Blowup of Accidental Images as a Passageway to Discovery: Insights into the Interaction between Hydroxyapatite Nanoparticles and Human Mesenchymal Stem Cells


{kind=link}
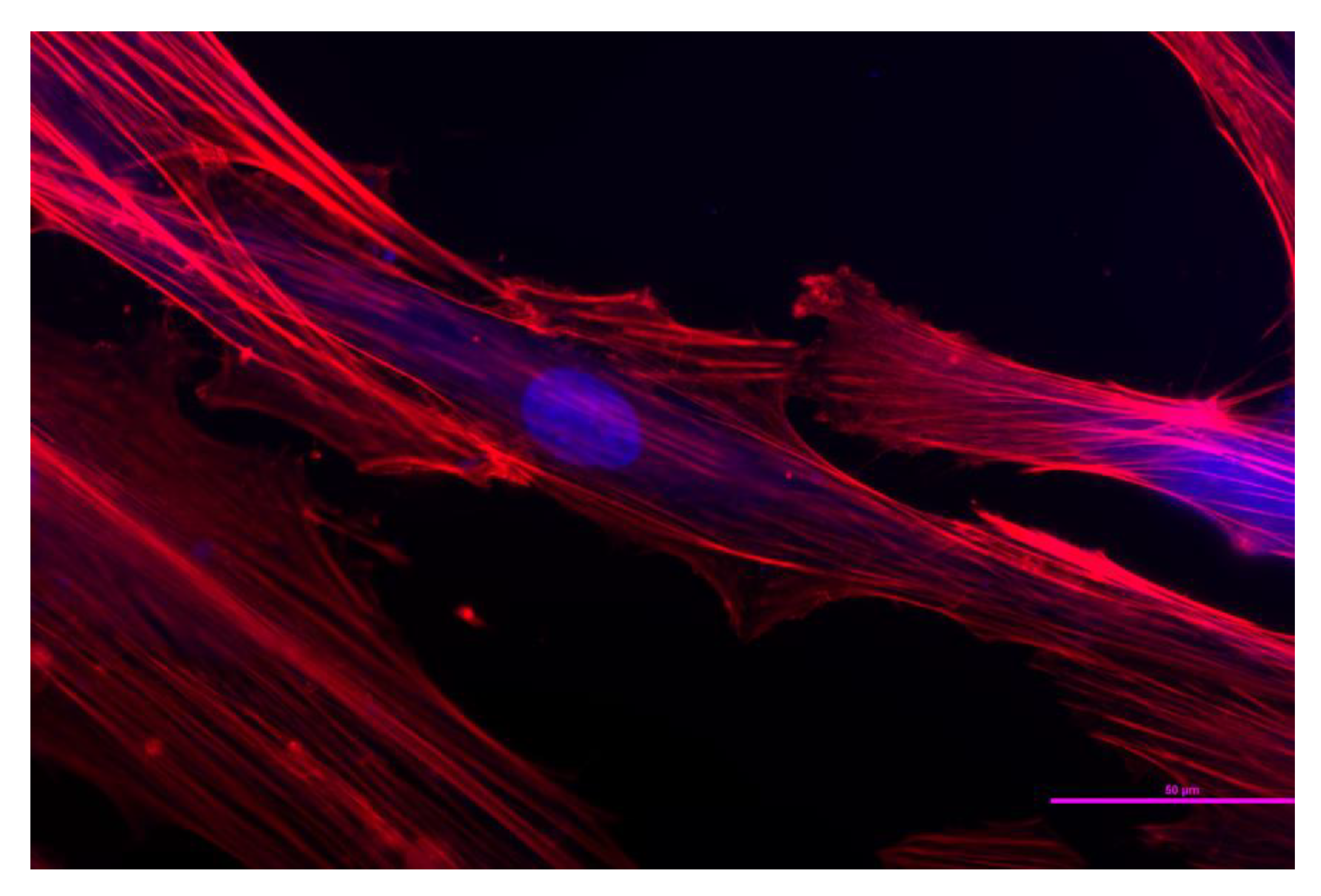{kind=link}
{kind=link}
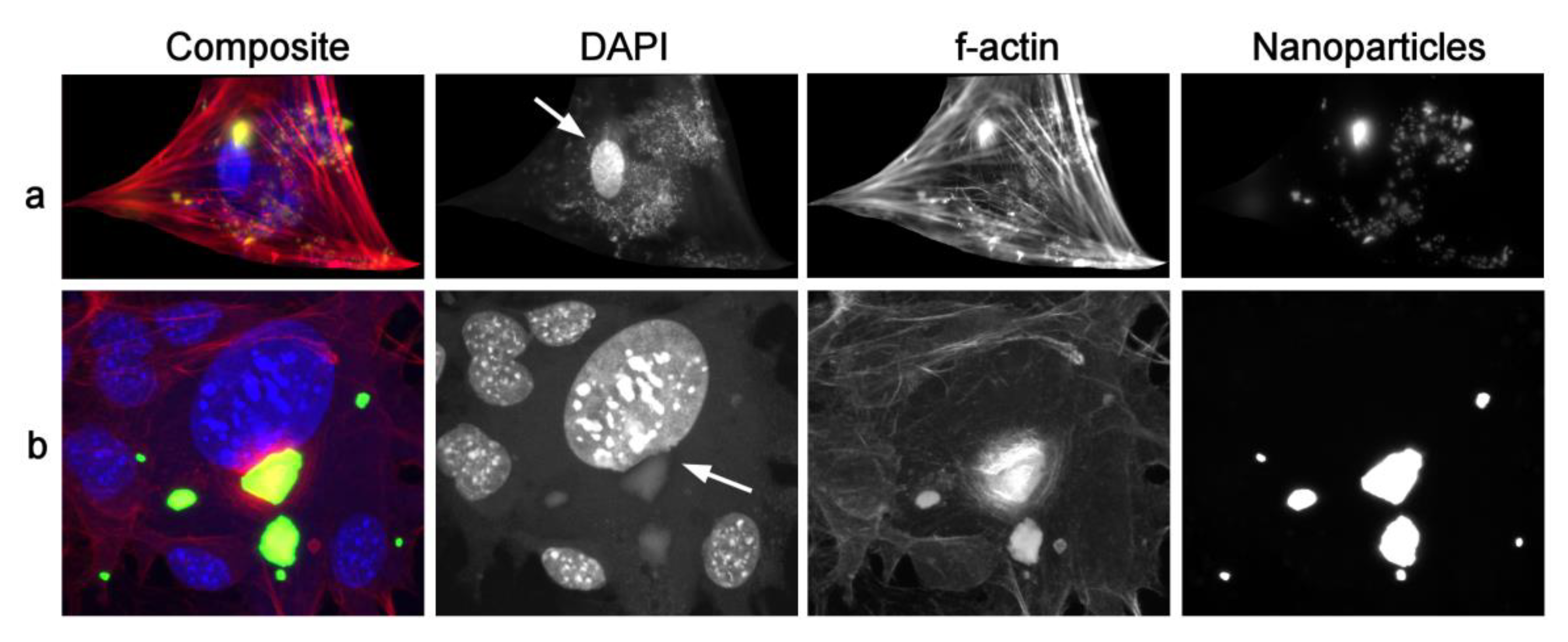{kind=link}
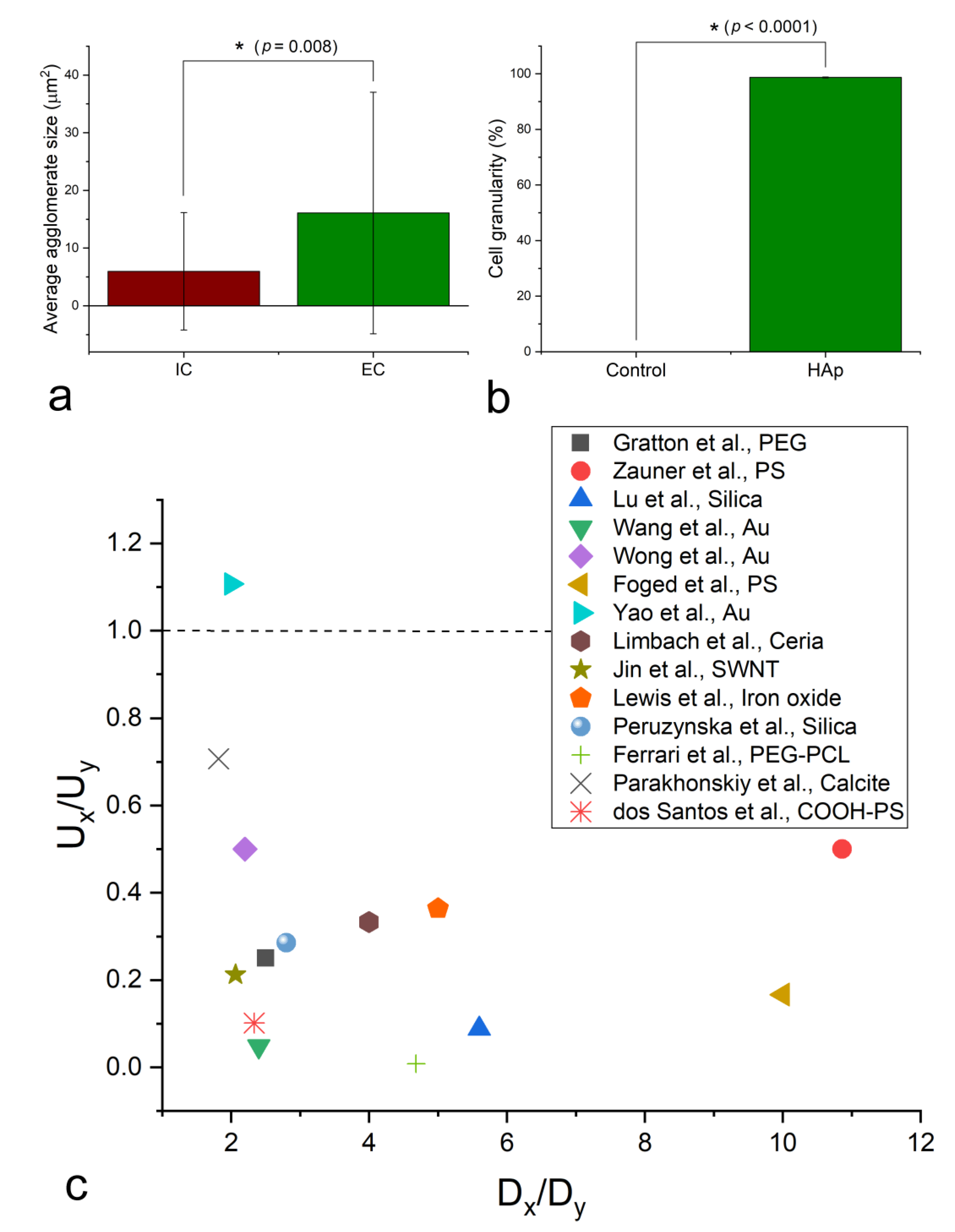{kind=link}
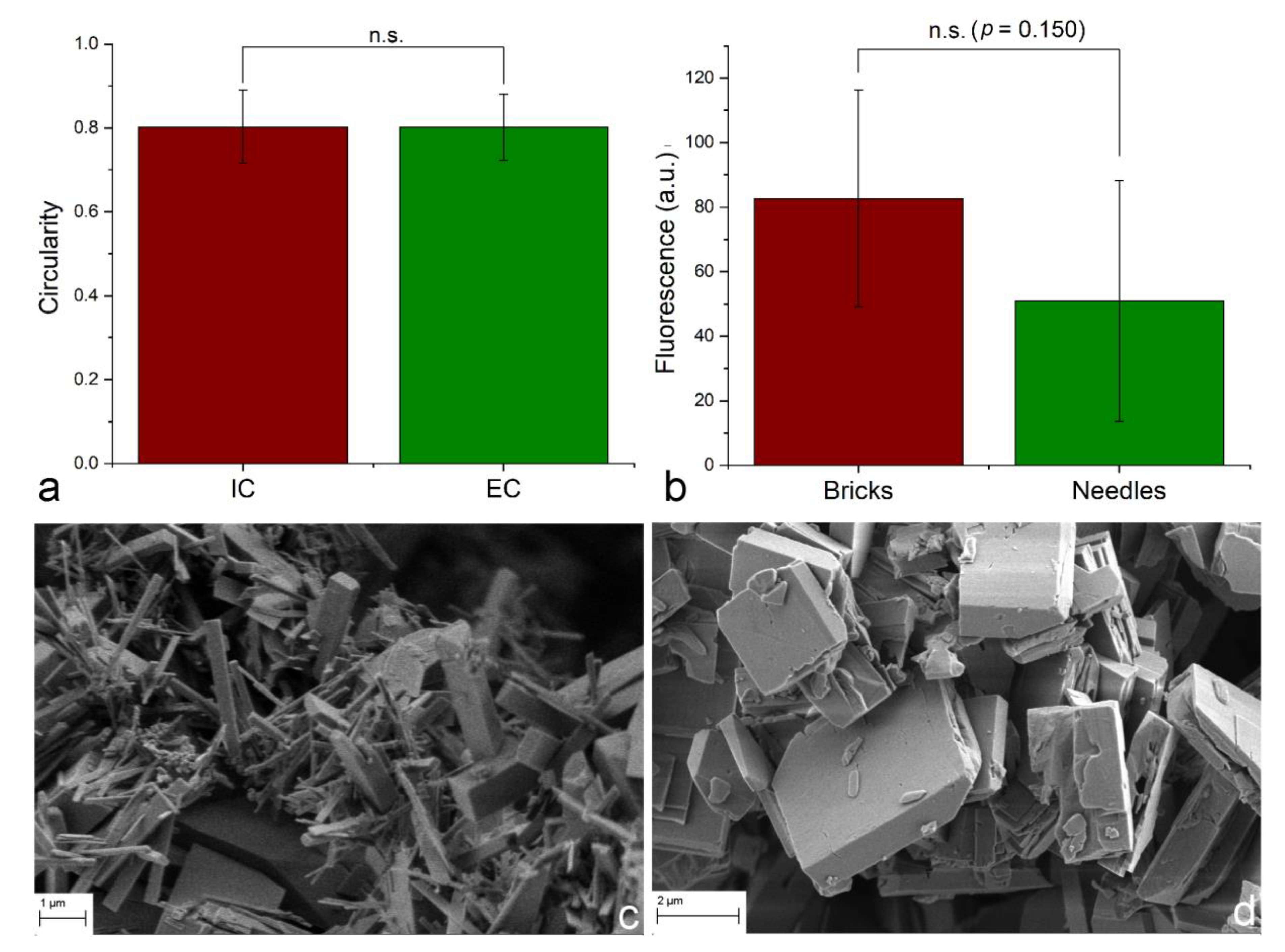{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Nanoparticle Synthesis
2.2. Cell Culture
2.3. AFM and SEM Imaging
2.4. Immunofluorescent Staining
2.5. Cell Viability Assay
2.6. Flow Cytometry
3. Results and Discussion
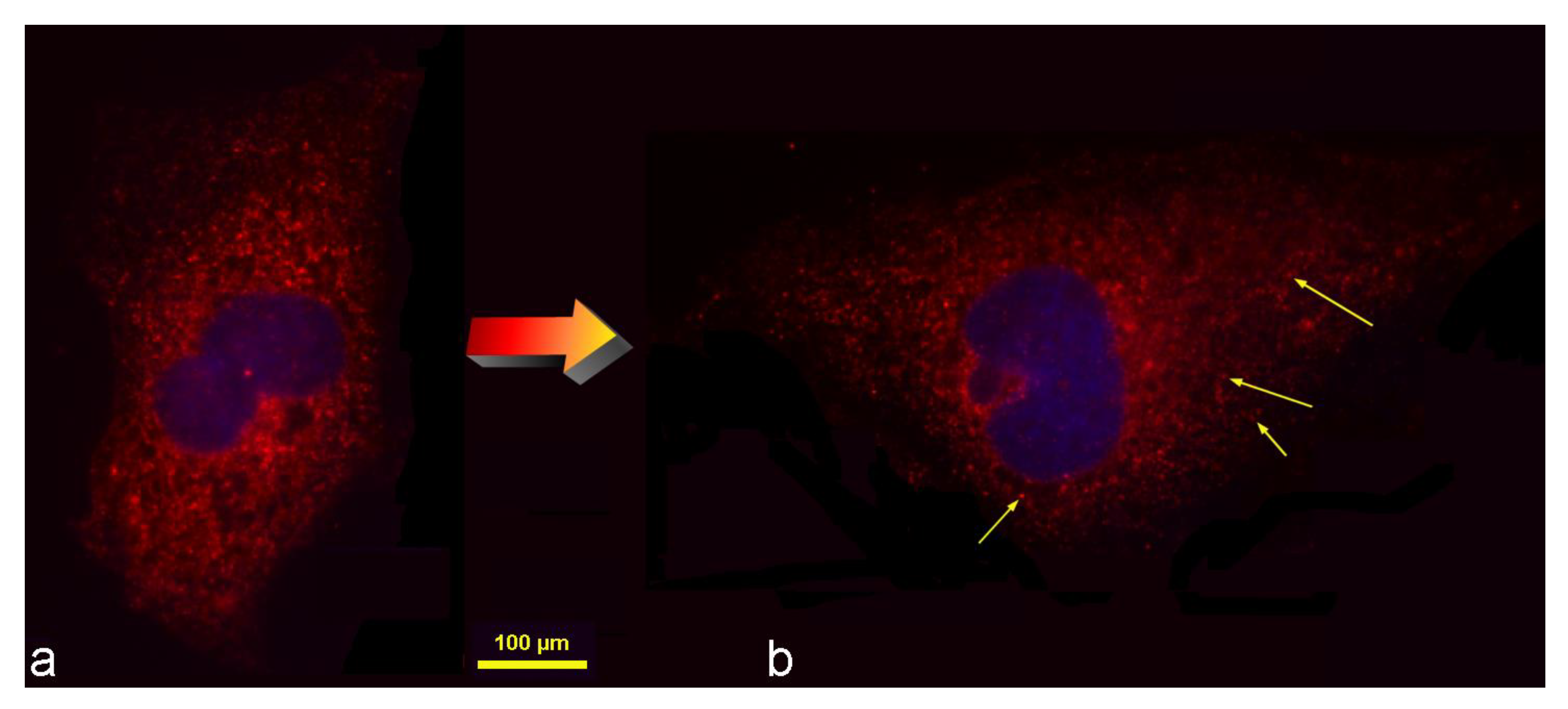
3.1. Smaller Particle Conglomerates Are More Likely to Be Internalized by the Cells
3.2. More Irregular Particle Clusters Are Equally Likely to Be Internalized as the Less Irregular Ones
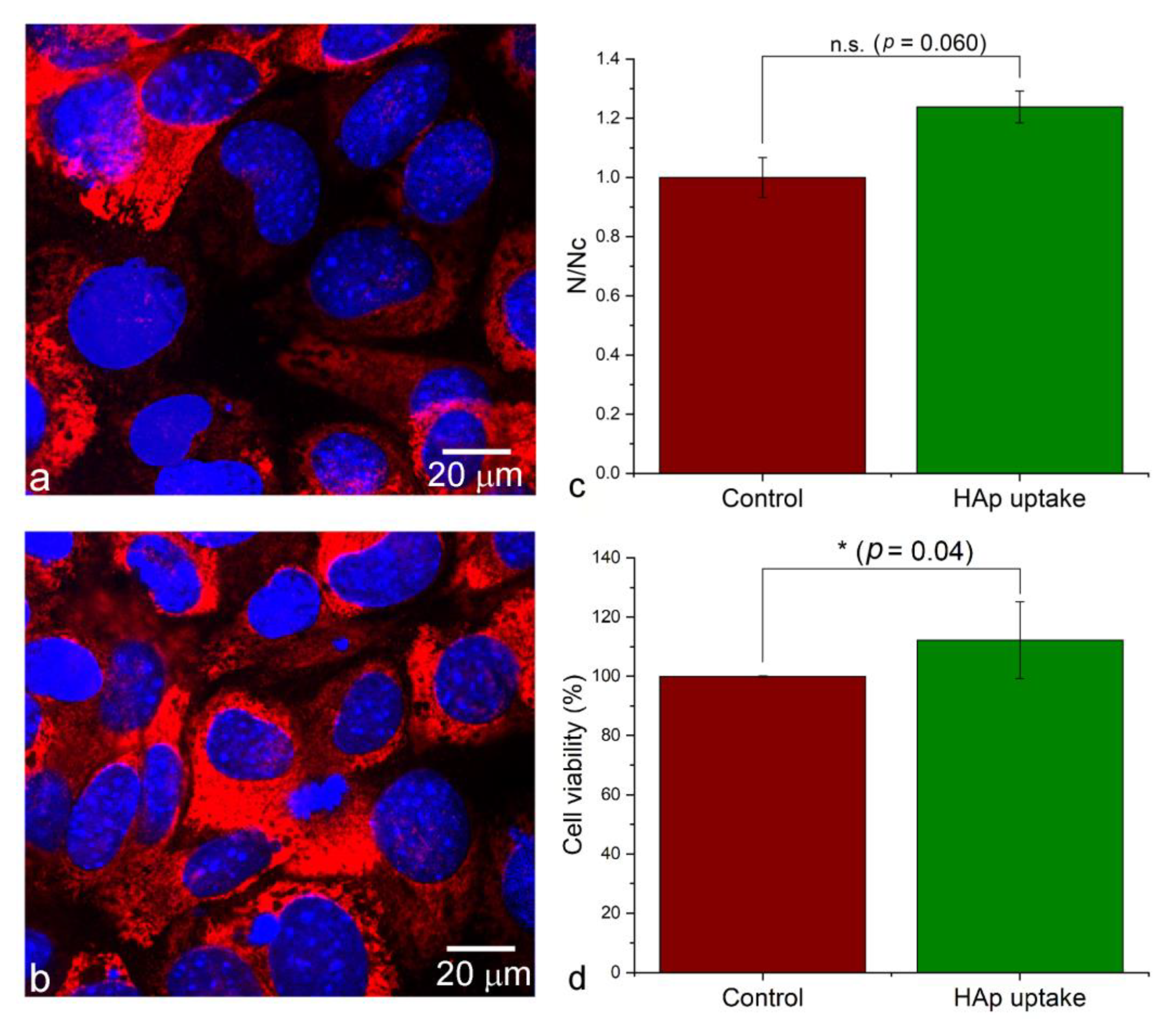
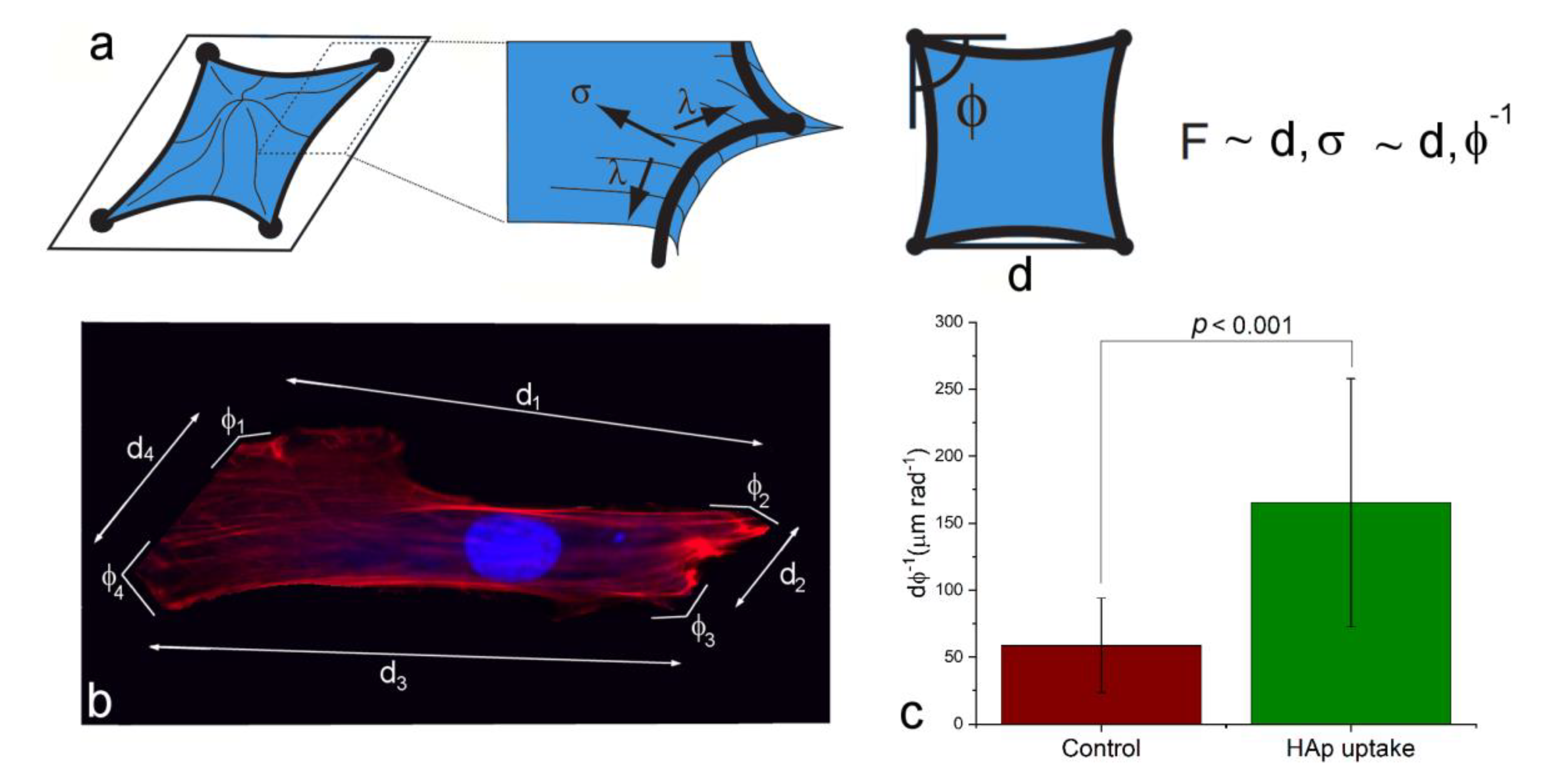
3.3. Uptake of HAp Nanoparticles Can Facilitate the Cell Adhesion and Proliferation on the Surface
4. Conclusions
Funding
Acknowledgments
Conflicts of Interest
References
- Poormoghadam, D.; Almasi, A.; Ashrafizadeh, M.; Vishkaei, A.S.; Reyazat, S.M.; Tavakol, S. The particle size of drug nanocarriers dictates the fate of neurons; critical points in neurological therapeutics. Nanotechnology 2020, 31, 335101. [Google Scholar] [CrossRef] [PubMed]
- Perez-Garnes, M.; Gutierrez-Salmeron, M.; Morales, V.; Chocarro-Calvo, A.; Sanz, R.; Garcia-Jimenez, C.; Garcia-Munoz, R.A. Engineering hollow mesoporous silica nanoparticles to increase cytotoxicity. Mat. Sci. Eng. C 2020, 112, 110935. [Google Scholar] [CrossRef] [PubMed]
- Shanei, A.; Akbari-Zadeh, H.; Attaran, N.; Salamat, M.R.; Baradaran-Ghahfarokhi, M. Effect of targeted gold nanoparticles size on acoustic cavitation: An in vitro study on melanoma cells. Ultrasonics 2020, 102, 106061. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.-D.; Wu, D.; Shen, X.; Liu, P.-X.; Yang, N.; Zhao, B.; Zhang, H.; Sun, Y.-M.; Zhang, L.-A.; Fan, F.-Y. Size-dependent in vivo toxicity of PEG-coated gold nanoparticles. Int. J. Nanomed. 2011, 6, 2071–2081. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, B.J.; Liu, N.; Liu, Q.S.; Zhang, J.Q.; Zhou, Q.F.; Jiang, G.B. Silver nanoparticles induce size-dependent and particle-specific neurotoxicity to primary cultures of rat cerebral cortical neurons. Ecotoxicol. Environ. Saf. 2020, 198, 110674. [Google Scholar] [CrossRef] [PubMed]
- Arrieta-Sandoval, N.; Rojas, P.E.; Olivas-Armendariz, I.; Gomez, L.V.E.; Paz, J.H.F.; Cordero, B.M.E.; Gonzalez, C.R.A. Effect of Ag2S-BSA nanoparticle size on 3T3 fibroblast cell line cytotoxicity. J. Nanopart. Res. 2020, 22, 106. [Google Scholar] [CrossRef]
- Zhang, S.; Wu, S.; Shen, Y.R.; Xiao, Y.Q.; Gao, L.Z.; Shi, S.R. Cytotoxicity studies of Fe3O4 nanoparticles in chicken macrophage cells. R. Soc. Open Sci. 2020, 7, 191561. [Google Scholar] [CrossRef] [Green Version]
- Thiramanas, R.; Jiang, S.; Simon, J.; Landfester, K.; Mailander, V. Silica nanocapsules with different sizes and physicochemical properties as suitable nanocarriers for uptake in T-cells. Int. J. Nanomed. 2020, 15, 6069–6084. [Google Scholar] [CrossRef]
- Colaco, M.; Marques, A.P.; Jesus, S.; Duarte, A.; Borges, O. Safe-by-Design of Glucan nanoparticles: Size matters when assessing the immunotoxicity. Chem. Res. Toxicol. 2020, 33, 915–932. [Google Scholar] [CrossRef]
- Wroblewska-Wolna, A.M.; Harvie, A.J.; Rowe, S.F.; Critchley, K.; Butt, J.N.; Jeuken, L.J.C. Quantum dot interactions with and toxicity to Shewanella oneidensis MR-1. Nanotechnology 2020, 31, 134005. [Google Scholar] [CrossRef]
- Chen, Y.H.; D’Amario, C.; Gee, A.; Duong, H.T.T.; Shimoni, O.; Valenzuela, S.M. Dispersion stability and biocompatibility of four ligand-exchanged NaYF4:Yb, Er upconversion nanoparticles. Acta Biomater. 2020, 102, 384–393. [Google Scholar] [CrossRef] [PubMed]
- Bhattacharjee, S.; de Haan, L.H.; Evers, N.M.; Jiang, X.; Marcelis, A.T.; Zuilhof, H.; Rietjens, I.M.; Alink, G.M. Role of surface charge and oxidative stress in cytotoxicity of organic monolayer-coated silicon nanoparticles towards macrophage NR8383 cells. Part Fibre Toxicol. 2010, 7, 25. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tada-Oikawa, S.; Eguchi, M.; Yasuda, M.; Izouka, K.; Ikegami, A.; Vranic, S.; Boland, S.; Tran, L.; Ichihara, G.; Ichihara, S. Functionalized surface-charged SiO2 nanoparticles induce pro-inflammatory responses, but are not lethal to Caco-2 cells. Chem. Res. Toxicol. 2020, 33, 1226–1236. [Google Scholar] [CrossRef] [PubMed]
- Tomita, Y.; Rikimaru-Kaneko, A.; Hashiguchi, K.; Shirotake, S. Effect of anionic and cationic n-butylcyanoacrylate nanoparticles on NO and cytokine production in Raw264.7 cells. Immunopharmacol. Immunotoxicol. 2011, 33, 730–737. [Google Scholar] [CrossRef] [PubMed]
- Pavičić, I.; Milić, M.; Pongrac, I.M.; Ahmed, L.B.; Glavan, T.M.; Ilić, K.; Zapletal, E.; Ćurlin, M.; Mitrečić, D.; Vrček, I.V. Neurotoxicity of silver nanoparticles stabilized with different coating agents: In vitro response of neuronal precursor cells. Food Chem. Toxicol. 2020, 136, 110935. [Google Scholar] [CrossRef]
- Zhai, C.; Zhang, X.; Chen, J.; He, J.; Fei, H.; Liu, Y.; Luo, C.; Fan, W. The effect of cartilage extracellular matrix particle size on the chondrogenic differentiation of bone marrow mesenchymal stem cells. Regen. Med. 2019, 14, 663–680. [Google Scholar] [CrossRef]
- Yang, X.; Li, Y.; Liu, X.; Zhang, R.; Feng, Q. In Vitro Uptake of Hydroxyapatite Nanoparticles and Their Effect on Osteogenic Differentiation of Human Mesenchymal Stem Cells. Stem Cells Int. 2018, 2018, 2036176. [Google Scholar] [CrossRef] [Green Version]
- Kang, E.-S.; Song, I.; Kim, D.-S.; Lee, U.; Kim, J.-K.; Son, H.; Min, J.; Kim, T.-H. Size-dependent effects of graphene oxide on the osteogenesis of human adipose-derived mesenchymal stem cells. Colloids Surf B Biointerfaces 2018, 169, 20–29. [Google Scholar] [CrossRef]
- Yang, X.; Li, Y.; Huang, Q.; Liu, X.; Zhang, R.; Feng, Q. The effect of hydroxyapatite nanoparticles on adipogenic differentiation of human mesenchymal stem cells. J. Biomed. Mater. Res. Part A 2018, 106, 1822–1831. [Google Scholar] [CrossRef]
- Ko, W.-K.; Heo, D.N.; Moon, H.-J.; Lee, S.J.; Bae, M.S.; Lee, J.B.; Sun, I.-C.; Jeon, H.B.; Park, H.K.; Kwon, I.K. The effect of gold nanoparticle size on osteogenic differentiation of adipose-derived stem cells. J. Colloid Interface Sci. 2015, 438, 68–76. [Google Scholar] [CrossRef]
- Lv, L.; Liu, Y.; Zhang, P.; Zhang, X.; Liu, J.; Chen, T.; Su, P.; Li, H.; Zhou, Y. The nanoscale geometry of TiO2 nanotubes influences the osteogenic differentiation of human adipose-derived stem cells by modulating H3K4 trimethylation. Bio-Mater. 2015, 39, 193–205. [Google Scholar] [CrossRef] [PubMed]
- Weissenboeck, M.; Stein, E.; Undt, G.; Ewers, R.; Lauer, G.; Turhani, D. Particle size of hydroxyapatite granules calcified from red algae affects the osteogenic potential of human mesenchymal stem cells in vitro. Cells Tissues Organs 2006, 182, 79–88. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.; Guo, Y.Q.; He, X.; Chen, H.Q. Effect of Titanium Particle Size on Osteogenic Differentiation of Bone Marrow–Derived Mesenchymal Stem Cells. Key Eng. Mater. 2011, 474, 1939–1942. [Google Scholar] [CrossRef]
- Smith, P. M. Train; Alfred A. Knopf: New York, NY, USA, 2015. [Google Scholar]
- Uskoković, V. A Collection of Micrographs: Where Science and Art Meet. Technoetic Arts J. Specul. Res. 2010, 7, 231–248. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Uskoković, T.; Uskoković, E.; Wu, V.; Uskoković, V. Calcium Phosphate and Senescence of Orange Jubilees in the Summertime. ACS Appl. Bio Mater. 2020, 3, 3770–3784. [Google Scholar] [CrossRef]
- Uskoković, E.; Uskoković, T.; Wu, V.M.; Uskoković, V. … And All the World a Dream: Memory Outlining the Mysterious Temperature-Dependency of Crystallization of Water, aka the Mpemba Effect. Substantia 2020, 4, 59–117. [Google Scholar]
- Uskoković, V. Ion-Doped Hydroxyapatite: An Impasse or the Road to Follow? Ceram. Int. 2020, 46, 11443–11465. [Google Scholar] [CrossRef]
- Medawar, P. Is the Scientific Paper a Fraud? Listener, 12 September 1963; 377–378. [Google Scholar]
- Uskoković, V.; Batarni, S.S.; Schweicher, J.; King, A.; Desai, T.A. Effect of Calcium Phosphate Particle Shape and Size on their Antibacterial and Osteogenic Activity in the Delivery of Antibiotics in vitro. ACS Appl. Mater. Interfaces 2013, 5, 2422–2431. [Google Scholar] [CrossRef]
- Sekiya, I.; Larson, B.L.; Smith, J.R.; Pochampally, R.; Cui, J.G.; Prockop, D.J. Expansion of human adult stem cells from bone marrow stroma: Conditions that maximize the yields of early progenitors and evaluate their quality. Stem Cells 2002, 20, 530–541. [Google Scholar] [CrossRef]
- Sherer, N.M.; Lehmann, M.J.; Jimenez-Soto, L.F.; Ingmundson, A.; Horner, S.M.; Cicchetti, G.; Allen, P.G.; Pypaert, M.; Cunningham, J.M.; Mothes, W. Visualization of retroviral replication in living cells reveals building into multivesicular bodies. Traffic 2003, 4, 785–801. [Google Scholar] [CrossRef]
- Cox, E.P. A method of assigning numerical and percentage values to the degree of roundness of sand grains. J. Paleontol. 1927, 1, 179–183. [Google Scholar]
- Bagheri, G.; Bonadonna, C. Aerodynamics of Volcanic Particles: Characterization of Size, Shape, and Settling Velocity. In Volcanic Ash; Mackie, S., Cashman, K., Ricketts, H., Rust, A., Watson, M., Eds.; Elsevier: Amsterdam, The Netherlands, 2016. [Google Scholar]
- Geng, Y.; Dalhaimer, P.; Cai, S.; Tsai, R.; Tewari, M.; Minko, T.; Discher, D.E. Shape effects of filaments versus spherical particles in flow and drug delivery. Nat. Nanotechnol. 2007, 2, 249–255. [Google Scholar] [CrossRef] [PubMed]
- Treuel, L.; Jiang, X.; Nienhaus, G.U. New views on cellular uptake and trafficking of manufactured nanoparticles. J. Royal Soc. Interface 2013, 10, 20120939. [Google Scholar] [CrossRef] [PubMed]
- Loos, C.; Syrovets, T.; Musyanovych, A.; Mailander, V.; Landfester, K.; Nienhaus, G.U.; Simmet, T. Functionalized polystyrene nanoparticles as a platform for studying bio-nano interactions. Beilstein. J. Nanotechnol. 2014, 5, 2403–2412. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Florencio-Silva, R.; Sasso, G.R.S.; Sasso-Cerri, E.; Simões, M.J.; Cerri, P.S. Biology of Bone Tissue: Structure, Function, and Factors That Influence Bone Cells. Biomed. Res. Int. 2015, 2015, 421746. [Google Scholar] [CrossRef] [Green Version]
- Rau, J.V.; Fosca, M.; Fadeeva, I.V.; Kalay, S.; Culha, M.; Raucci, M.G.; Fasolino, I.; Ambrosio, L.; Antoniac, I.V.; Uskoković, V. Tricalcium Phosphate Bone Cement Supplemented with Boron Nitride Nanotubes with Enhanced Biological Properties. Mater. Sci. Eng. C Mater. Biol. Appl. 2020, 114, 111044. [Google Scholar] [CrossRef]
- Thorpe, A.A.; Creasey, S.; Sammon, C.; le Maitre, C.L. Hydroxyapatite nanoparticle injectable hydrogel scaffold to support osteogenic differentiation of human mesenchymal stem cells. Eur. Cell Mater. 2016, 5, 1–23. [Google Scholar] [CrossRef]
- Xu, S.J.; Qiu, Z.Y.; Wu, J.J.; Kong, X.D.; Weng, X.S.; Cui, F.Z.; Wang, X.M. Osteogenic Differentiation Gene Expression Profiling of hMSCs on Hydroxyapatite and Mineralized Collagen. Tissue Eng. Part A 2016, 22, 170–181. [Google Scholar] [CrossRef]
- Pino, A.M.; Rosen, C.J.; Rodríguez, J.P. In Osteoporosis, differentiation of mesenchymal stem cells (MSCs) improves bone marrow adipogenesis. Biol. Res. 2012, 45, 279–287. [Google Scholar] [CrossRef] [Green Version]
- Ma, C.; Tian, X.; Kim, J.P.; Xie, D.; Ao, X.; Shan, D.; Lin, Q.; Hudock, M.R.; Bai, X.; Yang, J. Citrate-based materials fuel human stem cells by metabonegenic regulation. PNAS 2018, 115, 11741–11750. [Google Scholar] [CrossRef] [Green Version]
- Hoh, J.H.; Schoenenberger, C.A. Surface morphology and mechanical properties of MDCK monolayers by atomic force microscopy. J. Cell Sci. 1994, 107, 1105–1114. [Google Scholar] [PubMed]
- Swanson, J.A.; Watts, C. Macropinocytosis. Trends Cell Biol. 1995, 5, 424–428. [Google Scholar] [CrossRef]
- Rejman, J.; Oberle, V.; Zuhorn, I.S.; Hoekstra, D. Size-dependent internalization of particles via the pathways of clathrin- and caveolae-mediated endocytosis. Biochem. J. 2004, 377 Pt 1, 159–169. [Google Scholar] [CrossRef]
- Yi, X.; Shi, X.H.; Gao, H.J. A universal law for cell uptake of one-dimensional nanomaterials. Nano Lett. 2014, 14, 1049–1055. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, S.; Gao, H.; Bao, G. Physical principles of nanoparticle cellular endocytosis. ACS Nano 2015, 9, 8655–8671. [Google Scholar] [CrossRef] [Green Version]
- De Saint-Exupery, A. The Little Prince; Reynal & Hitchcock: New York, NY, USA, 1943. [Google Scholar]
- Guduru, R. In Situ AFM Imaging of Nanoparticle-Cellular Membrane Interactions for a Drug Delivery Study. Master of Science Thesis, Florida International University, Miami, FL, USA, 2011. [Google Scholar]
- Uskoković, V. When 1 + 1 > 2: Nanostructured Composite Materials for Hard Tissue Engineering Applications. Mater. Sci. Eng. C Mater. Biol. Appl. 2015, 57, 434–451. [Google Scholar] [CrossRef] [Green Version]
- Choi, J.S.; Cao, J.; Naeem, M.; Noh, J.; Hasan, N.; Choi, H.K.; Yoo, J.W. Size-controlled biodegradable nanoparticles: Preparation and size-dependent cellular uptake and tumor cell growth inhibition. Coll. Surf. B 2014, 122, 545–551. [Google Scholar] [CrossRef]
- He, C.; Hu, Y.; Yin, L.; Tang, C.; Yin, C. Effects of particle size and surface charge on cellular uptake and biodistribution of polymeric nanoparticles. Biomaterials 2010, 31, 3657–3666. [Google Scholar] [CrossRef]
- Chono, S.; Tanino, T.; Seki, T.; Morimoto, K. Uptake characteristics of liposomes by rat alveolar macrophages: Influence of particle size and surface mannose modification. J. Pharm. Pharmacol. 2007, 59, 75–80. [Google Scholar] [CrossRef]
- Dalzon, B.; Torres, A.; Reymond, S.; Gallet, B.; Saint-Antonin, F.; Collin-Faure, V.; Moriscot, C.; Fenel, D.; Schoehn, G.; Aude-Garcia, C.; et al. Influences of Nanoparticles Characteristics on the Cellular Responses: The Example of Iron Oxide and Macrophages. Nanomaterials 2020, 10, 226. [Google Scholar] [CrossRef] [Green Version]
- Gao, H.; Shi, W.; Freund, L.B. Mechanics of receptor-mediated endocytosis. Proc. Natl. Acad. Sci. USA 2005, 102, 9469–9474. [Google Scholar] [CrossRef] [Green Version]
- Shang, L.; Nienhaus, K.; Jiang, X.; Yang, L.; Landfester, L.; Mailander, V.; Simmet, T.; Nienhaus, G.U. Nanoparticles interactions with live cells: Quantitative fluorescence microscopy of nanoparticles size effects. Beilstein J. Nanotechnol. 2014, 5, 2388–2397. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chithrani, B.D.; Chan, W.C.W. Elucidating the mechanism of cellular uptake and removal of protein-coated gold nanoparticles of different sizes and shapes. Nano Lett. 2007, 7, 1542–1550. [Google Scholar] [CrossRef] [PubMed]
- Osaki, F.; Kanamori, T.; Sando, S.; Sera, T.; Aoyama, Y. A quantum dot conjugated sugar ball and its cellular uptake. On the size effects of endocytosis in the subviral region. J. Am. Chem. Soc. 2004, 126, 6520–6521. [Google Scholar] [CrossRef] [PubMed]
- Gratton, S.E.A.; Ropp, P.A.; Pohlhaus, P.D.; Luft, J.C.; Madden, V.J.; Napier, M.E.; DeSimone, J.M. The effect of particle design on cellular internalization pathways. Proc. Natl. Acad. Sci. USA 2008, 105, 11613–11618. [Google Scholar] [CrossRef] [Green Version]
- Lu, F.; Wu, S.; Hung, Y.; Mou, C.Y. Size effect on cell uptake in well-suspended, uniform mesoporous silica nanoparticles. Small 2009, 5, 1408–1413. [Google Scholar] [CrossRef] [PubMed]
- Wong, A.C.; Wright, D.W. Size-dependent cellular uptake of DNA functionalized gold nanoparticles. Small 2016, 12, 5592–5600. [Google Scholar] [CrossRef]
- Yao, M.; He, L.; McClements, D.J.; Xiao, H. Uptake of gold nanoparticles by intestinal epithelial cells: Impact of particle size on their absorption, accumulation, and toxicity. J. Agric. Food Chem. 2015, 63, 8044–8049. [Google Scholar] [CrossRef]
- Limbach, L.K.; Li, Y.; Grass, R.N.; Brunner, T.J.; Hintermann, M.A.; Muller, M.; Gunther, D.; Stark, W.J. Oxide nanoparticle uptake in human lung fibroblasts: Effects of particle size, agglomeration, and diffusion at low concentrations. Environ. Sci. Technol. 2005, 39, 9370–9376. [Google Scholar] [CrossRef]
- Lewis, E.E.L.; Child, H.W.; Hursthouse, A.; Stirling, D.; McCully, M.; Paterson, D.; Mullin, M.; Berry, C.C. The influence of particle size and static magnetic fields on the uptake of magnetic nanoparticles into three dimensional cell-seeded collagen gel cultures. J. Biomed. Mat. Res. 2015, 103, 1294–1301. [Google Scholar] [CrossRef] [Green Version]
- Wang, S.H.; Lee, C.W.; Chiou, A.; Wei, P.K. Size-dependent endocytosis of gold nanoparticles studied by three-dimensional mapping of plasmonic scattering images. J. Nanobiotechnol. 2010, 8, 33. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Foged, C.; Brodin, B.; Frokjaer, S.; Sundblad, A. Particle size and surface charge affect particle uptake by human dendritic cells in an in vitro model. Int. J. Pharm. 2005, 298, 315–322. [Google Scholar] [CrossRef] [PubMed]
- Zauner, W.; Farrow, N.A.; Haines, A.M.R. In vitro uptake of polystyrene microspheres: Effect of particle size, cell line and cell density. J. Controlled Release 2001, 71, 39–51. [Google Scholar] [CrossRef]
- Peruzynska, M.; Cendrowski, K.; Barylak, M.; Roginska, D.; Tarnowski, M.; Tkacz, M.; Kurzawski, M.; Machalinski, B.; Mijowska, E.; Drozdzik, M. Study on size effect of the silica nanospheres with solid core and mesoporous shell on cellular uptake. Biomed. Mater. 2015, 10, 065012. [Google Scholar] [CrossRef]
- Ferrari, R.; Lupi, M.; Colombo, C.; Morbidelli, M.; D’Incalci, M.; Moscatelli, D. Investigation of size, surface charge, PEGylation degree and concentration on the cellular uptake of polymer nanoparticles. Coll. Surf. B 2014, 123, 639–647. [Google Scholar] [CrossRef]
- Dos Santos, T.; Varela, J.; Lynch, I.; Salvati, A.; Dawson, K.A. Quantitative assessment of the comparative nanoparticle-uptake efficiency of a range of cell lines. Small 2011, 7, 3341–3349. [Google Scholar] [CrossRef] [Green Version]
- Jin, H.; Heller, D.A.; Sharma, R.; Strano, M.S. Size-dependent cellular uptake and expulsion of single-walled carbon nanotubes: Single particle tracking and a generic uptake model for nanoparticles. ACS Nano 2009, 3, 149–158. [Google Scholar] [CrossRef]
- Parakhonskiy, B.; Zyuzin, M.V.; Yashchenok, A.; Carregal-Romero, S.; Rejman, J.; Mohwald, H.; Parak, W.J.; Skirtach, A.G. The influence of the size and aspect ratio of anisotropic, porous CaCO3 particles on their uptake by cells. J. Nanobiotechnol. 2015, 12, 53. [Google Scholar] [CrossRef] [Green Version]
- Kashani, A.S.; Badilescu, S.; Piekny, A.; Packirisamy, M. Differing Affinities of Gold Nanostars and Nanospheres toward HeLa and HepG2 Cells: Implications for Cancer Therapy. ACS Appl. Nano Mater. 2020, 3, 4114–4126. [Google Scholar] [CrossRef]
- Zhang, Z.K.; Liu, C.R.; Li, C.; Wu, W.; Jiang, X.Q. Shape Effects of Cylindrical versus Spherical Unimolecular Polymer Nanomaterials on in Vitro and in Vivo Behaviors. Research 2019, 2019, 2391484. [Google Scholar] [CrossRef] [Green Version]
- Zhuang, J.; Wang, D.D.; Li, D.; Yang, Y.Q.; Lu, Y.; Wu, W.; Wu, W.; Qi, J.P. The influence of nanoparticle shape on bilateral exocytosis from Caco-2 cells. Chin. Chem. Lett. 2018, 29, 1815–1818. [Google Scholar] [CrossRef]
- Xu, D.; Wan, Y.X.; Li, Z.H.; Wang, C.B.; Zou, Q.X.; Du, C.; Wang, Y.J. Tailorable hierarchical structures of biomimetic hydroxyapatite micro/nano particles promoting endocytosis and osteogenic differentiation of stem cells. Biomater. Sci. 2020, 8, 3286–3300. [Google Scholar] [CrossRef] [PubMed]
- Yang, G.; Liu, H.; Hu, X.; Chen, Z.; Friis, T.E.; Wang, J.; Xiao, Y.; Zhang, S. Bio-inspired hybrid nanoparticles promote vascularized bone regeneration in a morphology-dependent manner. Nanoscale 2017, 9, 5794–5805. [Google Scholar] [CrossRef] [PubMed]
- Nambara, K.; Nikura, K.; Mitomo, H.; Ninomiya, T.; Takeuchi, C.; Wei, J.; Matsuo, Y.; Ijiro, K. Reverse size dependences of the cellular uptake of triangular and spherical gold nanoparticles. Langmuir 2016, 32, 12559–12567. [Google Scholar] [CrossRef]
- Farge, E.; Devaux, P.F. Shape changes of giant liposomes induced by an asymmetric transmembrane distribution of phospholipids. Biophys. J. 1992, 61, 347–357. [Google Scholar] [CrossRef] [Green Version]
- Uskoković, V.; Drofenik, M. Reverse Micelles: Inert Nano-Reactors or Physico-Chemically Active Guides of the Capped Reactions. Adv. Colloid Interface Sci. 2007, 133, 23–34. [Google Scholar] [CrossRef]
- Champion, J.A.; Mitragotri, S. Shape induced inhibition of phagocytosis of polymer particles. Pharm. Res. 2009, 26, 244–249. [Google Scholar] [CrossRef] [Green Version]
- Uskoković, V. Nanostructured Platforms for the Sustained and Local Delivery of Antibiotics in the Treatment of Osteomyelitis. Crit. Rev. Ther. Drug Carr. Syst. 2015, 32, 1–59. [Google Scholar] [CrossRef]
- Rasel, M.A.I.; Singh, S.; Nguyen, T.D.; Afara, I.O.; Gu, Y. Impact of Nanoparticle Uptake on the Biophysical Properties of Cell for Biomedical Engineering Applications. Sci. Rep. 2019, 9, 5859. [Google Scholar] [CrossRef] [Green Version]
- Yin, M.; Xu, W.G.; Cui, B.C.; Dai, H.L.; Han, Y.C.; Yin, Y.X.; Li, S.P. Effects of the interaction between hydroxyapatite nanoparticles and hepatoma cells. J. Wuhan Univ. Technol. Mat. Sci. Ed. 2014, 29, 635–642. [Google Scholar] [CrossRef]
- Yin, M.; Yin, Y.; Han, Y.; Dai, H.; Li, S. Effects of Uptake of Hydroxyapatite Nanoparticles into Hepatoma Cells on Cell Adhesion and Proliferation. J. Nanomater. 2014, 2014, 731897. [Google Scholar] [CrossRef] [Green Version]
- Wiegand, T.; Fratini, M.; Frey, F.; Yserentant, K.; Liu, Y.; Weber, E.; Galior, K.; Ohmes, J.; Braun, F.; Herten, D.P.; et al. Forces during cellular uptake of viruses and nanoparticles at the ventral side. Nat. Commun. 2020, 11, 32. [Google Scholar] [CrossRef] [PubMed]
- Bischofs, I.B.; Schmidt, S.S.; Schwarz, U.S. Effect of Adhesion Geometry and Rigidity on Cellular Force Distributions. Phys. Rev. Lett. 2009, 103, 048101. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tonelli, F.M.P.; Santos, A.K.; Gomes, D.A.; da Silva, S.L.; Gomes, K.N.; Ladeira, L.O.; Resende, R.R. Stem cells and calcium signaling. Adv. Exp. Med. Biol. 2012, 740, 891–916. [Google Scholar] [PubMed] [Green Version]
- Uskoković, V.; Desai, T.A. Does Translational Symmetry Matter on the Micro Scale? Fibroblastic and Osteoblastic Interactions with the Topographically Distinct Poly(ε-Caprolactone)/Hydroxyapatite Thin Films. ACS Appl. Mater. Interfaces 2014, 6, 13209–13220. [Google Scholar] [CrossRef]
- Hanna, H.; Mir, L.M.; Andre, F.M. In vitro osteoblastic differentiation of mesenchymal stem cells generates cell layers with distinct properties. Stem Cell Res. Ther. 2018, 9, 203. [Google Scholar] [CrossRef]
- Ghiasi, B.; Sefidbakht, Y.; Mozaffari-Jovin, S.; Gharachloo, B.; Mehraria, M.; Khodadadi, A.; Rezaei, M.; Ranaei-Siadat, S.O.; Uskoković, V. Hydroxyapatite as a Biomaterial–A Gift that Keeps on Giving. Drug Dev. Ind. Pharm. 2020, 46, 1035–1062. [Google Scholar] [CrossRef]
- Khan, M.A.; Wu, V.M.; Ghosh, S.; Uskoković, V. Gene Delivery Using Calcium Phosphate Nanoparticles: Optimization of the Transfection Process and the Effects of Citrate and Poly(L-Lysine) as Additives. J. Colloid Interface Sci. 2016, 471, 48–58. [Google Scholar] [CrossRef] [Green Version]
- Wu, V.M.; Uskoković, V. Waiting for Aπαταο: 250 Years Later. Found. Sci. 2019, 24, 617–640. [Google Scholar] [CrossRef]
- Wu, V.M.; Uskoković, V. Is There a Relationship between Solubility and Resorbability of Different Calcium Phosphate Phases in vitro? Biochim. Biophys. Acta Gen. Subj. 2016, 1860, 2157–2168. [Google Scholar] [CrossRef] [Green Version]
- Hallermann, S. Calcium channels for endocytosis. J. Physiol. 2014, 592 (Pt 16), 3343–3344. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, L.Y.; Chen, H.M.; Xie, J.; Becton, M.; Wang, X.Q. Interplay of Nanoparticle Rigidity and Its Translocation Ability through Cell Membrane. J. Phys. Chem. B 2019, 123, 8923–8930. [Google Scholar] [CrossRef] [PubMed]
- Uskoković, V. The Role of Hydroxyl Channel in Defining Selected Physicochemical Peculiarities Exhibited by Hydroxyapatite. RSC Adv. 2015, 5, 36614–36633. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rey, C.; Combes, C.; Drouet, C.; Glimcher, M.J. Bone mineral: Update on chemical composition and structure. Osteoporos. Int. 2009, 20, 1013–1021. [Google Scholar] [CrossRef] [Green Version]
- Uskoković, V.; Tang, S.; Nikolić, M.G.; Marković, S.; Wu, V.M. Calcium Phosphate Nanoparticles as Intrinsic Inorganic Antimicrobials: In Search of the Key Particle Property. Biointerphases 2019, 14, 031001. [Google Scholar] [CrossRef]
- Wu, V.M.; Mickens, J.; Uskoković, V. Bisphosphonate-Functionalized Calcium Phosphate Nanoparticles for the Delivery of the Bromodomain Inhibitor JQ1 in the Treatment of Osteosarcoma. ACS Appl. Mater. Interfaces 2017, 9, 25887–25904. [Google Scholar] [CrossRef]
- Pernal, S.P.; Wu, V.M.; Uskoković, V. Hydroxyapatite as a Vehicle for the Selective Effect of Superparamagnetic Iron Oxide Nanoparticles against Human Glioblastoma Cells. ACS Appl. Mater. Interfaces 2017, 9, 39283–39302. [Google Scholar] [CrossRef]
- Shi, X.X.; Zhou, K.; Huang, F.; Zhang, J.; Wang, C. Endocytic mechanisms and osteoinductive profile of hydroxyapatite nanoparticles in human umbilical cord Wharton’s jelly-derived mesenchymal stem cells. Int. J. Nanomed. 2018, 13, 1457–1470. [Google Scholar] [CrossRef] [Green Version]
- Uskoković, V. Celeste’s Plight: What Can Film Teach Natural Science? Film Int. 2019, 17, 69–88. [Google Scholar] [CrossRef]









Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Uskoković, V. Blowup of Accidental Images as a Passageway to Discovery: Insights into the Interaction between Hydroxyapatite Nanoparticles and Human Mesenchymal Stem Cells. Appl. Sci. 2020, 10, 8204. https://doi.org/10.3390/app10228204
Uskoković V. Blowup of Accidental Images as a Passageway to Discovery: Insights into the Interaction between Hydroxyapatite Nanoparticles and Human Mesenchymal Stem Cells. Applied Sciences. 2020; 10(22):8204. https://doi.org/10.3390/app10228204
Chicago/Turabian StyleUskoković, Vuk. 2020. "Blowup of Accidental Images as a Passageway to Discovery: Insights into the Interaction between Hydroxyapatite Nanoparticles and Human Mesenchymal Stem Cells" Applied Sciences 10, no. 22: 8204. https://doi.org/10.3390/app10228204
APA StyleUskoković, V. (2020). Blowup of Accidental Images as a Passageway to Discovery: Insights into the Interaction between Hydroxyapatite Nanoparticles and Human Mesenchymal Stem Cells. Applied Sciences, 10(22), 8204. https://doi.org/10.3390/app10228204