Microbial and Geochemical Investigation down to 2000 m Deep Triassic Rock (Meuse/Haute Marne, France)

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
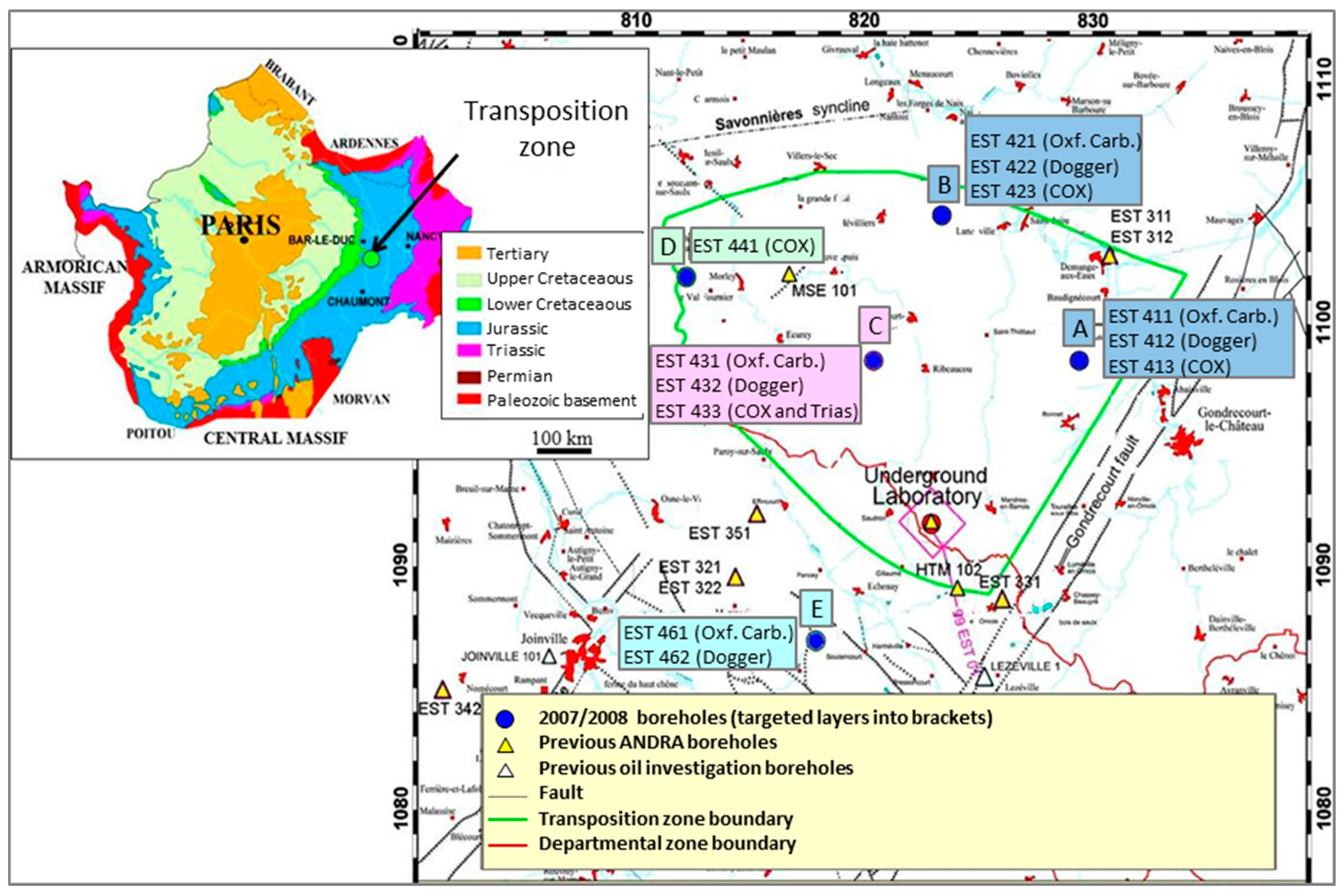
2.1. Site, Drilling and Sampling
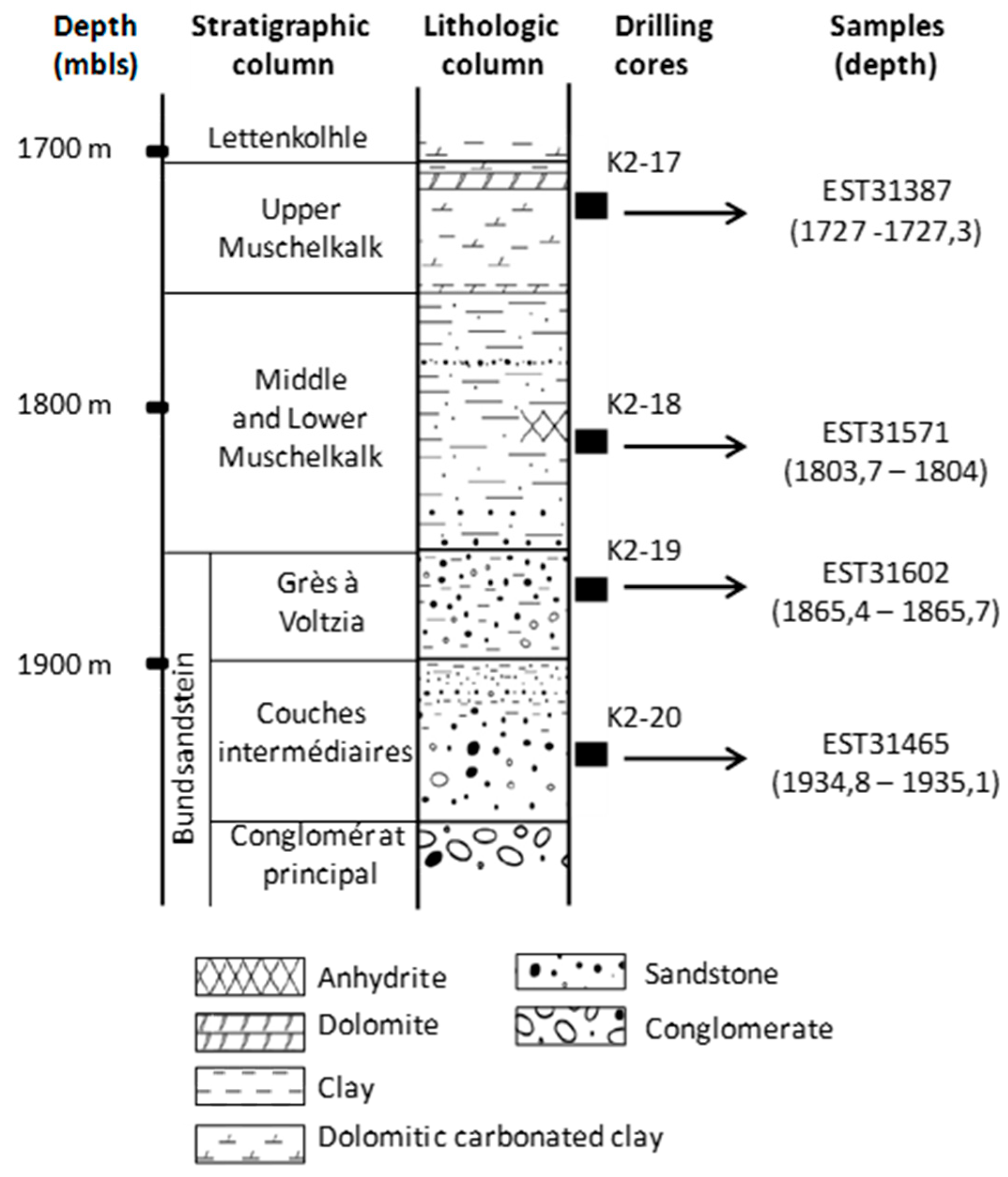
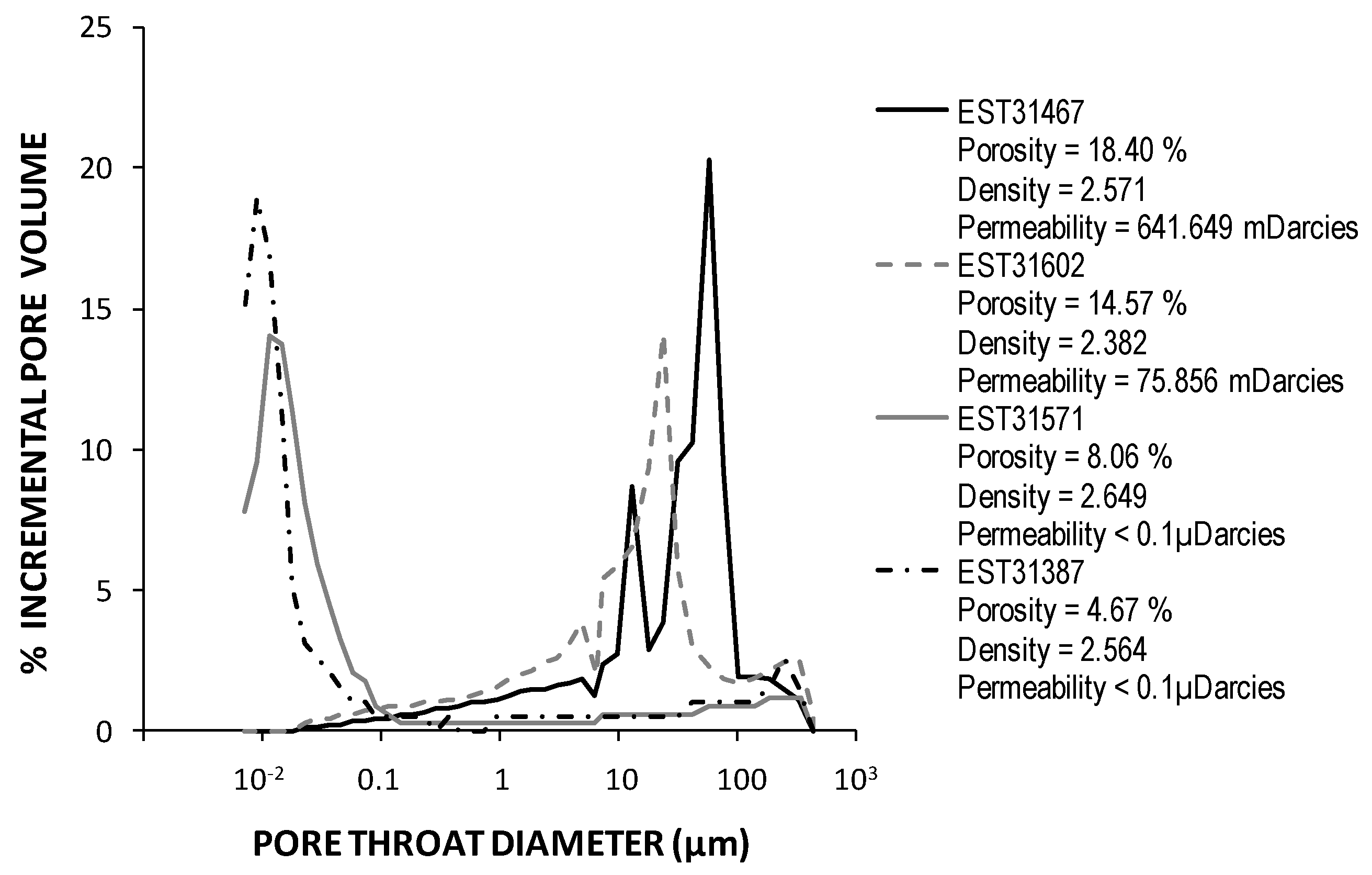
2.2. Geochemical Analyses and Mineralogical Characterisation of the Triassic Cores
2.3. Microbial Enrichments
2.4. DNA Extraction and Optimisation
2.5. PCR Amplification of 16S rRNA Gene Sequences
2.6. DGGE Analysis of Bacterial Diversity
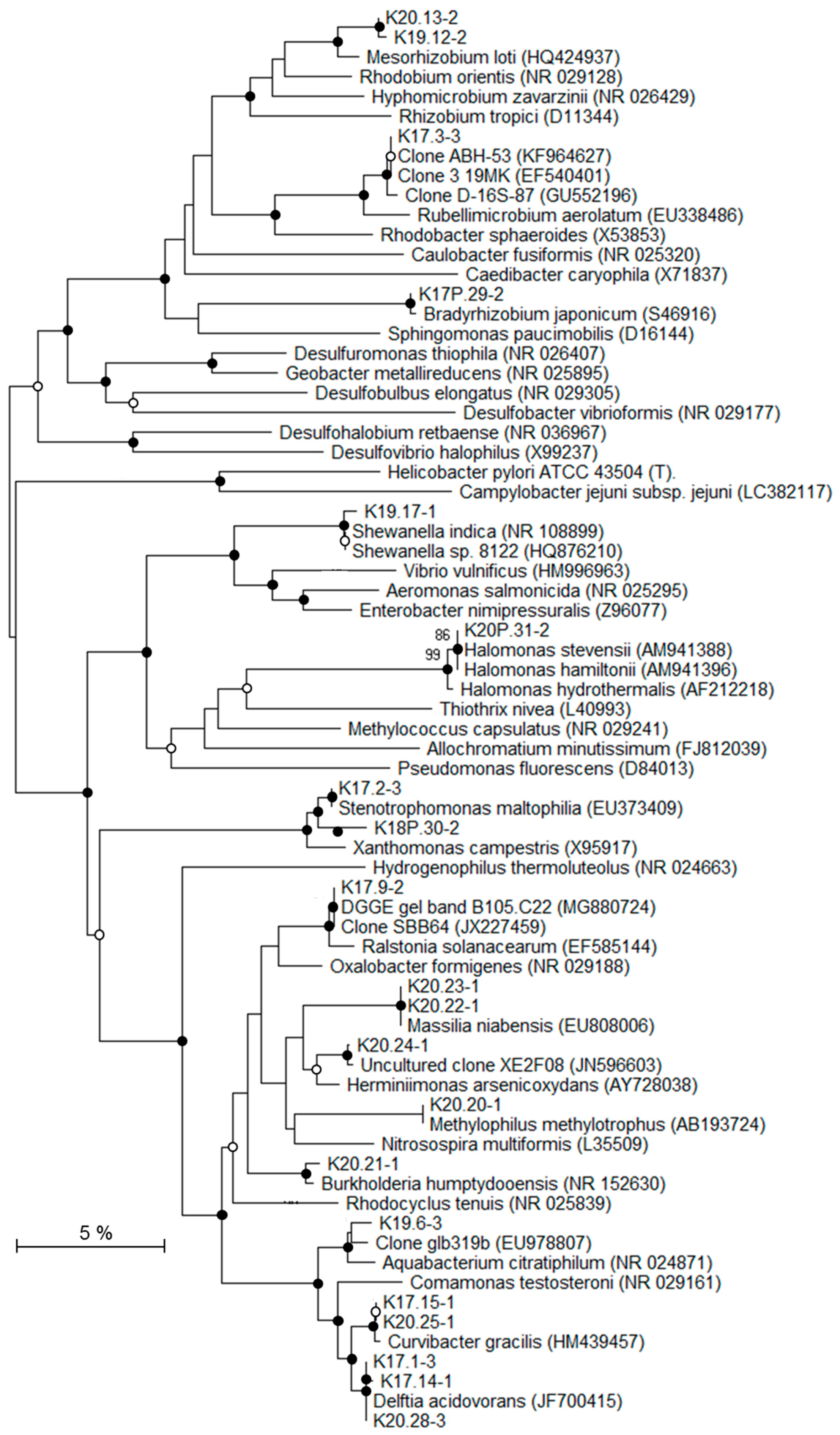
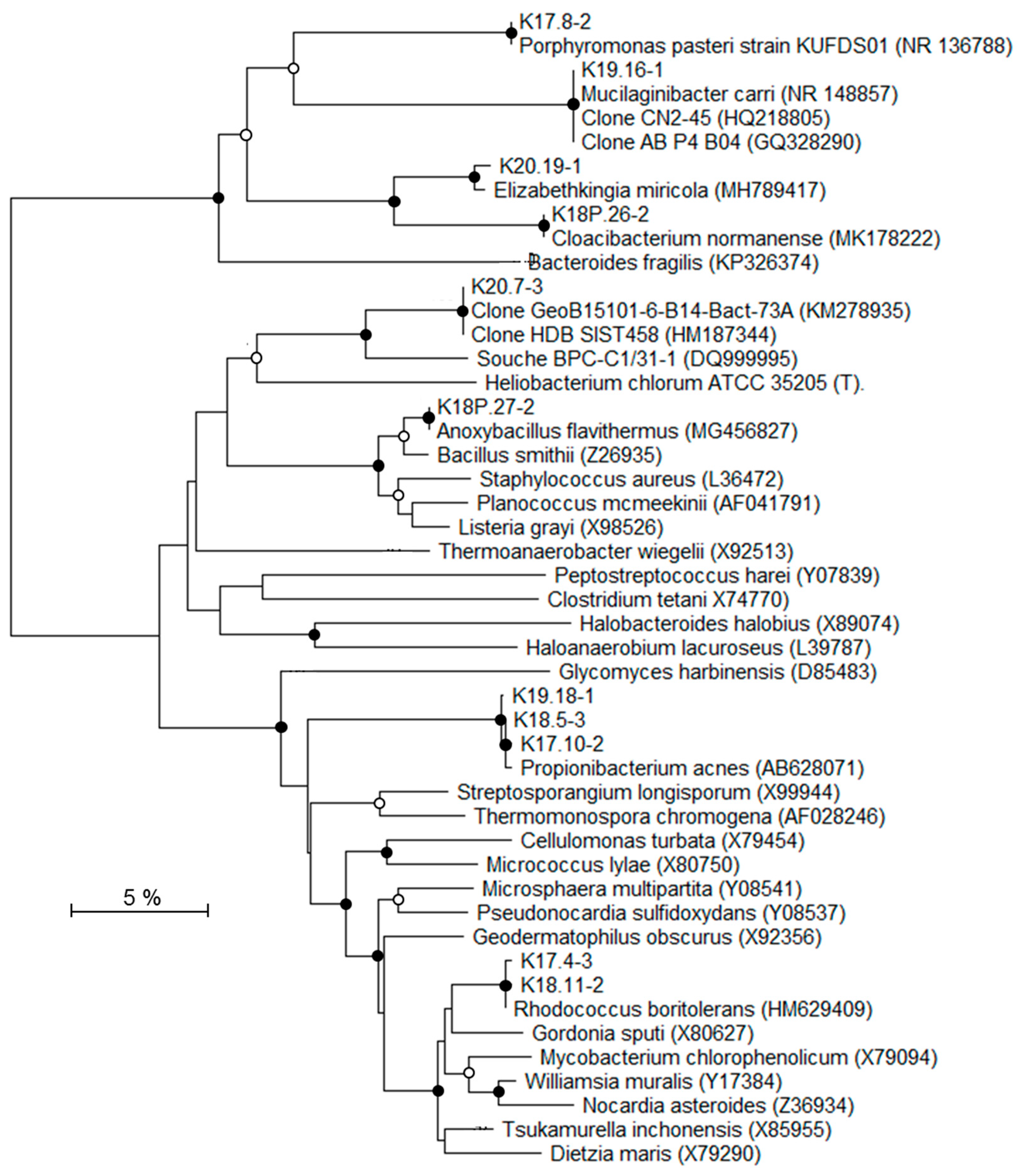
2.7. Phylogenetic Analysis
3. Results
3.1. Characterisation of Deep Triassic Samples
3.2. Exclusion of Potential Contamination Sources
3.3. Microbial Enrichments from Triassic Sandstones
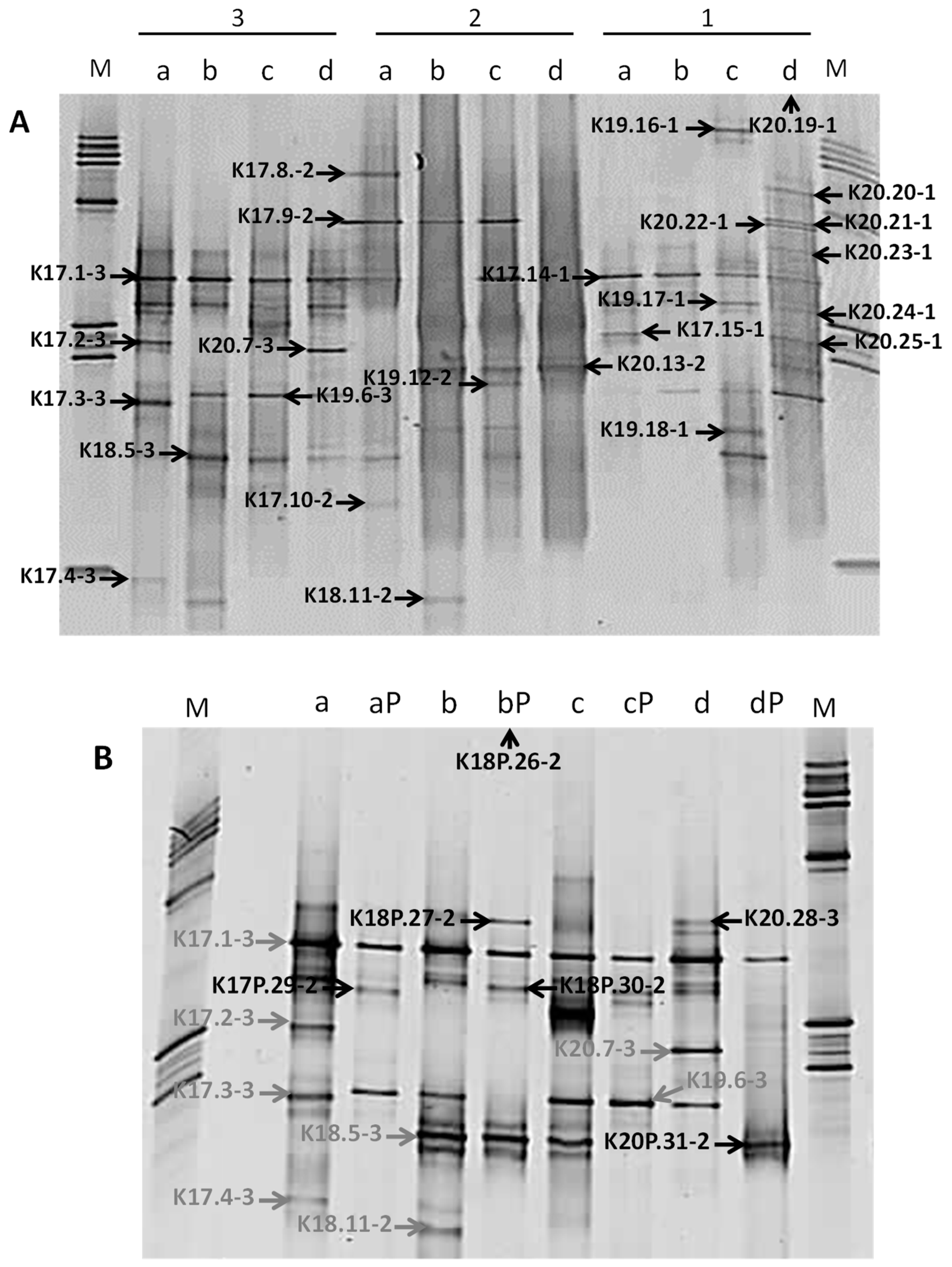
3.4. Molecular Biodiversity of Deep Triassic Formations
4. Discussion
4.1. Life in Deep Geological Formations
4.2. Origins of the Indigenous Bacterial Community
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Kashefi, K.; Lovley, D.R. Extending the upper temperature limit for life. Science 2003, 301, 934. [Google Scholar] [CrossRef] [PubMed]
- Takai, K.; Nakamura, K.; Toki, T.; Tsunogai, U.; Miyazaki, M.; Miyazaki, J.; Hirayama, H.; Nakagawa, S.; Nunoura, T.; Horikoshi, K. Cell proliferation at 122 °C and isotopically heavy CH4 production by a hyperthermophilic methanogen under high-pressure cultivation. Proc. Natl. Acad. Sci. USA 2008, 105, 10949–10954. [Google Scholar] [CrossRef] [PubMed]
- Amend, J.P.; Teske, A. Expanding frontiers in deep subsurface microbiology. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2005, 219, 131–155. [Google Scholar] [CrossRef]
- Miranda-Tello, E.; Fardeau, M.-L.; Thomas, P.; Ramirez, F.; Casalot, L.; Cayol, J.-L.; Garcia, J.-L.; Ollivier, B. Petrotoga mexicana sp. nov., a novel thermophilic, anaerobic and xylanolytic bacterium isolated from an oil-producing well in the Gulf of Mexico. Int. J. Syst. Evol. Microbiol. 2004, 54, 169–174. [Google Scholar] [CrossRef] [PubMed]
- Szewzyk, U.; Szewzyk, R.; Stenström, T.A. Thermophilic, anaerobic bacteria isolated from a deep borehole in granite in Sweden. Proc. Natl. Acad. Sci. USA 1994, 91, 1810–1813. [Google Scholar] [CrossRef]
- Bar-On, Y.M.; Phillips, R.; Milo, R. The biomass distribution on Earth. Proc. Natl. Acad. Sci. USA 2018. [Google Scholar] [CrossRef] [PubMed]
- Magnabosco, C.; Lin, L.-H.; Dong, H.; Bomberg, M.; Ghiorse, H.; Stan-Lotter, H.; Pedersen, K.; Kieft, T.L.; van Heerden, E.; Onstott, T.C. The biomass and biodiversity of the continental subsurface. Nat. Geosci. 2018, 11, 707–717. [Google Scholar] [CrossRef]
- Fredrickson, J.K.; Balkwill, D.L. Geomicrobial processes and biodiversity in the deep terrestrial subsurface. Geomicrobiol. J. 2006, 23, 345–356. [Google Scholar] [CrossRef]
- Oren, A. Bioenergetic aspects of halophilism. Microbiol. Mol. Biol. Rev. 1999, 63, 334–348. [Google Scholar]
- Krumholz, L.R.; McKinley, J.P.; Ulrich, G.A.; Suflita, J.M. Confined subsurface microbialcommunities in Cretaceous rock. Nature 1997, 386, 64–66. [Google Scholar] [CrossRef]
- Magot, M.; Ollivier, B.; Patel, B.K.C. Microbiology of petroleum reservoirs. Antonie Leeuwenhoek 2000, 77, 103–116. [Google Scholar] [CrossRef]
- Pedersen, K. Exploration of deep intraterrestrial microbial life: Current perspectives. FEMS Microbiol. Lett. 2000, 185, 9–16. [Google Scholar] [CrossRef] [PubMed]
- D’Hondt, S.; Jørgensen, B.B.; Miller, D.J.; Batzke, A.; Blake, R.; Cragg, B.A.; Cypionka, H.; Dickens, G.R.; Ferdelman, T.; Hinrichs, K.-U.; et al. Distributions of microbial activities in deep subseafloor sediments. Science 2004, 306, 2216–2221. [Google Scholar] [CrossRef] [PubMed]
- DeLong, E.F. Microbial life breathes deep. Science 2004, 306, 2198–2200. [Google Scholar] [CrossRef] [PubMed]
- Basso, O.; Lascourrèges, J.-F.; Jarry, M.; Magot, M. The effect of cleaning and disinfecting the sampling well on the microbial communities of deep subsurface water samples. Environ. Microbiol. 2005, 7, 13–21. [Google Scholar] [CrossRef] [PubMed]
- Fardeau, M.-L.; Goulhen, F.; Bruschi, M.; Khelifi, N.; Cayol, J.-L.; Ignatiadis, I.; Guyot, F.; Ollivier, B. Archaeoglobus fulgidus and Thermotoga elfii, thermophilic isolates from deep geothermal water of the Paris Basin. Geomicrobiol. J. 2009, 26, 119–130. [Google Scholar] [CrossRef]
- Onstott, T.C.; Moser, D.P.; Pfiffner, S.M.; Fredrickson, J.K.; Brockman, F.J.; Phelps, T.J.; White, D.C.; Peacock, A.; Balkwill, D.; Hoover, R.; et al. Indigenous and contaminant microbes in ultradeep mines. Environ. Microbiol. 2003, 5, 1168–1191. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Purkamo, L.; Bomberg, M.; Nyyssönen, M.; Kukkonen, I.; Ahonen, L.; Itävaara, M. Heterotrophic communities supplied by ancient organic carbon predominate in deep Fennoscandian bedrock fluids. Microb. Ecol. 2015, 69, 319–332. [Google Scholar] [CrossRef] [PubMed]
- Onstott, T.; Phelps, T.J.; Colwell, F.; Ringelberg, D.; White, D.; Boone, D.; Mckinley, J.P.; Stevens, T.O.; Long, P.E.; Balkwill, D. Observations pertaining to the origin and ecology of microorganisms recovered from the deep subsurface of Taylorsville Basin, Virginia. Geomicrobiol. J. 1998, 15, 353–385. [Google Scholar] [CrossRef]
- Zhang, G.; Dong, H.; Xu, Z.; Zhao, D.; Zhang, C. Microbial diversity in ultra-high-pressure rocks and fluids from the chinese continental scientific drilling project in China. Appl. Environ. Microbiol. 2005, 71, 3213–3227. [Google Scholar] [CrossRef]
- Colwell, F.S.; Onstott, T.C.; Delwiche, M.E.; Chandler, D.; Fredrickson, J.K.; Yao, Q.-J.; McKinley, J.P.; Boone, D.R.; Griffiths, R.; Phelps, T.J.; et al. Microorganisms from deep, high temperature sandstones: Constraints on microbial colonization. FEMS Microbiol. Rev. 1997, 20, 425–435. [Google Scholar] [CrossRef]
- Landrein, P.; Vigneron, G.; Delay, V.; Lebon, P.; Pagel, M. Lithologie, hydrodynamisme et thermicité dans le système sédimentaire multicouche recoupé par les forages Andra de Montiers-sur-Saulx (Meuse). Bull. de la Société Géologique de Fr. 2013, 184, 519–543. [Google Scholar] [CrossRef]
- Brunet, M.-F.; Le Pichon, V. Subsidence of the Paris Basin. J. Geophys. Res. 1982, 87, 8547–8560. [Google Scholar] [CrossRef]
- Poulain, S.; Sergeant, C.; Simonoff, M.; Le Marrec, C.; Altmann, S. Microbial investigations in Opalinus Clay, an argillaceous formation under evaluation as a potential host rock for a radioactive waste repository. Geomicrobiol. J. 2008, 25, 240–249. [Google Scholar] [CrossRef]
- NF T 90-008 2001. Water Quality—Determination of pH; AFNOR: Paris, France, 2001; ISBN 2-12-213141-1. (In French) [Google Scholar]
- ISO 11732 2005. Water Quality—Measurement of Ammonium—Method by Flow Analysis (CFA and FIA) and Spectrometrical Detection; AFNOR: Paris, France, 2005; ISBN 2-12-213141-1. (In French) [Google Scholar]
- ISO 9963-1 1994. Water Quality—Determination of Alkalinity—Part 1: Determination of Total and Composite Alkalinity; AFNOR: Paris, France, 1994; ISBN 2-12-213141-1. (In French) [Google Scholar]
- ISO 10304-1 2010. Water Quality—Measurement of Dissolved Anions by Ionic Chromatography in Liquid Phase—Part 1: Measurement of Bromide, Chloride, Fluoride, Nitrate, Nitrite, Phosphate and Sulphate; AFNOR: Paris, France, 2010; ISBN 2-12-213141-1. (In French) [Google Scholar]
- NF T 01-041 1975. Atomic Absorption Spectrometry—Use Introduction; AFNOR: Paris, France, 1975; ISBN 2-12-213141-1. (In French) [Google Scholar]
- Copard, Y.; Di-Giovanni, C.; Martaud, T.; Albéric, P.; Olivier, J.-E. Using Rock-Eval 6 pyrolysis for tracking fossil organic carbon in modern environments: Implications for the roles of erosion and weathering. Earth Surf. Process. Landf. 2006, 31, 135–153. [Google Scholar] [CrossRef]
- EN ISO 11885. Water Quality—Determination of Selected Elements by Inductively Coupled Plasma Optical Emission Spectrometry (ICP-OES); AFNOR: Paris, France, 2009. (In French) [Google Scholar]
- Blanc, P.; Lassin, A.; Piantone, P.; Azaroual, M.; Jacquemet, N.; Fabbri, A.; Gaucher, E.C. Thermoddem: A geochemical database focused on low temperature water/rock interactions and waste materials. Appl. Geochem. 2012, 27, 2107–2116. [Google Scholar] [CrossRef]
- Parkhurst, D.L.; Appelo, C.A.J. User’s Guide to Phreeqc (Version 2)—A Computer Program for Speciation, Batch-Reaction, One-Dimensional Transport, and Inverse Geochemical Calculations; Water-Resources Investigations Report; U.S. Department of the Interior, USGS: Denver, CO, USA, 1999; pp. 99–4259.
- Miller, T.L.; Wolin, M.J. A serum bottle modification of the Hungate technique for cultivating obligate anaerobes. Appl. Microbiol. 1974, 27, 985–987. [Google Scholar] [PubMed]
- Kallmeyer, J.; Ferdelman, T.G.; Jansen, K.H.; Jørgensen, B.B. A high-pressure thermal gradient block for investigating microbial activity in multiple deep-sea samples. J. Microbiol. Methods 2003, 55, 165–172. [Google Scholar] [CrossRef]
- Lerner, A.; Shor, Y.; Vinokurov, A.; Okon, Y.; Jurkevitch, E. Can denaturing gradient gel electrophoresis (DGGE) analysis of amplified 16s rDNA of soil bacterial populations be used in forensic investigations? Soil Biol. Biochem. 2006, 38, 1188–1192. [Google Scholar] [CrossRef]
- Lane, D.J. 16S/23S rRNA sequencing. In Nucleic Acid Techniques in Bacterial Systematics; Stackebrandt, E., Goodfellow, M., Eds.; Wiley Press: Chichester, UK, 1991; pp. 130–141. [Google Scholar]
- Schäfer, H.; Muyzer, G. Denaturing gradient gel electrophoresis in marine microbial ecology. Mar. Microbiol. 2001, 30, 425–468. [Google Scholar]
- Baker, G.C.; Smith, J.J.; Cowan, D.A. Review and re-analysis of domain-specific 16S primers. J. Microbiol. Methods 2003, 55, 541–555. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zumstein, E.; Moletta, R.; Godon, J.-J. Examination of two years of community dynamics in an anaerobic bioreactor using fluorescence polymerase chain reaction (PCR) single-strand conformation polymorphism analysis. Environ. Microbiol. 2000, 2, 1462–2920. [Google Scholar] [CrossRef]
- Huber, T.; Faulkner, G.; Hugenholtz, P. Bellerophon: A program to detect chimeric sequences in multiple sequence alignments. Bioinformatics 2004, 20, 2317–2319. [Google Scholar] [CrossRef] [PubMed]
- Hall, T. BioEdit: A user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucleic Acids Symp. Ser. 1999, 41, 95–98. [Google Scholar]
- Jukes, T.H.; Cantor, C.R. Evolution of protein molecules. In Mammalian Protein Metabolism; Munro, H.N., Ed.; Academic Press: New York, NY, USA, 1969; pp. 21–132. [Google Scholar]
- Saitou, N.; Nei, M. The neighbor-joining method: A new method for reconstructing phylogenetic trees. Mol. Biol. Evol. 1987, 4, 406–425. [Google Scholar]
- Felsenstein, J. Confidence limits on phylogenies: An approach using the bootstrap. Evolution 1985, 39, 783–791. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Katz, A.J.; Thompson, A.H. Quantitative prediction of permeability in porous rock. Phys. Rev. Ser. B 1986, 34, 8179–8191. [Google Scholar] [CrossRef]
- Rebeix, R.; Le Gal La Salle, C.; Verdoux, P.; Gianesinni, S.; Lancelot, J.; Michelot, J.-L.; Noret, A.; Monvoisin, G.; Simler, R. Tracing the origin of water and solute transfers in deep groundwater from Oxfordian, Dogger and Trias formations in the east of the Paris Basin-France. Phys. Chem. Earth 2011, 36, 1496–1510. [Google Scholar] [CrossRef]
- Barton, H.A.; Taylor, N.M.; Lubbers, B.R.; Pemberton, A.C. DNA extraction from low-biomass carbonate rock: An improved method with reduced contamination and the low-biomass contaminant database. J. Microbiol. Methods 2006, 66, 21–31. [Google Scholar] [CrossRef]
- Gérard, E.; Moreira, D.; Philippot, P.; van Kranendonk, M.J.; López-García, P. Modern subsurface bacteria in pristine 2.7 Ga-old fossil xtromatolite drillcore samples from the fortescue group, Western Australia. PLoS ONE 2009, 4, e5298. [Google Scholar] [CrossRef]
- Moissl, C.; Osman, S.; La Duc, M.T.; Dekas, A.; Brodie, E.; DeSantis, T.; Venkateswaran, K. Molecular bacterial community analysis of clean rooms where spacecraft are assembled. FEMS Microbiol. Ecol. 2007, 61, 509–521. [Google Scholar] [CrossRef] [Green Version]
- Hashim, S.O.; Delgado, O.; Hatti-Kaul, R.; Mulaa, F.J.; Mattiasson, B. Starch hydrolysing Bacillus halodurans isolates from a Kenyan soda lake. Biotechnol. Lett. 2004, 26, 823–828. [Google Scholar] [CrossRef] [PubMed]
- Ulfah, M.; Helianti, I.; Wahyuntari, B.; Nurhayati, N. Characterization of a new thermoalkalophilic xylanase-producing bacterial strain isolated from Cimanggu hot spring, west Java, Indonesia. Microbiol. Indones. 2012, 5, 139–143. [Google Scholar] [CrossRef]
- Reysenbach, A.-L.; Ehringer, M.; Hershberger, K. Microbial diversity at 83 °C in Calcite Springs, Yellowstone National Park: Another environment where the Aquificales. Extremophiles 2000, 4, 61–67. [Google Scholar] [PubMed]
- Chandler, D.P.; Brockman, F.J.; Fredrickson, J.K. Use of 16S rDNA clone libraries to study changes in a microbial community resulting from ex situ perturbation of a subsurface sediment. FEMS Microbiol. Rev. 1997, 20, 217–230. [Google Scholar] [CrossRef] [Green Version]
- Luna, G.M.; Dell’Anno, A.; Danovaro, R. DNA extraction procedure: A critical issue for bacterial diversity assessment in marine sediments. Environ. Microbiol. 2006, 8, 308–320. [Google Scholar] [CrossRef]
- Martin-Laurent, F.; Philippot, L.; Hallet, S.; Chaussod, R.; Germon, J.C.; Soulas, G.; Catroux, G. DNA extraction from soils: Old bias for new microbial diversity analysis methods. Appl. Environ. Microbiol. 2001, 67, 2354–2359. [Google Scholar] [CrossRef]
- Okubo, T.; Tsukui, T.; Maita, H.; Okamoto, S.; Oshima, K.; Fujisawa, T.; Saito, A.; Futamata, H.; Hattori, R.; Shimomura, Y. Complete genome sequence of Bradyrhizobium sp. S23321: Insights into symbiosis evolution in soil oligotrophs. Microbes Environ. 2012, 27, 306–315. [Google Scholar] [CrossRef]
- Marchesi, J.R.; Weightman, A.J.; Cragg, B.A.; Parkes, R.J.; Fry, J.C. Methanogen and bacterial diversity and distribution in deep gas hydrate sediments from the Cascadia Margin as revealed by 16S rRNA molecular analysis. FEMS Microbiol. Ecol. 2001, 34, 221–228. [Google Scholar] [CrossRef] [Green Version]
- Mergeay, M.; Monchy, S.; Vallaeys, T.; Auquier, V.; Benotmane, A.; Bertin, P.; Taghavi, S.; Dunn, J.; van der Lelie, D.; Wattiez, R. Ralstonia metallidurans, a bacterium specifically adapted to toxic metals: Towards a catalogue of metal-responsive genes. FEMS Microbiol. Rev. 2003, 27, 385–410. [Google Scholar] [CrossRef]
- Tiemeyer, A.; Link, H.; Weuster-Botz, D. Kinetic studies on autohydrogenotrophic growth of Ralstonia eutropha with nitrate as terminal electron acceptor. Appl. Microbiol. Biotechnol. 2007, 76, 75–81. [Google Scholar] [CrossRef] [PubMed]
- Grey, B.E.; Steck, T.R. The viable but nonculturable state of Ralstonia solanacearum may be involved in long-term survival and plant infection. Appl. Environ. Microbiol. 2001, 67, 3866–3872. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.M.; Cho, N.-T.; Yang, S.-H.; Arun, A.; Young, C.-C.; Sheu, S.-Y. Aquabacterium limnoticum sp. nov., isolated from a freshwater spring. Int. J. Syst. Evol. Microbiol. 2012, 62, 698–704. [Google Scholar] [CrossRef] [PubMed]
- Kalmbach, S.; Manz, W.; Wecke, J.; Szewzyk, U. Aquabacterium gen. nov., with description of Aquabacterium citratiphilum sp. nov., Aquabacterium parvum sp. nov. and Aquabacterium commune sp. nov., three in situ dominant bacterial species from the Berlin drinking water system. Int. J. Syst. Bacteriol. 1999, 49, 769–777. [Google Scholar] [CrossRef] [PubMed]
- Simon, C.; Wiezer, A.; Strittmatter, A.W.; Daniel, R. Phylogenetic diversity and metabolic potential revealed in a glacier ice metagenome. Appl. Environ. Microbiol. 2009, 75, 7519–7526. [Google Scholar] [CrossRef] [PubMed]
- Santelli, C.M.; Orcutt, B.N.; Banning, E.; Bach, W.; Moyer, C.L.; Sogin, M.L.; Staudigel, H.; Edwards, K.J. Abundance and diversity of microbial life in ocean crust. Nature 2008, 453, 653–656. [Google Scholar] [CrossRef] [PubMed]
- Gong, X.-C.; Liu, Z.-S.; Guo, P.; Chi, C.-Q.; Chen, J.; Wang, X.-B.; Tang, Y.-Q.; Wu, X.-L.; Liu, C.-Z. Bacteria in crude oil survived autoclaving and stimulated differentially by exogenous bacteria. PLoS ONE 2012, 7, e40842. [Google Scholar] [CrossRef] [PubMed]
- Christner, B.C.; Royston-Bishop, G.; Foreman, C.M.; Arnold, B.R.; Tranter, M.; Welch, K.A.; Lyons, W.B.; Tsapin, A.I.; Studinger, M.; Priscu, J.C. Limnological conditions in subglacial lake Vostok, Antarctica. Limnol. Oceanogr. 2006, 51, 2485–2501. [Google Scholar] [CrossRef]
- Colquhoun, J.A.; Heald, S.C.; Li, L.; Tamaoka, J.; Kato, C.; Horikoshi, K.; Bull, A.T. Taxonomy and biotransformation activities of some deep-sea actinomycetes. Extremophiles 1998, 2, 269–277. [Google Scholar] [CrossRef]
- Kevbrin, V.V.; Zengler, K.; Lysenko, A.M.; Wiegel, J. Anoxybacillus kamchatkensis sp. nov., a novel thermophilic facultative aerobic bacterium with a broad pH optimum from the Geyser valley, Kamchatka. Extremophiles 2005, 9, 391–398. [Google Scholar] [CrossRef] [PubMed]
- Willerslev, E.; Cooper, A. Ancient DNA. Proc. R. Soc. B 2005, 272, 3–16. [Google Scholar] [CrossRef] [PubMed]
- Lin, X.; Kennedy, D.; Fredrickson, J.; Bjornstad, B.; Konopka, A. Vertical stratification of subsurface microbial community composition across geological formations at the Hanford Site. Environ. Microbiol. 2012, 14, 414–425. [Google Scholar] [CrossRef]
- Eloe, E.A.; Shulse, C.N.; Fadrosh, D.W.; Williamson, S.J.; Allen, E.E.; Bartlett, D.H. Compositional differences in particle-associated and free-living microbial assemblages from an extreme deep-ocean environment. Environ. Microbiol. Rep. 2011, 3, 449–458. [Google Scholar] [CrossRef]
- Elhottová, D.; Krištůfek, V.; Frouz, J.; Nováková, A.; Chroňáková, A. Screening for microbial markers in Miocene sediment exposed during open-cast brown coal mining. Antonie Leeuwenhoek 2006, 89, 459–463. [Google Scholar] [CrossRef] [PubMed]
- Wilson, M.S.; Herrick, J.B.; Jeon, C.O.; Hinman, D.E.; Madsen, E.L. Horizontal transfer of phnAc dioxygenase genes within one of two phenotypically and genotypically distinctive naphthalene-degrading guilds from adjacent soil environments. Appl. Environ. Microbiol. 2003, 69, 2172–2181. [Google Scholar] [CrossRef] [PubMed]
- Verma, P.; Pandey, P.K.; Gupta, A.K.; Kim, H.J.; Baik, K.S.; Seong, C.N.; Patole, M.S.; Shouche, Y.S. Shewanella indica sp. nov., isolated from sediment of the Arabian Sea. Int. J. Syst. Evol. Microbiol. 2011, 61, 2058–2064. [Google Scholar] [CrossRef] [PubMed]
- Kaye, J.Z.; Márquez, M.C.; Ventosa, A.; Baross, J.A. Halomonas neptunia sp. nov., Halomonas sulfidaeris sp. nov., Halomonas axialensis sp. nov. and Halomonas hydrothermalis sp. nov.: Halophilic bacteria isolated from deep-sea hydrothermal-vent environments. Int. J. Syst. Evol. Microbiol. 2004, 54, 499–511. [Google Scholar] [CrossRef]
- Lauer, A.; Sørensen, K.B.; Teske, A. Phylogenetic Characterization of Marine Benthic Archaea in Organic-Poor Sediments of the Eastern Equatorial Pacific Ocean (ODP Site 1225). Microorganisms 2016, 4, 32. [Google Scholar] [CrossRef]
- Frank, Y.A.; Kadnikov, V.V.; Gavrilov, S.N.; Banks, D.; Gerasimchuk, A.L.; Podosokorskaya, O.A.; Merkel, A.Y.; Chernyh, N.A.; Mardanov, A.V.; Ravin, N.V.; et al. Stable and Variable Parts of Microbial Community in Siberian Deep Subsurface Thermal Aquifer System Revealed in a Long-Term Monitoring Study. Front. Microbiol. 2016. [Google Scholar] [CrossRef]
- Bomberg, M.; Nyyssönen, M.; Nousiainen, A.; Hultman, J.; Paulin, L.; Auvinen, P.; Itävaara, M. Evaluation of Molecular Techniques in Characterization of Deep Terrestrial Biosphere. Open J. Ecol. 2014, 4, 468–487. [Google Scholar] [CrossRef]
- Jenneman, G.E.; McInerney, M.J.; Knapp, R.M. Microbial penetration through nutrient-saturated berea sandstone. Appl. Environ. Microbiol. 1985, 50, 383–391. [Google Scholar] [PubMed]
- Alain, K.; Querellou, J. Cultivating the uncultured: Limits, advances and future challenges. Extremophiles 2009, 13, 583–594. [Google Scholar] [CrossRef] [PubMed]
- Ollivier, B.; Alazard, D. The oil reservoir ecosystem. In Handbook of Hydrocarbon Microbiology: Microbial Interactions with Hydrocarbons, Oils, Fats and Related Hydrophobic Substrates and Products; Timmis, K.N., Ed.; Springer: Berlin, Bermany, 2010; pp. 2262–2268. [Google Scholar]
- McGenity, T.J.; Gemmell, R.T.; Grant, W.D.; Stan-Lotter, H. Origins of halophilic microorganisms in ancient salt deposits. Environ. Microbiol. 2000, 2, 243–250. [Google Scholar] [CrossRef] [PubMed]
- Blaise, T.; Barbarand, J.; Kars, M.; Ploquin, F.; Aubourg, C.; Bribaud, B.; Cathelineau, M.; El Albani, A.; Gautheron, C.; Izart, A.; et al. Reconstruction of low burial (<100 °C) in sedimentary basins: A comparison of geothermometer sensibility in the intracontinental Paris Basin. Marine Petroleum Geol. 2014, 53, 71–87. [Google Scholar] [CrossRef]
- Wilhelm, A.; Larter, S.R.; Head, I.; Farrimond, P.; di-Primio, R.; Zwach, C. Biodegradation of oil in uplifted basins prevented by deep-burial sterilization. Nature 2001, 411, 1034–1037. [Google Scholar] [CrossRef] [PubMed]
- Yamane, K.; Maki, H.; Nakayama, T.; Nakajima, T.; Nomura, N.; Uchiyama, H.; Kitaoka, M. Diversity and similarity of microbial communities in petroleum crude oils produced in Asia. Biosc. Biotechnol. Biochem. 2008, 72, 2831–2839. [Google Scholar] [CrossRef] [PubMed]
- Fish, S.A.; Shepherd, T.J.; McGenity, T.J.; Grant, W.D. Recovery of 16S ribosomal RNA gene fragments from ancient halite. Nature 2002, 417, 432–436. [Google Scholar] [CrossRef] [PubMed]
- Gales, G.; Tsesmetzis, N.; Neria, I.; Alazard, D.; Coulon, S.; Lomans, B.P.; Morin, D.; Ollivier, B.; Borgomano, J.; Joulian, C. Preservation of ancestral Cretaceous microflora recovered from a hypersaline oil reservoir. Sci. Rep. 2016, 6, 22960. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Inagaki, F.; Okada, H.; Tsapin, A.I.; Nealson, K.H. Microbial survival: The Paleome: A Sedimentary Genetic Record of Past Microbial Communities. Astrobiology 2005, 5, 141–153. [Google Scholar] [CrossRef] [PubMed]
- Marty, B.; Dewonck, S.; France-Lanord, C. Geochemical evidence for efficient aquifer isolation over geological timeframes. Nature 2003, 425, 55–58. [Google Scholar] [CrossRef] [PubMed]
- Celle-Jeanton, H.; Huneau, F.; Travi, Y.; Edmunds, W.M. Twenty years of groundwater evolution in the Triassic sandstone aquifer of Lorraine: Impacts on baseline water quality. Appl. Geochem. 2009, 24, 1198–1213. [Google Scholar] [CrossRef]
- Fontes, J.C.; Matray, J.M. Geochemistry and origin of formation brines from the Paris Basin, France: 1. Brines associated with Triassic salts. Chem. Geol. 1993, 109, 149–175. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Metabolic Guilds | Minimum Media | Yeast Extract (g·L−1) | Peptone (g·L−1) | HEPES (mM) | Energy Sources (mM) | Electron Acceptors (mM) | L-Cysteine HCl·H2O (mM) | Na2S·9H2O (mM) | NaHCO3(mM) | |
|---|---|---|---|---|---|---|---|---|---|---|
| Aerobic | ||||||||||
| Chemoorganotrophs | AEM1 | 1 | 0.4 | 2 | 50 | Glucose (10) | O2 | 12 | ||
| AE | 2 | 5 | 5 | Glucose (20) | O2 | |||||
| AAEM1 | 3 | 5 | 5 | O2 | 48 | |||||
| Anaerobic | ||||||||||
| Fermentative | ANM1 | 1 | 0.4 | 2 | 50 | Glucose (10) | 12 | |||
| F1 | 2 | 5 | 5 | Glucose (20) | 2 | 20 | ||||
| F2 | 2+ | 1 | 1 | Fructose (10) | 20 | |||||
| AANM1 | 3 | 5 | 5 | 48 | ||||||
| Nitrate reducers | IRD1 | 4 | 1 | Glucose (20) | NaNO3 (20) | 24 | ||||
| Thiosulfate reducers | IRD2 | 4 | Lactate (20) | Na2S2O3·5H2O (20) | 2.85 | 2 | 24 | |||
| IRD3 | 4 | 0.2 | 5 | H2 (2 bar) | Na2S2O3·5H2O (20) | 2.85 | 2 | 24 | ||
| Sulfate reducers | ANM5 | 1 | 0.2 | Lactate (10) | Na2SO4 (14) | 2.85 | 2 | 12 | ||
| L | 2 | 0.5 | 0.5 | Lactate (20) | Na2SO4 (20) | 2.85 | 2 | 1.7 | ||
| B | 2+ | 0.5 | Lactate (10) and pyruvate (10) | Na2SO4 (35) | 1.5 | 20 | ||||
| HL | 2 | 0.5 | 0.5 | H2 (2 bar) | Na2SO4 (20) | 2.85 | 2 | 20 | ||
| Methanogens | M | 2 | 0.5 | 0.5 | Methanol (40) | CO2 | 2.85 | 2 | 20 | |
| C | 2+ | 0.5 | Methanol (40) | CO2 | 2.85 | 1.5 | 20 | |||
| IRD4 | 4 | TMA a-HCl (50) | CO2 | 2.85 | 2 | 24 | ||||
| IRD5 | 4 | H2 (2 bar) | CO2 | 2.85 | 2 | 24 | ||||
| ANM4 | 1 | 0.2 | CO2, carbonate | CO2 | 2.85 | 2 | 12 | |||
| HM | 2 | 0.5 | 0.5 | H2 (2 bar) | CO2 | 2.85 | 2 | 20 | ||
| Other lithotrophs | ANM3 | 1 | 0.1 | CO2 | AQDS b (20) | 12 | ||||
| Others, on reconstituted pore waters | EP-18 | 5 | 1 | 0.95 | ||||||
| EP-17 | 6 | 1 | 1.55 | |||||||
| SS | Sea salt (Sigma) | 1 | 2 | |||||||
| Sample | Depth (mbsl) | Cl | Br | SO4 | F | NH4 | Na | Ca | K | Mg | |
|---|---|---|---|---|---|---|---|---|---|---|---|
| Cores | |||||||||||
| K2-17 | EST31387 | 1727 | 17.80 | 0.08 | 24.91 | 0.05 | 0.00831 | 165.22 | 63 | 0.5 | 1.9 |
| K2-18 | EST31571 | 1823 | 39.27 | 0.18 | 22.60 | 0.16 | 0.014 | 191.30 | 71 | 1.2 | 1.2 |
| K2-19 | EST31602 | 1885 | 61.10 | 0.29 | 8.69 | 0.11 | 0.04 | 121.74 | 8.75 | 1.6 | 1.2 |
| K2-20 | EST31467 | 1956 | 68.76 | 0.34 | 0.12 | 0.11 | 0.00942 | 104.35 | 0.8 | 1.2 | 1.3 |
| Groundwater | |||||||||||
| EST03257 | 1900 | 2579.13 | 15.46 | 12.18 | nd | nd | 1976.97 | 167.2 | 55.12 | 147.77 |
| Phylogenetic Group | Isolates and Denaturing Gradient Gel Electrophoresis (DGGE) Bands | Closest Identified Relative (Sequence Identity) | Isolation Sources |
|---|---|---|---|
| Actinobacteria | |||
| C5a | Williamsia (99%) | Soils, Human | |
| C2 | Micrococcus (99%) | Cold environment, Human, deep marine sediments | |
| C5b | Salinibacterium (99%) | Human, marine environment | |
| C10e | Agrococcus jenensis (99%) | Alkaline environment, Human, limestone | |
| C4 | Cellulomonas (99%) | Sediments, permafrost, soils | |
| C9, C10d | Microbacterium oxydans (100%) | Black shales, cold environment, Human | |
| Firmicutes | |||
| C3 | Sporosarcina (99%) | Sediment, permafrost, soil | |
| C6, C10b, C10c | Staphylococcus epidermidis (100%) | Soil, sea water, sediments, Human | |
| C8 | Brevibacterium (99%) | Marine sediments, soil, rocks | |
| C12, C15 | Bacillus (99%) | Soils, sediments | |
| DFO-AE7-1/2 | Bacillus halodurans (99%) | Soil, hot spring, oceanic crust | |
| DFO-AE11-1/2 | Bacillus polygoni (99%) | Leguminous shrub, sandstones | |
| DFW-AE7-6a, DFO-AE7-6c | Aerococcus viridans (99%) | Human, sea water | |
| Alphaproteobacteria | |||
| DFW-AE7-1/2a | Sphingobium yanoikuyae (99%) | Drinking water, Human | |
| DFW-AE7-7a | Methylobacterium extorquens (99%) | Contaminated environments, clean room | |
| CTRL1 | Ochrobactrum anthropi (99%) | Soils, sediments, Human | |
| DFW-AE7-6b | Sphingomonas (100%) | soil, Human | |
| DFW-AE7-4, DFO-AE7-6a, DFO-AE7-6b | Sphingomonas (99%) | Industrial plants, clean room, Human | |
| Betaproteobacteria | |||
| DFW-AE7-7b, DFW-AE7-7c | Massilia (99%) | Soil and contaminated soils, Human, geothermal spring | |
| CTRL2, CTRL6 | Delftia acidovorans (100%) | Sewage treatment and activated sludge, soils, marine sediments, rocks | |
| Gammaproteaobacteria | |||
| C7a | Pseudomonas (99%) | Glaciers, Human, soils | |
| C10a | Acinetobacter lwoffii (99%) | Compost, water, Human | |
| C7b, C16, CTRL3, CTRL4 | Stenotrophomonas maltophilia (99%) | Plants, Human, lake, young oceanic crust | |
| Phylogenetic Group | DGGE Bands | Samples (Depth [mbsl]) | Closest Related Genus or Environmental Clone (% Blast Similarity) | Deep Bacterial Community [*] | |||
|---|---|---|---|---|---|---|---|
| K2-20 (1935) | K2-19 (1865) | K2-18 (1804) | K2-17 (1727) | ||||
| α-Proteobacteria | |||||||
| K20.13-2 K19.12-2 | × | × × | × | Mesorhizobium (99%) | Endogenous [73] | ||
| K17P.29-2 | × | Bradyrhizobium (100%) | Potentially endogenous [58,59] | ||||
| K17.3-3 | × | Uncultured Rebullimicrobium clone (99%) | Endogenous | ||||
| β-Proteobacteria | |||||||
| K17.9-2 | × | × | × | Ralstonia (99%) | Potentially endogenous [60,61,62] | ||
| K20.21-1 | × | Burkholderia (99%) | Endogenous [74] | ||||
| K19.6-3 | × | × | × | Aquabacterium (98%) | Potentially endogenous [63,64,65] | ||
| K17.15-1 K20.25-1 | × | × | × | Curvibacter gracilis (99%) Uncultured ocean crust clone EPR4055-N3-Bc85 (98%) | Potentially endogenous [66] | ||
| K17.1-3 | × | × | × | × | Delftia (99%) | Allochtonous | |
| K17.14-1 | × | × | × | ||||
| K20.28-3 | |||||||
| K20.22-1 K20.23-1 | × | Massilia (99%) | Allochtonous | ||||
| K20.24-1 | × | Uncultured β-proteobacterium clone XE2F08 (99%) | Endogenous | ||||
| K20.20-1 | × | Methylophilus (99%) | Potentially endogenous [67,68] | ||||
| γ-Proteobacteria | |||||||
| K19.17-1 | × | × | × | × | Shewanella (99%) | Endogenous [76] | |
| K17.2-3 K18P.30-2 | × | × | × | Stenotrophomonas (99%) | Allochtonous | ||
| K20P.31-2 | × | Halomonas (98%) | Endogenous [77] | ||||
| Bacteroidetes | |||||||
| K20.19-1 | × | Elizabethkingia (99%) | Allochtonous | ||||
| K18P.26-2 | × | Cloacibacterium (99%) | Allochtonous | ||||
| K19.16-1 | × | Mucilaginibacter (99%) | Allochtonous | ||||
| K17.8-2 | × | Porphyromonas (99%) | Allochtonous | ||||
| Firmicutes | |||||||
| K18P.27-2 | × | Anoxybacillus (99%) | Potentially endogenous [70] | ||||
| K20.17-3 | × | Uncultured subsurface clone HDB_SIST458 (100%) | Endogenous [71,72] | ||||
| Actinobacteria | |||||||
| K19.18-1 K18.5-3 K17.10-2 | × | × × | × × | × × | Propionibacterium (99%) | Allochtonous | |
| K17.4-3 K18.11-2 | × | × | Rhodococcus (99%) | Potentially endogenous [69] | |||
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Leblanc, V.; Hellal, J.; Fardeau, M.-L.; Khelaifia, S.; Sergeant, C.; Garrido, F.; Ollivier, B.; Joulian, C. Microbial and Geochemical Investigation down to 2000 m Deep Triassic Rock (Meuse/Haute Marne, France). Geosciences 2019, 9, 3. https://doi.org/10.3390/geosciences9010003
Leblanc V, Hellal J, Fardeau M-L, Khelaifia S, Sergeant C, Garrido F, Ollivier B, Joulian C. Microbial and Geochemical Investigation down to 2000 m Deep Triassic Rock (Meuse/Haute Marne, France). Geosciences. 2019; 9(1):3. https://doi.org/10.3390/geosciences9010003
Chicago/Turabian StyleLeblanc, Vanessa, Jennifer Hellal, Marie-Laure Fardeau, Saber Khelaifia, Claire Sergeant, Francis Garrido, Bernard Ollivier, and Catherine Joulian. 2019. "Microbial and Geochemical Investigation down to 2000 m Deep Triassic Rock (Meuse/Haute Marne, France)" Geosciences 9, no. 1: 3. https://doi.org/10.3390/geosciences9010003
APA StyleLeblanc, V., Hellal, J., Fardeau, M. -L., Khelaifia, S., Sergeant, C., Garrido, F., Ollivier, B., & Joulian, C. (2019). Microbial and Geochemical Investigation down to 2000 m Deep Triassic Rock (Meuse/Haute Marne, France). Geosciences, 9(1), 3. https://doi.org/10.3390/geosciences9010003