Fossilized Endolithic Microorganisms in Pillow Lavas from the Troodos Ophiolite, Cyprus


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
Geological Setting and Sampled Localities
2. Materials and Methods
2.1. Microscopy
2.2. Environmental Scanning Electron Microscopy
2.3. Raman Spectroscopy
2.4. Microthermometry
3. Results
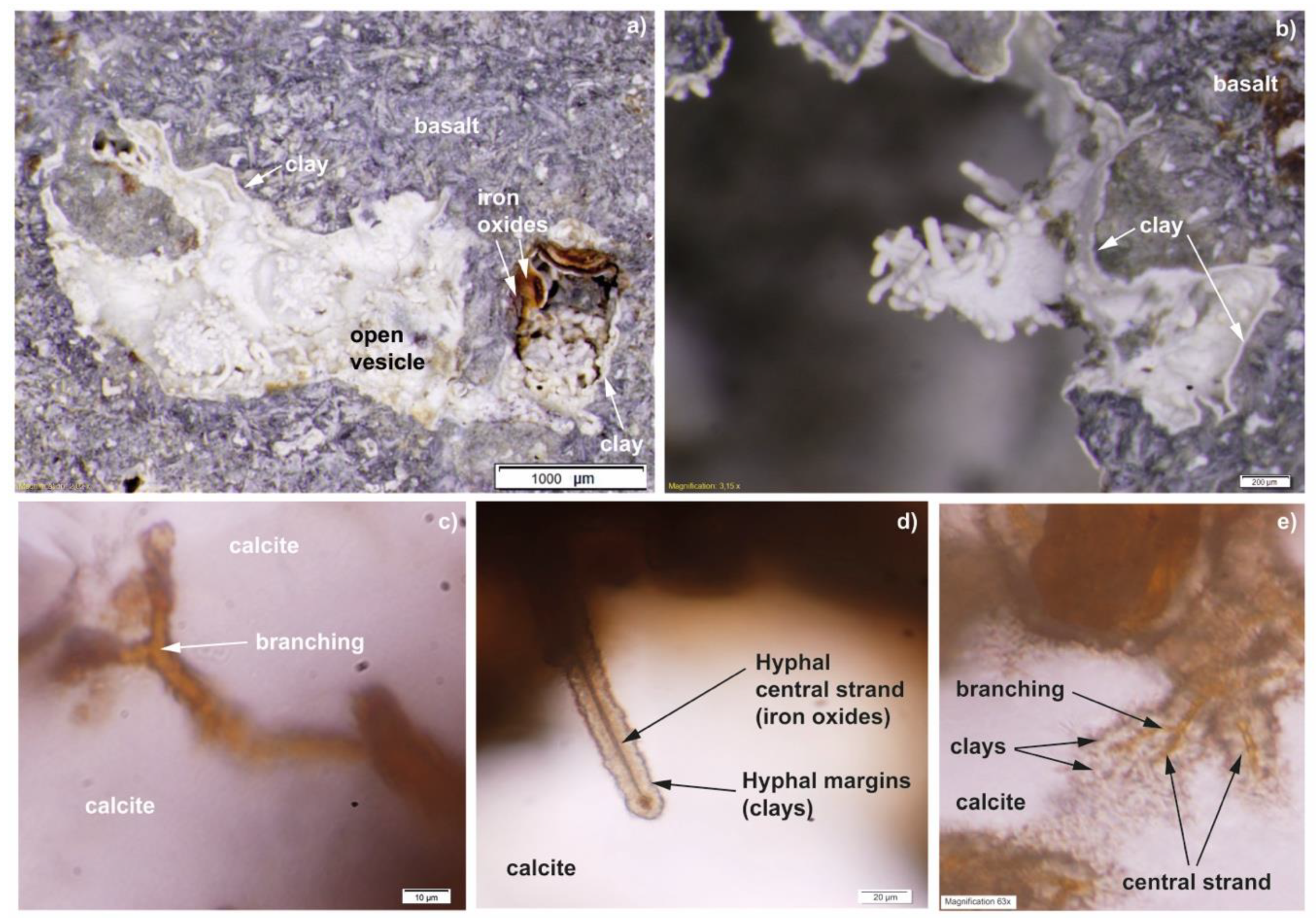
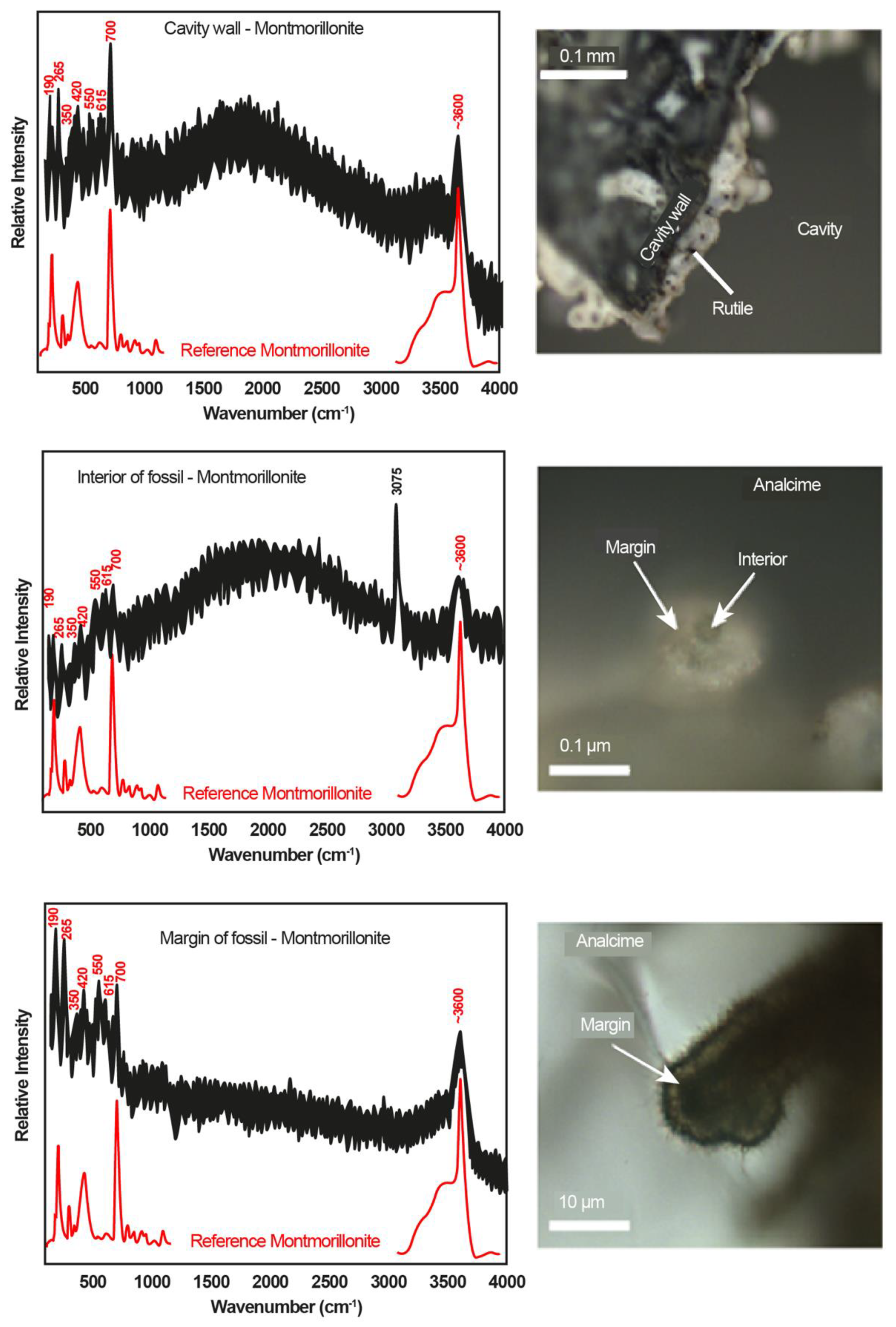
3.1. Mineralogy and Filamentous Structures
3.2. Fluid Inclusions
4. Discussion
4.1. Biogenicity
4.2. Hydrothermal Conditions of the Habitats
4.3. Mobilization of Elements
4.4. Fossilization
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Schrenk, M.O.; Huber, J.A.; Edwards, K.J. Microbial Provinces in the Subseafloor. Ann. Rev. Mar. Sci. 2009, 2, 279–304. [Google Scholar] [CrossRef] [PubMed]
- Orcutt, B.N.; Bach, W.; Becker, K.; Fisher, A.T.; Hentscher, M.; Toner, B.M.; Wheat, C.G.; Edwards, K.J. Colonization of subsurface microbial observatories deployed in young ocean crust. ISME J. 2011, 5, 692. [Google Scholar] [CrossRef] [PubMed]
- Mason, O.U.; Nakagawa, T.; Rosner, M.; Van Nostrand, J.D.; Zhou, J.; Maruyama, A.; Fisk, M.R.; Giovannoni, S.J. First Investigation of the Microbiology of the Deepest Layer of Ocean Crust. PLOS ONE 2010, 5, e15399. [Google Scholar] [CrossRef] [PubMed]
- Orcutt, B.; Wheat, C.G.; Edwards, K.J. Subseafloor Ocean Crust Microbial Observatories: Development of FLOCS (FLow-through Osmo Colonization System) and Evaluation of Borehole Construction Materials. Geomicrobiol. J. 2010, 27, 143–157. [Google Scholar] [CrossRef]
- Lever, M.A.; Rouxel, O.; Alt, J.C.; Shimizu, N.; Ono, S.; Coggon, R.M.; Shanks, W.C.; Lapham, L.; Elvert, M.; Prieto-Mollar, X.; et al. Evidence for microbial carbon and sulfur cycling in deeply buried ridge flank basalt. Science 2013, 339, 1305–1308. [Google Scholar] [CrossRef]
- Jungbluth, S.P.; Lin, H.-T.; Cowen, J.P.; Glazer, B.T.; Rappé, M.S. Phylogenetic diversity of microorganisms in subseafloor crustal fluids from Holes 1025C and 1026B along the Juan de Fuca Ridge flank. Front. Microbiol. 2014, 5, 119. [Google Scholar] [CrossRef] [Green Version]
- Ivarsson, M.; Bengtson, S.; Skogby, H.; Belivanova, V.; Marone, F. Fungal colonies in open fractures of subseafloor basalt. Geo-Marine Lett. 2013, 33, 233–243. [Google Scholar] [CrossRef] [Green Version]
- Ivarsson, M.; Peckmann, J.; Tehler, A.; Broman, C.; Bach, W.; Behrens, K.; Reitner, J.; Böttcher, M.E.; Ivarsson, L.N. Zygomycetes in Vesicular Basanites from Vesteris Seamount, Greenland Basin—A New Type of Cryptoendolithic Fungi. PLOS ONE 2015, 10, e0133368. [Google Scholar] [CrossRef]
- Schumann, G.; Manz, W.; Reitner, J.; Lustrino, M. Ancient Fungal Life in North Pacific Eocene Oceanic Crust. Geomicrobiol. J. 2004, 21, 241–246. [Google Scholar] [CrossRef]
- Bengtson, S.; Ivarsson, M.; Astolfo, A.; Belivanova, V.; Broman, C.; Marone, F.; Stampanoni, M. Deep-biosphere consortium of fungi and prokaryotes in Eocene subseafloor basalts. Geobiology 2014, 12, 489–496. [Google Scholar] [CrossRef]
- Ivarsson, M.; Bengtson, S.; Neubeck, A. The igneous oceanic crust—Earth’s largest fungal habitat? Fungal Ecol. 2016, 20, 249–255. [Google Scholar] [CrossRef]
- Ivarsson, M.; Bengtson, S.; Skogby, H.; Lazor, P.; Broman, C.; Belivanova, V.; Marone, F. A Fungal-Prokaryotic Consortium at the Basalt-Zeolite Interface in Subseafloor Igneous Crust. PLOS ONE 2015, 10, e0140106. [Google Scholar] [CrossRef] [PubMed]
- Bengtson, S.; Sallstedt, T.; Belivanova, V.; Whitehouse, M. Three-dimensional preservation of cellular and subcellular structures suggests 1.6 billion-year-old crown-group red algae. PLoS Biol. 2017, 15, e2000735. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McMahon, S.; Parnell, J. The deep history of Earth’s biomass. J. Geol. Soc. 2018, 175, 716–720. [Google Scholar] [CrossRef]
- Ivarsson, M.; Holm, N.G.; Neubeck, A. The deep biosphere of the subseafloor igneous crust. In Handbook of Environmental Chemistry; Demina, L., Galkin, S., Eds.; Springer: Cham, Switzerland, 2016; Volume 50. [Google Scholar]
- Osozawa, S.; Shinjo, R.; Lo, C.-H.; Jahn, B.-M.; Hoang, N.; Sasaki, M.; Ishikawa, K.; Kano, H.; Hoshi, H.; Xenophontos, C.; et al. Geochemistry and geochronology of the Troodos ophiolite: An SSZ ophiolite generated by subduction initiation and an extended episode of ridge subduction? Lithosphere 2012, 4, 497–510. [Google Scholar] [CrossRef]
- Furnes, H.; Muehlenbachs, K.; Tumyr, O.; Torsvik, T.; Xenophontos, C. Biogenic alteration of volcanic glass from the Troodos ophiolite, Cyprus. J. Geol. Soc. 2001, 158, 75–84. [Google Scholar] [CrossRef]
- Furnes, H.; Banerjee, N.R.; Staudigel, H.; Muehlenbachs, K.; McLoughlin, N.; De Wit, M.; Van Kranendonk, M. Comparing petrographic signatures of bioalteration in recent to Mesoarchean pillow lavas: Tracing subsurface life in oceanic igneous rocks. Precambrian Res. 2007, 158, 156–176. [Google Scholar] [CrossRef]
- Pearce, J.A.; Lippard, S.J.; Roberts, S. Characteristics and tectonic significance of supra-subduction zone ophiolites. Geol. Soc. London, Spec. Publ. 2008, 16, 77–94. [Google Scholar] [CrossRef]
- Moores, E.M.; Vine, F.J. The Troodos Massif, Cyprus and other Ophiolites as Oceanic Crust: Evaluation and Implications. Philos. Trans. R. Soc. A: Math. Phys. Eng. Sci. 1971, 268, 443–467. [Google Scholar] [CrossRef]
- Ivarsson, M. Advantages of doubly polished thin sections for the study of microfossils in volcanic rock. Geochem. Trans. 2006, 7, 5. [Google Scholar] [CrossRef]
- Downs, R.T. The RRUFF Project: An integrated study of the chemistry, crystallography, Raman and infrared spectroscopy of minerals. In Program and Abstracts of the 19th General Meeting of the International Mineralogical Association in Kobe, Japan, 2006; International Mineralogical Association: Kobe, Japan, 2006. [Google Scholar]
- Shepherd, T.J.; Rankin, A.H.; Alderton, D.H. A Practical Guide to Fluid Inclusion Studies; Chapman & Hall: London, UK, 1985; p. 239. [Google Scholar]
- Bishop, J.L.; Murad, E. Characterization of minerals and biogeochemical markers on Mars: A Raman and IR spectroscopic study of montmorillonite. J. Raman Spectrosc. 2004, 35, 480–486. [Google Scholar] [CrossRef]
- Wang, A.; Freeman, J.J.; Jolliff, B.L. Understanding the Raman spectral features of phyllosilicates. J. Raman Spectrosc. 2015, 46, 829–845. [Google Scholar] [CrossRef]
- Charpentier, D.; Buatier, M.; Jacquot, E.; Gaudin, A.; Wheat, C. Conditions and mechanism for the formation of iron-rich Montmorillonite in deep sea sediments (Costa Rica margin): Coupling high resolution mineralogical characterization and geochemical modeling. Geochim. Cosmochim. Acta 2011, 75, 1397–1410. [Google Scholar] [CrossRef] [Green Version]
- Orange, D.; Knitile, E.; Farber, D.; Williams, Q. Raman spectroscopy of crude oils and hydrocarbon fluid inclusions: A feasibility study. Miner. Spectrosc. 1996, 5, 65–81. [Google Scholar]
- Lin-Vien, D.; Colthup, N.B.; Fateley, W.G.; Grasselli, J.G.; Lin-Vien, D.; Colthup, N.B.; Fateley, W.G.; Grasselli, J.G. The Handbook of Infrared and Raman Characteristic Frequencies of Organic Molecules; Elsevier Science: Amsterdam, The Netherlands, 1991; ISBN 0080571166, 9780080571164. [Google Scholar]
- Freeman, J.J.; Wang, A.; Kuebler, K.E.; Jolliff, B.L.; Haskin, L.A. Characterization of Natural Feldspars by Raman Spectroscopy for Future Planetary Exploration. Can. Miner. 2008, 46, 1477–1500. [Google Scholar] [CrossRef]
- Frogner, P.; Broman, C.; Lindblom, S. Weathering detected by Raman spectroscopy using AI-ordering in albite. Chem. Geol. 1998, 151, 161–168. [Google Scholar] [CrossRef]
- Frost, R.L.; Lopez, A.; Theiss, F.L.; Romano, A.W.; Scholz, R. A vibrational spectroscopic study of the silicate mineral analcime—Na2(Al4SiO4O12)·2H2O—A natural zeolite. Spectrochim. Acta Part A: Mol. Biomol. Spectrosc. 2014, 133, 521–525. [Google Scholar] [CrossRef]
- Burkhard, M. Calcite twins, their geometry, appearance and significance as stress-strain markers and indicators of tectonic regime: a review. J. Struct. Geol. 1993, 15, 351–368. [Google Scholar] [CrossRef] [Green Version]
- Bodnar, R.J. Fluid Inclusions—Analysis and Interpretation; Anderson, A., Marshall, D., Eds.; Mineralogical Association of Canada: Quebec City, QC, Canada, 2003. [Google Scholar]
- Gibson, E.; McKay, D.; Thomas-Keprta, K.; Wentworth, S.; Westall, F.; Steele, A.; Romanek, C.; Bell, M.; Toporski, J. Life on Mars: evaluation of the evidence within Martian meteorites ALH84001, Nakhla, and Shergotty. Precambrian Res. 2001, 106, 15–34. [Google Scholar] [CrossRef]
- Buick, R. Microfossil Recognition in Archean Rocks: An Appraisal of Spheroids and Filaments from a 3500 M.Y. Old Chert-Barite Unit at North Pole, Western Australia. Palaios 1991, 5, 441–459. [Google Scholar] [CrossRef]
- Schopf, J.W.; Walter, M.R. Archean microfossils: New evidence of ancient microorganisms. In Earth’s Earliest Biosphere, Its Origin and Evolution; Schopf, J.W., Ed.; Princeton University Press: Princeton, NJ, USA, 1983. [Google Scholar]
- Ivarsson, M.; Bengtson, S.; Belivanova, V.; Stampanoni, M.; Marone, F.; Tehler, A. Fossilized fungi in subseafl oor Eocene basalts. Geology 2012, 40, 163–166. [Google Scholar] [CrossRef]
- Peckmann, J.; Bach, W.; Behrens, K.; Reitner, J. Putative cryptoendolithic life in Devonian pillow basalt, Rheinisches Schiefergebirge, Germany. Geobiology 2008, 6, 125–135. [Google Scholar] [CrossRef] [PubMed]
- Eickmann, B.; Bach, W.; Kiel, S.; Reitner, J.; Peckmann, J. Evidence for cryptoendolithic life in Devonian pillow basalts of Variscan orogens, Germany. Palaeogeogr. Palaeoclim. Palaeoecol. 2009, 283, 120–125. [Google Scholar] [CrossRef]
- Orcutt, B.N.; Sylvan, J.B.; Knab, N.J.; Edwards, K.J. Microbial Ecology of the Dark Ocean above, at, and below the Seafloor†. Microbiol. Mol. Boil. Rev. 2011, 75, 361–422. [Google Scholar] [CrossRef]
- Pirajno, F. Hydrothermal Processes and Mineral Systems. Hydrothermal Processes and Mineral Systems 2009.
- Ivarsson, M.; Bach, W.; Broman, C.; Neubeck, A.; Bengtson, S. Fossilized Life in Subseafloor Ultramafic Rocks. Geomicrobiol. J. 2018, 35, 460–467. [Google Scholar] [CrossRef]
- Schulz, H.N.; Jørgensen, B.B. Big Bacteria. Ann. Rev. Microbiol. 2002, 55, 105–137. [Google Scholar] [CrossRef]
- Webster, J.; Weber, R. Introduction to Fungi, 3rd ed.; Cambridge University Press: Cambridge, UK, 1981; ISBN 9780521807395. [Google Scholar]
- Goodfellow, M.; Kämpfer, P.; Busse, H.J.; Trujillo, M.E.; Suzuki, K.I.; Ludwig, W.; Whitman, W.B. (Eds.) Busse Bergey’s Manual of Systematic Bacteriology; Springer: New York, NJ, USA, 2012; Volume 5. [Google Scholar]
- Crosby, C.H.; Bailey, J.V. The role of microbes in the formation of modern and ancient phosphatic mineral deposits. Front. Microbiol. 2012, 3, 241. [Google Scholar] [CrossRef] [Green Version]
- Sindhu, S.S.; Phour, M.; Choudhary, S.R.; Chaudhary, D. Phosphorus Cycling: Prospects of Using Rhizosphere Microorganisms for Improving Phosphorus Nutrition of Plants. In Geomicrobiology and Biogeochemistry; Springer: Berlin, Germany, 2014; pp. 199–237. [Google Scholar]
- Raven, P.; Johnson, G.; Mason, K.; Losos, J.; Singer, S. Biology, 11th ed.; McGraw-Hill: New York, NY, USA, 2017; ISBN 1259188132, 9781259188138. [Google Scholar]
- Fru, E.C.; Hemmingsson, C.; Holm, M.; Chiu, B.; Iñiguez, E. Arsenic-induced phosphate limitation under experimental Early Proterozoic oceanic conditions. Earth Planet. Sci. Lett. 2016, 434, 52–63. [Google Scholar] [CrossRef]
- Candela, P.A. Ores in the Earth’s Crust. In The Crust; Rudick, R.L., Ed.; Elsevier Science: Amsterdam, The Netherlands, 2005; p. 702. [Google Scholar]
- Spooner, E.T.C. Cu-pyrite mineralization and seawater convection in oceanic crust-the ophiolitic ore deposits of Cyprus. Geol. Assoc. Can. 1980, 20, 685–704. [Google Scholar]
- Cann, J.; Gillis, K. Hydrothermal insights from the Troodos Ophiolite, Cyprus. In Hydrogeology of the Oceanic Lithosphere; Cambridge University Press: New York, NY, USA, 2004; pp. 272–310. [Google Scholar]
- Pálmason, G.; Arnórsson, S.; Fridleifsson, I.B.; Kristmannsdóttir, H.; Saemundsson, K.; Stefánsson, V.; Steingrímsson, B.; Tómasson, J.; Kristjánsson, L. The Iceland crust: Evidence from drillhole data on structure and processes. Deep Drill. Results Atl. Ocean: Ocean Crust 2011, 2, 43–65. [Google Scholar]
- Odin, G. Introduction to the Study of Green Marine Clays. Dev. Sedimentol. 1988, 45, 1–3. [Google Scholar]
- Beyssac, O.; Rouzaud, J.-N.; Goffé, B.; Brunet, F.; Chopin, C. Graphitization in a high-pressure, low-temperature metamorphic gradient: a Raman microspectroscopy and HRTEM study. Contrib. Miner. Pet. 2002, 143, 19–31. [Google Scholar] [CrossRef]
- Beyssac, O.; Goffé, B.; Chopin, C.; Rouzaud, J.N. Raman spectra of carbonaceous material in metasediments: a new geothermometer. J. Metamorph. Geol. 2002, 20, 859–871. [Google Scholar] [CrossRef]
- Huang, E.-P.; Huang, E.; Yu, S.-C.; Chen, Y.-H.; Lee, J.-S.; Fang, J.-N. In situ Raman spectroscopy on kerogen at high temperatures and high pressures. Phys. Chem. Miner. 2010, 37, 593–600. [Google Scholar] [CrossRef]
- Pimenta, M.A.; Dresselhaus, G.; Dresselhaus, M.S.; Cançado, L.G.; Jorio, A.; Saito, R. Studying disorder in graphite-based systems by Raman spectroscopy. Phys. Chem. Chem. Phys. 2007, 9, 1276–1290. [Google Scholar] [CrossRef]
- Maheshwari, R.; Bharadwaj, G.; Bhat, M.K. Thermophilic Fungi: Their Physiology and Enzymes. Microbiol. Mol. Biol. Rev. 2003, 64, 461–488. [Google Scholar] [CrossRef]
- Ivarsson, M.; Broman, C.; Lindblom, S.; Holm, N. Fluid inclusions as a tool to constrain the preservation conditions of sub-seafloor cryptoendoliths. Planet. Space Sci. 2009, 57, 477–490. [Google Scholar] [CrossRef]
- Stakes, D.S.; Scheidegger, K.F. Temporal variations in secondary minerals from Nazca plate basalts, diabases, and microgabbros. From Rodinia to Pangea: The Lithotectonic Record of the Appalachian Region 1981, 154, 109–130. [Google Scholar]
- Alt, J.C.; Honnorez, J. Alteration of the upper oceanic crust, DSDP site 417: mineralogy and chemistry. Contrib. Miner. Pet. 1984, 87, 149–169. [Google Scholar] [CrossRef]
- Robb, L. Introduction to Ore-Forming Processes; John Wiley & Sons: Hoboken, NJ, USA, 2005; ISBN 0632063785. [Google Scholar]
- Henin, S. Synthesis of Clay Minerals at Low Temperatures. Clays Clay Miner. 2006, 4, 54–60. [Google Scholar] [CrossRef]
- Mariner, R.H.; Surdam, R.C. Alkalinity and Formation of Zeolites in Saline Alkaline Lakes. Science 2006, 170, 977–980. [Google Scholar] [CrossRef]
- Savage, D.; Rochelle, C.; Moore, Y.; Milodowski, A.; Bateman, K.; Bailey, D.; Mihara, M. Analcime reactions at 25–90 °C in hyperalkaline fluids. Miner. Mag. 2001, 65, 571–587. [Google Scholar] [CrossRef]
- Gudbrandsson, S.; Wolff-Boenisch, D.; Gislason, S.R.; Oelkers, E.H. An experimental study of crystalline basalt dissolution from 2⩽pH⩽11 and temperatures from 5 to 75 °C. Geochim. Cosmochim. Acta 2011, 75, 5496–5509. [Google Scholar] [CrossRef]
- Rousk, J.; Brookes, P.C.; Bååth, E. Contrasting Soil pH Effects on Fungal and Bacterial Growth Suggest Functional Redundancy in Carbon Mineralization. Appl. Environ. Microbiol. 2009, 75, 1589–1596. [Google Scholar] [CrossRef] [Green Version]
- Reitner, J.; Schumann, G.; Pedersen, K. Fungi in subterranean environments. In Fungi in Biogeochemical Cycles; Cambridge University Press: Cambridge, UK, 2006; p. 377. ISBN 9780511550522. [Google Scholar]
- Benson, S.A. Chapter 2 The bacterial outer membrane and surface permeability. Dev. Biol. 1998, 9, 15–22. [Google Scholar]
- Rodríguez-Navarro, A. Potassium transport in fungi and plants. Biochim. Biophys. Acta 2000, 1469, 1–30. [Google Scholar] [CrossRef]
- Gadd, G.M. Geomycology: biogeochemical transformations of rocks, minerals, metals and radionuclides by fungi, bioweathering and bioremediation. Mycol. Res. 2007, 111, 3–49. [Google Scholar] [CrossRef]
- Gadd, G.M. Metals, minerals and microbes: Geomicrobiology and bioremediation. Microbiology 2010, 156, 609–643. [Google Scholar] [CrossRef]
- Vaughan, D.J.; Pattrick, R.A.D.; Wogelius, R.A. Minerals, metals and molecules: ore and environmental mineralogy in the new millennium. Miner. Mag. 2002, 66, 653–676. [Google Scholar] [CrossRef]
- Brown, G.E.; Trainor, T.P.; Chaka, A.M. Geochemistry of Mineral Surfaces and Factors Affecting Their Chemical Reactivity. In Chemical Bonding at Surfaces and Interfaces; Elsevier: Amsterdam, The Netherlands, 2008; pp. 457–509. [Google Scholar]
- Bernal, J.D. The Physical Basis of Life; Routledge and Paul: London, UK, 1951; p. 597. [Google Scholar]
- Clarkson, E. Genetic Takeover and the Mineral Origins of Life. By A. G. Cairns-Smith (first published 1982, first paperback edition 1987). Cambridge University Press. ISBN 0 521 34682 7. Genet. Res. 1988, 51, 167–168. [Google Scholar] [CrossRef]
- Hashizume, H. Role of Clay Minerals in Chemical Evolution and the Origins of Life. In Clay Minerals in Nature—Their Characterization, Modification and Application; IntechOpen: London, UK, 2012. [Google Scholar] [Green Version]
- Smith, S.; Read, D. Mycorrhizal Symbiosis, 3rd ed.; Academic Press: Cambridge, MA, USA, 2008; ISBN 9780123705266. [Google Scholar]









© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Carlsson, D.-T.; Ivarsson, M.; Neubeck, A. Fossilized Endolithic Microorganisms in Pillow Lavas from the Troodos Ophiolite, Cyprus. Geosciences 2019, 9, 456. https://doi.org/10.3390/geosciences9110456
Carlsson D-T, Ivarsson M, Neubeck A. Fossilized Endolithic Microorganisms in Pillow Lavas from the Troodos Ophiolite, Cyprus. Geosciences. 2019; 9(11):456. https://doi.org/10.3390/geosciences9110456
Chicago/Turabian StyleCarlsson, Diana-Thean, Magnus Ivarsson, and Anna Neubeck. 2019. "Fossilized Endolithic Microorganisms in Pillow Lavas from the Troodos Ophiolite, Cyprus" Geosciences 9, no. 11: 456. https://doi.org/10.3390/geosciences9110456
APA StyleCarlsson, D. -T., Ivarsson, M., & Neubeck, A. (2019). Fossilized Endolithic Microorganisms in Pillow Lavas from the Troodos Ophiolite, Cyprus. Geosciences, 9(11), 456. https://doi.org/10.3390/geosciences9110456