Google Scholar] [CrossRef]
- Hamer, K.; Furness, R. Sexing Great Skuas Catharacta skua by discriminant analysis using external measurements. Ring. Migr. 1991, 12, 16–22. [Google Scholar] [CrossRef]
- Weimerskirch, H.; Jouventin, P. Population dynamics of the wandering albatross, Diomedea exulans, of the Crozet Islands: Causes and consequences of the population decline. Oikos 1987, 49, 315–322. [Google Scholar] [CrossRef]
- Prince, P.A.; Wood, A.G.; Barton, T.; Croxall, J.P. Satellite tracking of wandering albatrosses (Diomedea exulans) in the South Atlantic. Antarct. Sci. 1992, 4, 31–36. [Google Scholar] [CrossRef]
- Stahl, J.C.; Sagar, P.M. Foraging strategies and migration of southern Buller’s albatrosses Diomedea b. bulleri breeding on the Solander Islands, New Zealand. J. R. Soc. N. Z. 2000, 30, 319–334. [Google Scholar] [CrossRef]
- Svärdson, G. Competition and habitat selection in birds. Oikos 1949, 1, 157–174. [Google Scholar] [CrossRef]
- Charnov, E.L. Optimal foraging, the marginal value theorem. Theor. Popul. Biol. 1976, 9, 129–136. [Google Scholar] [CrossRef] [Green Version]
- Morris, D.W. Toward an ecological synthesis: A case for habitat selection. Oecologia 2003, 136, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Suryan, R.M.; Anderson, D.J.; Shaffer, S.A.; Roby, D.D.; Tremblay, Y.; Costa, D.P.; Sievert, P.R.; Sato, F.; Ozaki, K.; Balogh, G.R.; et al. Wind, waves, and wing loading: Morphological specialization may limit range expansion of endangered albatrosses. PLoS ONE 2008, 3, e4016. [Google Scholar] [CrossRef] [PubMed]
- Whittow, G. Black-Footed Albatross (Diomedea nigripes); The American Ornithologists’ Union: Chicago, IL, USA, 1993. [Google Scholar]
- Ashmole, N. Seabird ecology and the marine environment. In Avian Biology; Farner, D.S., King, J.R., Eds.; London Academic Press: Cambridge, MA, USA, 1971; pp. 223–286. [Google Scholar]
- Ballance, L.; Pitman, R.; Reilly, S. Seabird community structure along a productivity gradient: Importance of competition and energetic constraint. Ecology 1997, 78, 1502–1518. [Google Scholar] [CrossRef]
- Awkerman, J.; Fukuda, A.; Higuchi, H.; Anderson, D. Foraging activity and submesoscale habitat use of waved albatrosses Phoebastria irrorata during chick-brooding period. Mar. Ecol. Prog. Ser. 2005, 291, 289–300. [Google Scholar] [CrossRef]
- Catry, P.; Phillips, R.A.; Croxall, J.P.; Ruckstuhl, K.; Neuhaus, P. Sexual segregation in birds: Patterns, processes and implications for conservation. In Sexual Segregation in Vertebrates: Ecology of the Two Sexes; Cambridge University Press: Cambridge, UK, 2006; pp. 351–378. [Google Scholar]
- Weimerskirch, H.; Le Corre, M.; Gadenne, H.; Pinaud, D.; Kato, A.; Ropert-Coudert, Y.; Bost, C.A. Relationship between reversed sexual dimorphism, breeding investment and foraging ecology in a pelagic seabird, the masked booby. Oecologia 2009, 161, 637–649. [Google Scholar] [CrossRef] [PubMed]
- Keddy, P.A. Competition; Chapman & Hall: London, UK, 1989. [Google Scholar]
- Amundsen, P.A.; Knudsen, R.; Klemetsen, A. Intraspecific competition and density dependence of food consumption and growth in Arctic charr. J. Anim. Ecol. 2007, 76, 149–158. [Google Scholar] [CrossRef] [PubMed]
- Ashmole, N.P. The regulation of numbers of tropical oceanic birds. Ibis 1963, 103, 458–473. [Google Scholar] [CrossRef]
- Lewis, S.; Sherratt, T.N.; Hamer, K.C.; Wanless, S. Evidence of intra-specific competition for food in a pelagic seabird. Nature 2001, 412, 816–819. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variable | Male | Female | t-Test | p-Value | ||||
|---|---|---|---|---|---|---|---|---|
| SD | CV | SD | CV | |||||
| Cranial length (mm) | 181.80 | 4.10 | 2.26 | 173.61 | 3.92 | 2.26 | −11.84 | <0.001 |
| Bill length (mm) | 113.88 | 2.64 | 2.32 | 107.56 | 2.93 | 2.72 | −13.05 | <0.001 |
| Nostril length (mm) | 85.66 | 2.14 | 2.50 | 81.19 | 2.69 | 3.32 | −10.52 | <0.001 |
| Cranial width (mm) | 54.15 | 1.56 | 2.89 | 51.67 | 2.00 | 3.86 | −7.91 | <0.001 |
| Bill height (mm) | 33.57 | 0.91 | 2.71 | 32.01 | 1.27 | 3.96 | −8.08 | <0.001 |
| Bill width (mm) | 26.20 | 1.19 | 4.55 | 25.12 | 1.34 | 5.32 | −4.90 | <0.001 |
| Tarsus length (mm) | 93.14 | 2.29 | 2.46 | 89.61 | 2.52 | 2.81 | −8.44 | <0.001 |
| Closed wing length (cm) | 52.03 | 1.05 | 2.01 | 51.40 | 1.26 | 2.45 | −3.10 | 0.002 |
| Opened wing length (cm) | 97.46 | 2.07 | 2.12 | 95.58 | 2.16 | 2.26 | −5.14 | <0.001 |
| Wingspan length (cm) | 212.66 | 4.14 | 1.95 | 207.63 | 4.72 | 2.28 | −6.50 | <0.001 |
| Body mass (kg) | 2.86 | 0.34 | 11.79 | 2.44 | 0.23 | 9.54 | −8.55 | <0.001 |
| Variable | PC1 | PC2 |
|---|---|---|
| Cranial length | −0.38 | 0.14 |
| Bill length | −0.37 | 0.23 |
| Nostril length | −0.37 | 0.23 |
| Cranial width | −0.27 | 0.25 |
| Bill height | −0.29 | 0.28 |
| Bill width | −0.27 | 0.21 |
| Tarsus length | −0.33 | 0.13 |
| Closed wing length | −0.23 | −0.60 |
| Opened wing length | −0.30 | −0.46 |
| Wingspan length | −0.34 | −0.33 |
| Standard deviation | 2.32 | 1.17 |
| Proportion of Variance | 0.54 | 0.14 |
| Cumulative Proportion | 0.54 | 0.67 |
| Variable | Estimate | Std. Error | z Value | Pr (>|z|) |
|---|---|---|---|---|
| (Intercept) | −12.466 | 2.765 | −4.508 | 0 |
| Bill length | 8.661 | 2.624 | 3.301 | 0.001 |
| Bill height | 5.911 | 2.662 | 2.221 | 0.026 |
| Cranial width | 3.612 | 1.851 | 1.952 | 0.051 |
| Tarsus length | 4.234 | 2.254 | 1.878 | 0.06 |
| Sex | n | No. of Trips | Trip Length (km) | Maximum Trips Distance (km) | ||
|---|---|---|---|---|---|---|
| SD | SD | |||||
| Males | 16 | 96 | 2270.5 | 3849.60 | 401.00 | 415.80 |
| Females | 20 | 148 | 1874.7 | 2808.20 | 420.90 | 491.50 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hernández Montoya, J.C.; Juárez-Rodríguez, M.; Méndez-Sánchez, F.; Aguirre-Muñoz, A.; Rojas-Mayoral, E.; Íñigo-Elias, E.; Galina-Tessaro, P.; Arnaud, G.; Ortega-Rubio, A. Sexual Dimorphism and Foraging Trips of the Laysan Albatross (Phoebastria immutabilis) on Guadalupe Island. Animals 2019, 9, 364. https://doi.org/10.3390/ani9060364
Hernández Montoya JC, Juárez-Rodríguez M, Méndez-Sánchez F, Aguirre-Muñoz A, Rojas-Mayoral E, Íñigo-Elias E, Galina-Tessaro P, Arnaud G, Ortega-Rubio A. Sexual Dimorphism and Foraging Trips of the Laysan Albatross (Phoebastria immutabilis) on Guadalupe Island. Animals. 2019; 9(6):364. https://doi.org/10.3390/ani9060364
Chicago/Turabian StyleHernández Montoya, Julio César, Maricela Juárez-Rodríguez, Federico Méndez-Sánchez, Alfonso Aguirre-Muñoz, Evaristo Rojas-Mayoral, Eduardo Íñigo-Elias, Patricia Galina-Tessaro, Gustavo Arnaud, and Alfredo Ortega-Rubio. 2019. "Sexual Dimorphism and Foraging Trips of the Laysan Albatross (Phoebastria immutabilis) on Guadalupe Island" Animals 9, no. 6: 364. https://doi.org/10.3390/ani9060364
APA StyleHernández Montoya, J. C., Juárez-Rodríguez, M., Méndez-Sánchez, F., Aguirre-Muñoz, A., Rojas-Mayoral, E., Íñigo-Elias, E., Galina-Tessaro, P., Arnaud, G., & Ortega-Rubio, A. (2019). Sexual Dimorphism and Foraging Trips of the Laysan Albatross (Phoebastria immutabilis) on Guadalupe Island. Animals, 9(6), 364. https://doi.org/10.3390/ani9060364