Effect of Acute Heat Stress on the mRNA Levels of Cytokines in Broiler Chickens Subjected to Embryonic Thermal Manipulation


Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Egg Procurement and Incubation Conditions
2.2. Rearing and Exposure to Acute Heat Stress (AHS)
2.3. RNA Extraction and cDNA Synthesis
2.4. Real-Time RT-qPCR Analysis
2.5. Plasma Analysis
2.6. Statistical Analysis
3. Results
3.1. Effect of Thermal Manipulation (TM) on the Hatchability Rate and Post-Hatch Body Weights of Broiler Chickens
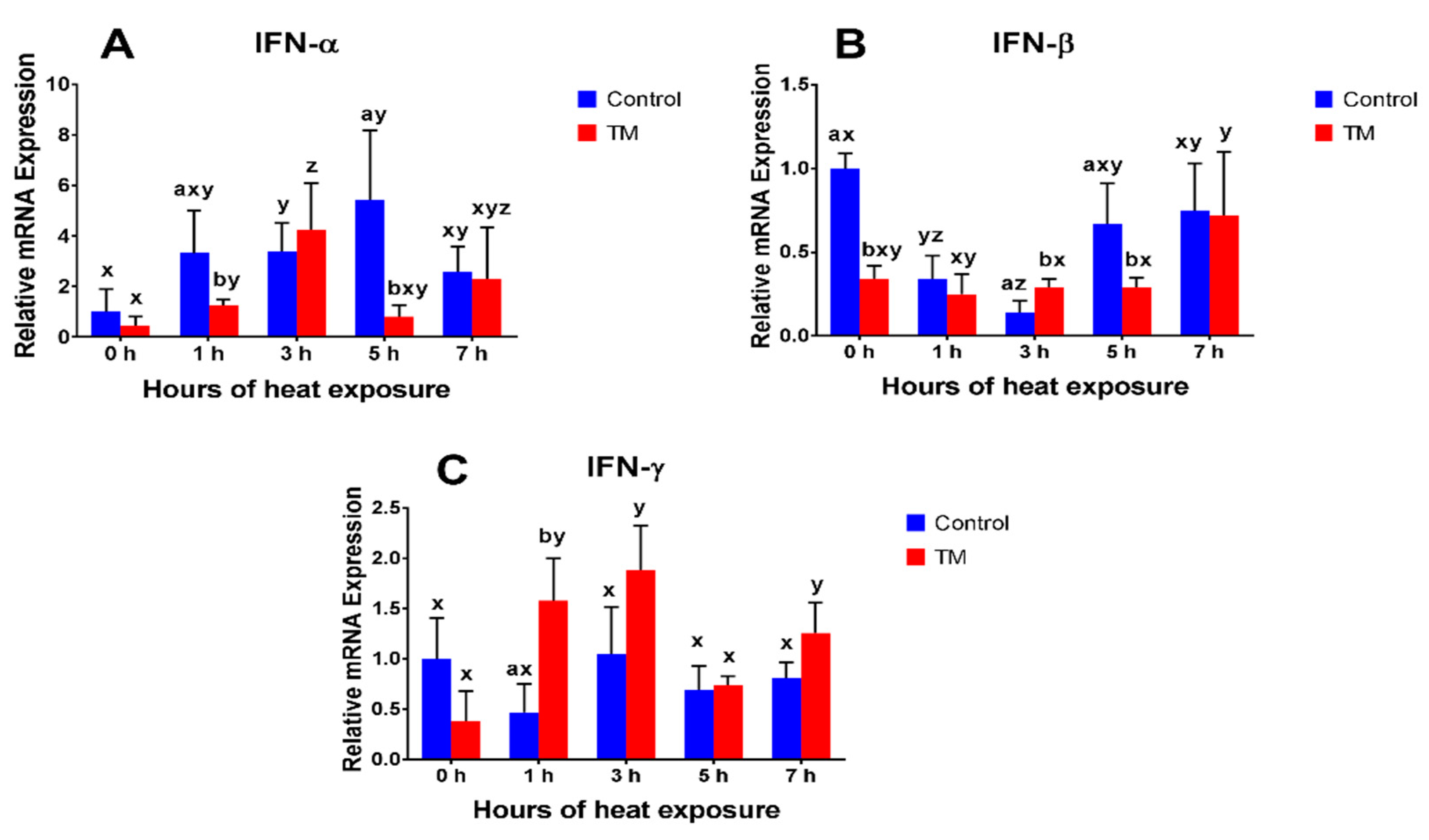
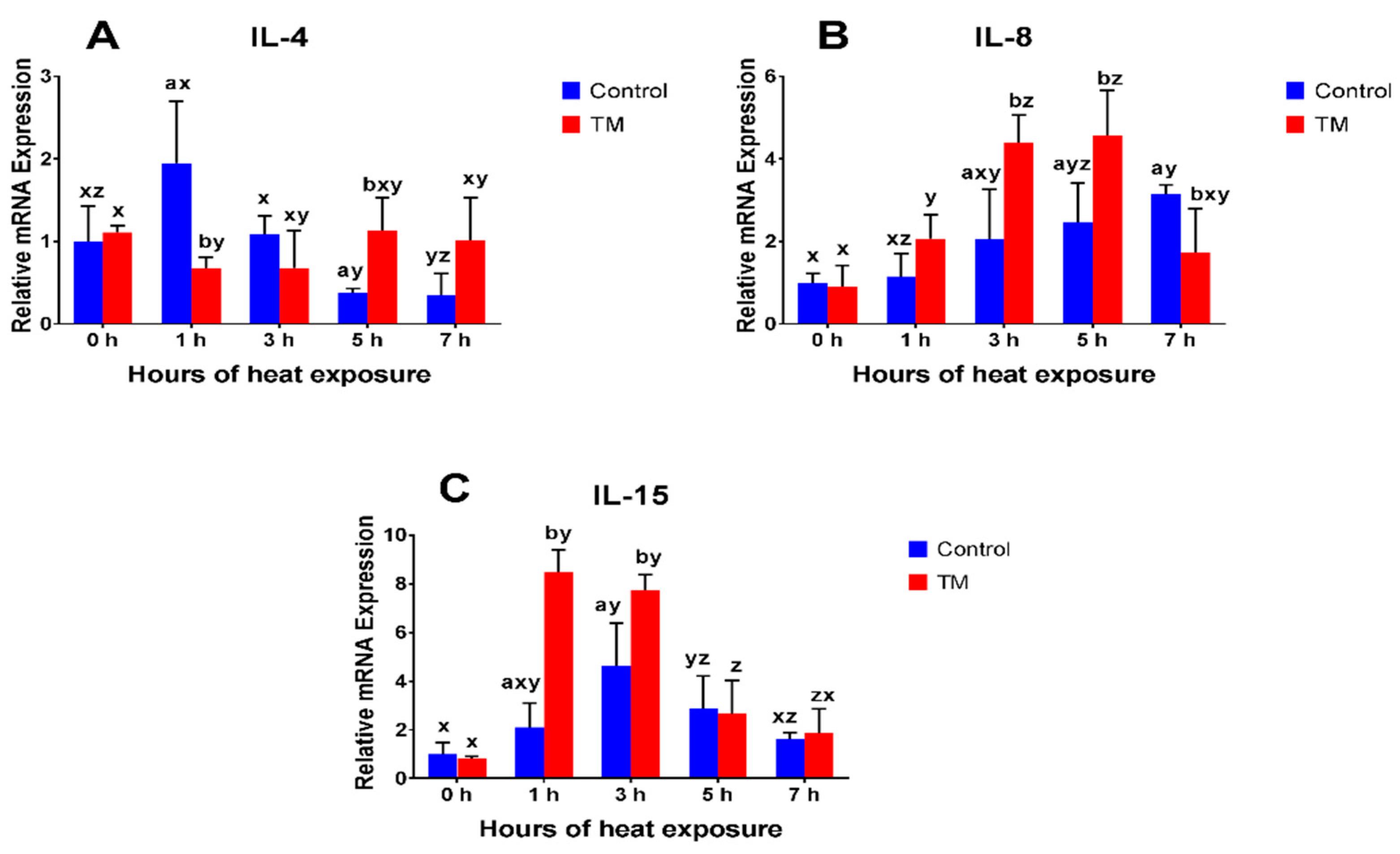
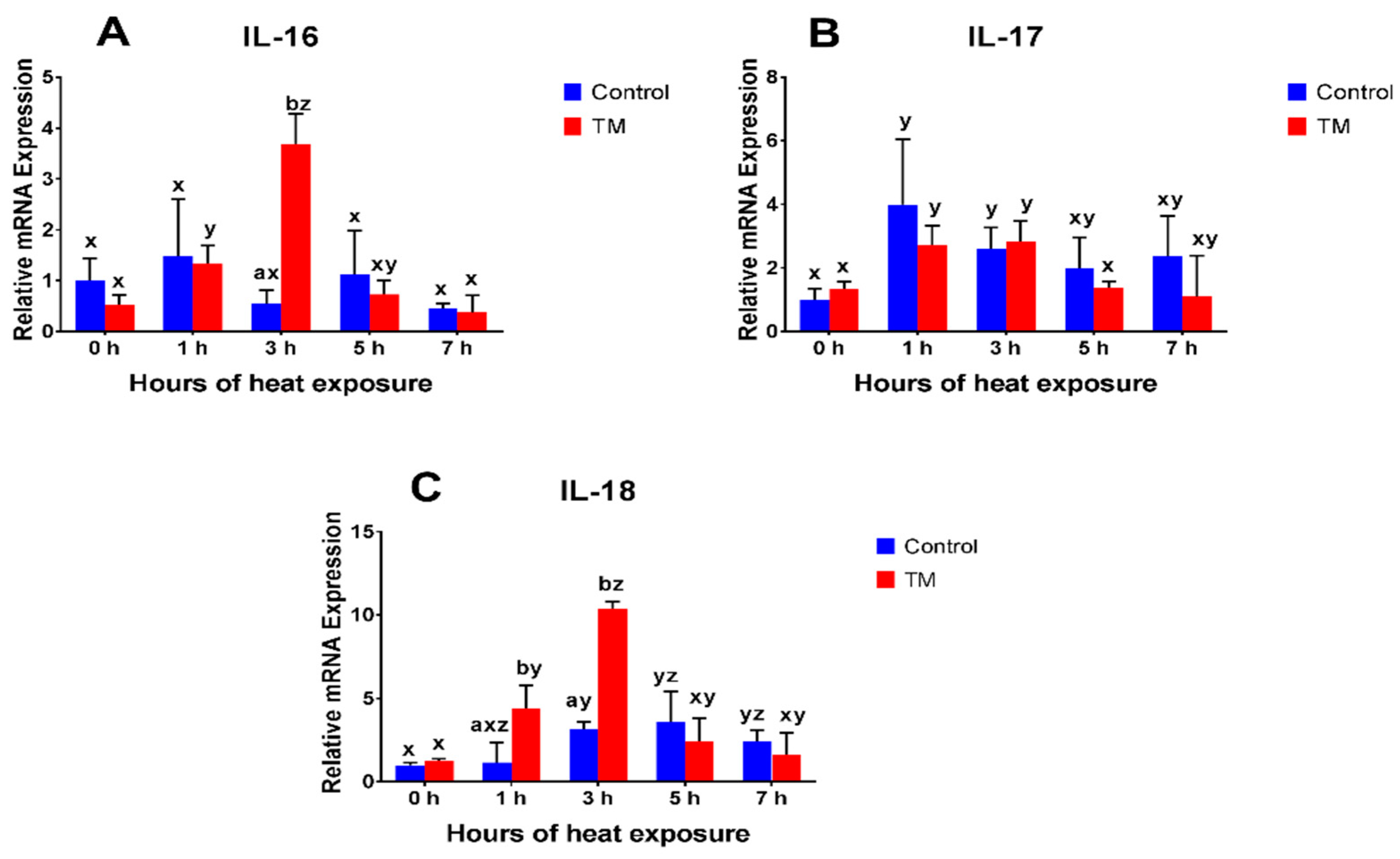
3.2. Effect of Thermal Manipulation (TM) and Acute Heat Stress (AHS) on mRNA Levels of Splenic Cytokines
3.2.1. IFN-α
3.2.2. IFN-β
3.2.3. IFN-γ
3.2.4. IL-4
3.2.5. IL-8
3.2.6. IL-15
3.2.7. IL-16
3.2.8. IL-17
3.2.9. IL-18
3.3. Effect of Thermal Manipulation (TM) and Acute Heat Stress (AHS) on IL-4, IL-8, and IFN-γ Plasma Levels
3.3.1. IL-4
3.3.2. IL-8
3.3.3. IFN-γ
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Nardone, A.; Ronchi, B.; Lacetera, N.; Ranieri, M.S.; Bernabucci, U. Effects of climate changes on animal production and sustainability of livestock systems. Livest. Sci. 2010, 130, 57–69. [Google Scholar] [CrossRef]
- Mashaly, M.; Hendricks, G.L.; Kalama, M.A.; Gehad, A.E.; Abbas, A.O.; Patterson, P.H. Effect of Heat Stress on Production Parameters and Immune Responses of Commercial Laying Hens. Poult. Sci. 2004, 83, 889–894. [Google Scholar] [CrossRef] [PubMed]
- Quinteiro-Filho, W.; Ribeiro, A.; Ferraz-de-Paula, V.; Pinheiro, M.; Sakai, M.; Sa, L.; Ferreira, A.; Palermo-Neto, J. Heat stress impairs performance parameters, induces intestinal injury, and decreases macrophage activity in broiler chickens. Poult. Sci. 2010, 89, 1905–1914. [Google Scholar] [CrossRef] [PubMed]
- Herman, J.P.; Cullinan, W.E. Neurocircuitry of stress: Central control of the hypothalamo-pituitary-adrenocortical axis. Trends Neurosci. 1997, 20, 78–84. [Google Scholar] [CrossRef]
- Ohtsu, H.; Yamazaki, M.; Abe, H.; Murakami, H.; Toyomizu, M. Heat Stress Modulates Cytokine Gene Expression in the Spleen of Broiler Chickens. J. Poult. Sci. 2015, 52, 282–287. [Google Scholar] [CrossRef] [Green Version]
- Quinteiro-Filho, W.M.; Calefi, A.S.; Cruz, D.S.G.; Aloia, T.P.A.; Zager, A.; Astolfi-Ferreira, C.S.; Ferreira, J.A.P.; Sharif, S.; Palermo-Neto, J. Heat stress decreases expression of the cytokines, avian β-defensins 4 and 6 and Toll-like receptor 2 in broiler chickens infected with Salmonella Enteritidis. Vet. Immunol. Immunopathol. 2017, 186, 19–28. [Google Scholar] [CrossRef] [PubMed]
- Giansanti, F.; Giardi, M.; Botti, D. Avian cytokines-an overview. Curr. Pharm. Des. 2006, 12, 3083–3099. [Google Scholar] [CrossRef]
- Davison, F.; Kaspers, B.; Schat, K.A.; Kaiser, P. Avian Immunology; Academic Press: New York, NY, USA, 2011. [Google Scholar]
- Kaiser, P. Advances in avian immunology—Prospects for disease control: A review. Avian Pathol. 2010, 39, 309–324. [Google Scholar] [CrossRef]
- Kaiser, P.; Stäheli, P. Avian cytokines and chemokines. In Avian Immunology, 2nd ed.; Elsevier: Amsterdam, The Netherlands, 2014; pp. 189–204. [Google Scholar]
- Hietbrink, F.; Koenderman, L.; Rijkers, G.T.; Leenen, L.P. Trauma: The role of the innate immune system. World J. Emerg. Surg. 2006, 1, 15. [Google Scholar] [CrossRef]
- Heled, Y.; Fleischmann, C.; Epstein, Y. Cytokines and their role in hyperthermia and heat stroke. J. Basic Clin. Physiol. Pharmacol. 2013, 24, 85–96. [Google Scholar] [CrossRef]
- Varasteh, S.; Braber, S.; Akbari, P.; Garssen, J.; Fink-Gremmels, J. Differences in Susceptibility to Heat Stress along the Chicken Intestine and the Protective Effects of Galacto-Oligosaccharides. PLoS ONE 2015, 10, e0138975. [Google Scholar] [CrossRef]
- Song, Z.; Cheng, K.; Zhang, L.; Wang, T. Dietary supplementation of enzymatically treated Artemisia annua could alleviate the intestinal inflammatory response in heat-stressed broilers. J. Therm. Biol. 2017, 69, 184–190. [Google Scholar] [CrossRef]
- Al-Zghoul, M.B.; Saleh, K.M.; Ababneh, M.M.K. Effects of pre-hatch thermal manipulation and post-hatch acute heat stress on the mRNA expression of interleukin-6 and genes involved in its induction pathways in 2 broiler chicken breeds. Poult. Sci. 2018, pey499. [Google Scholar] [CrossRef]
- Piestun, Y.; Shinder, D.; Ruzal, M.; Halevy, O.; Brake, J.; Yahav, S. Thermal manipulations during broiler embryogenesis: Effect on the acquisition of thermotolerance. Poult. Sci. 2008, 87, 1516–1525. [Google Scholar] [CrossRef]
- Molenaar, R.; Reijrink, I.A.M.; Meijerhof, R.; Van den Brand, H. Meeting embryonic requirements of broilers throughout incubation: A review. Rev. Bras. Cienc. Avic. 2010, 12, 137–148. [Google Scholar] [CrossRef]
- Al-Rukibat, R.K.; Al-Zghoul, M.B.; Hananeh, W.M.; Al-Natour, M.Q.; Abu-Basha, E.A. Thermal manipulation during late embryogenesis: Effect on body weight and temperature, thyroid hormones, and differential white blood cell counts in broiler chickens. Poult. Sci. 2017, 96, 234–240. [Google Scholar] [CrossRef]
- Al-Zghoul, M.B.; Ismail, Z.B.; Dalab, A.E.; Al-Ramadan, A.; Althnaian, T.A.; Al-Ramadan, S.Y.; Ali, A.M.; Albokhadaim, I.F.; Al Busadah, K.A.; Eljarah, A.; et al. Hsp90, Hsp60 and HSF-1 genes expression in muscle, heart and brain of thermally manipulated broiler chicken. Res. Vet. Sci. 2015, 99, 105–111. [Google Scholar] [CrossRef]
- Al-Zghoul, M.B.; El-Bahr, S.M.; Al-Rukibat, R.K.; Dalab, A.E.; Althnaian, T.A.; Al-Ramadan, S.Y. Biochemical and molecular investigation of thermal manipulation protocols during broiler embryogenesis and subsequent thermal challenge. BMC Vet. Res. 2015, 11, 292–300. [Google Scholar] [CrossRef]
- Al-Zghoul, M.B.; Dalab, A.E.; Yahya, I.E.; Althnaian, T.A.; Al-Ramadan, S.Y.; Ali, A.M.; Albokhadaim, I.F.; El-Bahr, S.M.; Al Busadah, K.A.; Hannon, K.M. Thermal manipulation during broiler chicken embryogenesis: Effect on mRNA expressions of Hsp108, Hsp70, Hsp47 and Hsf-3 during subsequent post-hatch thermal challenge. Res. Vet. Sci. 2015, 103, 211–217. [Google Scholar] [CrossRef]
- Al-Zghoul, M.B.; Dalab, A.E.; Ababneh, M.M.; Jawasreh, K.I.; Al Busadah, K.A.; Ismail, Z.B. Thermal manipulation during chicken embryogenesis results in enhanced Hsp70 gene expression and the acquisition of thermotolerance. Res. Vet. Sci. 2013, 95, 502–507. [Google Scholar] [CrossRef]
- de Barros Moreira Filho, A.L.; de Oliveira, C.J.B.; de Oliveira, H.B.; Campos, D.B.; Guerra, R.R.; Costa, F.G.P.; Givisiez, P.E.N. High incubation temperature and threonine dietary level improve ileum response against post-hatch salmonella enteritidis inoculation in broiler chicks. PloS one 2015, 10, e0131474. [Google Scholar] [CrossRef]
- Rajkumar, U.; Shanmugam, M.; Rajaravindra, K.S.; Vinoth, A.; RAO, S.R. Effect of increased incubation temperature on juvenile growth, immune and serum biochemical parameters in selected chicken populations. Indian J. Anim. Sci. 2015, 85, 1328–1333. [Google Scholar]
- Liu, J.; Yan, X.; Li, Q.; Wang, G.; Liu, H.; Wang, J.; Li, L.; Du, X.; Han, C.; He, H. Thermal manipulation during the middle incubation stage has a repressive effect on the immune organ development of Peking ducklings. J. Therm. Biol. 2013, 38, 520–523. [Google Scholar] [CrossRef]
- Shanmugasundaram, R.; Wick, M.; Lilburn, M.S. Effect of embryonic thermal manipulation on heat shock protein 70 expression and immune system development in Pekin duck embryos. Poult. Sci. 2018. [Google Scholar] [CrossRef]
- Shanmugasundaram, R.; Wick, M.; Lilburn, M. Effect of embryonic thermal manipulation on heat shock protein 70 (HSP70) expression and subsequent immune response to post-hatch lipopolysaccharide challenge in Pekin ducklings. Poult. Sci. 2018. [Google Scholar] [CrossRef]
- Wigley, P.; Kaiser, P. Avian cytokines in health and disease. Rev. Bras. Cienc. Avic. 2003, 5, 1–14. [Google Scholar] [CrossRef]
- Eming, S.A.; Hammerschmidt, M.; Krieg, T.; Roers, A. Interrelation of immunity and tissue repair or regeneration. Semin. Cell Dev. Biol. 2009, 20, 517–527. [Google Scholar]
- Zaboli, G.R.; Rahimi, S.; Shariatmadari, F.; Torshizi, M.A.K.; Baghbanzadeh, A.; Mehri, M. Thermal manipulation during Pre and Post-Hatch on thermotolerance of male broiler chickens exposed to chronic heat stress. Poult. Sci. 2016, 96, 478–485. [Google Scholar] [CrossRef]
- Lourens, A.; Van den Brand, H.; Meijerhof, R.; Kemp, B. Effect of eggshell temperature during incubation on embryo development, hatchability, and posthatch development. Poult. Sci. 2005, 84, 914–920. [Google Scholar] [CrossRef]
- Al-Zghoul, M.B.; Alliftawi, A.R.S.; Saleh, K.M.M.; Jaradat, Z.W. Expression of digestive enzyme and intestinal transporter genes during chronic heat stress in the thermally manipulated broiler chicken. Poultry science 2019, pez249. [Google Scholar] [CrossRef]
- Piestun, Y.; Shinder, D.; Ruzal, M.; Halevy, O.; Yahav, S. The effect of thermal manipulations during the development of the thyroid and adrenal axes on in-hatch and post-hatch thermoregulation. J. Therm. Biol. 2008, 33, 413–418. [Google Scholar] [CrossRef]
- Collin, A.; Berri, C.; Tesseraud, S.; Rodon, F.E.R.; Skiba-Cassy, S.; Crochet, S.; Duclos, M.J.; Rideau, N.; Tona, K.; Buyse, J. Effects of thermal manipulation during early and late embryogenesis on thermotolerance and breast muscle characteristics in broiler chickens. Poult. Sci. 2007, 86, 795–800. [Google Scholar] [CrossRef]
- Al-Zghoul, M.B.; Al-Natour, M.Q.; Dalab, A.S.; Alturki, O.I.; Althnaian, T.I.I.I.; Al-Ramadan, S.Y.; Hannon, K.M.; El-Bahr, S.M. Thermal Manipulation Mid-term Broiler Chicken Embryogenesis: Effect on Muscle Growth Factors and Muscle Marker Genes. Rev. Bras. Cienc. Avic. 2016, 18, 607–618. [Google Scholar] [CrossRef] [Green Version]
- Piestun, Y.; Halevy, O.; Yahav, S. Thermal manipulations of broiler embryos—The effect on thermoregulation and development during embryogenesis. Poult. Sci. 2009, 88, 2677–2688. [Google Scholar] [CrossRef]
- Hartley, R.S.; Bandman, E.; Yablonka-Reuveni, Z. Skeletal muscle satellite cells appear during late chicken embryogenesis. Dev. Biol. 1992, 153, 206–216. [Google Scholar] [CrossRef] [Green Version]
- Halevy, O.; Yahav, S.; Rozenboim, I. Enhancement of meat production by environmental manipulations in embryo and young broilers. Worlds Poult. Sci. J. 2006, 62, 485–497. [Google Scholar]
- Piestun, Y.; Yahav, S.; Halevy, O. Thermal manipulation during embryogenesis affects myoblast proliferation and skeletal muscle growth in meat-type chickens. Poult. Sci. 2015, 94, 2528–2536. [Google Scholar] [CrossRef]
- Leon, L.R. Heat stroke and cytokines. Prog. Brain Res. 2007, 162, 481–524. [Google Scholar]
- Xie, X.; Liang, C.; Li, M.; Chen, Z. Effects of Gaba on the Thymus Cytokines of Wenchang Chickens Submitted to Heat Stress. Rev. Bras. Cienc. Avic. 2017, 19, 143–150. [Google Scholar] [CrossRef] [Green Version]
- Mosmann, T.R.; Cherwinski, H.; Bond, M.W.; Giedlin, M.A.; Coffman, R.L. Two types of murine helper T cell clone. I. Definition according to profiles of lymphokine activities and secreted proteins. J. Immunol. 1986, 136, 2348–2357. [Google Scholar]
- Degen, W.G.; van Daal, N.; Rothwell, L.; Kaiser, P.; Schijns, V.E. Th1/Th2 polarization by viral and helminth infection in birds. Vet. Microbiol. 2005, 105, 163–167. [Google Scholar] [CrossRef] [Green Version]
- Eming, S.A.; Krieg, T.; Davidson, J.M. Inflammation in wound repair: Molecular and cellular mechanisms. J. Investig. Dermatol. 2007, 127, 514–525. [Google Scholar] [CrossRef]
- Streetz, K.; Luedde, T.; Manns, M.; Trautwein, C. Interleukin 6 and liver regeneration. Gut 2000, 47, 309–312. [Google Scholar] [CrossRef] [Green Version]
- Rennekampff, H.-O.; Hansbrough, J.F.; Kiessig, V.; Doré, C.; Sticherling, M.; Schröder, J.-M. Bioactive interleukin-8 is expressed in wounds and enhances wound healing. J. Surg. Res. 2000, 93, 41–54. [Google Scholar] [CrossRef]
- Bosch, I.; Xhaja, K.; Estevez, L.; Raines, G.; Melichar, H.; Warke, R.V.; Fournier, M.V.; Ennis, F.A.; Rothman, A.L. Increased production of interleukin-8 in primary human monocytes and in human epithelial and endothelial cell lines after dengue virus challenge. J. Virol. 2002, 76, 5588–5597. [Google Scholar] [CrossRef]
- Crouser, E.D.; Shao, G.; Julian, M.W.; Macre, J.E.; Shadel, G.S.; Tridandapani, S.; Huang, Q.; Wewers, M.D. Monocyte activation by necrotic cells is promoted by mitochondrial proteins and formyl peptide receptors. Crit. Care Med. 2009, 37, 2000. [Google Scholar] [CrossRef]
- Shini, S.; Kaiser, P. Effects of stress, mimicked by administration of corticosterone in drinking water, on the expression of chicken cytokine and chemokine genes in lymphocytes. Stress 2009, 12, 388–399. [Google Scholar] [CrossRef]
- Shini, S.; Shini, A.; Kaiser, P. Cytokine and chemokine gene expression profiles in heterophils from chickens treated with corticosterone. Stress 2010, 13, 185–194. [Google Scholar] [CrossRef]





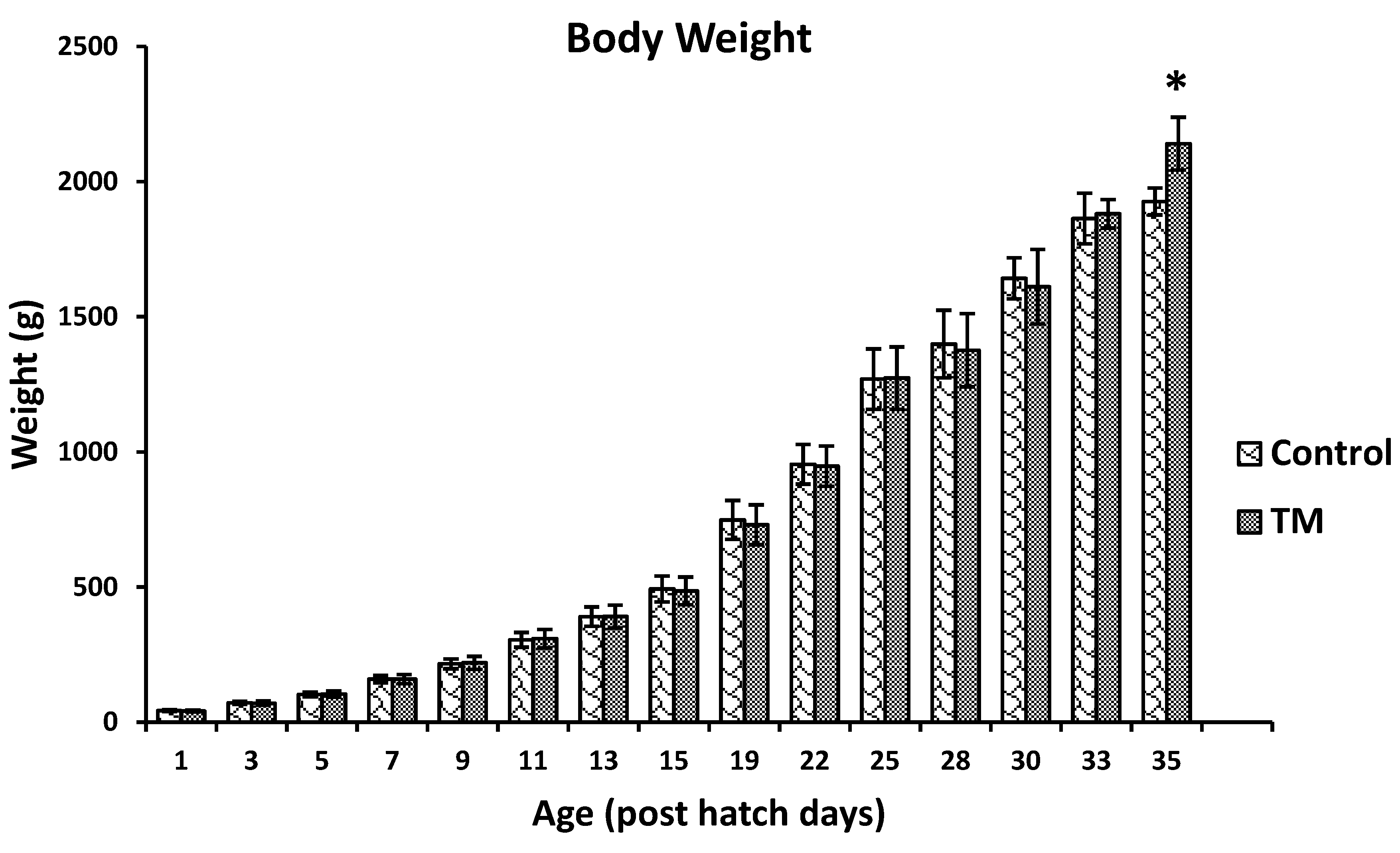{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ingredient (% As fed) | Starter Diet | Grower Diet |
|---|---|---|
| Day 1–21 | Day 22–35 | |
| Corn | 57 | 64 |
| Soybean meal | 32 | 25 |
| Concentrate | 6 | 5.5 |
| Soybean oil | 1 | 2 |
| Dicalcium phosphate | 1.1 | 0.90 |
| Limestone | 1.5 | 1.5 |
| Mineral-vitamin premix a | 0.75 | 0.50 |
| Salt (NaCl) | 0.25 | 0.25 |
| DL-Methionine | 0.23 | 0.21 |
| L-Lysine HCl | 0.17 | 0.14 |
| Calculated nutritive value (g kg−1) DM b | ||
| AMEn (kcal kg−1) c | 3120 | 3185 |
| Crude protein (DM %) | 22.3 | 19.2 |
| Ether Extract (DM %) | 4.9 | 7.8 |
| Gene | Sequence (5′-3′) |
|---|---|
| 28S rRNA | F: CCTGAATCCCGAGGTTAACTATT |
| R: GAGGTGCGGCTTATCATCTATC | |
| IL-4 | F: GAGAGCATCCGGATAGTGAATG |
| R: TGTGGAGGCTTTGCATAAGAG | |
| IL-8 | F: CTGCGGTGCCAGTGCATTA |
| R: AGCACACCTCTCTTCCATCC | |
| IL-15 | F: GTGGTCAGACGTTCTGAAAGAT |
| R: CAGGTTCCTGGCATTCTATATCC | |
| IL-16 | F: GGAACAAAGCAGCCCAGTTC |
| R: GGCTGTGGTGTGCACCTGTA | |
| IL-17 | F: CTCCGATCCCTTATTCTCCTC |
| R: AAGCGGTTGTGGTCCTCAT | |
| IL-18 | F: AGGTGAAATCTGGCAGTGGAAT |
| R: TGAAGGCGCGGTGGTTT | |
| IFN-α | F: ATGCCACCTTCTCTCACGAC |
| R: AGGCGCTGTAATCGTTGTCT | |
| IFN-β | F: CCTCAACCAGATCCAGCATTA |
| R: TAGTTGTTGTGCCGTAGGAAG | |
| IFN-γ | F: CAAGTCAAAGCCGCACATC |
| R: CGCTGGATTCTCAAGTCGTT |
| IL-4 (pg/mL) | 0 h | 1 h | 3 h | 5 h | 7 h |
|---|---|---|---|---|---|
| Control | 227.01 ± 2.81 ax | 301.19 ± 74.13 axy | 304.67 ± 8.61 by | 246.70 ± 17.83 bx | 250.30 ± 44.22 axy |
| TM | 205.21 ± 84.48 ax | 219.88 ± 23.90 ax | 243.25 ± 33.06 ax | 284.74 ± 23.06 ax | 248.69 ± 72.82 ax |
| IL-8 (pg/mL) | 0 h | 1 h | 3 h | 5 h | 7 h |
| Control | 30.35 ± 4.68 bx | 33.48 ± 5.26 bxy | 33.94 ± 8.64 bxy | 30.91 ± 6.20 bxy | 38.17 ± 7.36 ay |
| TM | 21.37 ± 4.65 ax | 26.18 ± 6.91 axz | 41.01 ± 3.55 ay | 38.72 ± 4.53 ay | 31.12 ± 9.04 az |
| IFN-γ (pg/mL) | 0 h | 1 h | 3 h | 5 h | 7 h |
| Control | 179.18 ± 48.18 bx | 155.88 ± 63.06 bx | 183.36 ± 84.59 ax | 179.39 ± 69.86 ax | 204.39 ± 41.40 bx |
| TM | 99.58 ± 46.31 ax | 93.23 ± 45.77 ax | 189.23 ± 32.46 ay | 199.92 ± 58.76 ay | 108.00 ± 52.19 ax |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Saleh, K.M.M.; Al-Zghoul, M.B. Effect of Acute Heat Stress on the mRNA Levels of Cytokines in Broiler Chickens Subjected to Embryonic Thermal Manipulation. Animals 2019, 9, 499. https://doi.org/10.3390/ani9080499
Saleh KMM, Al-Zghoul MB. Effect of Acute Heat Stress on the mRNA Levels of Cytokines in Broiler Chickens Subjected to Embryonic Thermal Manipulation. Animals. 2019; 9(8):499. https://doi.org/10.3390/ani9080499
Chicago/Turabian StyleSaleh, Khaled M. M., and Mohammad B. Al-Zghoul. 2019. "Effect of Acute Heat Stress on the mRNA Levels of Cytokines in Broiler Chickens Subjected to Embryonic Thermal Manipulation" Animals 9, no. 8: 499. https://doi.org/10.3390/ani9080499
APA StyleSaleh, K. M. M., & Al-Zghoul, M. B. (2019). Effect of Acute Heat Stress on the mRNA Levels of Cytokines in Broiler Chickens Subjected to Embryonic Thermal Manipulation. Animals, 9(8), 499. https://doi.org/10.3390/ani9080499