1. Introduction
The Triassic (Carnian–Norian, ca. 225–235 Ma) Luning Formation of west–central Nevada has been the focus of spirited debate [
1,
2,
3,
4,
5] regarding its depositional environment and paleoecology (
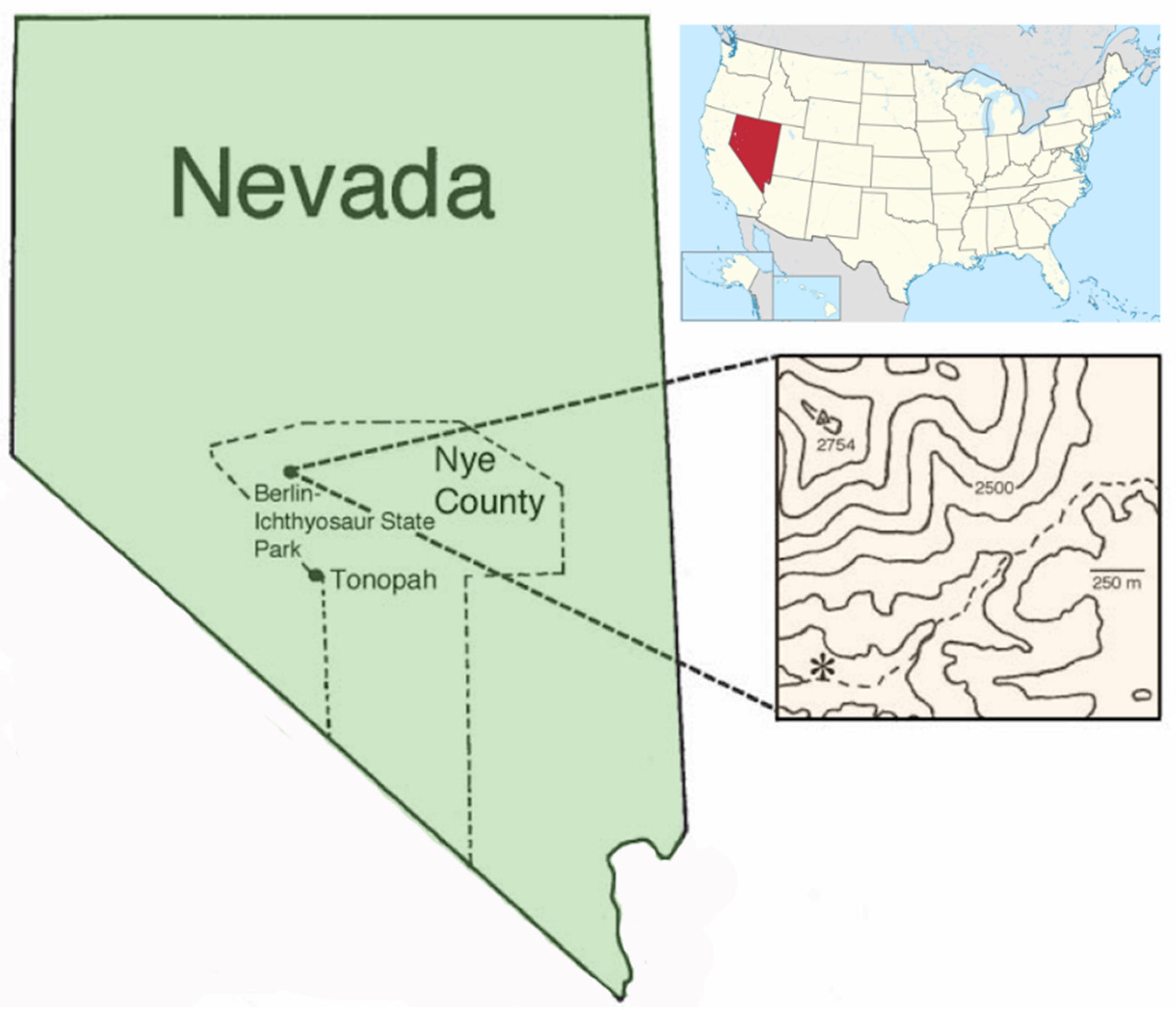
Figure 1 and
Figure 2).
Repeated attempts have been made to determine the paleobathymetry of the Shaly Limestone Member of the Luning Formation of Berlin-Ichthyosaur State Park (BISP), central Nevada, USA. Further analysis is required to help ascertain the cause of death and reasons for postmortem emplacement in virtually the same spot of seven to nine large carcasses of the ichthyosaur
Shonisaurus popularis. Camp [
6,
7] originally suggested death by stranding on a Panthalassic shoreline. Holger [
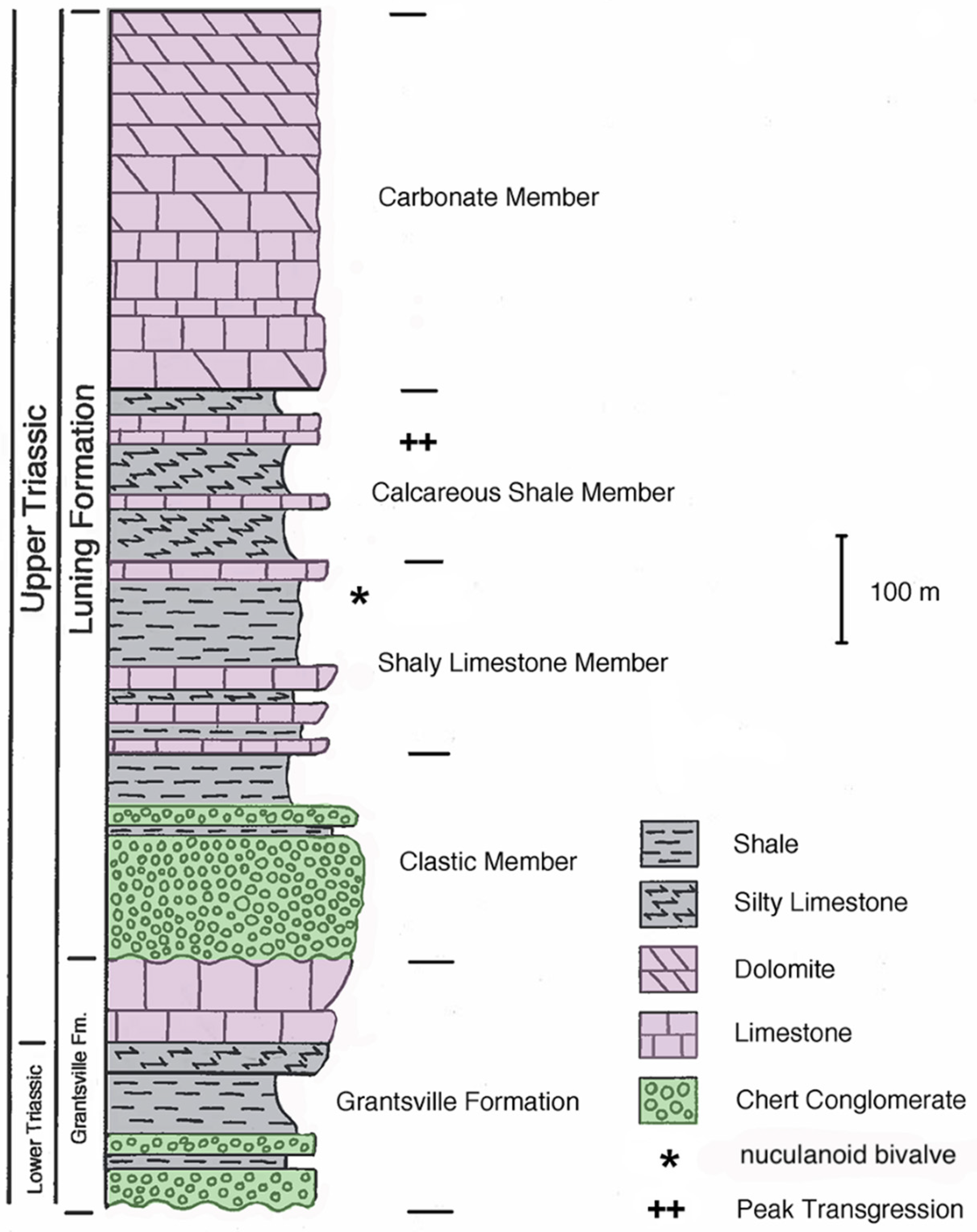
1] challenged this hypothesis by showing that the rock matrix in question was deposited in deep water, thus falsifying the ichthyosaur mass stranding hypothesis. The ichthyosaurs occur in the Shaly Limestone Member of the Luning Formation, close to the peak transgressive phase of marine sediment deposition in the region (
Figure 2).
Apart from rejecting a shoreline interpretation, Holger showed some uncertainty regarding the paleobathymetry of the Luning Formation. In 1989, Holger and Hanger [
8] argued that limestone matrix characteristics of the Luning Formation suggested “deposition in a warm, productive, carbonate shelf setting within the photic zone, subject to marked fluctuations in oxygen content and current activity” (p. 249). Subsequently, Holger [
9] argued that her Biofacies 2 of the Luning Formation was deposited in a relatively deep-water, “stagnant outer shelf” basin. Holger [
1] argued further that the Luning Formation may have been deposited at a site of major upwelling, thus providing a high productivity hunting area for the shonisaurs. Bottjer [
10] argued in favor of a deep-water interpretation, noting that “in the Late Triassic, the largest ichthyosaurs ever known roamed the proto-Pacific Ocean, feeding along the offshore margins of carbonate banks” (p. 249).
Direct fossil evidence of deep-water organisms from the Luning Formation has been difficult to acquire. In a precursor to the present study, McMenamin et al. [
2] thought that they had recovered the remains of a giant deep-sea amphipod, but the fossil turned out to be the tail of a decapod [
4] that could potentially represent a deep-water form but alas was not diagnostic for ancient water depth.
The cause of death of the shonisaurs remains unresolved, and their emplacement in a deep-water site remains a major paleontological anomaly. The marine reptile remains at BISP are too closely spaced (overlapping in some cases) to represent mere passive accumulation of skeletons in a sediment-starved basinal setting.
In a recent study of the Luning Formation at BISP, Kelley et al. [
11] argued that there was no evidence for a significant environmental perturbation that could be blamed for the deaths of the ichthyosaurs. Kelley et al. [
11] further maintained that the ichthyosaurs at BISP were deposited between fair-weather wave base and maximum storm wave base, in other words, between approximately 15 and 90 m depth. This relatively shallow depth estimate was presented as support for the hypothesis that the adult shonisaurs died as part of a philopatric group engaging in reproductive behavior [
11]. This concept was first proposed by Massare [
12], a work that is curiously not cited in Reference 11. Unfortunately, the Massare [
12] and Kelley et al. [
11] scenario provides neither a killing mechanism for the ichthyosaurs nor a method for sequentially transporting their bodies to virtually the same spot on the seafloor—a problem that is magnified if BISP indeed represents a deep-water site. As shown by the evidence presented below, the Kelley et al. [
11] depth estimate is too shallow by at least a factor of ten, making it highly unlikely that the ichthyosaurs perished as part of a group engaged in geographically localized reproductive behavior. The BISP ichthyosaurs and the fossils described here are both derived from the Shaly Limestone Member of the Luning Formation, although meter-level stratigraphic precision of either occurrence is not available at this time [
3]. This stratigraphic imprecision, however, does not invalidate the conclusions that follow.
2. Methods
Bivalves and other mollusks are common in the Shaly Limestone Member of the Luning Formation but are usually preserved as steinkerns that lack both taxonomically diagnostic surface ornamentation and detailed morphology of the hinge structure. Due to this limited mode of preservation, there are significant gaps in our knowledge of the Late Triassic bivalve, gastropod and cephalopod faunas of the Shaly Limestone Member of the Luning Formation. The clams Septocardia and Myophoria are present and sometimes abundant in the Shaly Limestone Member of the Luning Formation. These are usually not interpreted as deep-water taxa, but their depth range(s) in the Late Triassic are not known with precision, and it is possible that they represent eurybathic taxa. In contrast, the mollusks reported below represent, in comparison with their most similar living forms, animals that are considered here to not be eurybathic but rather depth-diagnostic within the range of a few hundred meters.
Mount Holyoke College paleontological field expeditions in 2013–2014 recovered limestone samples from the Shaly Limestone Member of the Luning Formation from sites near Berlin-Ichthyosaur State Park, Nevada. Acetic acid maceration of field sample 12 of 23 May 2014 produced a single specimen of a left (opisthogyrate condition) nuculanoid valve preserved by silicification (
Figure 3,
Figure 4,
Figure 5 and
Figure 6).
This is a rare mode of preservation in the Shaly Limestone Member (most of its clam fossils are preserved as steinkerns). The fossil (IGM 5010a) is taxonomically significant considering that both its valve interior and exterior, as well as its hinge teeth series, are preserved. A second acid maceration from the Luning Formation (field sample 4 of 15 May 2013) produced silicified malacostracan chelae [
4] and other fossils.
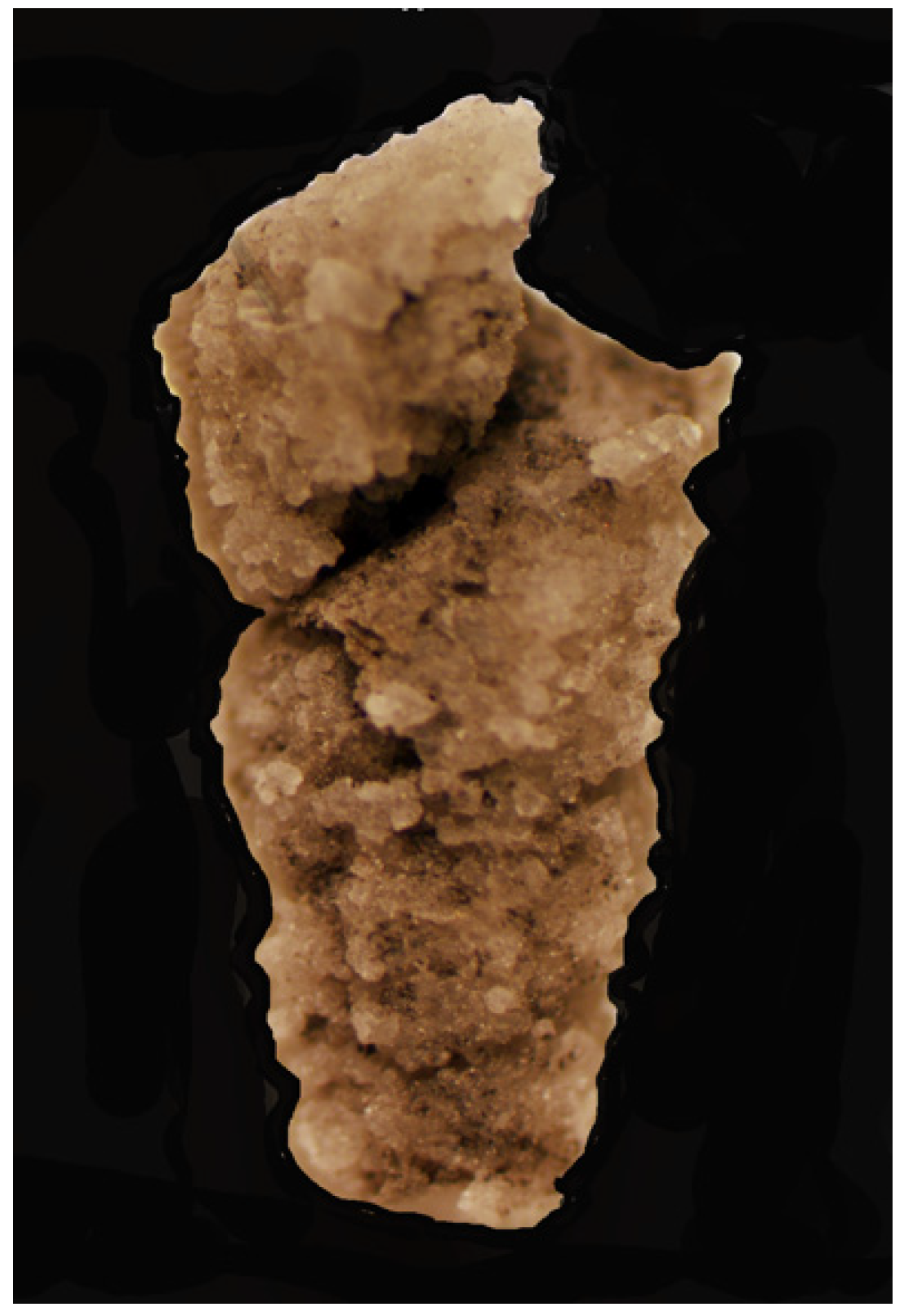
The same maceration sample (field sample 12 of 23 May 2014) that yielded the nuculanoid bivalve produced a specimen of an abyssochrysoid gastropod (IGM 5010b). This represents the first known abyssochrysoid gastropod from the Luning Formation (
Figure 7,
Figure 8,
Figure 9,
Figure 10,
Figure 11,
Figure 12 and
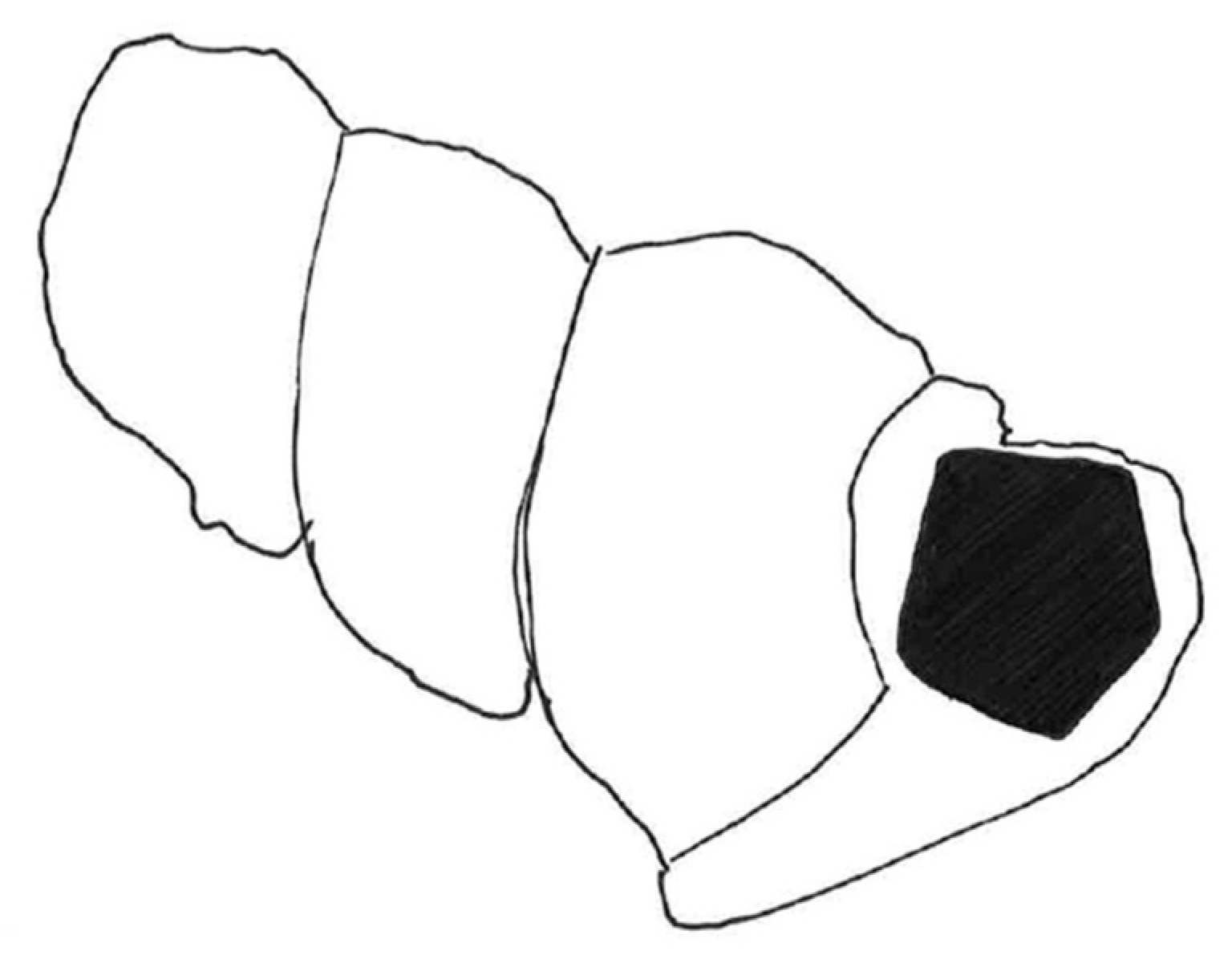
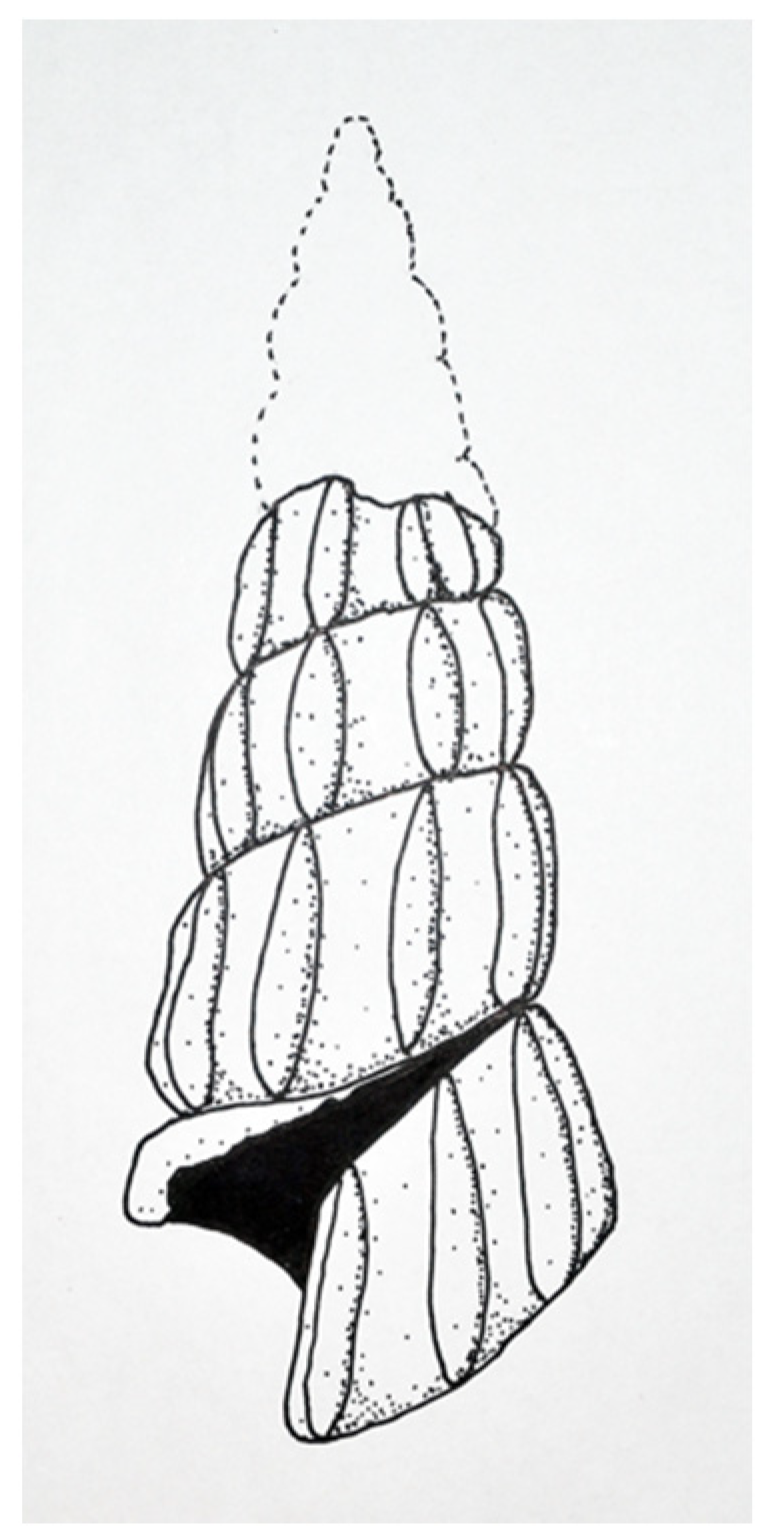
Figure 13). Gastropods are common in the Shaly Limestone Member of the Luning Formation but are preserved primarily as steinkerns that lack taxonomically useful surface ornamentation and morphology. The silicified Luning Formation gastropod is identified here as a member of the Abyssochrysoidae. This specimen shows a classic variation on the high-spired gastropod shell form, known as wentletrap, a word taken from the Dutch word wenteltrap, meaning “spiral staircase”. The specimen represents a high-spired abyssochrysoid gastropod (greatest dimension of preserved shell: 4.0 mm; greatest preserved whorl width: 1.8 mm) with varix-like axial ribs in wentletrap configuration, nine to ten per whorl. The shell’s apical angle is 13–23°. The last whorl separates slightly from the previous whorl, forming a channel. The axial ribs are relatively straight and parallel to the coiling axis. The aperture opening is slightly polygonal/pentameral.
The key character for classification of the fossil gastropod is the shape of the aperture. The aperture has a slightly polygonal/pentagonal outline (
Figure 10 and
Figure 11), similar to that seen in modern abyssochrysoids [
13], such as
Abyssochrysos melanioides, where a polygonal (square to pentagonal) aperture is seen in the specimen illustrated in Houbrick [
13] and in
Abyssochrysos melvilli, also shown in Reference [
13]. The living species most similar to the fossil snail (also with a pentagonal aperture) is
Abyssochrysos brasilianus [
14].
Abyssochrysos melanioides is also quite similar to the fossil snail; however, in
A. brasilianus, the spacing of its varix-like axial ribs near the apex is more similar to the spacing seen in the fossil.
The rock materials and associated fossils were collected under the rubric of the United States Department of the Interior, Bureau of Land Management, Paleontological Resources (Use Permit N-92224).
4. Large Triassic Cephalopod
In 2011, the author and Dianna L Schulte McMenamin proposed that the BISP Lagerstätte represented a site of sequential predation on the shonisaurs [
19]. Furthermore, they [
19] argued that the site represented a gigantic midden formed by an equally large but unknown extinct cephalopod. This hypothesis, known as “Triassic Kraken”, received a burst of media attention and trended on social media but was not well received by some parts of the paleontological community, evidently because the hypothesis appeared to make fantastic claims about a putative giant Triassic mollusk for which there was no direct fossil evidence. The controversy became so intense, involving calumnious statements, vandalism of the senior author’s Wikipedia entry and ad hominem attacks against the authors, that it complicated efforts to complete the research.
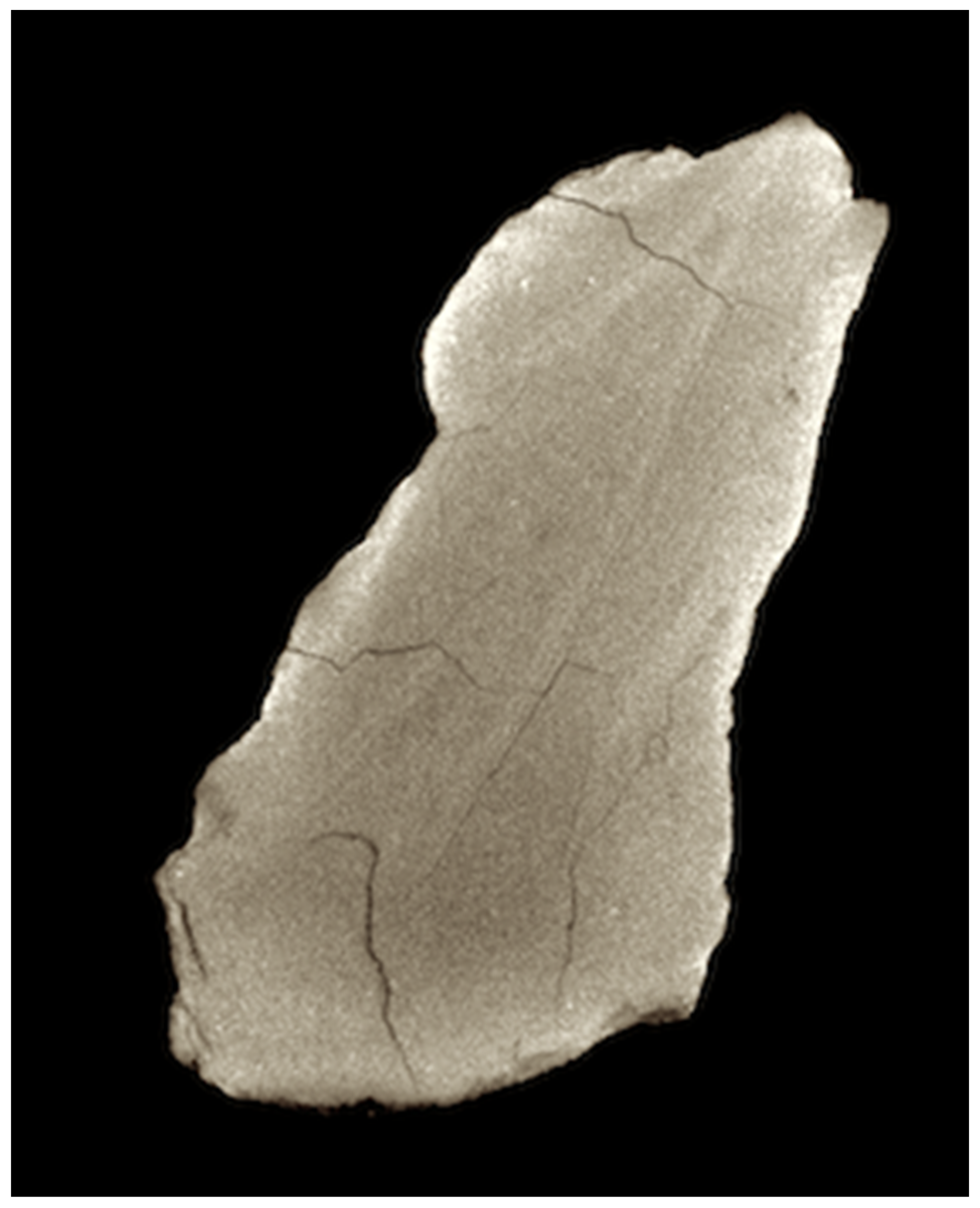
Direct fossil evidence for the giant cephalopod was presented by McMenamin and Schulte McMenamin in 2013 [
20]. The fossil from the Shaly Limestone Member of the Luning Formation is interpreted here as the rostrum [
21] of the upper beak of a cephalopod of great size (
Figure 14 and
Figure 15). The fossil is unique and unlike any other specimens collected from the Luning Formation. An alternative explanation, namely, that the specimen represents part of the hinge of a ramonalinid clam [
22], is not plausible because the specimen has a flaring V-shaped, rather than roughly parallel, fabric as would be expected for a ramonalinid hinge. Furthermore, remains of ramonalinids are otherwise unknown from the Luning Formation. Ramonalinids are a large, robust and gregarious type of myalinoid bivalve that would not be easily missed in the field.
Further evidence supporting the midden hypothesis for the BISP ichthyosaur death assemblage has since become available. McMenamin [
23,
24] presented a hydrodynamic statistical analysis showing that the vertebral centra could not have been arranged into their existing configuration by current action; some other type of agency must have been responsible for the arrangement of bones observed at the site. The probability that the biserial Specimen U arrangement of vertebral centra was produced by current action is zero [
23,
24]. Using an analogy with modern octopuses that are able to prey on both sharks and seagulls, paleoecological considerations suggest the activity of a giant cephalopod capable of preying upon
Shonisaurus [
24] individuals up to a particular size limit. Interestingly, the ichthyosaurs at BISP appear to be size-selected, and all are approximately the same size. As larger ichthyosaurs are known from the Luning Formation (in prep.), it seems reasonable to infer the prey ichthyosaurs were confined to a particular size range that the large cephalopod could subdue. An intact shonisaur vertebral column from an animal belonging to this same size class occurs outside of the front entrance to the Fossil House at BISP [
24]. This vertebral column has been stripped of its appendicular skeletal elements and thus resembles a scaled-up version of the cartilaginous remains of a modern shark vertebral column after it has been stripped of flesh by a predatory octopus. Arrangements of prey remains to form pictorial representations, part of the original Triassic Kraken proposal [
19], have now been observed among prey remains manipulated by the modern octopuses of Mo’orea, French Polynesia [
25].
The living species most similar to the Triassic Kraken is the colossal squid (
Mesonychoteuthis hamiltoni). Interestingly, the preferred depth range of adult
Mesonychoteuthis is at the mesopelagic/bathypelagic depth boundary [
26]; in other words, a depth of approximately 1000 m. Finally, in one of the best preserved Nevada shonisaurs [
24], the rib cage is crushed and the backbone broken in a peculiar fashion that once again shows that the occurrences at BISP stand in stark contrast to the typical ichthyosaur carcass fall [
27].
An estimate of the size of the Triassic Kraken may be made from the upper beak rostrum fossil shown in
Figure 14 and
Figure 15. The length of the beak rostrum fossil is 48 mm. A comparable beak rostrum section on a large specimen of the Humbolt squid (
Dosidicus gigas) has a length of 4 mm.
Dosidicus gigas can reach a 1.5 m mantle length. This gives:
In a typical squid, the mantle length (ML)/total length (TL) ratio is 0.629. We may use this ratio to estimate the total length of the Triassic Kraken as follows:
This value, 28.6 m total length, is close to the original 30 m total length estimate published in 2011 by McMenamin and Schulte McMenamin [
19].
5. Conclusions
A thick-shelled, smooth-surfaced nuculanoid (possibly neilonellid) bivalve from the Late Triassic of Luning Formation, Nevada, provides support for the deep marine interpretation of the depositional environment of the Shaly Limestone Member of the Luning Formation. As such, it supports Holger’s [
1] deep-water interpretation over both Camp’s [
6,
7] strandline interpretation and a recently published hypothesis of deposition of the Luning Formation above the storm wave base [
8]. The closest living relatives of the Triassic nuculanoid clam inhabit marine water depths from 500 to 2500+ m, with the most similar modern clam occurring at 878–933 m. The Luning Formation fossil clam is thus estimated here to have lived at approximately 1000 m water depth.
An abyssochrysoid gastropod was recovered from the same maceration sample that produced the Triassic clam. The closest living relative of the abyssochrysoid gastropod inhabits marine depths of 620–1540 m. The snail is thus also inferred here to have lived at a water depth of approximately 1000 m. The two types of fossil mollusks, nuculanoids and abyssochrysoids, are both considered to belong to relict taxa. The occurrences of the modern taxa (Chile and Brazil, respectively) are not too distant on a global scale (especially considering the former existence of the Tethyan seaway) from the fossil locality in Nevada. Thus, the modern and ancient shellfish could conceivably be very closely related as all are New World mollusks. Although likely not conspecific with comparable modern forms (considering the age disparity), both the clam and the snail may have an ancestor–descendant relationship with living forms, and thus may represent precursors to the geologically long-lived nuculanoid/provannid (abyssochrysoid) faunal assemblage [
28]. As the living forms inhabit a zone between the archibenthal transit and upper abyssal depths (the boundary separating these zones is 1000 m depth [
15]), a comparable depth of habitat is quite plausible for the Triassic clam–gastropod pair.
In a remarkable development for the paleontological sciences, the weight of evidence now favors the existence of an enormous Triassic cephalopod (“Triassic Kraken”) that lived at depth off the west coast of Triassic North America. This cephalopod was responsible for preying (using an analogy with the behavior of modern cephalopods) upon shonisaurs that ventured into its bathypelagic realm. Whether the ichthyosaurs were exhibiting group behaviors or not is beside the point; a killing mechanism for the ichthyosaurs is logically required, and attritional deaths cannot explain the characteristics of the ichthyosaur death assemblage observed at BISP. Perhaps not coincidentally, the preferred depth of the largest modern cephalopod, Mesonychoteuthis, is around 1000 m.
In conclusion, the Shady Limestone Member of the Luning Formation represents deposition offshore from a Late Triassic carbonate bank system. The bathymetric profile dropped off to deeper water fairly rapidly as one proceeded offshore, a situation with some similarities to the continental borderland basins off the west coast of modern-day California, with the exception that the latter represent a temperate siliciclastic rather a tropical carbonate depositional system.
6. Systematic Paleontology
Repository Data: IGM, Institute of Geology Museum, Departmento de Paleontología, Cuidad Universitaria, Delegacíon de Coyoacán, 04510, México.
Class: Bivalvia.
Subclass: Protobranchia.
Order: Nuculanida.
Superfamily: Nuculanoidea.
Remarks: Within the Nuculanoidea, members of the Neilonellidae have thick shells, are subovate/subtrigonal/subquadrate, are not rostrate, have siphons and have teeth with a noticeable gap under their umbos [
16]. Within the family,
Neilonella is known to be amphidetic, showing a thin external ligament groove.
Nuculanoid clam.
Figures: 3–6.
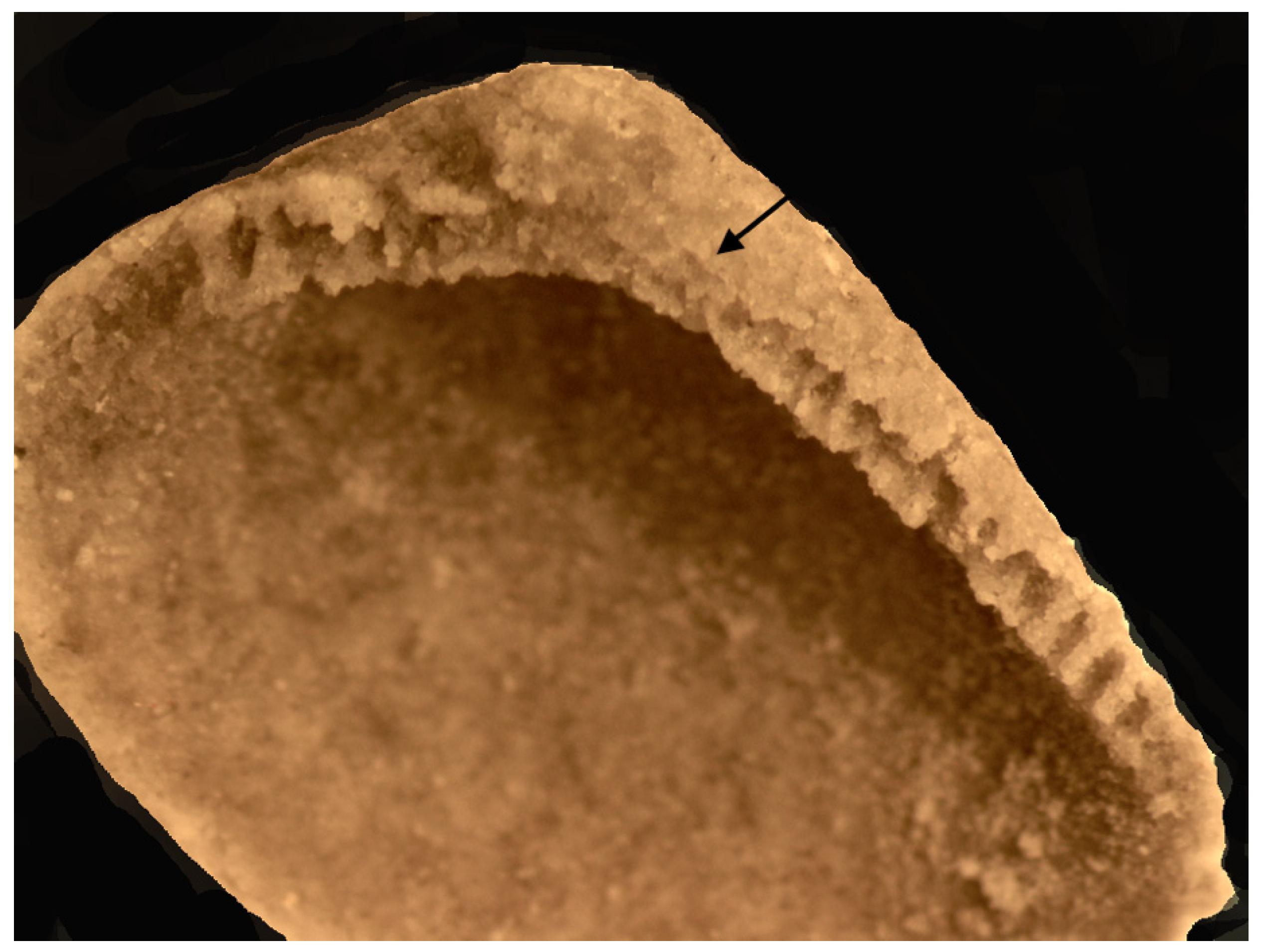
Material: One specimen, a silicified left valve (IGM 5010a), was recovered using acetic acid maceration of Field sample 12 of 23 May 2014. The specimen was collected under a permit from the United States Department of the Interior, Bureau of Land Management, Paleontological Resources Use Permit N-92224 issued to Mark McMenamin.
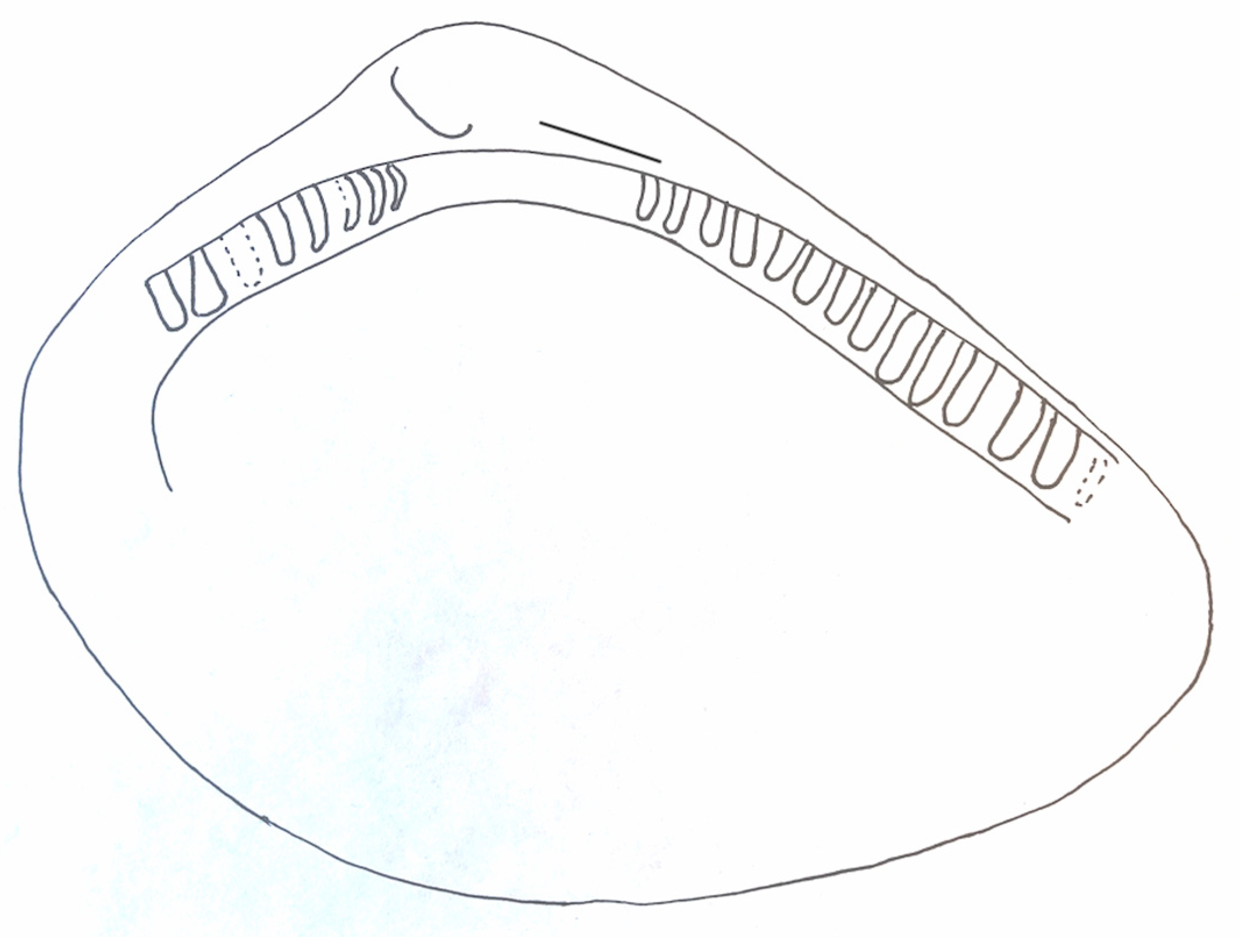
Description: A thick-shelled nuculanoid (possibly neilonellid) bivalve with an ovate outline that is slightly elongated posteriorly. The outer surface of the valve is smooth and lacking in ornamentation or growth lines. The hinge is amphidetic. A resilifer is absent. The umbo is broad and relatively large. The antero-dorsal margin is slightly concave and relatively short. The postero-dorsal margin is long and gently convex. The hinge plate is wide, with robust teeth in an anterior and posterior series (8 hinge teeth are present on the postero-dorsal margin, and 13–14 teeth are visible on the antero-dorsal margin), separated by an apparently edentulous gap. The hinge teeth at the postero-dorsal margin are generally straight but may be inclined to slightly curved. Both the antero-dorsal and postero-dorsal margins slightly overhang the hinge plate, forming a thin shelf. The adductor scars and pallial line/sinus are unknown.
Remarks: The preservation of this specimen, plus the fact that there is only a single valve currently available, precludes the establishment of a new genus and species. The Triassic fossil certainly belongs to the superfamily Nuculanoidea and could very well be a member of the Neilonellidae family due to the fact that it lacks a resilifer [
18]. A more thorough taxonomic assignment must await additional material. This isolated valve may have been subjected to postmortem transport; however, the fact that it co-occurs with an abyssochrysoid gastropod suggests that both fossils were not displaced far from their point of origin.
The hinge teeth vary in width and robustness. The specimen is amphidetic, showing a thin external ligament groove (
Figure 5 and
Figure 6). Living species (or undescribed living species) similar to the Triassic fossil are
Neilonella mexicana,
Neilonella corpulenta,
Neilonella brunnea,
Pseudoneilonella sp. A [
18] and
Malletia species A [
18].
Pseudoneilonella sp. A is the most similar to the Triassic fossil as its teeth vary in width and robustness, and it also has a very wide hinge plate. The Triassic clam differs from
Neilonella mexicana,
Neilonella corpulenta and
Neilonella brunnea as it lacks both outer surface ornamentation and chevron-shaped hinge teeth.
Locality: Field sample 12 of 23 May 2014, Late Triassic, Norian Stage (
Tropites welleri-
Stikinoceras kerri zones), Shaly Limestone Member of the Luning Formation, Nye County, Nevada; a float sample probably belonging to Biofacies 2 [
1]. This biofacies is thought to have been deposited in a relatively deep-water, “stagnant outer shelf” basin [
1]. The bivalve
Septocardia sp., a widespread and morphologically variable form, occurs in Biofacies 2 but is more abundant in Holger’s [
1] Biofacies 1.
The petrology of the Shaly Limestone Member of the Luning Formation limestone sample (12 of 23 May 2014) is as follows: The limestone is a grey, almost bluish grey, slightly muddy-looking wackestone–packstone. Acetic acid maceration of this sample, in addition to producing the nuculanoid clam, also produced silicified bivalve and gastropod fragments. In the thin section, the rock shows a fabric of weathered-looking allochem grains (peloids) and small fossils, with the fabric crossed by thin white calcite veins and occasional stylolites. Microlaminations occur in places. Patches of silicification occur in the larger shell fragments. Fossils seen in the thin section include fossil and whole bivalve valves (petrographic thin section 12 of 23 May 2014B, near thickest vein), gastropods (petrographic thin section 12 of 23 May 2014B, near crossing white veins) and rare echinoderm fragments (stereom lattice, petrographic thin section 12 of 23 May 2014A near the big “bullseye” fossil (possibly Isocrinus sp.)).
Clade: Caenogastropoda.
Superfamily: Abyssochrysoidea.
Family: Abyssochrysoidae.
Abyssochrysoid gastropod.
Figures: 7–13.
Description: A silicified, high-spired abyssochrysoid gastropod (IGM 5010b; greatest dimension of preserved shell: 4.0 mm; greatest preserved whorl width: 1.8 mm) with varix-like axial ribs in wentletrap configuration, nine to ten per whorl. The shell’s apical angle is 13–23°. The last whorl separates slightly from the previous whorl, forming a channel. The axial ribs are relatively straight and parallel to the coiling axis and do not show prominent swellings on either end of each rib. The aperture opening is slightly polygonal/pentameral.
Remarks: The living species most similar to the fossil snail and sharing its slightly polygonal aperture and varix-like axial rib spacing is
Abyssochrysos brasilianus [
14]. The polygonal aperture in the Nevada Triassic fossil snail forms, at the corner of the polygonal generating curve closest to the coiling axis, a 112° angle (
Figure 10 and
Figure 11); this is quite different from the gentle sigmoidal curve in the same position as seen in the Mesozoic high-spired procerithiid snail
Protofusus.
Abyssochrysos melanioides is also quite similar to the Nevada Triassic fossil snail; however, in
A. brasilianus, the spacing of its axial ribs near the apex is more similar to the spacing seen in the fossil. Abyssochrysoids are considered a relict group of gastropods [
13].
Locality: As for previous specimen.
Class: Cephalopoda.
Giant cephalopod (“Triassic Kraken”).
Figures: 14 and 15.
Description: Permineralized rostral end of upper beak, black to dark gray in color, of an extinct cephalopod of unusually large proportions. The specimen consists of layers of permineralized organic material tightly packed to form a lamellate cone that is roughly triangular in cross-section. This structure expands proximally to form an opening V structure (broken where it connected to the rest of the beak), with layers converging distally to form the tip of the beak. Micro-CT scan data confirm that the layers meet to form an acute rostral tip (
Figure 15). Longitudinal ridges occur on the outer surfaces. Converging striae with offset sections and spall fractures are visible on the surface of the specimen.
Remarks: The longitudinal ridges on this specimen are interpreted here as taphonomic features associated with slight contraction of the organic matter (not biomineralized) of the beak during permineralization. The converging striae with offset sections and spall fractures are similar to those seen in beak specimens taken from the giant morph of the Humbolt squid (Dosidicus gigas). Dosidicus, however, lacks the longitudinal ridges as seen in the fossil.





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}














