Living Dendrolitic Microbial Mats in Hamelin Pool, Shark Bay, Western Australia

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
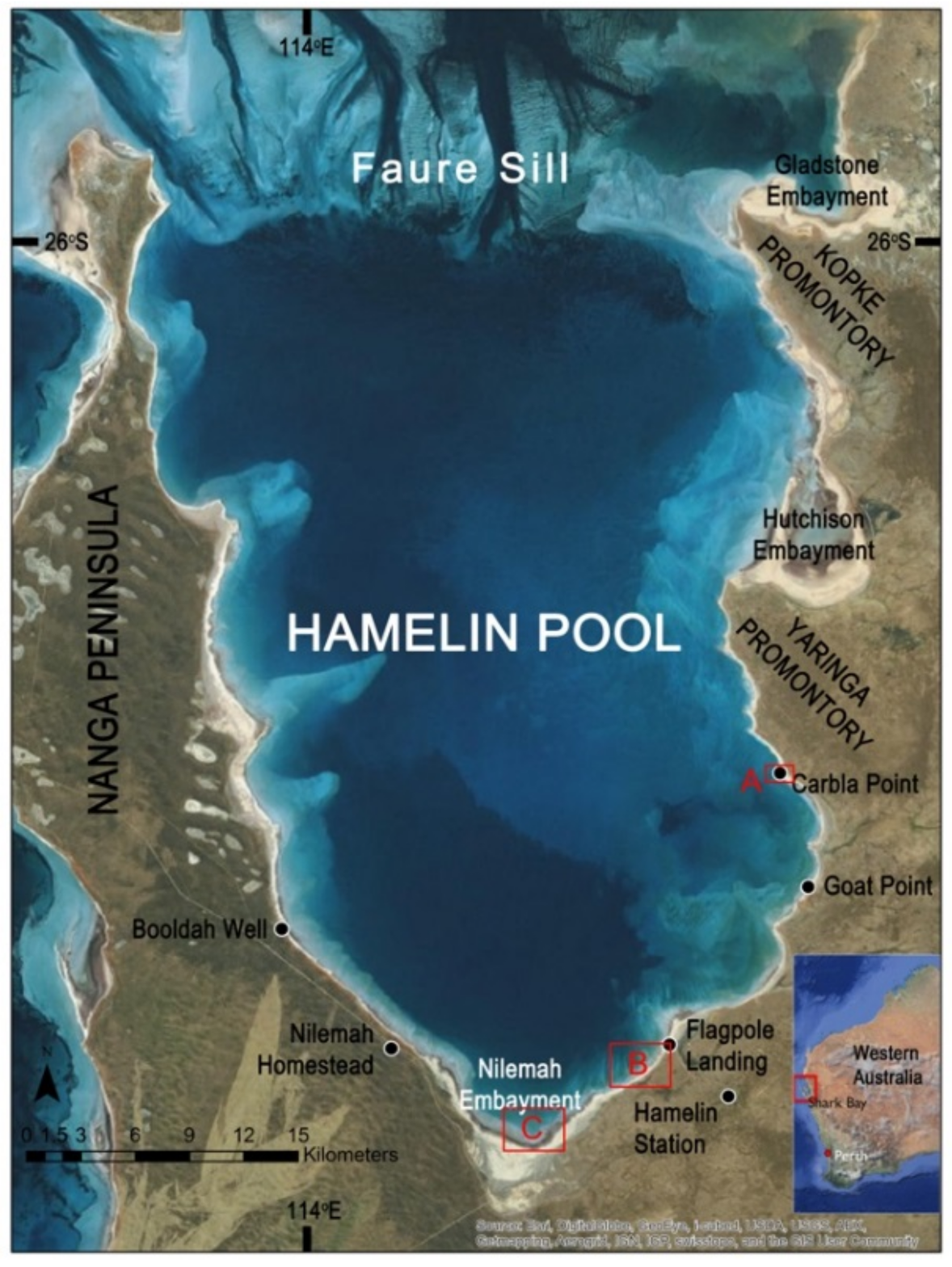
2.1. Location
2.2. Field Studies, Sampling, and Analysis
3. Results
3.1. Distribution
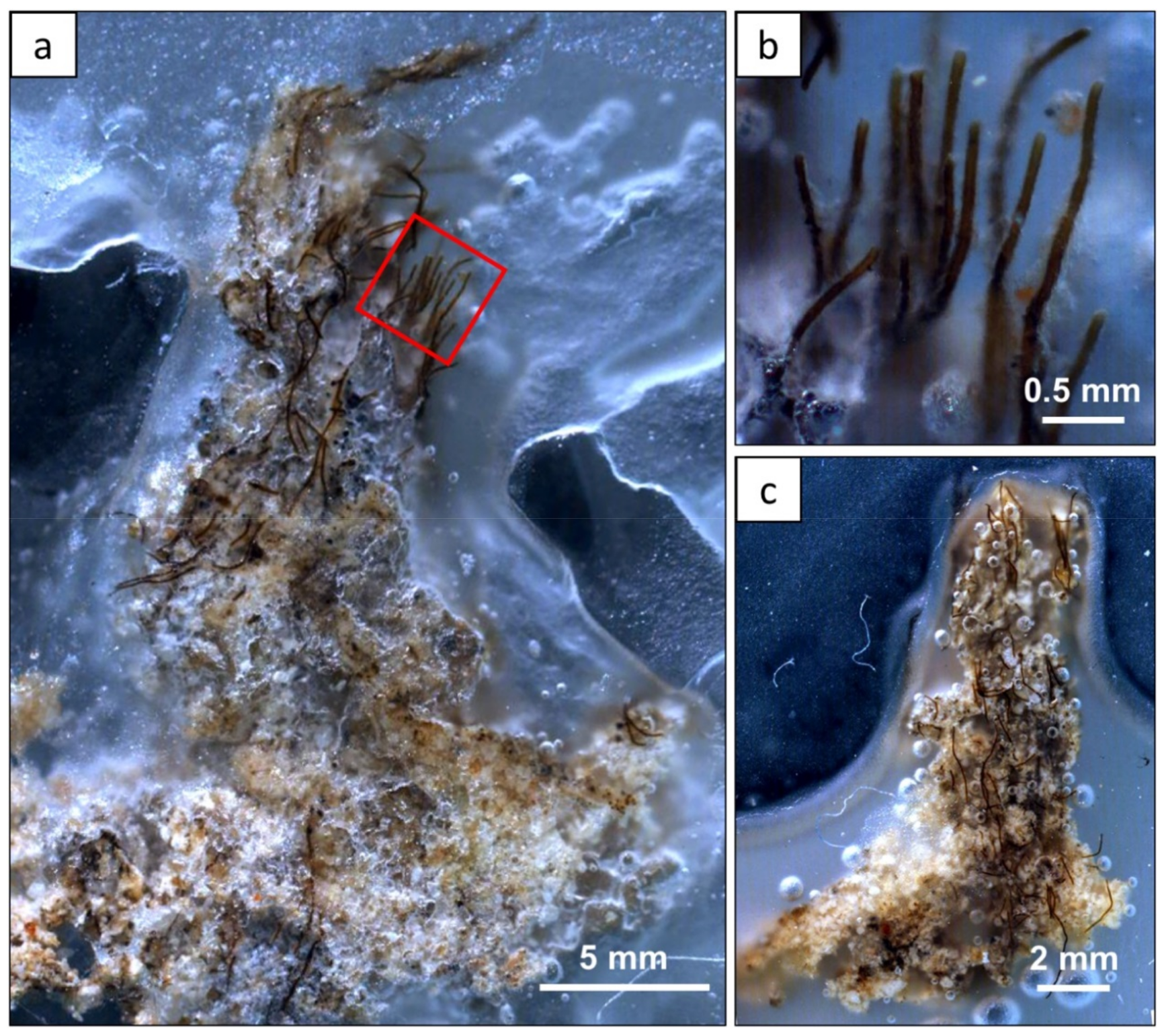
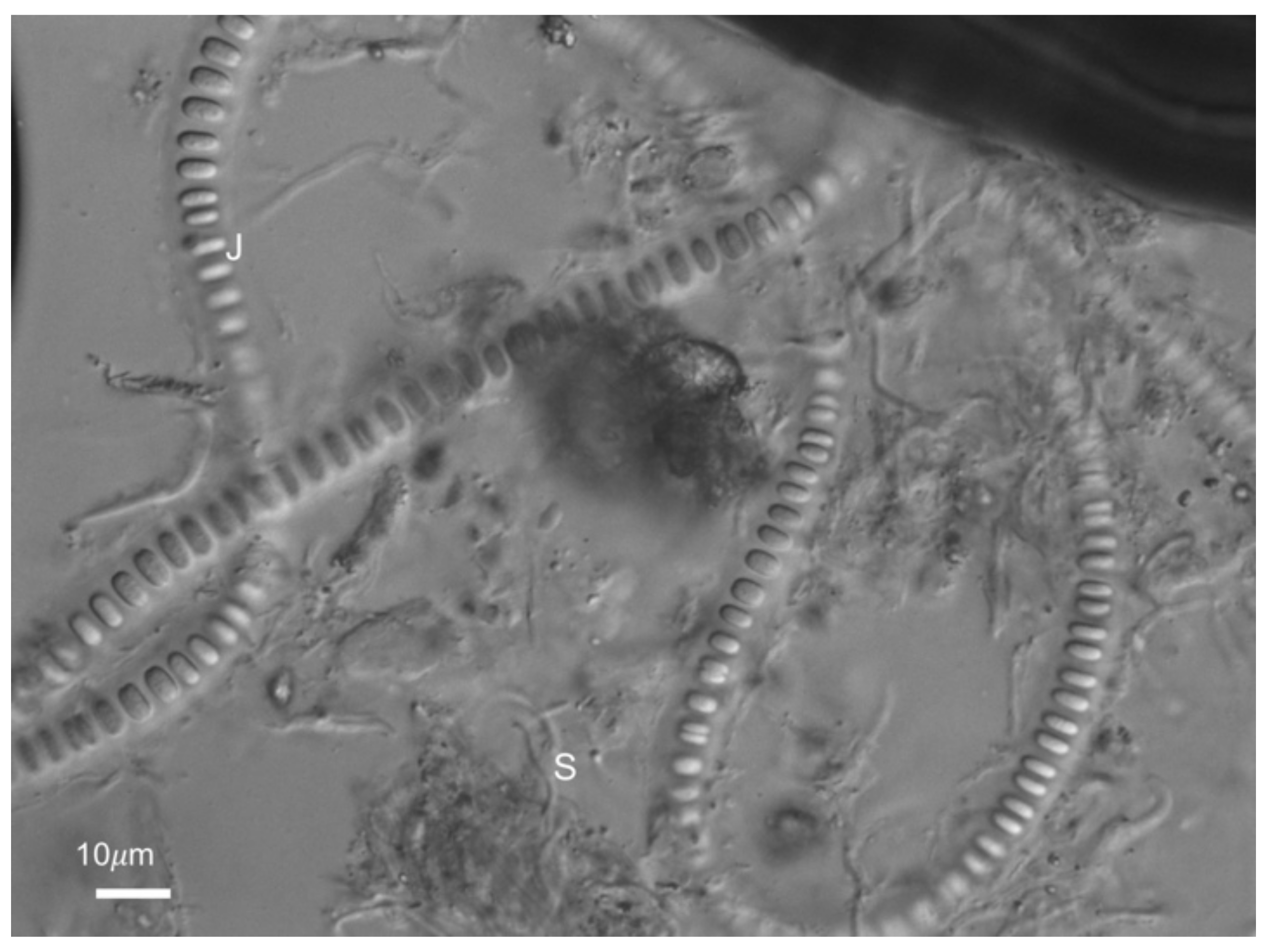
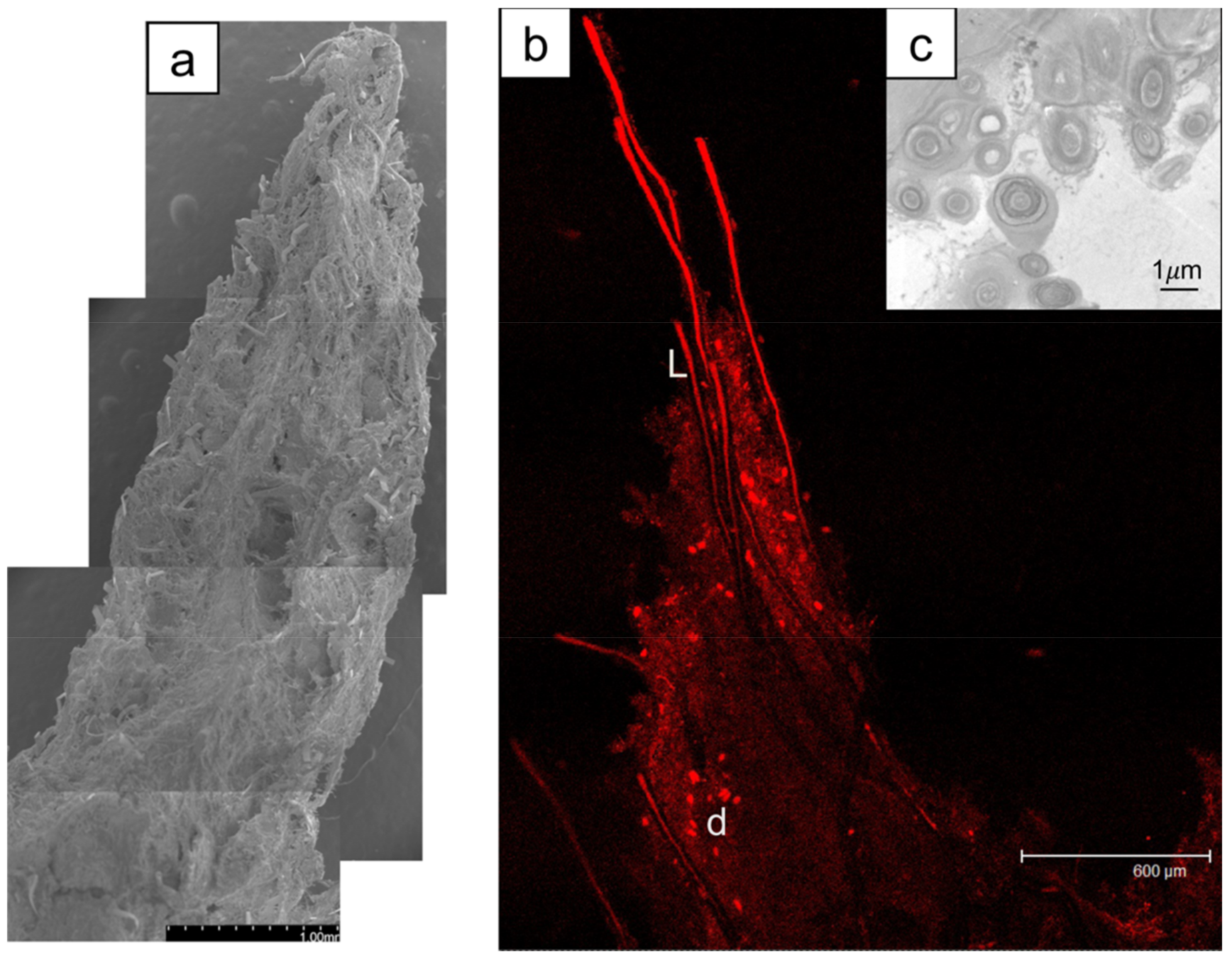
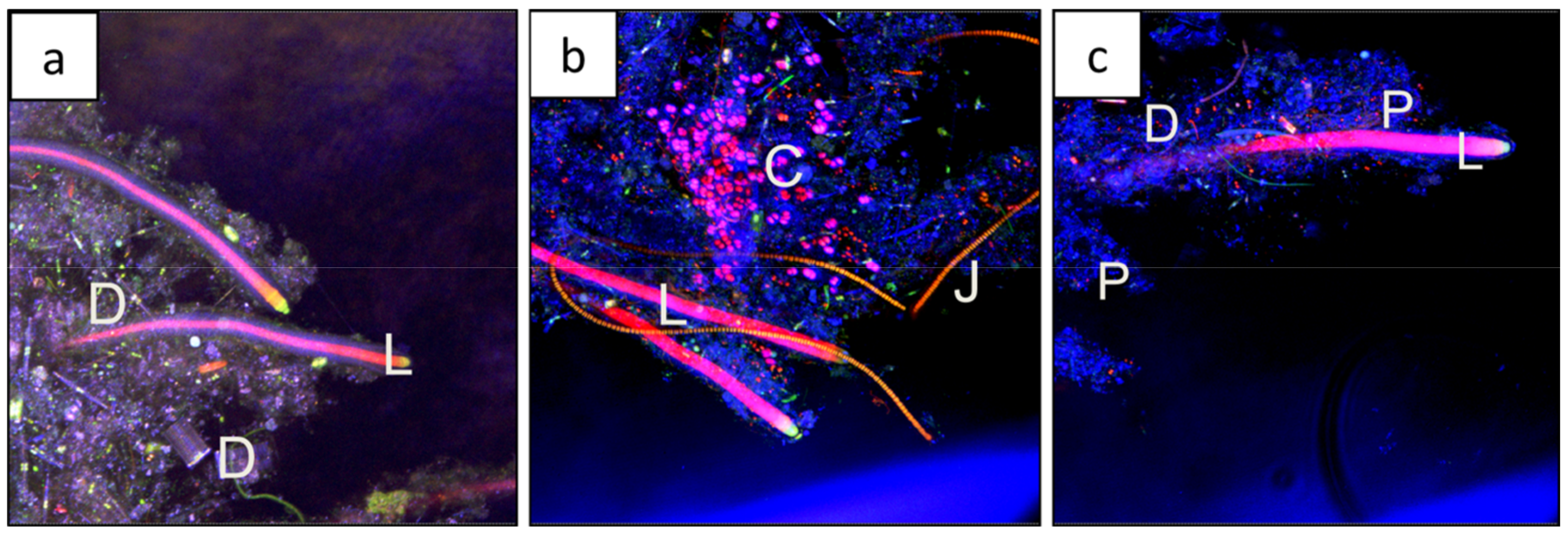
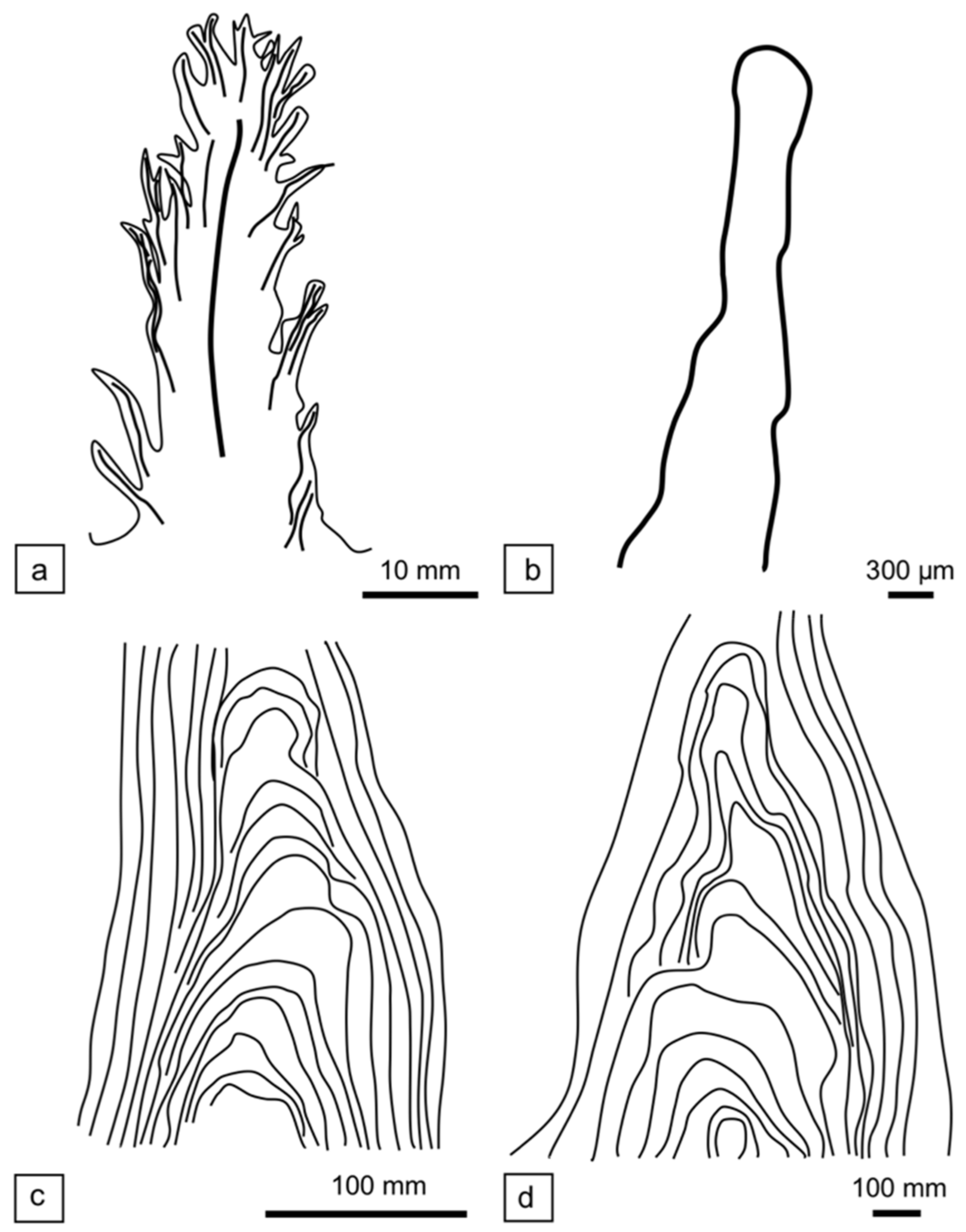
3.2. Morphology and Microbial Composition
4. Discussion
4.1. Shrub Formation and Comparison to Pinnacle and Tufted Microbial Mats
4.2. Modularity
5. Implications and Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Knoll, A.H. The early evolution of eukaryotes—A geological perspective. Science 1992, 256, 622–627. [Google Scholar] [CrossRef] [PubMed]
- Burne, R.V.; Moore, L.S. Microbialites: Organosedimentary deposits of benthic microbial communities. Palaios 1997, 2, 241–254. [Google Scholar] [CrossRef]
- Riding, R. Microbialites, stromatolites, and thrombolites. In Encyclopedia of Geobiology; Springer: Dordrecht, The Netherlands, 2011; pp. 635–654. [Google Scholar]
- Semikhatov, M.A.; Gebelein, C.D.; Cloud, P.; Awramik, S.M.; Benmore, W.C. Stromatolite morphogenesis—Progress and problems. Can. J. Earth Sci. 1979, 16, 992–1015. [Google Scholar] [CrossRef]
- Ginsburg, R.N. Controversies about stromatolites: Vices and virtues. In Controversies in Modern Geology; Academic Press: London, UK, 1991; pp. 25–36. [Google Scholar]
- Awramik, S.M. The history and significance of stromatolites. In Early Organic Evolution; Schidlowski, M., Golubic, S., Kimberley, M.M., McKirdy, D.M., Trudinger, P.A., Eds.; Springer: Berlin, Germany, 1992; pp. 435–449. [Google Scholar]
- Grotzinger, J.P.; Knoll, A.H. Stromatolites in Precambrian carbonates: Evolutionary mileposts or environmental dipsticks? Annu. Rev. Earth Planet. Sci. 1999, 27, 313–358. [Google Scholar] [CrossRef] [PubMed]
- Brasier, M.; McLoughlin, N.; Green, O.; Wacey, D. A fresh look at the fossil evidence for early Archaean cellular life. Philos. Trans. R. Soc. B 2006, 361, 887–902. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McLoughlin, N.; Wilson, L.A.; Brasier, M.D. Growth of synthetic stromatolites and wrinkle structures in the absence of microbes–implications for the early fossil record. Geobiology 2008, 6, 95–105. [Google Scholar] [CrossRef] [PubMed]
- Shepard, R.N.; Sumner, D.Y. Undirected motility of filamentous cyanobacteria produces reticulate mats. Geobiology 2010, 8, 179–190. [Google Scholar] [CrossRef] [PubMed]
- Bosak, T.; Knoll, A.H.; Petroff, A.P. The meaning of stromatolites. Annu. Rev. Earth Planet. Sci. 2013, 41, 21–44. [Google Scholar] [CrossRef]
- Ibarra, Y.; Corsetti, F.A. Lateral comparative investigation of stromatolites: Astrobiological implications and assessment of scales of control. Astrobiology 2016, 16, 271–281. [Google Scholar] [CrossRef] [PubMed]
- Andres, M.S.; Reid, R.P. Growth morphologies of modern marine stromatolites: A case study from Highborne Cay, Bahamas. Sediment. Geol. 2006, 185, 319–328. [Google Scholar] [CrossRef]
- Awramik, S.M.; Semikhatov, M.A. The relationship between morphology, microstructure, and microbiota in three vertically intergrading stromatolites from the Gunflint Iron Formation. Can. J. Earth Sci. 1979, 16, 484–495. [Google Scholar] [CrossRef]
- Grotzinger, J.P.; Rothman, D.H. An abiotic model for stromatolite morphogenesis. Nature 1996, 383, 423–425. [Google Scholar] [CrossRef]
- Kah, L.C.; Knoll, A.H. Microbenthic distribution of Proterozoic tidal flats: Environmental and taphonomic considerations. Geology 1996, 24, 79–82. [Google Scholar] [CrossRef]
- Knoll, A.H.; Swett, K.; Burkhardt, E. Paleoenvironmental distribution of microfossils and stromatolites in the Upper Proterozoic Backlundtoppen Formation, Spitsbergen. J. Paleontol. 1989, 63, 129–145. [Google Scholar] [PubMed]
- Turner, E.C.; James, N.P.; Narbonne, G.M. Taphonomic control on microstructure in early Neoproterozoic reefal stromatolites and thrombolites. Palaios 2000, 15, 87–111. [Google Scholar] [CrossRef]
- McLoughlin, N.; Melezhik, V.A.; Brasier, A.T.; Medvedev, P.V. Palaeoproterozoic stromatolites from the Lomagundi-Jatuli interval of the Fennoscandian Shield. In Reading the Archive of Earth’s Oxygenation: Volume 3: Global Events and the Fennoscandian Arctic Russia—Drilling Early Earth Project; Melezhik, V., Prave, A.R., Hanski, E.J., Fallick, A.E., Lepland, A., Kump, L.R., Strauss, H., Eds.; Frontiers in Earth Sciences; Springer: Berlin/Heidelberg, Germany, 2013; pp. 1298–1351. [Google Scholar]
- Walter, M.R.; Bauld, J.; Brock, T.D. Microbiology and morphogenesis of columnar stromatolites (Conophyton, Vacerrila) from hot springs in Yellowstone National Park. In Stromatolites; Walter, M.R., Ed.; Elsevier: New York, NY, USA, 1976; pp. 273–310. [Google Scholar]
- Flannery, D.T.; Walter, M.R. Archean tufted microbial mats and the Great Oxidation Event: New insights into an ancient problem. Aust. J. Earth Sci. 2012, 59, 1–11. [Google Scholar] [CrossRef]
- Bartley, J.K.; Kah, L.C.; Frank, T.D.; Lyons, T.W. Deep-water microbialites of the Mesoproterozoic Dismal Lakes Group: Microbial growth, lithification, and implications for coniform stromatolites. Geobiology 2015, 13, 15–32. [Google Scholar] [CrossRef] [PubMed]
- Golubic, S.; Hofmann, H.J. Comparison of Holocene and mid-Precambrian Entophysalidaceae (Cyanophyta) in stromatolitic algal mats: Cell division and degradation. J. Paleontol. 1976, 50, 1074–1082. [Google Scholar]
- Ben-Jacob, E.; Levine, H. Self-engineering capabilities of bacteria. J. R. Soc. Interface 2006, 3, 197–214. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Howell, J.; Woo, J.; Chough, S.K. Dendroid morphology and growth patterns: 3-D computed tomographic reconstruction. Palaeogeogr. Palaeocl. 2011, 299, 335–347. [Google Scholar] [CrossRef]
- Suosaari, E.P.; Reid, R.P.; Playford, P.E.; Foster, J.S.; Stolz, J.F.; Casaburi, G.; Hagan, P.D.; Chirayath, V.; Macintyre, I.G.; Planavsky, N.J.; et al. New multi-scale perspectives on the stromatolites of Shark Bay, Western Australia. Sci. Rep. 2016, 20557. [Google Scholar] [CrossRef] [PubMed]
- Hofmann, H.J. Precambrian Remains in Canada: Fossils, Dubiofossils, and Pseudofossils. In Proceedings of the 24th International Geological Congress; International Geological Congress: Montreal, QC, Canada, 1972; pp. 20–30. Available online: https://openlibrary.org/publishers/24th_International_Geological_Congress (accessed on 10 June 2018).
- Awramik, S.M.; Grey, K. Stromatolites: Biogenicity, biosignatures, and bioconfusion. Astrobiol. Planet. Missions 2005, 5906, 59060P. [Google Scholar] [CrossRef]
- Riding, R. (Ed.) Classification of microbial carbonates. In Calcareous Algae and Stromatolites; Springer: Berlin, Germany, 1991; pp. 21–51. [Google Scholar]
- Riding, R. Microbial carbonates: The geological record of calcified bacterial-algal mats and biofilms. Sedimentology 2000, 47, 179–214. [Google Scholar] [CrossRef]
- Cloud, P.E.; Semikhatov, M.A. Proterozoic stromatolite zonation. Am. J. Sci. 1969, 267, 1017–1061. [Google Scholar] [CrossRef]
- Semikhatov, M.A. Experience in stromatolite studies in the USSR. Dev. Sediment. 1976, 20, 337–357. [Google Scholar]
- Zhu, S. An outline of studies on the Precambrian stromatolites of China. Precambrian Res. 1982, 18, 367–396. [Google Scholar]
- Semikhatov, M.A.; Raaben, M.E. Proterozoic stromatolite taxonomy and biostratigraphy. In Microbial Sediments; Awramik, S.M., Riding, R., Eds.; Springer: Berlin, Germany, 2000; pp. 295–306. [Google Scholar]
- Grey, K.; Hill, A.C.; Calver, C. Biostratigraphy and stratigraphic subdivision of Cryogenian successions of Australia in a global context. Geo. Soc. Mem. 2011, 36, 113–134. [Google Scholar] [CrossRef]
- Zaitseva, T.S.; Semikhatov, M.A.; Gorokhov, I.M.; Sergeev, V.N.; Kuznetsov, A.B.; Ivanovskaya, T.A.; Melnikov, N.N.; Konstantinova, G.V. Isotopic geochronology and biostratigraphy of Riphean deposits of the Anabar Massif, North Siberia. Stratigr. Geol. Correl. 2016, 24, 549–574. [Google Scholar] [CrossRef]
- Shapiro, R.S.; Awramik, S.M. Microbialite morphostratigraphy as a tool for correlating Late Cambrian–Early Ordovician sequences. J. Geol. 2000, 108, 171–180. [Google Scholar] [CrossRef] [PubMed]
- Logan, B.W.; Hoffman, P.; Gebelein, C.D. Algal mats, cryptalgal fabrics, and structures, Hamelin Pool, Western Australia. In Evolution and Diagenesis of Quaternary Carbonate Sequences, Shark Bay, Western Australia; Logan, B.W., Read, J.F., Hagan, G.M., Hoffman, P., Brown, R.G., Woods, P.J., Gebelein, C.D., Eds.; American Association of Petroleum Geologists: Tulsa, OK, USA, 1974; Volume 22, pp. 140–193. [Google Scholar]
- Horodyski, R.J.; Bloeser, B.; Haar, S.V. Laminated algal mats from a coastal lagoon, Laguna Mormona, Baja California, Mexico. J. Sediment. Res. 1977, 47, 680–696. [Google Scholar]
- Grey, K.; Moore, L.; Burne, R.; Pierson, B.; Bauld, J. Lake Thetis, Western Australia: An example of saline lake sedimentation dominated by benthic microbial processes. Mar. Freshw. Res. 1990, 41, 275–300. [Google Scholar] [CrossRef]
- Ferris, F.G.; Thompson, J.B.; Beveridge, T.J. Modern freshwater microbialites from Kelly Lake, British Columbia, Canada. Palaios 1997, 12, 213–219. [Google Scholar] [CrossRef]
- Reid, R.P.; Visscher, P.T.; Decho, A.W.; Stolz, J.F. The role of microbes in accretion, lamination and early lithification of modern marine stromatolites. Nature 2000, 406, 989–992. [Google Scholar] [CrossRef] [PubMed]
- Gischler, E.; Gibson, M.A.; Oschmann, W. Giant holocene freshwater microbialites, Laguna Bacalar, Quintana Roo, Mexico. Sedimentology 2008, 55, 1293–1309. [Google Scholar] [CrossRef]
- Couradeau, E.; Benzerara, K.; Moreira, D.; Gérard, E.; Kaźmierczak, J.; Tavera, R.; López-García, P. Prokaryotic and eukaryotic community structure in field and cultured microbialites from the alkaline Lake Alchichica (Mexico). PLoS ONE 2011, 6, e28767. [Google Scholar] [CrossRef] [PubMed]
- Rowland, S.M.; Shapiro, R.S. Reef Patterns and Environmental Influences in the Cambrian and Earliest Ordovician; SEPM: Tulsa, OK, USA, 2002; Volume 72, pp. 95–128. [Google Scholar]
- Ibarra, Y.; Corsetti, F.A.; Cheetham, M.I.; Feakins, S.J. Were fossil spring-associated carbonates near Zaca Lake, Santa Barbara, California deposited under an ambient or thermal regime? Sediment. Geol. 2014, 301, 15–25. [Google Scholar] [CrossRef]
- Shapiro, R.S.; Rigby, J.K. First occurrence of an in situ anthaspidellid sponge in a dendrolite mound (Upper Cambrian; Great Basin, USA). J. Paleontol. 2004, 78, 645–650. [Google Scholar] [CrossRef]
- Grey, K. (Geological Survey of Western Australia, Perth, Western Australia, Australia); Awramik, S.M. (University of Santa Barbara, Santa Barbara, California, USA); Unpublished data from Stromatolite Handbook, 2017.
- Cloud, P.; Wright, L.A.; Williams, E.G.; Diehl, P.; Walter, M.R. Giant stromatolites and associated vertical tubes from the Upper Proterozoic Noonday Dolomite, Death Valley region, eastern California. Geol. Soc. Am. Bull. 1974, 85, 1869–1882. [Google Scholar] [CrossRef]
- Corsetti, F.A.; Grotzinger, J.P. Origin and significance of tube structures in Neoproterozoic post-glacial cap carbonates: Example from Noonday Dolomite, Death Valley, United States. Palaios 2005, 20, 348–362. [Google Scholar] [CrossRef]
- Folk, R.L.; Chafetz, H.S. Pisoliths (Pisoids) in Quaternary Travertines of Tivoli, Italy. In Coated Grains; Springer: Berlin/Heidelberg, Germany, 1983; p. 474. [Google Scholar]
- Chafetz, H.S.; Guidry, S.A. Bacterial shrubs, crystal shrubs, and ray-crystal shrubs: Bacterial vs. abiotic precipitation. Sediment. Geol. 1999, 126, 57–74. [Google Scholar] [CrossRef]
- Gandin, A.; Capezzuoli, E. Travertine: Distinctive depositional fabrics of carbonates from thermal spring systems. Sedimentology 2014, 61, 264–290. [Google Scholar] [CrossRef]
- Bradley, J.A.; Daille, L.K.; Trivedi, C.B.; Bojanowski, C.L.; Stamps, B.W.; Stevenson, B.S.; Nunn, H.S.; Johnson, H.A.; Loyd, S.J.; Berelson, W.M.; et al. Carbonate-rich dendrolitic cones: Insights into a modern analog for incipient microbialite formation, Little Hot Creek, Long Valley Caldera, California. NPJ Biofilms Microb. 2017, 3, 32. [Google Scholar] [CrossRef] [PubMed]
- Playford, P.E.; Cockbain, A.E.; Berry, P.F.; Roberts, A.P.; Haines, P.W.; Brooke, B.P. The Geology of Shark Bay. Geol. Surv. Western Aust. Bull. 2013, 146, 281. [Google Scholar]
- Logan, B.W.; Cebulski, D.E. Sedimentary environments of Shark Bay, Western Australia. In Carbonate Sedimentation and Environments, Shark Bay, Western Australia; Logan, B.W., Davies, G.R., Read, J.F., Cebulski, D.E., Eds.; American Association of Petrolium Geologists: Tulsa, OK, USA, 1970; Volume 13, pp. 1–37. [Google Scholar]
- Suosaari, E.P.; Reid, R.P.; Abreu Araujo, T.A.; Playford, P.E.; Holley, D.K.; McNamara, J.K.; Eberli, G.P. Environmental pressures influencing living stromatolites in Hamelin Pool, Shark Bay, Western Australia. Palaios 2016, 31, 483–496. [Google Scholar] [CrossRef]
- Burne, R.V.; Johnson, K. Sea level variation and the zonation of microbialites in Hamelin Pool, Shark Bay, Western Australia. Mar. Freshw. Res. 2012, 63, 994–1004. [Google Scholar] [CrossRef]
- Stolz, J.F.; Reid, R.P.; Visscher, P.T.; Decho, A.W.; Norman, R.S.; Aspden, R.J.; Bowlin, E.M.; Franks, J.; Foster, J.S.; Paterson, D.M.; et al. The microbial communities of the modern marine stromatolites at Highborne Cay, Bahamas. Atoll Res. Bull. 2009, 567, 1–29. [Google Scholar] [CrossRef]
- Franks, J.; Reid, R.P.; Aspen, R.J.; Underwood, G.J.C.; Paterson, D.M.; Prufert-Bebout, L.; Stolz, J.F. Ooid accreting diatom communities from the modern marine stromatolites at Highborne Cay, Bahamas. In Microbial Mats; Seckbach, J., Oren, A., Eds.; Springer: Berlin, Germany, 2010; pp. 277–285. [Google Scholar]
- Stolz, J.F.; Feinstein, T.N.; Salsi, J.; Visccher, P.T.; Reid, R.P. TEM analysis of microbial mediated sedimentation and lithification in a modern marine stromatolite. Am. Min. 2001, 86, 826–833. [Google Scholar] [CrossRef]
- Kawaguchi, T.; Decho, A.W. In-situ microspatial imaging using two-photon and confocal laser scanning microscopy of bacteria and its extracellular polymeric secretions (EPS) within marine stromatolites. Mar. BioTechnol. 2002, 4, 127–131. [Google Scholar] [CrossRef] [PubMed]
- Paterson, D.M.; Aspden, R.J.; Visscher, P.T.; Consalvey, M.; Andres, M.S.; Decho, A.W.; Stolz, J.; Reid, R.P. Light-Dependant Biostabilisation of Sediments by Stromatolite Assemblages. PLoS Biol. 2008, 3, e3176. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Golubic, S. Organisms that Build Stromatolites. Dev. Sediment. 1976, 20, 113–126. [Google Scholar]
- Papineau, D.; Walker, J.J.; Mojzsis, S.J.; Pace, N.R. Composition and structure of microbial communities from stromatolites of Hamelin Pool in Shark Bay, Western Australia. Appl. Environ. Microbiol. 2005. [Google Scholar] [CrossRef] [PubMed]
- Allen, M.A.; Goh, F.; Burns, B.P.; Neilan, B.A. Bacterial, archaeal and eukaryotic diversity of smooth and pustular microbial mat communities in the hypersaline lagoon of Shark Bay. Geobiology 2009, 7, 82–96. [Google Scholar] [CrossRef] [PubMed]
- Goh, F.; Allen, M.A.; Leuko, S.; Kawaguchi, T.; Decho, A.W.; Burns, B.P.; Neilan, B.A. Determining the specific microbial populations and their spatial distribution within the stromatolite ecosystem of Shark Bay. ISME J. 2009. [Google Scholar] [CrossRef] [PubMed]
- Ruvindy, R.; White, R.A., III; Neilan, B.A.; Burns, B.P. Unravelling core microbial metabolisms in the hypersaline microbial mats of Shark Bay using high-throughput metagenomics. ISME J. 2015. [Google Scholar] [CrossRef] [PubMed]
- Wong, L.H.; Smith, D.L.; Visscher, P.T.; Burns, B.P. Niche differentiation of bacterial communities at a millimeter scale in Shark Bay microbial mats. Sci. Rep. 2015. [Google Scholar] [CrossRef] [PubMed]
- Suosaari, E.P. (Bush Heritage Australia, Hamelin Pool, Western Australia, Australia); Unpublished data for discrete pH water measurements across Hamelin Pool 2014–2017 (environmental monitoring), 2018.
- Awramik, S.M.; Vanyo, J.P. Heliotropism in modern stromatolites. Science 1986, 231, 1279–1281. [Google Scholar] [CrossRef] [PubMed]
- Castenholz, R.W.; Garcia-Pichel, F. Cyanobacterial responses to UV radiation. In Ecology of Cyanobacteria II; Springer: Dordrecht, The Netherlands, 2012; pp. 481–499. [Google Scholar]
- Moon, Y.J.; Kim, S.I.; Chung, Y.H. Sensing and responding to UV-A in cyanobacteria. Int. J. Mol. Sci. 2012, 13, 16303–16332. [Google Scholar] [CrossRef] [PubMed]
- Whale, G.F.; Walsby, A.E. Motility of the cyanobacterium Microcoleus chthonoplastes in mud. Br. Phycol. J. 1984, 19, 117–123. [Google Scholar] [CrossRef]
- Petroff, A.P.; Wu, T.D.; Liang, B.; Mui, J.; Guerquin-Kern, J.L.; Vali, H.; Rothman, D.H.; Bosak, T. Reaction–diffusion model of nutrient uptake in a biofilm: Theory and experiment. J. Theor. Biol. 2011, 289, 90–95. [Google Scholar] [CrossRef] [PubMed]
- Petroff, A.P.; Sim, M.S.; Maslov, A.; Krupenin, M.; Rothman, D.H.; Bosak, T. Biophysical basis for the geometry of conical stromatolites. Proc. Natl. Acad. Sci. USA 2010, 107, 9956–9961. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Decho, A.W.; Norman, R.S.; Visscher, P.T. Quorum sensing in natural environments: Emerging views from microbial mats. Trends Microbiol. 2010, 18, 73–80. [Google Scholar] [CrossRef] [PubMed]
- Castenholz, R.W. The behavior of Oscillatoria terebriformis in hot springs. J. Phycol. 1968, 4, 132–139. [Google Scholar] [CrossRef] [PubMed]
- De Philippis, R.; Margheri, M.C.; Materassi, R.; Vincenzini, M. Potential of unicellular cyanobacteria from saline environments as exopolysaccharide producers. Appl. Environ. Microbiol. 1998, 64, 1130–1132. [Google Scholar] [PubMed]
- Jones, B.; Renaut, R.W.; Rosen, M.R. Microbial biofacies in hot-spring sinters; a model based on Ohaaki Pool, North Island, New Zealand. J. Sediment. Res. 1998, 68, 413–434. [Google Scholar] [CrossRef]
- Grey, K. (Geological Survey of Western Australia, Perth, Western Australia, Australia); Personal communication, 2017.
- Shapiro, J.A. Thinking about bacterial populations as multicellular organisms. Annu. Rev. Microbiol. 1998, 52, 81–104. [Google Scholar] [CrossRef] [PubMed]
- Jones, B.; Renaut, R.W.; Rosen, M.R.; Andsdell, K.M. Coniform stromatolites from geothermal systems, North Island, New Zealand. Palaios 2002, 17, 84–103. [Google Scholar] [CrossRef]
- Hickman, A.H.; Van Kranendonk, M.J.; Grey, K. State Geoheritage Reserve R50149 (Trendall Reserve), North Pole, Pilbara Craton, Western Australia—Geology and Evidence for Early Archean Life; Record 2011/10; Geological Survey of Western Australia: Perth, Australia, 2011; 32p. [Google Scholar]
- Grey, K. (Geological Survey of Western Australia, Perth, Western Australia, Australia); Unpublished data from Pamelup Pond: pinnacle mats, 1990.
- Bauld, J.; D’Amelio, E.; Farmer, J.D. Modern microbial mats. In The Proterozoic Biosphere: An Interdisciplinary Study; Schopf, J.W., Klein, C., Eds.; Cambridge University Press: New York, NY, USA, 1992; pp. 261–269. [Google Scholar]
- Krumbein, W.E.; Cohen, Y. Primary production, mat formation and lithification: Contribution of oxygenic and facultative anoxygenic cyanobacteria. In Fossil Algae; Springer: Berlin/Heidelberg, Germany, 1977; pp. 37–56. [Google Scholar]
- Krumbein, W.E. Calcification by Bacteria and Algae. Stud. Environ. Sci. 1979, 3, 47–68. [Google Scholar]
- Park, R.K. The preservation potential of some recent stromatolites. Sedimentology 1977, 24, 485–506. [Google Scholar] [CrossRef]
- Grotzinger, J.P. Geochemical model for Proterozoic stromatolite decline. Am. J. Sci. 1990, 290, 80–103. [Google Scholar]
- Andrews, J.H. Bacteria as modular organisms. Annu. Rev. Microbiol. 1998, 52, 105–126. [Google Scholar] [CrossRef] [PubMed]
- Shapiro, J.A.; Dworkin, M. Bacteria as Multicellular Organisms; Oxford University Press: Oxford, UK, 1997. [Google Scholar]
- Levine, H.; Ben-Jacob, E. Physical schemata underlying biological pattern formation—Examples, issues and strategies. Phys. Biol. 2004, 1, 14–22. [Google Scholar] [CrossRef] [PubMed]









© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Suosaari, E.P.; Awramik, S.M.; Reid, R.P.; Stolz, J.F.; Grey, K. Living Dendrolitic Microbial Mats in Hamelin Pool, Shark Bay, Western Australia. Geosciences 2018, 8, 212. https://doi.org/10.3390/geosciences8060212
Suosaari EP, Awramik SM, Reid RP, Stolz JF, Grey K. Living Dendrolitic Microbial Mats in Hamelin Pool, Shark Bay, Western Australia. Geosciences. 2018; 8(6):212. https://doi.org/10.3390/geosciences8060212
Chicago/Turabian StyleSuosaari, Erica P., Stanley M. Awramik, R. Pamela Reid, John F. Stolz, and Kathleen Grey. 2018. "Living Dendrolitic Microbial Mats in Hamelin Pool, Shark Bay, Western Australia" Geosciences 8, no. 6: 212. https://doi.org/10.3390/geosciences8060212
APA StyleSuosaari, E. P., Awramik, S. M., Reid, R. P., Stolz, J. F., & Grey, K. (2018). Living Dendrolitic Microbial Mats in Hamelin Pool, Shark Bay, Western Australia. Geosciences, 8(6), 212. https://doi.org/10.3390/geosciences8060212