Transcriptomic Analysis Provides Novel Insights into Heat Stress Responses in Sheep

 ,
,  ,
,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals and Sample Collection
2.2. Detection of Thyroid-Related Hormones in Sheep Serum
2.3. RNA Extraction and Quality Inspection
2.4. cDNA Library Construction and Sequencing
2.5. Raw Data Preprocessing and Alignment
2.6. Identification and Analysis of Differentially Expressed Genes
2.7. Functional Enrichment Analysis of Differentially Expressed Genes
2.8. Verification of Differentially Expressed Genes
2.9. Statistical Analysis
3. Results
3.1. Influence of Heat Stress on Serum T3 and T4 Levels of Sheep
3.2. Summary of Sheep Pen THI and Sequencing Data
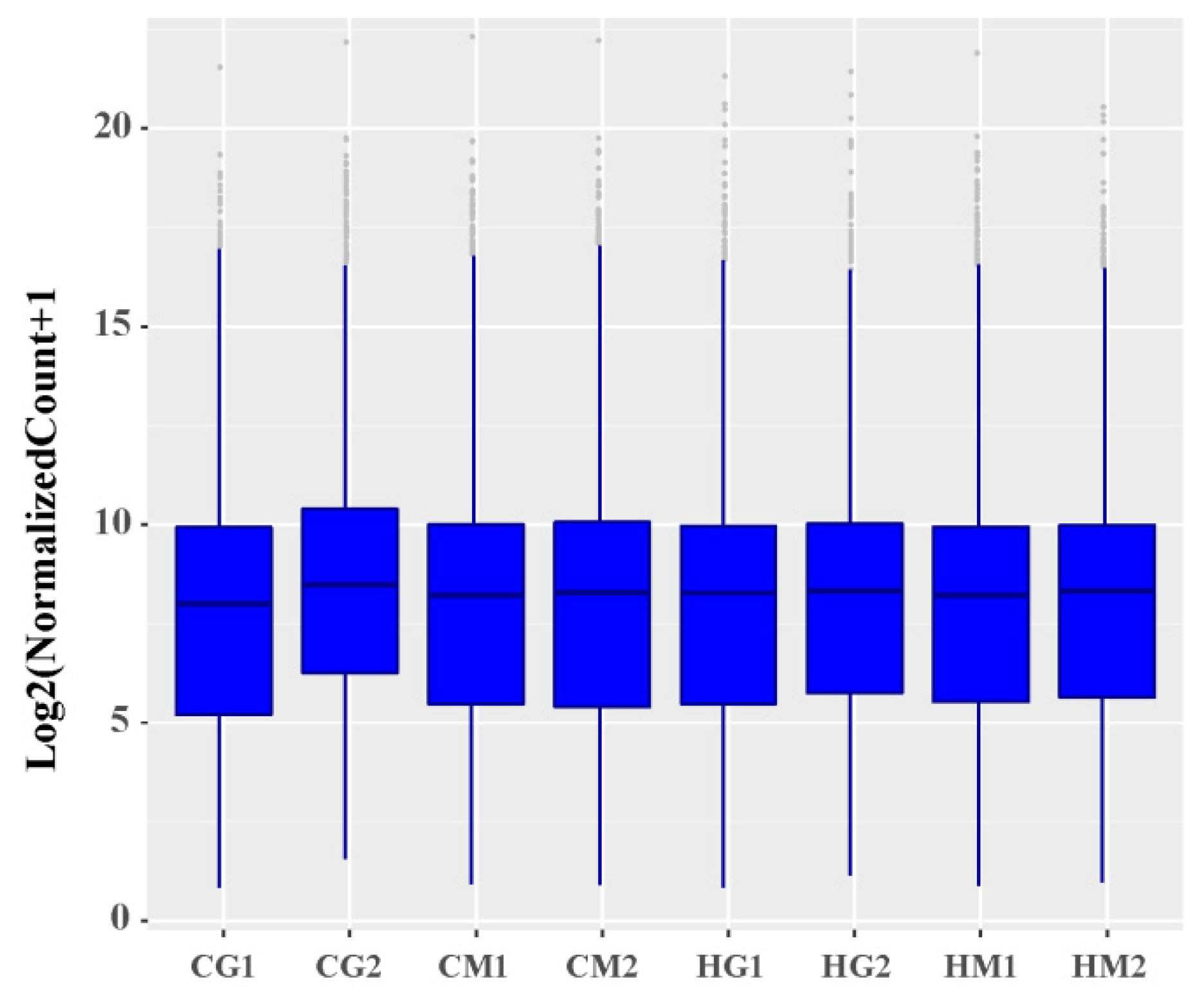
3.3. Analysis of Overall Gene Expression Levels
3.4. Screening and Clustering Analysis of Differentially Expressed Genes
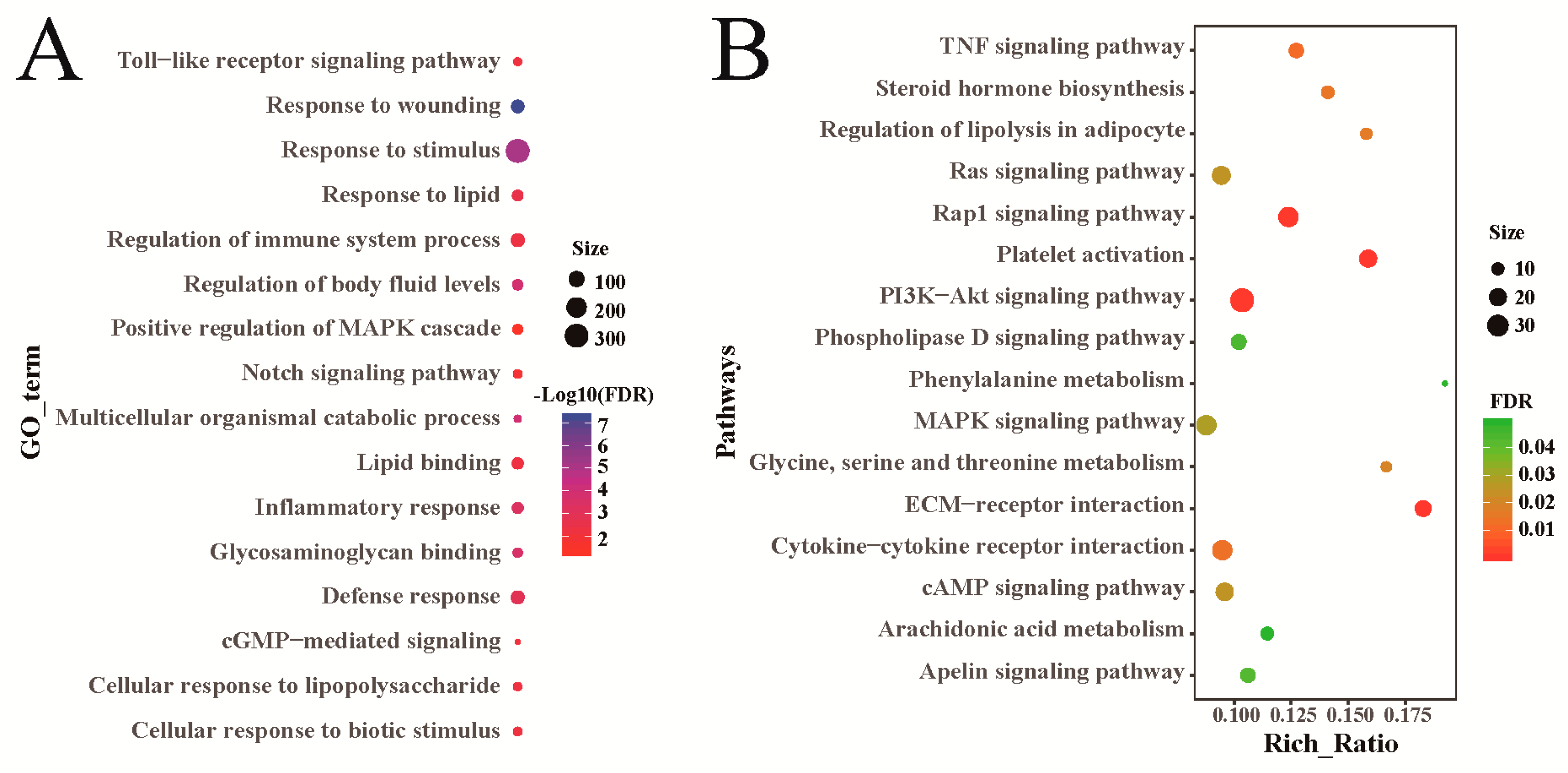
3.5. Functional Enrichment Analysis of Differentially Expressed Genes
3.6. Validation of RNA-Seq Data by qRT-PCR
4. Discussion
4.1. T3 and T4
4.2. Regulation of Body Temperature
4.3. Regulation of Stress Reactions
4.4. Regulation of Energy Metabolism
4.5. Regulation of Immunoreactions
5. Conclusions
Availability of Supporting Data
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| ACTB | β-actin |
| ANGPT2 | angiopoietin 2 |
| cAMP | cyclic adenosine monophosphate |
| cGMP | cyclic guanosinc monophosphate |
| FPKM | fragments per kilobase of transcript per million fragments mapped |
| GO | gene ontology |
| HSP70 | heat stress protein 70 |
| HTR4 | 5-Hydroxytryptamine receptor 4 |
| IL1R | interleukinl type 1 receptor |
| KEGG | kyoto encyclopedia of genes and genomes |
| MAPK | mitogen-activated protein kinase |
| NPR1 | natriuretic peptide receptor 1 |
| PI3K-AKt | phosphatidylinositol 3-kinase and protein kinase B |
| PKA | protein kinase A |
| qRT-PCR | quantitative real time PCR |
| RH | relative humidity |
| SLC13A5 | sodium-dependent citrate transporter member 5 |
| Td | temperature |
| THI | temperature-humidity index |
References
- Pantoja, M.H.A.; Esteves, S.N.; Jacinto, M.A.C.; Pezzopane, J.R.M.; Paz, C.C.P.; Silva, J.; Lourenco Junior, J.B.; Brandao, F.Z.; Moura, A.B.B.; Romanello, N.; et al. Thermoregulation of male sheep of indigenous or exotic breeds in a tropical environment. J. Therm. Biol. 2017, 69, 302–310. [Google Scholar] [CrossRef] [PubMed]
- Vilas Boas Ribeiro, B.P.; Lanferdini, E.; Palencia, J.Y.P.; Lemes, M.A.G.; Teixeira de Abreu, M.L.; de Souza Cantarelli, V.; Ferreira, R.A. Heat negatively affects lactating swine: A meta-analysis. J. Therm. Biol. 2018, 74, 325–330. [Google Scholar] [CrossRef]
- Cui, Y.; Hao, Y.; Li, J.; Bao, W.; Li, G.; Gao, Y.; Gu, X. Chronic heat stress induces immune response, oxidative stress response, and apoptosis of finishing pig liver: A proteomic approach. Int. J. Mol. Sci. 2016, 17, 393. [Google Scholar] [CrossRef]
- Daramola, J.O.; Adeloye, A.A. Physiological adaptation to the humid tropics with special reference to the West African Dwarf (WAD) goat. Trop. Anim. Health Prod. 2009, 41, 1005–1016. [Google Scholar] [CrossRef] [PubMed]
- Marai, I.F.M.; Habeeb, A.A.M.; Gad, A.E. Reproductive traits of female rabbits as affected by heat stress and lighting regime under subtropical conditions of Egypt. Anim. Sci. 2004, 78, 119–127. [Google Scholar] [CrossRef]
- Li, F.K.; Yang, Y.; Jenna, K.; Xia, C.H.; Lv, S.J.; Wei, W.H. Effect of heat stress on the behavioral and physiological patterns of Small-tail Han sheep housed indoors. Trop. Anim. Health Prod. 2018, 50, 1893–1901. [Google Scholar] [CrossRef]
- Rathwa, S.D.; Vasava, A.A.; Pathan, M.M.; Madhira, S.P.; Patel, Y.G.; Pande, A.M. Effect of season on physiological, biochemical, hormonal, and oxidative stress parameters of indigenous sheep. Vet. World 2017, 10, 650–654. [Google Scholar] [CrossRef] [Green Version]
- Das, R.; Sailo, L.; Verma, N.; Bharti, P.; Saikia, J.; Imtiwati; Kumar, R. Impact of heat stress on health and performance of dairy animals: A review. Vet. World 2016, 9, 260–268. [Google Scholar] [CrossRef]
- De, K.; Kumar, D.; Saxena, V.K.; Thirumurugan, P.; Naqvi, S.M.K. Effect of high ambient temperature on behavior of sheep under semi-arid tropical environment. Int. J. Biometeorol. 2017, 61, 1269–1277. [Google Scholar] [CrossRef]
- Narayan, E.; Sawyer, G.; Parisella, S. Faecal glucocorticoid metabolites and body temperature in Australian merino ewes (Ovis aries) during summer artificial insemination (AI) program. PLoS ONE 2018, 13, e0191961. [Google Scholar] [CrossRef]
- Dahl, G.E.; Tao, S.; Monteiro, A.P.A. Effects of late-gestation heat stress on immunity and performance of calves. J. Dairy Sci. 2016, 99, 3193–3198. [Google Scholar] [CrossRef]
- Saelao, P.; Wang, Y.; Gallardo, R.A.; Lamont, S.J.; Dekkers, J.M.; Kelly, T.; Zhou, H. Novel insights into the host immune response of chicken Harderian gland tissue during Newcastle disease virus infection and heat treatment. BMC Vet. Res. 2018, 14, 280. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Gerstein, M.; Snyder, M. RNA-Seq: A revolutionary tool for transcriptomics. Nat. Rev. Genet. 2009, 10, 57–63. [Google Scholar] [CrossRef] [PubMed]
- Marai, I.F.M.; El-Darawany, A.A.; Fadiel, A.; Abdel-Hafez, M.A.M. Physiological traits as affected by heat stress in sheep—A review. Small Rumin. Res. 2007, 71, 1–12. [Google Scholar] [CrossRef]
- Baumgard, L.H.; Rhoads, R.P., Jr. Effects of heat stress on postabsorptive metabolism and energetics. Annu. Rev. Anim. Biosci. 2013, 1, 311–337. [Google Scholar] [CrossRef] [PubMed]
- Salleh, S.M.; Mazzoni, G.; Lovendahl, P.; Kadarmideen, H.N. Gene co-expression networks from RNA sequencing of dairy cattle identifies genes and pathways affecting feed efficiency. BMC Bioinform. 2018, 19, 513. [Google Scholar] [CrossRef]
- Horodyska, J.; Wimmers, K.; Reyer, H.; Trakooljul, N.; Mullen, A.M.; Lawlor, P.G.; Hamill, R.M. RNA-seq of muscle from pigs divergent in feed efficiency and product quality identifies differences in immune response, growth, and macronutrient and connective tissue metabolism. BMC Genom. 2018, 19, 791. [Google Scholar] [CrossRef]
- Khan, S.; Wu, S.B.; Roberts, J. RNA-sequencing analysis of shell gland shows differences in gene expression profile at two time-points of eggshell formation in laying chickens. BMC Genom. 2019, 20, 89. [Google Scholar] [CrossRef]
- Koluman, N.; Daskiran, I. Effects of ventilation of the sheep house on heat stress, growth and thyroid hormones of lambs. Trop. Anim. Health Prod. 2011, 43, 1123–1127. [Google Scholar] [CrossRef]
- Todini, L.; Malfatti, A.; Valbonesi, A.; Trabalza-Marinucci, M.; Debenedetti, A. Plasma total T3 and T4 concentrations in goats at different physiological stages, as affected by the energy intake. Small Rumin. Res. 2007, 68, 285–290. [Google Scholar] [CrossRef]
- Cox, B.; Lee, T.F. Further evidence for a physiological role for hypothalamic dopamine in thermoregulation in the rat. J. Physiol. 1980, 300, 7–17. [Google Scholar] [CrossRef] [PubMed]
- Boulant, J.A. Role of the preoptic-anterior hypothalamus in thermoregulation and fever. Clin. Infect. Dis. 2000, 31 (Suppl. 5), S157–S161. [Google Scholar] [CrossRef] [PubMed]
- Lucas, G. Serotonin receptors, type 4: A new hope? Curr. Drug Targets 2009, 10, 1085–1095. [Google Scholar] [CrossRef] [PubMed]
- Jin, Z.Y.; Ku, B.S.; Yao, H.Y.; Li, Z.H.; Rao, Y. Relationship between the function of 5-HT1A,2 receptors and waking-sleeping stages. Chin. Pharmacol. Bull. 2000, 16, 208–211. [Google Scholar]
- Frische, E.W.; Zwartkruis, F.J. Rap1, a mercenary among the Ras-like GTPases. Dev. Biol. 2010, 340, 1–9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, C.; Ramirez, A.; Cui, B.; Ding, J.; Delcroix, J.D.; Valletta, J.S.; Liu, J.J.; Yang, Y.; Chu, S.; Mobley, W.C. A functional dynein-microtubule network is required for NGF signaling through the Rap1/MAPK pathway. Traffic 2007, 8, 1503–1520. [Google Scholar] [CrossRef] [PubMed]
- Kam, C.Y.; Dubash, A.D.; Magistrati, E.; Polo, S.; Satchell, K.J.F.; Sheikh, F.; Lampe, P.D.; Green, K.J. Desmoplakin maintains gap junctions by inhibiting Ras/MAPK and lysosomal degradation of connexin-43. J. Cell Biol. 2018, 217, 3219–3235. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, Z.Z.; Tschopp, O.; Baudry, A.; Dummler, B.; Hynx, D.; Hemmings, B.A. Physiological functions of protein kinase B/Akt. Biochem. Soc. Trans. 2004, 32, 350–354. [Google Scholar] [CrossRef] [PubMed]
- Borrie, S.C.; Brems, H.; Legius, E.; Bagni, C. Cognitive dysfunctions in intellectual disabilities: The contributions of the Ras-MAPK and PI3K-AKT-mTOR pathways. Annu. Rev. Genom. Hum. Genet. 2017, 18, 115–142. [Google Scholar] [CrossRef]
- Lee, H.W.; Rhee, D.K.; Kim, B.O.; Pyo, S. Inhibitory effect of sinigrin on adipocyte differentiation in 3T3-L1 cells: Involvement of AMPK and MAPK pathways. Biomed. Pharmacother. 2018, 102, 670–680. [Google Scholar] [CrossRef]
- Wu, M.; Liu, D.; Zeng, R.; Xian, T.; Lu, Y.; Zeng, G.; Sun, Z.; Huang, B.; Huang, Q. Epigallocatechin-3-gallate inhibits adipogenesis through down-regulation of PPARgamma and FAS expression mediated by PI3K-AKT signaling in 3T3-L1 cells. Eur. J. Pharmacol. 2017, 795, 134–142. [Google Scholar] [CrossRef] [PubMed]
- Reverte-Salisa, L.; Sanyal, A.; Pfeifer, A. Role of cAMP and cGMP signaling in brown fat. In Handbook of Experimental Pharmacology; Springer: Cham, Switzerland, 2018; Volume 251, pp. 161–182. [Google Scholar]
- Huang, H.Y.; Zhao, G.P.; Liu, R.R.; Li, Q.H.; Zheng, M.Q.; Li, S.F.; Liang, Z.; Zhao, Z.H.; Wen, J. Brain natriuretic peptide stimulates lipid metabolism through its receptor NPR1 and the glycerolipid metabolism pathway in chicken adipocytes. Biochemistry 2015, 54, 6622–6630. [Google Scholar] [CrossRef] [PubMed]
- Rudnicki, M.; Abdifarkosh, G.; Rezvan, O.; Nwadozi, E.; Roudier, E.; Haas, T.L. Female mice have higher angiogenesis in perigonadal adipose tissue than males in response to high-fat diet. Front. Physiol. 2018, 9, 1452. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.Y.; Jiang, J.; Ma, H.M. Molecular characterization, tissue expression profile, and SNP analysis of porcine SLC13A5. Genet. Mol. Res. 2015, 14, 16090–16101. [Google Scholar] [CrossRef] [PubMed]
- Balasubramanian, S.P.; Azmy, I.A.; Higham, S.E.; Wilson, A.G.; Cross, S.S.; Cox, A.; Brown, N.J.; Reed, M.W. Interleukin gene polymorphisms and breast cancer: A case control study and systematic literature review. BMC Cancer 2006, 6, 188. [Google Scholar] [CrossRef] [PubMed]
- Zininga, T.; Ramatsui, L.; Shonhai, A. Heat shock proteins as immunomodulants. Molecules 2018, 23, 2846. [Google Scholar] [CrossRef] [PubMed]
- Binder, R.J. Heat-shock protein-based vaccines for cancer and infectious disease. Expert Rev. Vaccines 2008, 7, 383–393. [Google Scholar] [CrossRef]
- Shevtsov, M.; Multhoff, G. Heat shock protein-peptide and HSP-based immunotherapies for the treatment of cancer. Front. Immunol. 2016, 7, 171. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Item | Sex | Number | Control Group | Heat Stress Group |
|---|---|---|---|---|
| T3 (ng/mL) | ♂ | 5 | 3.92 ± 0.041 A | 1.24 ± 0.020 B |
| ♀ | 5 | 3.89 ± 0.045 A | 1.28 ± 0.016 B | |
| T4 (ng/mL) | ♂ | 5 | 82.92 ± 1.55 A | 41.89 ± 1.23 B |
| ♀ | 5 | 76.05 ± 1.17 A | 42.73 ± 0.74 B |
| Items | CG1 | CG2 | CM1 | CM2 | HG1 | HG2 | HM1 | HM2 |
|---|---|---|---|---|---|---|---|---|
| Raw reads | 69,008,628 | 39,367,262 | 69,141,276 | 68,952,966 | 78,420,396 | 51,776,410 | 76,392,674 | 60,428,044 |
| Clean reads | 68,367,484 | 38,903,468 | 68,478,224 | 68,463,512 | 77,762,862 | 51,308,882 | 75,471,938 | 59,933,578 |
| Clean ratio (%) | 99.07 | 98.82 | 99.04 | 99.29 | 99.16 | 99.1 | 98.79 | 99.18 |
| GC content (%) | 49.82 | 50.09 | 51.19 | 50.92 | 51.18 | 51.04 | 50.81 | 51.25 |
| Q20 (%) | 98.65 | 98.6 | 98.5 | 98.7 | 98.65 | 98.5 | 98.1 | 98.6 |
| Q30 (%) | 95.8 | 95.75 | 95.35 | 95.9 | 95.8 | 95.35 | 94.4 | 95.55 |
| Total mapped | 65,863,115 (96.34%) | 36,273,376 (93.24%) | 65,257,697 (95.30%) | 65,546,575 (95.74%) | 73,440,220 (94.44%) | 48,208,876 (93.96%) | 71,214,523 (94.36%) | 56,581,847 (94.41%) |
| Multiple mapped | 6,565,779 (9.60%) | 2,743,494 (7.05%) | 6,088,334 (8.89%) | 6,757,881 (9.87%) | 7,483,120 (9.62%) | 6,034,386 (11.76%) | 9,336,634 (12.37%) | 5,753,481 (9.60%) |
| Unique mapped | 59,297,336 (86.73%) | 33,529,882 (86.19%) | 59,169,363 (86.41%) | 58,788,694 (85.87%) | 65,957,100 (84.82%) | 42,174,490 (82.20%) | 61,877,889 (81.99%) | 50,828,366 (84.81%) |
| Unmapped | 2,504,369 (3.66%) | 2,630,092 (6.76%) | 3,220,527 (4.70%) | 2,916,937 (4.26%) | 4,322,642 (5.56%) | 3,100,006 (6.04%) | 4,257,415 (5.64%) | 3,351,731 (5.59%) |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lu, Z.; Chu, M.; Li, Q.; Jin, M.; Fei, X.; Ma, L.; Zhang, L.; Wei, C. Transcriptomic Analysis Provides Novel Insights into Heat Stress Responses in Sheep. Animals 2019, 9, 387. https://doi.org/10.3390/ani9060387
Lu Z, Chu M, Li Q, Jin M, Fei X, Ma L, Zhang L, Wei C. Transcriptomic Analysis Provides Novel Insights into Heat Stress Responses in Sheep. Animals. 2019; 9(6):387. https://doi.org/10.3390/ani9060387
Chicago/Turabian StyleLu, Zengkui, Mingxing Chu, Qing Li, Meilin Jin, Xiaojuan Fei, Lin Ma, Liping Zhang, and Caihong Wei. 2019. "Transcriptomic Analysis Provides Novel Insights into Heat Stress Responses in Sheep" Animals 9, no. 6: 387. https://doi.org/10.3390/ani9060387
APA StyleLu, Z., Chu, M., Li, Q., Jin, M., Fei, X., Ma, L., Zhang, L., & Wei, C. (2019). Transcriptomic Analysis Provides Novel Insights into Heat Stress Responses in Sheep. Animals, 9(6), 387. https://doi.org/10.3390/ani9060387