3.1. Macro- and Micromineral Contents of Yellow-Fleshed Cassava Roots Across Genotypes, Locations, and Sets 1 and 2
The macro- and micromineral contents of yellow-fleshed cassava root across all the genotypes and locations are presented in
Table 1. The most abundant mineral in cassava storage roots was potassium (K), with concentrations ranging from 4300 to 26,000 mg/kg with an overall mean of 12,116 ± 3543.58 mg/kg for trial 1, and ranged from 6300 to 38,000 mg/kg with a mean value of 17,883.60 ± 4898.77 mg/kg for trial 2. However, the least abundant mineral was sodium (Na), with concentrations ranging from 15.68 to 2100 mg/kg with an overall mean of 228.22 ± 316.74 mg/kg for trial 1, and from 11.47 to 186.73 mg/kg with a total mean value of 39.19 ± 23.41 mg/kg for trial 2. The results also showed that trial 2 had higher concentrations of calcium (Ca), magnesium (Mg), and potassium (K) than trial 1. Still, trial 1 sets had a higher content of Na, P (phosphorous), and S (sulphur). It could be deduced that the storage roots of cassava could be a good source of these macroelements, especially potassium. Diets rich in minerals are essential for proper growth and development [
11]. This result agrees with Aro et al. [
16], who reported that the highest mineral content of the cassava samples studied was K (269 mg/kg), but contradicts with Bamidele et al. [
17]. Among the mineral compositions of the four cassava samples they studied, Mg was the highest mineral found in all four cassava samples investigated, with values ranging between 185.1 and 321.5 mg/kg. Manano et al. [
18] studied some local and improved high-yielding and cassava mosaic disease (CMD)-resistant cassava varieties grown in Uganda. The findings of this study indicated that the levels of minerals differed with the cassava variety. The cassava yellow root genotypes for both trials and 1 and 2 had very low Na, and this indicates that these genotypes could form part of a low-sodium diet. However, K regulates the osmotic pressure within the cell, and is involved in cell membrane transport and the activation of several glycolytic and respiratory enzymes. Potassium intake in a regular diet is 2–5.9 g/day. The minimum daily requirement is estimated to be 782 mg. It implies that cassava, with a high level of K and low Na, might be an ideal crop to consume to reduce hypertension, although this needs further research evidence.
The most abundant mineral in cassava storage roots was manganese (Mn), with concentrations ranging from 7.390 to 39.990 mg/kg with an overall mean of 16.511 ± 6.200 mg/kg for trial 1, and ranging from 4.540 to 42.680 mg/kg with a mean value of 13.383 ± 4.698 mg/kg for trial 2. However, the least abundant minerals were molybdenum (Mo) and cobalt (Co), with concentrations ranging from 0.600 to 0.700 mg/kg with an overall mean of 0.694 ± 0.025 and 0.675 ± 0.043 mg/kg for trial 1, and ranging from 0.600 to 3.000 mg/kg with total mean values of 0.708 ± 0.237 and 0.696 ± 0.239 mg/kg for trial 2, respectively. The results also showed that all the minerals in trial 2 had higher concentrations than trial 1, as found for macrominerals. It could be deduced that the storage roots of cassava could be a good source of these microelements, especially Mn.
3.2. Mean Squares (MS) of Macro- and Microelements of Yellow-Fleshed Cassava Root
The effects of genotypes, maturity, and method of sampling on the macro- and microelements of yellow-fleshed cassava roots are presented in
Table 2 and
Table 3. For trial 1, the ANOVA showed that genotype, method, and genotype × maturity interaction had a strong significant (
p < 0.001) effect on all the macrominerals studied. However, maturity had no significant effect (
p > 0.05) on Mg, Na, K, and S. Genotype × maturity interaction had a significant effect (
p < 0.001) on all the minerals except for Ca. Only Mg, S, and Na showed a significant difference (
p < 0.001) for genotype × method interaction. For maturity × method interaction, only Ca and Na showed a significant difference at (
p < 0.001). For the data set 2 trial, there was a highly significant effect (
p < 0.001) of genotype and genotype × maturity interaction on all the macroelements. Maturity and method had no significant effect (
p < 0.05) on Mg and Ca, respectively. Though, genotype × method interaction had a significant effect (at
p < 0.001) only on Na; maturity × method interaction showed no significant effect (
p < 0.05) across all the minerals investigated. Thus, for both trials 1 and 2, the independent variables (genotype and maturity) explained the variations observed for all the dependent variables (Ca, Mg, Na, K, P, and S) rather than the basic mean.
Table 4 showed the effects of genotype, maturity, and method of sampling on the microelements of yellow-fleshed cassava roots. For trial 1, the ANOVA showed that genotype, maturity, method, genotype × maturity interaction, genotype × method interaction, and maturity × method interaction had a strong significant effect (
p < 0.001) on all the minerals investigated except Al. However, genotype × method interaction had no significant effect (
p > 0.05) on all the minerals except for Ni. From the dataset 2 trial, the ANOVA showed that genotype, maturity, method, genotype × maturity interaction, genotype × method interaction, and maturity × method interaction had a strong significant effect (
p < 0.001) on all the minerals investigated except Co. However, the genotype × method interaction had no significant effect (
p < 0.05) on all the minerals investigated. The study also revealed that maturity × method interaction had a significant difference (
p < 0.001) on Mo, Ni, and Zn. Both maturity and genotype × maturity interaction had a significant difference at (
p < 0.001) on Fe, Mg, B, Ni, Zn, and Al. Thus, for both trials 1 and 2, the independent variables (genotype and maturity) explained the variations observed for all the dependent variables (Ca, Mg, Na, K, P, and S) than the basic mean.
3.3. Macromineral Concentrations of Yellow-Fleshed Cassava Roots by Genotype
The means of the macromineral concentrations of yellow cassava roots (trial set 1) by genotype are shown in
Table 4. For trial 1, significant differences (
p < 0.05) existed amongst the means of the macroelements for all the genotypes. However, genotypes 01/1224 (2033 mg/kg) and 01/1412 (1742.3 mg/kg) had the highest Ca concentrations among the samples investigated. The lowest Ca value (596.3 mg/kg) was found for TME1, the check variety. Genotypes 01/1368 (676.2 mg/kg) and 90/01554 (650.5 mg/kg) had the highest levels of Na in all the samples examined. However, the Na concentrations were found to be the lowest in genotype 94/0330 (44 mg/kg). For K, considerably higher concentrations were recorded in genotype 01/1662 (20160 mg/kg). However, the lowest K values were recorded in genotype 94/3030 (8206.7 mg/kg). The highest concentrations for P were recorded in genotype 01/1335 (1993.7 mg/kg). However, the P concentration was considerably lower in the 01/1371 (813.3 mg/kg), 01/1412 (807.7 mg/kg), 01/1413 (847.7 mg/kg), and 01/1442 (801 mg/kg) genotypes.
From the results for trial 2 in
Table 5, the study showed that genotype 01/1417 (2833.3 mg/kg) had the highest Ca content of all genotypes investigated. However, genotypes 01/1181 and 01/1206 showed a significantly higher Ca content than 30,572 (check) and 91/02324 (check). Although the highest Mg content was recorded in genotype 01/1417 (1679.7 mg/kg), its concentration was found to be the lowest in genotypes 01/1231 (753.7 mg/kg) and 01/1172 (721.7 mg/kg) from all the genotypes examined. The highest levels of Na content were recorded in genotype 01/1417 (73.9 mg/kg). The study revealed that the levels of K content in genotype 01/1206 (23,000 mg/kg) were higher than the rest of the genotypes. However, genotype 99/7558 (12,680 mg/kg) had the lowest levels of all the genotypes examined. For P, the study showed the highest concentration in the Z97/0474 (12,680 mg/kg) genotype of all the samples examined. However, genotypes 99/7558 (760 mg/kg) and 01/1296 (763.7 mg/kg) had the lowest levels of P content of all the genotypes. Genotype Z97/0474 (358 mg/kg) showed the highest levels of S content of all the samples studied. From the results above, all the genotypes with significantly higher mineral concentrations than the check genotypes could be recommended for breeding purposes. However, genotypes with no significant difference or significantly lower mineral concentrations than the check genotypes could not be recommended for such purposes as breeding.
From these results, it could be inferred that most of the genotypes had higher levels of mineral density than the check genotypes, and they could be advanced to the next stage in the breeding program. Additionally, there were significant differences in the macroelement concentrations among the genotypes. Most of the values obtained for the macroelements for all the genotypes compared well with what most researchers reported. Chiwona-Karltun et al. [
19] reported the macroelement content of the nine cassava varieties as follows: Potassium content ranged from 6810 to 12,200 mg/kg, and sodium ranged from 1720 (Nalumino variety) to 4590 mg/kg (Bangweulu variety). Otache et al. [
20] reported that the macroelements of the peel of the three cassava cultivars studied was a calcium content of 137.4 to 198.1 mg/kg), sodium of 137.0 to 142.3 mg/kg, magnesium of 128.0 to 148.1 mg/kg, and potassium of 787.2 to 1239.8 mg/kg. Besides, the authors also corroborated the finding from this study that cassava roots are rich in macroelements, especially potassium. However, the values of macroelements reported by Afoakwa et al. [
21] for six varieties obtained from the Crop Research Institute of Council for Scientific and Industrial Research (CSIR) in Ghana were lower than those obtained in this study and reported. The reported calcium content ranged from 0.06 to 1.60 mg 100 g
−1, sodium ranged from 0.25 to 0.37 mg 100 g
−1, magnesium content ranged from 1.35 to 2.52 mg 100 g
−1, and the potassium content ranged from 0.25 to 0.36 mg 100 g
−1. The difference in the values could be due to the genotypic differences and the environment. This study used yellow-fleshed genotypes, but most of the authors have used white-fleshed varieties. This implies that yellow-fleshed varieties are rich in macroelements compared with white-fleshed varieties. Additionally, it was observed in this study that most of the genotypes had the lowest sodium content among the macroelements, which was not in agreement with what some authors (Aro et al. [
16]; Afoakwa et al. [
21]; Otache et al. [
20]) reported for white-fleshed varieties. This could also be due to genetic variation in terms of yellow-fleshed versus white-fleshed varieties. However, Chavez et al. [
22] and Manano et al. [
17] reported sodium as the lowest macroelement in cassava roots, as found in this study, which reported a sodium content of 3.6 to 3.9 mg 100 g
−1.
3.4. Micromineral Concentrations of Yellow Cassava Roots by Genotype
The means of the microminerals concentrations of yellow cassava roots (Trial 1) by genotype are shown in
Table 6. There were significant differences (
p 0.05) on the levels of minerals that existed amongst the cassava varieties. Genotype 01/1235 (13.033 mg/kg) had the highest Fe concentration compared to the rest of the samples of cassava genotypes investigated. However, the Fe contents were the lowest in the check genotype 91/02324 (5.637 mg/kg). On the other hand, its concentrations were significantly higher in genotypes 01/1662, 01/1412, and 01/1610 compared to those found in the check genotypes (TME, 91/02324, and 30,572). Although genotypes 01/1662 (24.414 mg/kg), 01/1273 (24.800 mg/kg), and 01/1335 (24.823 mg/kg) had the highest Mn contents, its concentration was found to be the lowest in genotypes 95/03379 (9.544 mg/kg). On the other hand, genotypes 01/1224, 01/1663, 90/01554, 98/2135, 94/0330, 01/1413, 01/1331, 01/1277, and 01/1235 showed significantly higher contents of Mn than all the check genotypes. Genotype 01/1335 (2.698 mg/kg) showed the highest levels of B as compared to the rest of the samples examined. Its concentration was also found to be significantly higher in genotypes 01/1662, 01/1273, 01/1277, 01/1331, 01/1442, 01/1649, 90/01554, and 95/0379. However, the rest of the genotypes showed the least concentrations of B. Genotype 01/1662 (5.504 mg/kg) showed the highest levels of Cu as compared to the rest of the cassava genotypes investigated. However, its contents were the lowest in genotype 01/1235 (1.745 mg/kg). On the other hand, Cu contents were found to be significantly higher in genotype 98/2132. All the genotypes had significant amounts of Co and were significantly the same with the checks. For Ni, the highest levels were recorded in genotype 01/1273 (3.449 mg/kg). However, its levels were the lowest in genotype 01/1412 (1.449 mg/kg). On the other hand, genotype 01/1335 showed significantly higher levels of Ni than what was found in the checks. Zinc (Zn) had the highest levels in genotype 01/1115 (11.523 mg/kg). The results of the study also showed that genotypes 01/1273, 01/1442, 95/0379, 01/1662, and 01/1371 had significantly higher levels of Zn than those found in check genotypes.
The means of the micromineral concentrations of yellow cassava roots (Trial 2) by genotype are shown in
Table 7. There were significant differences in the microelement concentrations among the genotypes. Genotype 00/0028 (14.851 mg/kg) had the highest Fe concentration as compared to the rest of the cassava genotypes investigated. However, the Fe contents were found to be the lowest in the check genotype 91/02324 (5.539 mg/kg). Additionally, genotype 00/0028 (21.261 mg/kg) had the highest Mn content, while its concentration was found to be the lowest in genotypes 99/7558 (8.850 mg/kg) and 01/1172 (8.563 mg/kg). The study also revealed that genotypes 00/0028 (3.418 mg/kg) and 01/1181 (3.366 mg/kg) had the highest contents of B than the rest of the genotypes examined. However, its lowest concentrations were recorded in genotypes 01/1423 (2.105 mg/kg), 99/7558 (2.114 mg/kg), 01/1231 (2.106 mg/kg), 01/1551 (2.106 mg/kg), and check 91/0232 (2.107 mg/kg). For Cu, the highest concentrations were recorded in the check genotype 30,572 (8.131 mg/kg). However, its contents were the lowest in genotypes 01/1296 (3.061 mg/kg) and 01/1172 (3.326 mg/kg). All the genotypes recorded significant contents of both Mo and Co and were significantly the same with the checks. Nickel (Ni) had the highest concentration in genotype 00/0028 (3.325 mg/kg). All the genotypes showed significantly higher levels of the mineral concentration than the check genotypes, which are white-fleshed. Thus, the yellow-fleshed roots are richer in minerals than the white-fleshed roots. Besides, the genotypes that showed higher levels of mineral density than the check genotypes could be advanced to the next stage in the breeding program. Most of the values obtained for the microelements for all the genotypes compared well with what most researchers reported. Burns et al. [
23] found that the concentrations of iron in tubers from all cultivars studied (range from 8–24 mg/kg) were at the low end of the published range for cassava (3−140 mg/kg). In contrast, the range of tuber zinc concentrations, 8−19 mg/kg, straddled the published standard for cassava of 14 mg/kg [
23].
3.5. Effect of Maturity on the Macroelement and Microelement Contents of Yellow-Fleshed Cassava Roots
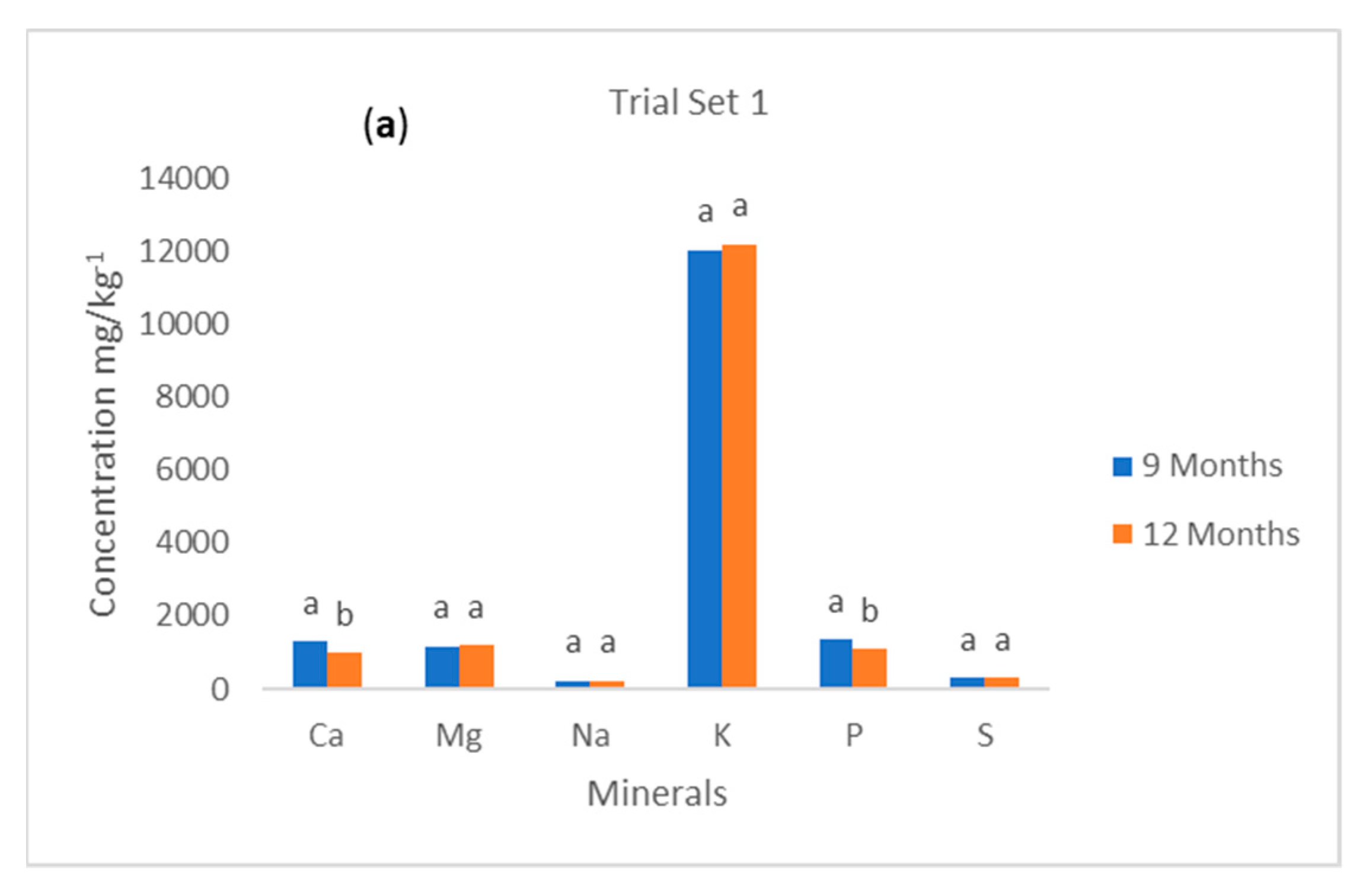
Figure 1a,b shows the concentrations of macrominerals in yellow-fleshed cassava roots by maturity (harvesting time) for both trials 1 and 2. For trial set 1, the study showed that there was a significant difference (
p < 0.05) in the mean values for the Ca and P contents in cassava roots harvested at 9 and 12 months, respectively. However, the concentrations of Ca and P were higher in the cassava roots that were harvested at the age of 9 months than those at 12 months. Thus, harvesting time had a strong influence on the Ca and P content of cassava roots. However, there was no significant difference (
p > 0.05) in the concentrations of Mg, Na, K, and S of cassava genotypes harvested at 9 and 12 months. For trial 2, the study showed that there was a significant difference (
p < 0.05) between the mean concentrations of Ca, Na, K, S, and P for cassava roots harvested at 9 and 12 months. However, the Mg concentration showed no difference in the mean value for cassava roots harvested at both maturity stages. The concentration of Ca, Na, K, and P were higher in the cassava roots that were sampled at 9 months than those at 12 months except for S, which was higher in the samples that matured at 12 months. It implies that harvesting time plays a major role in the concentration level of some macroelements in cassava roots while showing no effect on some. Additionally, it was established that harvesting time showed different effects on the two trials investigated. Thus, genotype plays a key role in the degree of the effects the harvesting time had on the macroelement concentrations of cassava roots. Richardson [
23] evaluated the macroelements of cassava sample materials that were harvested at nine months after planting and reported that the values of sodium ranged from 22.3 to 34.9 mg 100 g
−1. In contrast, the values for potassium ranged from 817.3 to 1301.2 mg 100 g
−1 [
24]. If compared with what was obtained in this study for cassava genotypes harvested at 9 months for both trials studied, the values of potassium in this study were higher and those of sodium lower. The difference could be due to the different genetic make-up of yellow-fleshed and white-fleshed cassava roots. This further confirmed that yellow-fleshed roots contain more macroelements than the white-fleshed roots, especially K. However, the values of the macroelements obtained at 12 months for trial 2 compared well with what Chavez et al. [
22] reported for cassava roots harvested at 10 to 11 months. He reported that samples had an average magnesium content of 1153 mg/kg, sodium averaged at 66.4 mg/kg, potassium averaged at 8903 mg/kg, phosphorus averaged at 1284 mg/kg, and sulphur averaged at 273 mg/kg dry weight. However, the macroelement values for trial 1 at 12 months showed higher values than what was reported in the literature [
22].
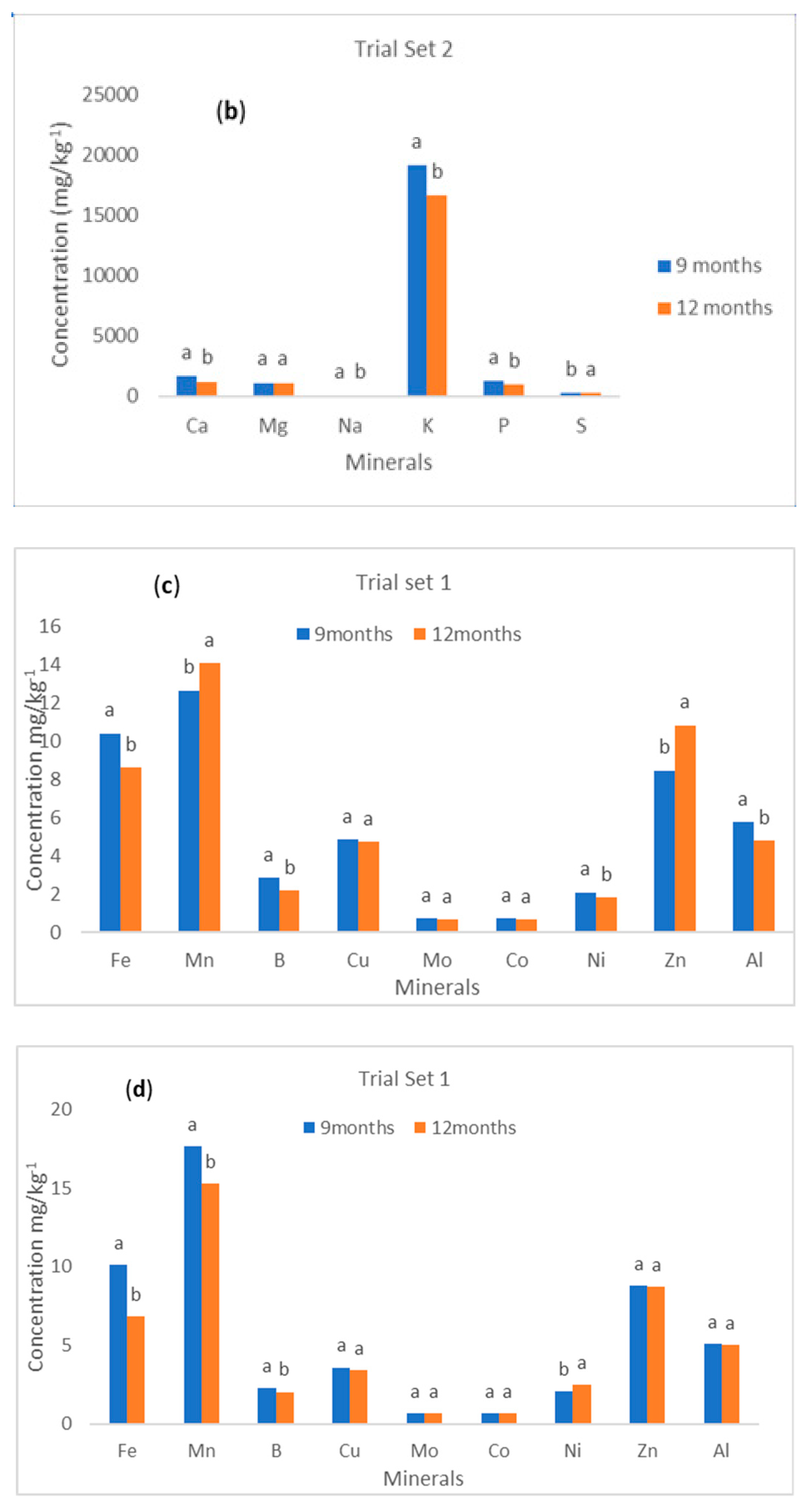
Figure 1c,d shows the levels of microelements in yellow-fleshed cassava roots by maturity (harvesting time) for both trial sets 1 and 2. For trial set 1, the study showed that there was a significant difference (
p < 0.05) in the concentration of Fe, Mn, B, and Ni at different maturity levels. For Fe, Mn, and B, higher concentrations were found in cassava roots that were harvested at 9 months. However, higher contents of Ni were recorded in cassava roots that were harvested at 12 months. The results of the study also revealed that there was no significant difference (
p > 0.05) in the contents of Cu, Mo, Co, Zn, and Al, regardless of the maturity stage. Thus, harvesting time had a strong influence on the contents of the minerals in cassava roots. For trial set 2, the study showed that there was a significant difference (
p < 0.05) in the concentration of Fe, Mn, B, Ni, Zn, and Al at different maturity stages. The concentrations of Fe, B, Ni, and Al were found to be higher in the cassava roots that were harvested at 9 months. However, Mn and Zn contents were higher in cassava roots that were harvested at 12 months. The study further revealed that there was no significant difference (
p > 0.05) in the contents of Cu, Mo, and Co regardless of the maturity stage. It implies that harvesting time plays a significant role in the concentration level of some microelements in the cassava roots while showing no effect on some. Additionally, it was established that harvesting time showed different effects on the two set trials investigated. Thus, genotype plays a vital role in the degree of effects the harvesting time showed on the microelement’s concentrations of cassava roots.
3.6. Effect of the Method of Sampling on the Distribution of the Macro- and Microelement Contents of Yellow-Fleshed Cassava Roots
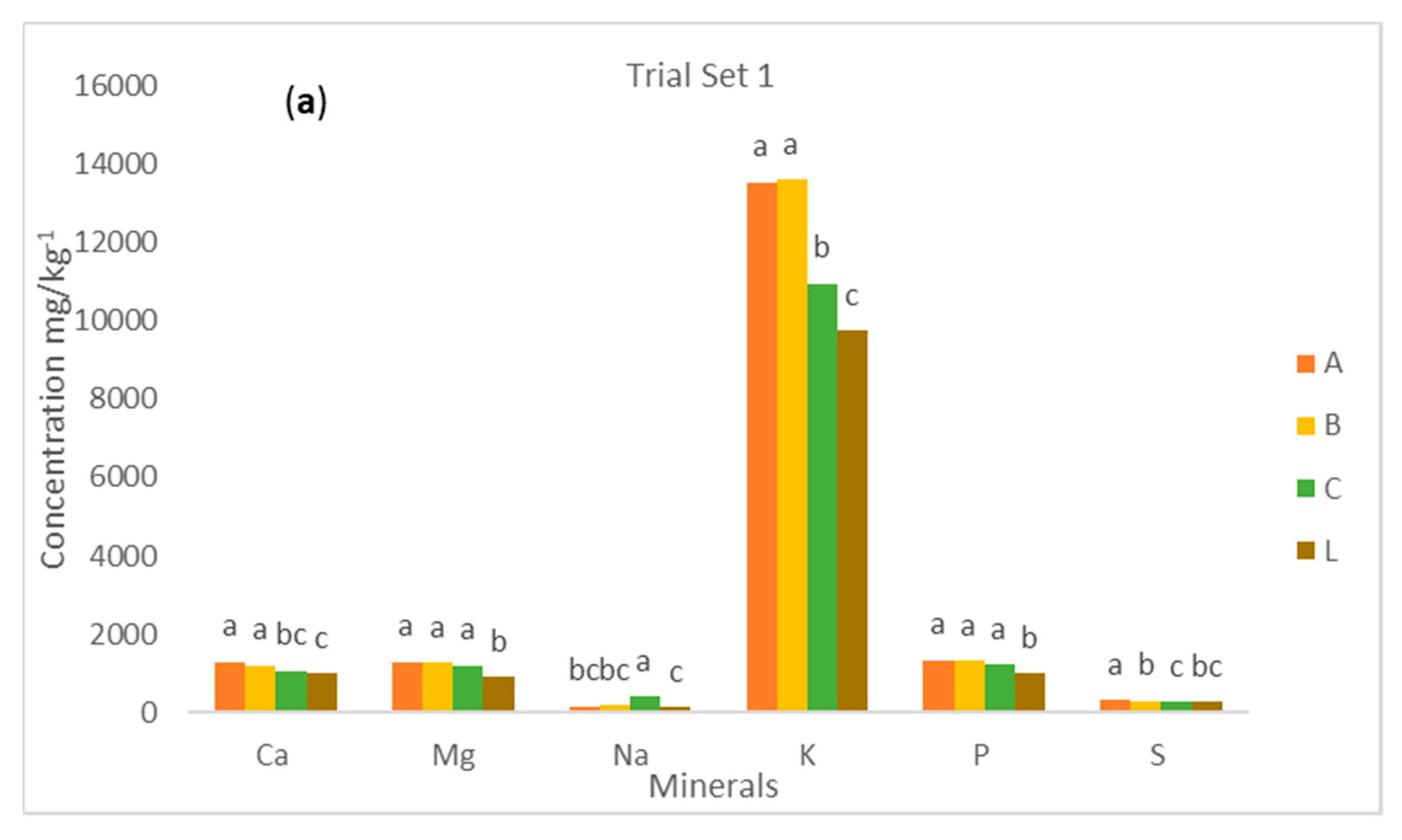
The mean values of the Ca, Mg, Na, K, P, and S content in yellow root cassava genotypes by sampling methods—method 1 (ABC) and method 2 (L) are presented in
Figure 2a,b. The table also gives information on the distribution of the macroelements from the proximal, middle, and distal parts of cassava roots. There was no significant (
p > 0.05) difference in the mean values of all macroelements studied in the proximal and middle parts of cassava roots except S, which showed a significant (
p < 0.05) difference. Additionally, there were significant (
p < 0.05) differences between the mean values of all the macroelements from the distal part and that of the proximal or middle part. It implies that macroelement concentrations are evenly distributed from the proximal to middle parts, but from the middle to the distal part, they are not evenly distributed. A similar pattern was observed for K, P, and S for set trial 2. However, a slightly different pattern was observed for the mean values of Ca, Mg, and Na, where there was no significant (
p < 0.05) difference observed for the proximal, middle, and distal sections of cassava roots. The results showed that the distribution of macroelements within the cassava is mineral dependent. It was also observed that the macroelement concentrations were lower at the distal part than the proximal or middle section. This could be due to the dilution effect because it has been established that the water distribution of cassava increases from the proximal to distal parts. However, we could conclude that the best part to sample macroelements in cassava roots should be from the proximal or middle part. The two methods were compared by averaging the values of each of the macroelements from the proximal, middle, and distal parts (method 1) and comparing them with the values of the macroelements from the longitudinal section of the cassava roots (method 2).
For both trials 1 and 2, there was a significant (
p < 0.05) difference in the mean values of all the macroelements except the Ca content of the set 2 trial. This shows that the concentration of the macroelements as a method is sampling dependent. It was observed that the mean value of all macroelements from method 1 was significantly (
p < 0.05) higher than the mean values from method 2 for both trials 1 and 2. The levels of Ca, Mg, K, and P were significantly higher in the proximal and middle portions of the samples in method ABC (method 1) than method L (method 2). However, the levels of Na and S were far higher in the distal and proximal portions in method ABC than method L, respectively. From this result, method 1 (where samples were taken from all parts and averaged) was found to be the best, but it is not cost-effective. Method 2 could be the best alternative because it is cost-effective but not as accurate as method 1. It can be recommended that using sampling method 1 to determine Ca, Mg, Na, K, P, and S contents in yellow root cassava is better than using method 2 as it gives a far higher value of the mineral content. For set trial 1, the study showed that there was a significant (
p < 0.05) difference in the mean values of macroelements. The levels of Ca, Mg, K, and P were significantly higher in the proximal and middle portions of the samples in method ABC than method L. However, the levels of Na and S were far higher in the distal and proximal portions in method ABC than method L, respectively. From this result, it can be recommended that using sampling method ABC to extract Ca, Mg, Na, K, P, and S contents in yellow root cassava is better than using method L as it gives a far higher value of the mineral content. This result was similar to Maziya-Dixon et al. [
13]. They observed that the mean cis- and trans-beta-carotene contents were generally highest in the proximal end and lowest in the distal end of yellow-fleshed cassava storage roots. However, they reported that there was no significant difference (
p > 0.05) between the two methods (the sectional and longitudinal) for the cis- and trans-beta-carotene content. The contrary results indicate that micronutrients are sampling dependent. The sampling method for minerals could be different for carotenes.
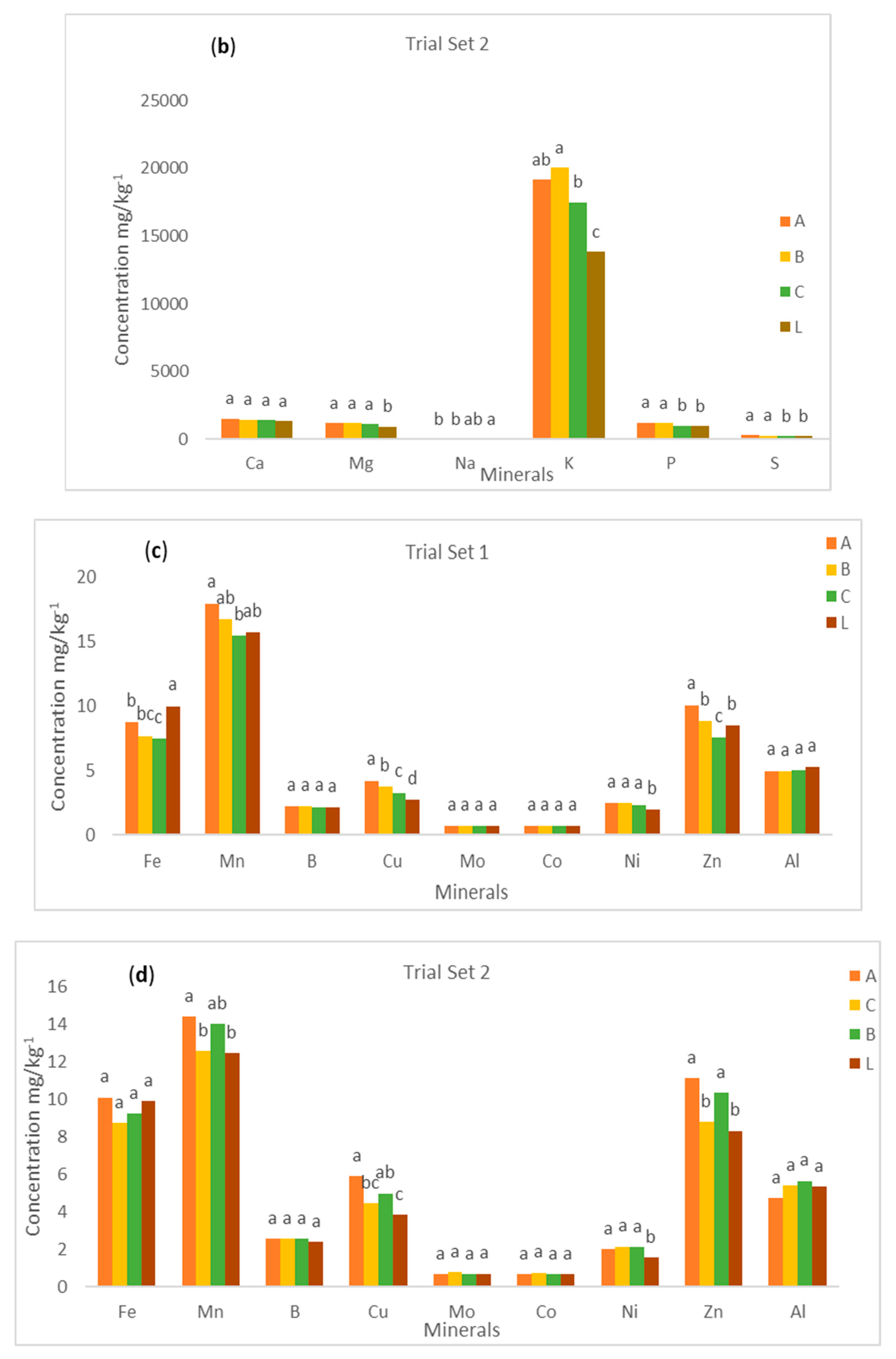
The mean values of Fe, Mn, B, Cu, Mo, Co, Ni, Zn, and Al contents in yellow root cassava genotypes by sampling methods—method 1 (ABC) and method 2 (L)—are presented in
Figure 2c,d. The figure also gives information on the distribution of the microelements from the proximal, middle, and distal parts of cassava roots. From trial set 1, there was a significant difference (
p < 0.05) in the mean values of Fe, Mn, Cu, and Zn elements in all the three sections (proximal, middle, and distal) of cassava root. A similar pattern was observed for Mn, and Cu in the trial set 2. The mean values for Zn showed a significant difference in the proximal and middle parts only. This result implies that the concentrations of these microelements are not evenly distributed from the proximal to the distal part of the cassava root. However, the study showed that there was no significant difference (
p > 0.05) in the mean values of B, Mo, Co, Ni, and Al elements in the three sections of cassava root. Similarly, the mean values for Fe, B, Mo, Co, Ni, and Al did not show any significant difference (
p > 0.05) in all three sections in the trial set 2. Further, the study revealed that there was no significant difference (
p > 0.05) in the mean values of all microelements in the proximal part of the cassava root. It implies that the concentrations of Fe, B, Mo, Co, Ni, and Al are evenly distributed across the three cassava root sections that were studied.
The results showed that the distribution of microelements in the cassava root is mineral dependent. It was also observed that the microelement concentrations were lower in the distal part than the proximal or middle section. This could be due to the dilution effect because it has been established that the water distribution of cassava increases from the proximal to the distal part. However, we could conclude that the best part to sample microelements in cassava roots from is the proximal or middle part. The two sampling methods were compared by averaging the values of each of the microelements from the proximal, middle, and distal parts (method 1) and comparing them with the values of the microelements from the longitudinal section of the cassava roots (method 2). For both trial 1 and 2, there was a significant (p < 0.05) difference in the mean values of all the microelements except the B, Mo, Co, and Al content of trial set 1. Additionally, there was a significant difference (p < 0.05) in the mean values for all the microelements except for Fe, B, Mo, Co, and Al in the trial set 2. This shows that the concentration of microelements in a method that is sampling dependent. It was observed that the mean value of all microelements from method 1 was significantly (p < 0.05) higher than the mean values from method 2 for both trial set 1 and 2. The levels of Mn, Cu, Ni, and Zn were found to be significantly higher in the proximal portions of the samples in method ABC (method 1) than method L (method 2). However, the study showed that Fe had far higher levels in method L (method 2) than method ABC (method 1) in the trial set 1.
Further, the results of the study revealed that the B, Mo, Co, and Al contents did not show any significant difference (p > 0.05) between the two sampling methods. For trial set 2, the contents of Mn, Cu, Ni, and Zn were found to be significantly higher in the proximal portions of the samples investigated in method ABC (method 1) than in method L (method 2). However, the study revealed that the levels of Fe, B, Mo, Co, and Al did not show any significant difference in the two sampling methods. As a result, any of the two methods could be used for the extraction of these microelements. It can also be recommended that using sampling method ABC (method 1) for extraction of Mn, Cu, Ni, and Zn contents in cassava roots is better than using method L (method 2).
3.7. Pearson Correlation Coefficients of Macro- and Micromineral Contents of Yellow-Fleshed Cassava Roots
The Pearson correlation coefficients of the macromineral content in yellow root cassava samples investigated are presented in
Table 8. From the dataset of trial 1, there was a significant positive (
p < 0.001) correlation between the K and P content (r = 0.513). This correlation shows the possibility of breeding yellow root cassava varieties with high K and P mineral contents. However, the results showed that there was a significant negative (
p < 0.05) correlation between the Na and K content (r = −0.271). Further, a significant (
p < 0.05) but weak correlation was observed between the P and Mg content (r = −0.062), P and Na content (r = −0.073), and S and Na content (r = −0.67). From the dataset of trial 2, the results showed a significant positive (
p < 0.001) correlation between the Ca and Mg content (r = 0.605), Ca and K content (r = 0.502), Mg and K content (r = 0.606), and K and P content (r = 0.636). However, the study showed a significant negative (
p < 0.05) correlation between the K and Na content (r = −0.126), and a significant (
p < 0.05) but weak correlation between P and Na (r = −0.052).
The Pearson correlation coefficients of the micromineral contents in the yellow root cassava samples investigated are presented in
Table 9. From the data set 1 trial, there was a significant positive (
p < 0.001) correlation between the Mn and B contents (r = 0.539). This correlation shows the possibility of breeding yellow root cassava varieties with high Mn and B mineral contents. However, the results of the study also showed that there was a significant negative (
p < 0.05) correlation between the Fe and Ni contents (r = −0.22), and Al and Ni contents (r = −0.228). Further, a significant (
p < 0.05) but a weak correlation was observed between the B and Mo contents (r = −0.161), Cu and Al contents (r = −0.186), as well as the Zn and Al contents (r = −0.141).
From the data set 2 trial, the results of the study showed a significant positive (p < 0.001) correlation between the Mo and Co contents (r = 0.991), B and Mo contents (r = 0.789), B and Co contents (r = 0.783), Fe and B contents (r = 0.663), Mn and Zn contents (r = 0.618), Fe and Co contents (r = 0.625), Co and Al contents (r = 0.604), Mo and Al contents (r = 0.603), Fe and Al contents (r = 0.536), and B and Al contents (r = 0.532). However, the study also showed that there was no correlation between the Al and Ni (r = 0.089), Al and Zn (r = 0.069), and Ni and Cu (r = 0.142) contents.
The observed strong positive correlation shows that those minerals could be bred together to increase their concentrations in yellow cassava roots. However, those minerals with a negative and significant correlation could be difficult to improve together in the breeding program as one will increase, and the other will decrease. Curiously, both for macro- and microminerals, r values >0.5 and lower p-values were found for set 2 than set 1, and the levels of minerals were higher than set 1.

 ,
, 




{kind=link}
{kind=link}
{kind=link}
{kind=link}