Jumping Locomotion Strategies: From Animals to Bioinspired Robots

 ,
, 


Abstract
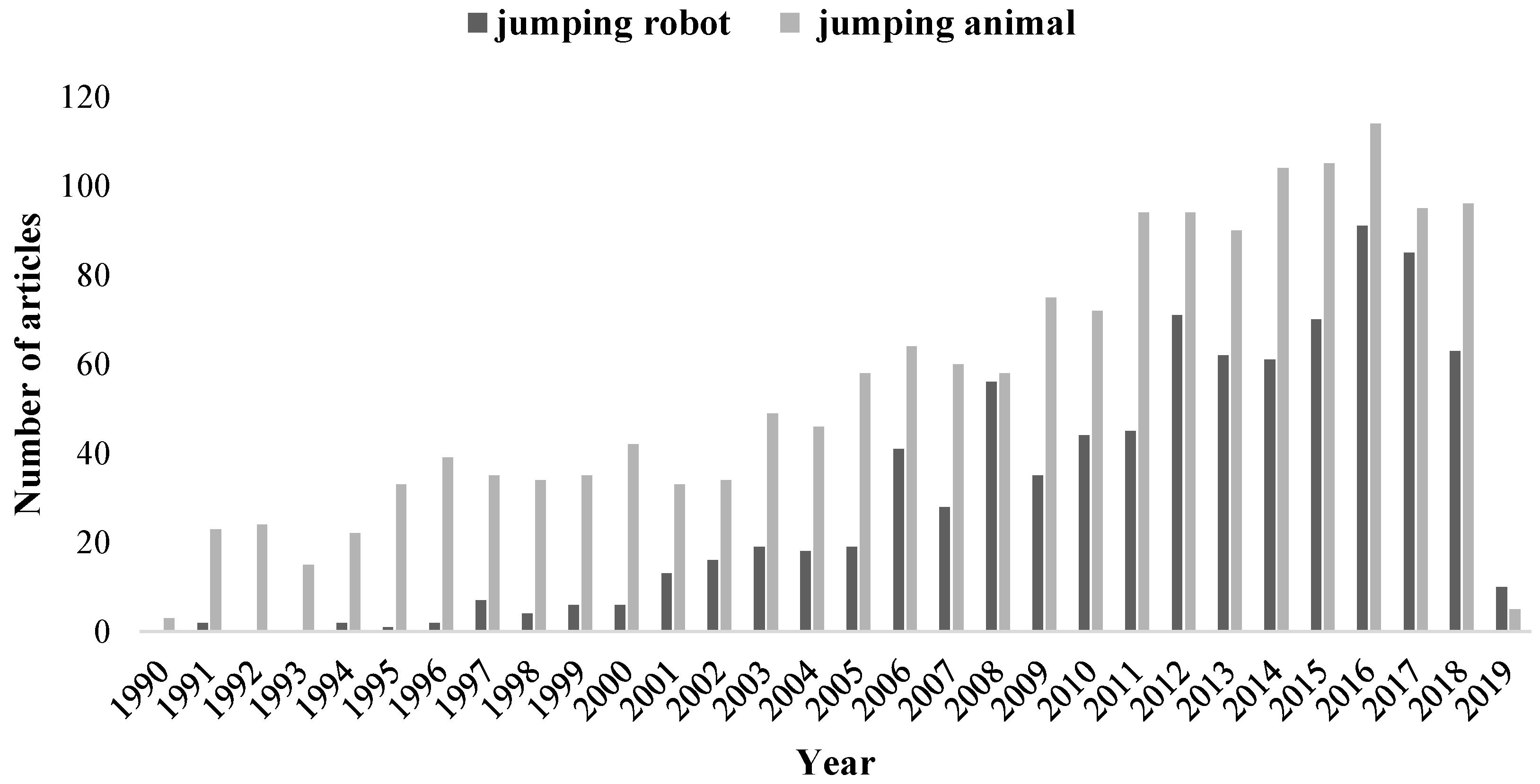
:1. Introduction
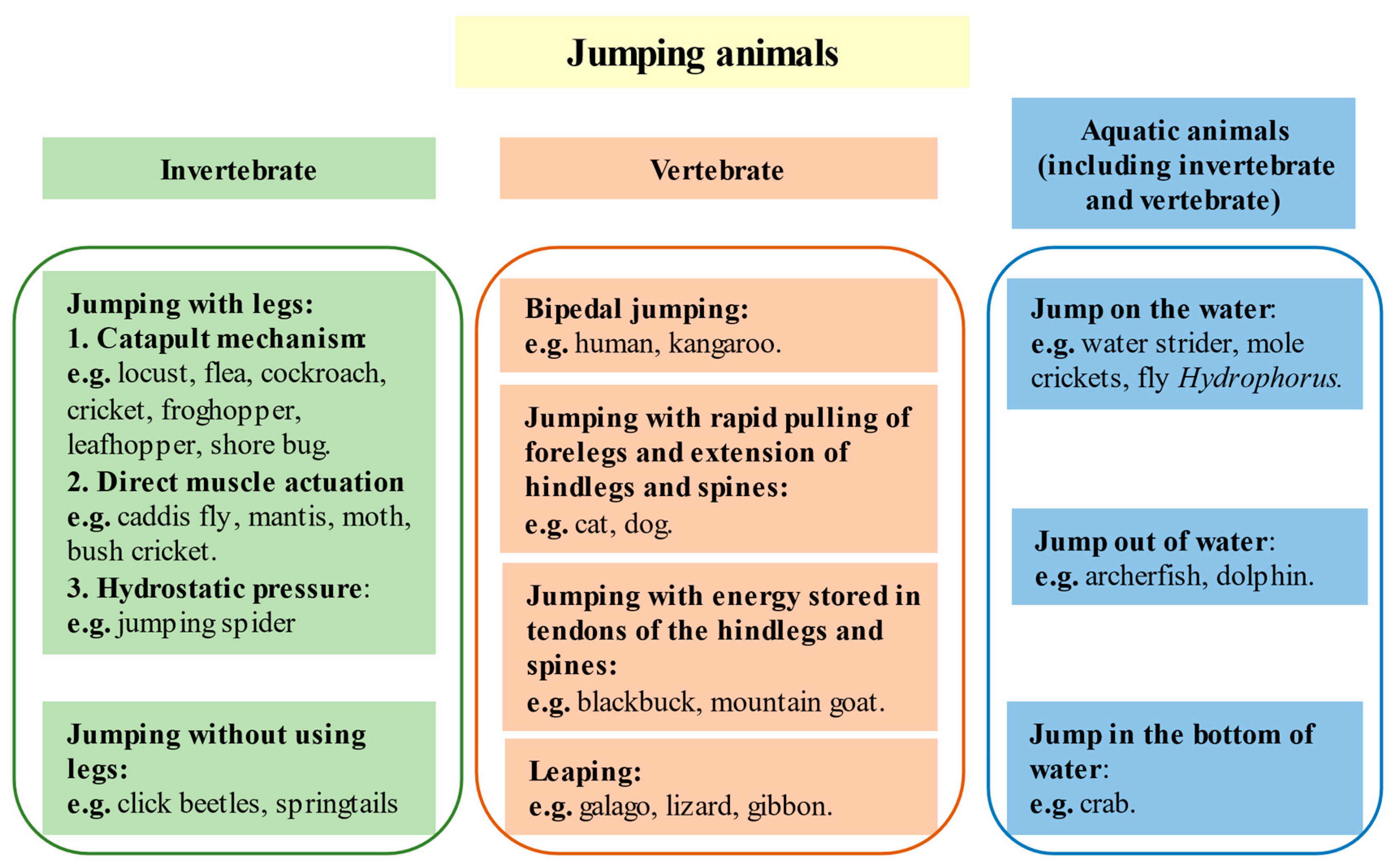
2. Jumping Animals and Biomimetic Miniature Jumping Robots
2.1. Invertebrates
2.1.1. Jumping Spider Inspired Jumping Robots
2.1.2. Locust Inspired Jumping Robots
2.1.3. Flea Inspiring Jumping Robot
2.1.4. Cricket Inspired Jumping Robots
2.1.5. Cockroach Inspired Jumping Robots
2.1.6. Froghopper Inspired Jumping Robots
2.1.7. Leafhopper Inspired Jumping Robots
2.2. Vertebrates
2.2.1. Frog Inspired Jumping Robots
2.2.2. Galago (Baby Monkey) Inspired Jumping Robots
2.2.3. Kangaroo-Inspired Jumping Robots
2.2.4. Human-Inspired Jumping Robots
2.2.5. Dog-Inspired Jumping Robots
2.3. Specialized Jumping Performance: Aquatic Environments
2.3.1. Water-Strider Inspired Jumping Robots
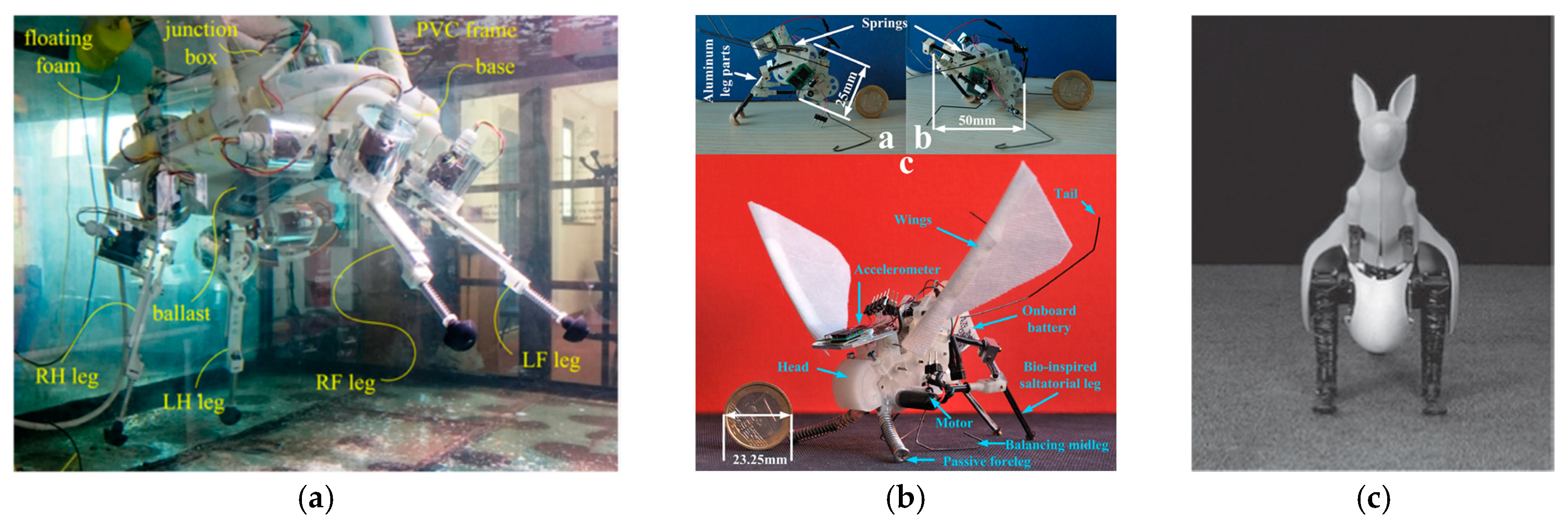
2.3.2. Dolphin-Inspired Jumping Robots
2.3.3. Crab Inspired Jumping Robots
3. Controllability of Take-Off in Jumping Animals and Robots
3.1. Take-off Angle
3.2. Take-off Direction

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Year | Name | Mass | Jumping Distance | Jumping Height | Actuators and Energy Storages | Highlights | Deficiencies | Aut 1 |
|---|---|---|---|---|---|---|---|---|
| 2020 | TAUB [99] | 43–46 g | N/A | 80–120 cm | Torsional spring | Single actuator, perform the flipping task after landing, reorient and jump in required direction | Jumping performance is decreased because of the added mechanism | Y |
| 2017 | Arachnabot [76] | 36 g | N/A | 11.5 cm | Fluidic air pressure | Pneumatics and electrically-actuated tendons capable of strong, dynamic, and rapid joint movement | Cannot control in air posture, landing, and re-jumping | N |
| 2017 | Salto-1P [160,161] | 98 g | >2 m | 125 cm | Torsional spring, thruster | Light weight, continuous jumping, high agility | No intermittent jumping, limited distance of wireless communication | Y |
| 2016 | Salto [158] | 100 g | N/A | 120 cm | Torsional spring | Light weight, two-stage jump for increased height, high agility | Need of artificial assistance, not able to jump forward | N |
| 2016 | Froghopper-inspired robot [135] | 3.06 g | N/A | N/A | SMA spring and linear spring | Added gears help realize synchronous jumping and change direction from −40° to 40° | Cannot control in air posture, landing, and re-jumping | N |
| 2016 | JumpRoACH [148] | 59.4 g | N/A | 1.1–1.62m | Latex rubber | Adjust both crawling speed and jumping take-off speed, height-adjustable via a novel active trigger | Landing and resetting abilities are poor | N |
| 2015 | Roly-poly Soft Robot [36] | 2.1 kg | N/A | 20 cm | Combustion | Soft material, combustion drive | Hard control on jumping direction, low jumping frequency | Y |
| 2014 | MultiMo-Bat [151] | 115.6 g | 232 cm | 305 cm | SMA wire, cable, clutch, linear spring | High jumping distance and height, components sharing, jumping with gliding | Impossibility to change take-off angle, lack of glide control and wings regulation | N |
| 2013 | MSU-tailbot [215] | 26.5 g | 90 cm | 87 cm | Spring | Wheel on the ground, control its body angle using an active tail to dynamically maneuver in midair for safe landings | Impossibility to change take-off angle and direction | Y |
| 2013 | Jumping Robotic Insect [121] | 34 mg | N/A | 30 cm | SMA actuators | Novel pop-up book MEMS process may eliminate the onerous assembly task caused by mesoscale components | Integrated untethered power is a challenging issue, cannot realize multiple jumping | N |
| 2013 | MSU-Jumper [214] | 23.5 g | 90 cm | 87 cm | Cable, rotation link, torsional spring | Light weight, minimum actuation, continuous steerable jumping | Attitude stabilization, no landing control | Y |
| 2013 | HAMR [132] | 2.8 g | N/A | N/A | piezoelectric actuators | Pop-up microelectromechanical systems (MEMS) fabrication process | Energy not on board, jumping height is relatively low | N |
| 2012 | Locust-Inspired [100] | 7 g | 100 cm | 71 cm | Linear spring | Mimic using springs and wire, springs for passive extensor muscles, and a wire as a flexor muscle | The in-air and landing are not considered, lack of autonomy | N |
| 2012 | GRILLO-III [38] | 22 g | 20 cm | 10 cm | Linear spring | continuous jumping, model-based jumping force optimization | Incomplete control systems, lack of autonomy | N |
| 2011 | EPFL [92,93] | 16.5 g | 30.2 cm | 12 cm | Torsional spring, cam | Infrared remote control, reduced impact force, jumping with gliding, high gliding distance | No pitch control, ineffective gliding when jumping from the ground | N |
| 2010 | Buckling [24] | 18 g | 95 cm | 15 cm | Elastic strip, snap | Elastic energy from bending and twisting, high impulsive force frequency | Need of external power supply, incomplete synchronization control | N |
| 2009 | Sand Flea [125] | N/A | 1 | 9 m | CO2 | Astonishing jumping performance and make 25 jumps | A haphazard aerial tumble would preclude it to send back useful video while in midair | Y |
| 2008 | 7 g Miniature [88] | 7 g | N/A | 138 cm | Torsional spring, cam. | Lightweight material, click mechanism, adjustable take-off angle, high endurance | Lack of self-righting mechanism and landing control |
3.3. Take-off Velocity
3.4. Take-off Stability
4. Aerial Righting
4.1. Aerial Righting in Jumping Animals
4.1.1. Aerial Righting by Way of Aerodynamic Torque
4.1.2. Aerial Righting by Way of Inertia
4.1.3. Using Dragline Silk
4.2. Aerial Righting in Jumping Robots
4.2.1. One/Two-DOF Balanced Tail
4.2.2. Gliding Wings
4.2.3. Propellers
5. Landing Buffering and Resetting Mechanisms in Animals and Robots
5.1. Active Resetting Using Support Legs
5.2. Passive Resetting Assisted by Flexible Frame
5.3. Gliding Wings Help Landing
5.4. Elastic Buffering Legs to Absorb Impact
6. Discussion
6.1. Issues to Still Be Addressed
6.1.1. Modeling
6.1.2. Materials
6.1.3. Mechanical Structure
6.1.4. Actuation Mechanisms
6.1.5. Miniaturization
6.1.6. Control Algorithms
6.1.7. High Energy Density
6.1.8. Trajectory Optimization
6.2. Prospects of Jumping Robots
7. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Gordon, M.S.; Blickhan, R.; Dabiri, J.O.; Videler, J.J. Animal Locomotion: Physical Principles and Adaptations; CRC Press; Taylor & Francis Group: Boca Raton, FL, USA, 2017. [Google Scholar] [CrossRef]
- Alexander, R.M. Principles of Animal Locomotion; Princeton University Press: Princeton, NJ, USA, 2002. [Google Scholar] [CrossRef]
- Biewener, A.; Patek, S. Animal Locomotion; Oxford University Press: Oxford, UK, 2018. [Google Scholar]
- Romano, D.; Donati, E.; Benelli, G.; Stefanini, C. A review on animal–robot interaction: From bio-hybrid organisms to mixed societies. Biol. Cybern. 2018, 113, 1–25. [Google Scholar] [CrossRef]
- Taylor, G.; Triantafyllou, M.S.; Tropea, C. Animal Locomotion; Springer: Des Moines, IA, USA, 2010. [Google Scholar] [CrossRef]
- Mammal’s Locomotion. Available online: http://mammals-locomotion.com (accessed on 20 August 2020).
- Sayyad, A.; Seth, B.; Seshu, P. Single-legged hopping robotics research—A review. Robotica 2007, 25, 587–613. [Google Scholar] [CrossRef]
- Kram, R.; Dawson, T.J. Energetics and biomechanics of locomotion by red kangaroos (Macropus rufus). Comp. Biochem. Physiol. Part B Biochem. Mol. Biol. 1998, 120, 41–49. [Google Scholar] [CrossRef]
- Scott, J. The locust jump: An integrated laboratory investigation. Adv. Physiol. Educ. 2005, 29, 21–26. [Google Scholar] [CrossRef]
- Armour, R.; Paskins, K.; Bowyer, A.; Vincent, J.; Megill, W. Jumping robots: A biomimetic solution to locomotion across rough terrain. Bioinspir. Biomim. 2007, 2, S65. [Google Scholar] [CrossRef] [Green Version]
- Wei, D.; Ge, W. Research on one bio-inspired jumping locomotion robot for search and rescue. Int. J. Adv. Robot. Syst. 2014, 11, 168. [Google Scholar] [CrossRef]
- Burdick, J.; Fiorini, P. Minimalist jumping robots for celestial exploration. Int. J. Robot. Res. 2003, 22, 653–674. [Google Scholar] [CrossRef]
- Kaplan, M.H.; Seifert, H.S. Hopping transporters for lunar exploration. J. Spacecr. Rocket. 1969, 6, 917–922. [Google Scholar] [CrossRef]
- Raibert, M.H. Legged Robots That Balance; MIT Press: Cambridge, MA, USA, 1986. [Google Scholar] [CrossRef]
- Berkemeier, M.D.; Fearing, R.S. Sliding and hopping gaits for the underactuated acrobot. IEEE Trans. Robot. Autom. 1998, 14, 629–634. [Google Scholar] [CrossRef]
- Rad, H.; Gregorio, P.; Buehler, M. Design, modeling and control of a hopping robot. In Proceedings of the 1993 IEEE/RSJ International Conference on Intelligent Robots and Systems, Yokohama, Japan, 26–30 July 1993; Volume 3, pp. 1778–1785. [Google Scholar] [CrossRef]
- M’Closkey, R.T.; Burdick, J.W. Periodic motions of a hopping robot with vertical and forward motion. Int. J. Robot. Res. 1993, 12, 197–218. [Google Scholar] [CrossRef]
- Hyon, S.H.; Emura, T. Quasi-periodic gaits of passive one-legged hopper. In Proceedings of the IEEE/RSJ International Conference on Intelligent Robots and Systems, Lausanne, Switzerland, 30 September–4 October 2002; Volume 3, pp. 2625–2630. [Google Scholar] [CrossRef]
- François, C.; Samson, C. A new approach to the control of the planar one-legged hopper. Int. J. Robot. Res. 1998, 17, 1150–1166. [Google Scholar] [CrossRef]
- Brown, B.; Zeglin, G. The bow leg hopping robot. In Proceedings of the 1998 IEEE International Conference on Robotics and Automation (Cat. No. 98CH36146), Leuven, Belgium, 20 May 1998; Volume 1, pp. 781–786. [Google Scholar] [CrossRef]
- Pratt, G.A. Legged robots at MIT: What’s new since Raibert? IEEE Robot. Autom. Mag. 2000, 7, 15–19. [Google Scholar] [CrossRef]
- Mochiyama, H.; Watari, M.; Fujimoto, H. A robotic catapult based on the closed elastica and its application to robotic tasks. In Proceedings of the 2007 IEEE/RSJ International Conference on Intelligent Robots and Systems, San Diego, CA, USA, 29 October–2 November 2007; pp. 1508–1513. [Google Scholar] [CrossRef]
- Yamada, A.; Watari, M.; Mochiyama, H.; Fujimoto, H. An asymmetric robotic catapult based on the closed elastica for jumping robot. In Proceedings of the 2008 IEEE International Conference on Robotics and Automation, Pasadena, CA, USA, 19–23 May 2008; pp. 232–237. [Google Scholar] [CrossRef]
- Yamada, A.; Mameda, H.; Mochiyama, H.; Fujimoto, H. A compact jumping robot utilizing snap-through buckling with bend and twist. In Proceedings of the 2010 IEEE/RSJ International Conference on Intelligent Robots and Systems, Taipei, Taiwan, 18–22 October 2010; pp. 389–394. [Google Scholar] [CrossRef]
- Yamada, A.; Watari, M.; Mochiyama, H.; Fujimoto, H. A robotic catapult based on the closed elastica with a high stiffness endpoint and its application to swimming tasks. In Proceedings of the 2008 IEEE/RSJ International Conference on Intelligent Robots and Systems, Nice, France, 22–26 September 2008; pp. 1477–1482. [Google Scholar] [CrossRef]
- Tsuda, T.; Mochiyama, H.; Fujimoto, H. Quick stair-climbing using snap-through buckling of closed elastica. In Proceedings of the 2012 International Symposium on Micro-NanoMechatronics and Human Science, Nagoya, Japan, 4–7 November 2012; pp. 368–373. [Google Scholar] [CrossRef]
- Scarfogliero, U.; Stefanini, C.; Dario, P. The use of compliant joints and elastic energy storage in bio-inspired legged robots. Mech. Mach. Theory 2009, 44, 580–590. [Google Scholar] [CrossRef]
- Wei, D.; Gao, T.; Li, Z.; Mo, X.; Zheng, S.; Zhou, C. Hybrid Inspired Research on the Flying-Jumping Locomotion of Locusts Using Robot Counterpart. Front. Neurorobot. 2019, 13, 87. [Google Scholar] [CrossRef]
- Sugiyama, Y.; Hirai, S. Crawling and jumping by a deformable robot. Int. J. Robot. Res. 2006, 25, 603–620. [Google Scholar] [CrossRef]
- Matsuyama, Y.; Hirai, S. Analysis of circular robot jumping by body deformation. In Proceedings of the 2007 IEEE International Conference on Robotics and Automation, Roma, Italy, 10–14 April 2007; pp. 1968–1973. [Google Scholar] [CrossRef]
- Sugiyama, Y.; Shiotsu, A.; Yamanaka, M.; Hirai, S. Circular/spherical robots for crawling and jumping. In Proceedings of the 2005 IEEE International Conference on Robotics and Automation, Barcelona, Spain, 18–22 April 2005; pp. 3595–3600. [Google Scholar] [CrossRef]
- Bartlett, N.W.; Tolley, M.T.; Overvelde, J.T.; Weaver, J.C.; Mosadegh, B.; Bertoldi, K.; Whitesides, G.M.; Wood, R.J. A 3D-printed, functionally graded soft robot powered by combustion. Science 2015, 349, 161–165. [Google Scholar] [CrossRef] [Green Version]
- Wang, H.; Luan, Y.; Oetomo, D.; Wang, Z. Design, analysis and experimental evaluation of a gas-fuel-powered actuator for robotic hoppers. IEEE ASME Trans. Mechatron. 2015, 20, 2264–2275. [Google Scholar] [CrossRef]
- Miao, Z.; Mo, J.; Li, G.; Ning, Y.; Li, B. Wheeled hopping robot with combustion-powered actuator. Int. J. Adv. Robot. Syst. 2018, 15, 1729881417745608. [Google Scholar] [CrossRef]
- Shepherd, R.F.; Stokes, A.A.; Freake, J.; Barber, J.; Snyder, P.W.; Mazzeo, A.D.; Cademartiri, L.; Morin, S.A.; Whitesides, G.M. Using explosions to power a soft robot. Angew. Chem. Int. Edit. 2013, 125, 2964–2968. [Google Scholar] [CrossRef]
- Loepfe, M.; Schumacher, C.M.; Lustenberger, U.B.; Stark, W.J. An untethered, jumping roly-poly soft robot driven by combustion. Soft Robot. 2015, 2, 33–41. [Google Scholar] [CrossRef]
- Hu, W.; Lum, G.Z.; Mastrangeli, M.; Sitti, M. Small-scale soft-bodied robot with multimodal locomotion. Nature 2018, 554, 81. [Google Scholar] [CrossRef]
- Li, F.; Liu, W.; Fu, X.; Bonsignori, G.; Scarfogliero, U.; Stefanini, C.; Dario, P. Jumping like an insect: Design and dynamic optimization of a jumping mini robot based on bio-mimetic inspiration. Mechatronics 2012, 22, 167–176. [Google Scholar] [CrossRef]
- Picardi, G.; Laschi, C.; Calisti, M. Model-based open loop control of a multigait legged underwater robot. Mechatronics 2018, 55, 162–170. [Google Scholar] [CrossRef]
- Scarfogliero, U.; Li, F.; Chen, D.; Stefanini, C.; Liu, W.; Dario, P. Jumping mini-robot as a model of scale effects on legged locomotion. In Proceedings of the 2007 IEEE International Conference on Robotics and Biomimetics, Sanya, China, 15–18 December 2007; pp. 853–858. [Google Scholar] [CrossRef]
- Graichen, K.; Hentzelt, S.; Hildebrandt, A.; Kärcher, N.; Gaißert, N.; Knubben, E. Control design for a bionic kangaroo. Control Eng. Pract. 2015, 42, 106–117. [Google Scholar] [CrossRef]
- Sutton, G.P.; Doroshenko, M.; Cullen, D.A.; Burrows, M. Take-off speed in jumping mantises depends on body size and a power-limited mechanism. J. Exp. Biol. 2016, 219, 2127–2136. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nabawy, M.R.; Sivalingam, G.; Garwood, R.J.; Crowther, W.J.; Sellers, W.I. Energy and time optimal trajectories in exploratory jumps of the spider Phidippus. Regius. Sci. Rep. 2018, 8, 7142. [Google Scholar] [CrossRef]
- Ribak, G.; Weihs, D. Jumping without using legs: The jump of the click-beetles (Elateridae) is morphologically constrained. PLoS ONE 2011, 6, e20871. [Google Scholar] [CrossRef] [Green Version]
- Christian, V.E. The jump of the springtails. Naturwissenschaften 1978, 65, 495–496. [Google Scholar] [CrossRef]
- Burrows, M.; Dorosenko, M. Jumping mechanisms in adult caddisflies (Insecta, Trichoptera). J. Exp. Biol. 2015, 218, 2764–2774. [Google Scholar] [CrossRef] [Green Version]
- Burrows, M.; Dorosenko, M. Jumping mechanisms and strategies in moths (Lepidoptera). J. Exp. Biol. 2015, 218, 1655–1666. [Google Scholar] [CrossRef] [Green Version]
- Mo, X.; Ge, W.; Romano, D.; Donati, E.; Benelli, G.; Dario, P.; Stefanini, C. Modelling jumping in Locusta migratoria and the influence of substrate roughness. Entomol. Gen. 2019, 38, 317–332. [Google Scholar] [CrossRef]
- Developing Flies Jump without Legs. Available online: https://phys.org/news/2018-01-flies-legs.html (accessed on 20 August 2020).
- This Legless Insect Can Jump 30 Times Its Body Length. Available online: http://www.sciencemag.org/news/2018/01/legless-insect-can-jump-30-times-its-body-length (accessed on 20 August 2020).
- Brown, R.H.J. Mechanism of locust jumping. Nature 1967, 214, 939. [Google Scholar] [CrossRef]
- Cofer, D.W.; Reid, J.; Zhu, Y.; Cymbalyuk, G.; Heitler, W.J.; Edwards, D.H. Role of the semi-lunar process in locust jumping. BMC Neurosci. 2007, 8, P12. [Google Scholar] [CrossRef] [Green Version]
- Snelling, E.P.; Becker, C.L.; Seymour, R.S. The effects of temperature and body mass on jump performance of the locust Locusta migratoria. PLoS ONE 2013, 8, e72471. [Google Scholar] [CrossRef] [Green Version]
- Bennet-Clark, H.C.; Lucey, E.C.A. The jump of the flea: A study of the energetics and a model of the mechanism. J. Exp. Biol. 1967, 47, 59–76. [Google Scholar] [CrossRef]
- Sutton, G.P.; Burrows, M. Biomechanics of jumping in the flea. J. Exp. Biol. 2011, 214, 836–847. [Google Scholar] [CrossRef] [Green Version]
- Picker, M.D.; Colville, J.F.; Burrows, M. A cockroach that jumps. Biol. Lett. 2012, 8, 390–392. [Google Scholar] [CrossRef] [Green Version]
- Bohn, H.; Picker, M.; Klass, K.; Colville, J. A jumping cockroach from South Africa. Saltoblattella montistabularis, gen. nov., spec. nov. (Blattodea: Blattellidae). Arthropod. Syst. Phylogeny 2010, 68, 53–69. [Google Scholar] [CrossRef]
- Tauber, E.R.A.N.; Camhi, J. The wind-evoked escape behavior of the cricket Gryllus bimaculatus: Integration of behavioral elements. J. Exp. Biol. 1995, 198, 1895–1907. [Google Scholar]
- Burrows, M. Jumping performance of froghopper insects. J. Exp. Biol. 2006, 209, 4607–4621. [Google Scholar] [CrossRef] [Green Version]
- Burrows, M. Morphology and action of the hind leg joints controlling jumping in froghopper insects. J. Exp. Biol. 2006, 209, 4622–4637. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burrows, M. Kinematics of jumping in leafhopper insects (Hemiptera, Auchenorrhyncha, Cicadellidae). J. Exp. Biol. 2007, 210, 3579–3589. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burrows, M. Anatomy of the hind legs and actions of their muscles during jumping in leafhopper insects. J. Exp. Biol. 2007, 210, 3590–3600. [Google Scholar] [CrossRef] [Green Version]
- Bonsignori, G.; Stefanini, C.; Scarfogliero, U.; Mintchev, S.; Benelli, G.; Dario, P. The green leafhopper, Cicadella viridis (Hemiptera, Auchenorrhyncha, Cicadellidae), jumps with near-constant acceleration. J. Exp. Biol. 2013, 216, 1270–1279. [Google Scholar] [CrossRef] [Green Version]
- Burrows, M. Jumping strategies and performance in shore bugs (Hemiptera, Heteroptera, Saldidae). J. Exp. Biol. 2009, 212, 106–115. [Google Scholar] [CrossRef] [Green Version]
- Burrows, M.; Morris, O. Jumping and kicking in bush crickets. J. Exp. Biol. 2003, 206, 1035–1049. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.; Liao, C.; Tsai, F.; Chi, K. More than a safety line: Jump-stabilizing silk of salticids. J. R. Soc. Interface 2013, 10, 20130572. [Google Scholar] [CrossRef] [Green Version]
- Weihmann, T.; Karner, M.; Full, R.J.; Blickhan, R. Jumping kinematics in the wandering spider Cupiennius salei. J. Comp. Physiol. A 2010, 196, 421–438. [Google Scholar] [CrossRef]
- Parry, D.A.; Brown, R.H.J. The jumping mechanism of salticid spiders. J. Exp. Biol. 1959, 36, 654–664. [Google Scholar] [CrossRef]
- Evans, M.E.G. The jump of the click beetle (Coleoptera, Elateridae)—A preliminary study. J. Zool. 1972, 167, 319–336. [Google Scholar] [CrossRef]
- Evans, M.E.G. The jump of the click beetle (Coleoptera: Elateridae)—Energetics and mechanics. J. Zool. 1973, 169, 181–194. [Google Scholar] [CrossRef]
- Bolmin, O.; Duan, C.; Urrutia, L.; Abdulla, A.M.; Hazel, A.M.; Alleyne, M.; Dunn, A.C.; Wissa, A. Pop! Observing and Modeling the Legless Self-righting Jumping Mechanism of Click Beetles. In Proceedings of the Conference on Biomimetic and Biohybrid Systems, Stanford, CA, USA, 26–28 July 2017; pp. 35–47. [Google Scholar] [CrossRef]
- Brackenbury, J.; Hunt, H. Jumping in springtails: Mechanism and dynamics. J. Zool. 1993, 229, 217–236. [Google Scholar] [CrossRef]
- Faraji, H.; Tachella, R.; Hatton, R.L. Aiming and vaulting: Spider inspired leaping for jumping robots. In Proceedings of the 2016 IEEE International Conference on Robotics and Automation, Stockholm, Sweden, 16–21 May 2016; pp. 2082–2087. [Google Scholar] [CrossRef]
- Tachella, R. Design and Development of a Salticid Inspired Jumping Robot. Master’s Thesis, Oregon State University, Corvallis, OR, USA, 2016. [Google Scholar]
- Hill, D.E. Targeted Jumps by Salticid Spiders (Araneae, Salticidae, Phidippus), Version 9; Peckhamia Epublications, 2006; pp. 1–28. [Google Scholar]
- Spröwitz, A.; Göttler, C.; Sinha, A.; Caer, C.; Öoztekin, M.U.; Petersen, K.; Sitti, M. Scalable pneumatic and tendon driven robotic joint inspired by jumping spiders. In Proceedings of the 2017 IEEE International Conference on Robotics and Automation, Singapore, 29 May–3 June 2017; pp. 64–70. [Google Scholar] [CrossRef]
- Zhu, Y.; Chen, L.; Liu, Q.; Qin, R.; Jin, B. Omnidirectional Jump of a Legged Robot Based on the Behavior Mechanism of a Jumping Spider. Appl. Sci. 2018, 8, 51. [Google Scholar] [CrossRef] [Green Version]
- Shield, S.; Fisher, C.; Patel, A. A spider-inspired dragline enables aerial pitch righting in a mobile robot. In Proceedings of the 2015 IEEE/RSJ International Conference on Intelligent Robots and Systems, Hamburg, Germany, 28 September–2 October 2015; pp. 319–324. [Google Scholar] [CrossRef]
- Albrecht, F.O. The Anatomy of the Migratory Locust; The Athlone Press: London, UK, 1953. [Google Scholar]
- Mo, X.; Romano, D.; Miraglia, M.; Ge, W.; Stefanini, C. Effect of substrates’ compliance on the jumping mechanism of Locusta Migratoria. Front. Bioeng. Biotechnol. 2020, 8, 661. [Google Scholar] [CrossRef]
- Romano, D.; Bloemberg, J.; Tannous, M.; Stefanini, C. Impact of aging and cognitive mechanisms on high-speed motor activation patterns: Evidence from an orthoptera-robot interaction. IEEE Trans. Med. Robot. Bionics 2020, 2, 292–296. [Google Scholar] [CrossRef]
- Mo, X.; Romano, D.; Milazzo, M.; Benelli, G.; Stefanini, C. Impact of different developmental instars on Locusta Migratoria jumping performance. Appl. Bionics Biomech. 2020, 2020, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Han, L.; Wang, Z.; Ji, A.; Dai, Z. The mechanics and trajectory control in locust jumping. J. Bionic Eng. 2013, 10, 194–200. [Google Scholar] [CrossRef]
- Chen, D.; Yin, J.; Chen, K.; Li, Z. Biomechanical and dynamic mechanism of locust take-off. Acta Mech. Sin. 2014, 30, 762–774. [Google Scholar] [CrossRef]
- Heitler, W.J. The locust jump: III. Structural specializations of the metathoracic tibiae. J. Exp. Biol. 1977, 67, 29–36. [Google Scholar]
- Heitler, W.J. Locust jump. Specialisations of the metathoracic femoral-tibial joint. J. Comp. Physiol. 1974, 89, 93–104. [Google Scholar] [CrossRef]
- Heitler, W.J.; Burrows, M. The locust jump. I. The motor programme. J. Exp. Biol. 1977, 66, 203–219. [Google Scholar]
- Kovac, M.; Fuchs, M.; Guignard, A.; Zufferey, J.C.; Floreano, D. A miniature 7 g jumping robot. In Proceedings of the 2008 IEEE International Conference on Robotics and Automation, Pasadena, CA, USA, 19–23 May 2008; pp. 373–378. [Google Scholar] [CrossRef] [Green Version]
- Kovac, M. Bioinspired Jumping Locomotion for Miniature Robotics. Ph.D. Thesis, Swiss Federal Institute of Technology in Lausanne, Lausanne, Switzerland, 2010. [Google Scholar] [CrossRef]
- Kovač, M.; Schlegel, M.; Zufferey, J.C.; Floreano, D. A miniature jumping robot with self-recovery capabilities. In Proceedings of the 2009 IEEE/RSJ International Conference on Intelligent Robots and Systems, St. Louis, MO, USA, 10–15 October 2009; pp. 583–588. [Google Scholar] [CrossRef] [Green Version]
- Vidyasagar, A.; Zufferey, J.C.; Floreano, D.; Kovač, M. Performance analysis of jump-gliding locomotion for miniature robotics. Bioinspir. Biomim. 2015, 10, 025006. [Google Scholar] [CrossRef]
- Kovač, M.; Fauria, O.; Zufferey, J.C.; Floreano, D. The EPFL jumpglider: A hybrid jumping and gliding robot with rigid or folding wings. In Proceedings of the 2011 IEEE International Conference on Robotics and Biomimetics, Phuket, Thailand, 7–11 December 2011; pp. 1503–1508. [Google Scholar] [CrossRef] [Green Version]
- Kovač, M.; Fauria, O.; Zufferey, J.C.; Floreano, D. The locomotion capabilities of the EPFL jumpglider: A hybrid jumping and gliding robot. In Proceedings of the 2011 IEEE International Conference on Robotics and Biomimetics, Phuket, Thailand, 7–11 December 2011; pp. 2249–2250. [Google Scholar] [CrossRef]
- Kovač, M.; Schlegel, M.; Zufferey, J.C.; Floreano, D. Steerable miniature jumping robot. Auton. Robot. 2010, 28, 295–306. [Google Scholar] [CrossRef] [Green Version]
- Zaitsev, V.; Gvirsman, O.; Ben-Hanan, U.; Weiss, A.; Ayali, A.; Kósa, G. Locust-inspired miniature jumping robot. In Proceedings of the 2015 IEEE/RSJ International Conference on Intelligent Robots and Systems, Hamburg, Germany, 28 September–2 October 2015; pp. 553–558. [Google Scholar] [CrossRef]
- Zaitsev, V.; Gvirsman, O.; Hanan, U.B.; Weiss, A.; Ayali, A.; Kosa, G. A locust-inspired miniature jumping robot. Bioinspir. Biomim. 2015, 10, 066012. [Google Scholar] [CrossRef]
- Hanan, U.B.; Weiss, A.; Zaitsev, V. Jumping efficiency of small creatures and its applicability in robotics. Procedia Manuf. 2018, 21, 243–250. [Google Scholar] [CrossRef]
- Beck, A.; Zaitsev, V.; Hanan, U.B.; Kosa, G.; Ayali, A.; Weiss, A. Jump stabilization and landing control by wing-spreading of a locust-inspired jumper. Bioinspir. Biomim. 2017, 12, 066006. [Google Scholar] [CrossRef]
- Weiss, A.; Zaitsev, V.; Nabi, N.; Ben-Hanan, U. Landing recovery and orientation control of a locust-inspired miniature jumping robot. Eng. Res. Express 2020, 2, 015017. [Google Scholar] [CrossRef]
- Nguyen, Q.V.; Park, H.C. Design and demonstration of a locust-like jumping mechanism for small-scale robots. J. Bionic Eng. 2012, 9, 271–281. [Google Scholar] [CrossRef]
- Zhang, J.; Song, G.; Li, Y.; Qiao, G.; Song, A.; Wang, A. A bio-inspired jumping robot: Modeling, simulation, design, and experimental results. Mechatronics 2013, 23, 1123–1140. [Google Scholar] [CrossRef]
- Zhang, J.; Song, G.; Qiao, G.; Li, Z.; Wang, W.; Song, A. A novel one-motor driven robot that jumps and walks. In Proceedings of the 2013 IEEE International Conference on Robotics and Automation, Karlsruhe, Germany, 6–10 May 2013; pp. 13–19. [Google Scholar] [CrossRef]
- Zhang, J.; Zhu, Y.; Wang, H.; Zhang, J. Design of a bio-inspired jumping robot for rough terrain. In Proceedings of the 2010 International Conference on Environmental Science and Information Application Technology, Wuhan, China, 17–18 July 2010; pp. 40–43. [Google Scholar] [CrossRef]
- Zhang, J.; Song, G.; Li, Z.; Qiao, G.; Sun, H.; Song, A. Self-righting, steering and take-off angle adjusting for a jumping robot. In Proceedings of the 2012 IEEE/RSJ International Conference on Intelligent Robots and Systems, Vilamoura, Portugal, 7–12 October 2012; pp. 2089–2094. [Google Scholar] [CrossRef]
- Zhang, J.; Song, G.; Qiao, G.; Li, Z.; Wang, A. A wireless sensor network system with a jumping node for unfriendly environments. Int. J. Distrib. Sens. Netw. 2012, 8, 568240. [Google Scholar] [CrossRef]
- Zhang, J.; Song, G.; Qiao, G.; Meng, T.; Sun, H. An indoor security system with a jumping robot as the surveillance terminal. IEEE Trans. Consum. Electr. 2011, 57, 1774–1781. [Google Scholar] [CrossRef]
- Chen, D.; Yin, J.; Huang, Y.; Zhao, K.; Wang, T. A hopping-righting mechanism analysis and design of the mobile robot. J. Braz. Soc. Mech. Sci. Eng. 2013, 35, 469–478. [Google Scholar] [CrossRef]
- Chen, D.; Yin, J.; Zhao, K.; Zheng, W.; Wang, T. Bionic mechanism and kinematics analysis of hopping robot inspired by locust jumping. J. Bionic Eng. 2011, 8, 429–439. [Google Scholar] [CrossRef]
- Chen, D.S.; Zhang, Z.Q.; Chen, K.W. Dynamic model and performance analysis of landing buffer for bionic locust mechanism. Acta Mech. Sin. 2016, 32, 551–565. [Google Scholar] [CrossRef]
- Chen, D.; Zhang, Z.; Chen, K. Legs attitudes determination for bionic locust robot based on landing buffering performance. Mech. Mach. Theory 2016, 99, 117–139. [Google Scholar] [CrossRef]
- Chen, D.; Chen, K.; Zhang, Z.; Zhang, B. Mechanism of locust air posture adjustment. J. Bionic Eng. 2015, 12, 418–431. [Google Scholar] [CrossRef]
- Chen, D.; Yin, J.; Chen, K.; Zhao, K.; Zhang, B. Prototype design and experimental study on locust air-posture righting. J. Bionic Eng. 2014, 11, 459–468. [Google Scholar] [CrossRef]
- Zhang, Z.; Yang, Q.; Gui, S.; Chang, B.; Zhao, J.; Yang, H.; Chen, D. Mechanism design for locust-inspired robot with one-DOF leg based on jumping stability. Mech. Mach. Theory 2019, 133, 584–605. [Google Scholar] [CrossRef]
- Cadiergues, M.C.; Joubert, C.; Franc, M. A comparison of jump performances of the dog flea, Ctenocephalides canis (Curtis, 1826) and the cat flea, Ctenocephalides felis (Bouché, 1835). Vet. Parasitol. 2000, 92, 239–241. [Google Scholar] [CrossRef]
- Rothschild, M.; Schlein, J. The jumping mechanism of Xenopsylla Cheopis. I. exoskeletal structures and musculature. Philos. Trans. R. Soc. Lond. B Biol. Sci. 1975, 271, 457–490. [Google Scholar] [CrossRef]
- Cullen, M.J. The jumping mechanism of Xenopsylla Cheopis II. The fine structure of the jumping muscle. Philos. Trans. R. Soc. Lond. B Biol. Sci. 1975, 271, 491–497. [Google Scholar] [CrossRef] [PubMed]
- Rothschild, M.L.; Schlein, J.; Parker, K.; Neville, C.; Sternberg, S. The jumping mechanism of Xenopsylla cheopis III. Execution of the jump and activity. Philos. Trans. R. Soc. Lond. B Biol. Sci. 1975, 271, 499–515. [Google Scholar] [CrossRef] [PubMed]
- Noh, M.; Kim, S.W.; An, S.; Koh, J.S.; Cho, K.J. Flea-inspired catapult mechanism for miniature jumping robots. IEEE Trans. Robot. 2012, 28, 1007–1018. [Google Scholar] [CrossRef]
- Jung, G.P.; Kim, J.S.; Koh, J.S.; Jung, S.P.; Cho, K.J. Role of compliant leg in the flea-inspired jumping mechanism. In Proceedings of the 2014 IEEE/RSJ International Conference on Intelligent Robots and Systems, Chicago, IL, USA, 14–18 September 2014; pp. 315–320. [Google Scholar] [CrossRef]
- Jung, G.P.; Choi, H.C.; Cho, K.J. The effect of leg compliance in multi-directional jumping of a flea-inspired mechanism. Bioinspir. Biomim. 2017, 12, 026006. [Google Scholar] [CrossRef] [PubMed]
- Koh, J.S.; Jung, S.P.; Wood, R.J.; Cho, K.J. A jumping robotic insect based on a torque reversal catapult mechanism. In Proceedings of the 2013 IEEE/RSJ International Conference on Intelligent Robots and Systems, Tokyo, Japan, 3–7 November 2013; pp. 3796–3801. [Google Scholar] [CrossRef]
- Jung, S.P.; Jung, G.P.; Koh, J.S.; Lee, D.Y.; Cho, K.J. Fabrication of Composite and Sheet Metal Laminated Bistable Jumping Mechanism. J. Mech. Robot. 2015, 7, 021010. [Google Scholar] [CrossRef]
- Koh, J.S.; Jung, S.P.; Noh, M.; Kim, S.W.; Cho, K.J. Flea inspired catapult mechanism with active energy storage and release for small scale jumping robot. In Proceedings of the 2013 IEEE International Conference on Robotics and Automation, Karlsruhe, Germany, 6–10 May 2013; pp. 26–31. [Google Scholar] [CrossRef]
- Urban Hopper. Available online: https://www.sandia.gov/research/robotics/unique_mobility/urban_hopper.html (accessed on 20 August 2020).
- Ackerman, E. Boston dynamics sand flea robot demonstrates astonishing jumping skills. IEEE Spectr. Robot. Blog. 2012, 2. [Google Scholar]
- Laksanacharoen, S.; Pollack, A.J.; Nelson, G.M.; Quinn, R.D.; Ritzmann, R.E. Biomechanics and simulation of cricket for microrobot design. In Proceedings of the IEEE International Conference on Robotics and Automation, San Francisco, CA, USA, 24–28 April 2000; Volume 2, pp. 1088–1094. [Google Scholar] [CrossRef]
- Birch, M.C.; Quinn, R.D.; Hahm, G.; Phillips, S.M.; Drennan, B.; Fife, A.; Verma, H.; Beer, R.D. Design of a cricket microrobot. In Proceedings of the IEEE International Conference on Robotics and Automation, San Francisco, CA, USA, 24–28 April 2000; Volume 2, pp. 1109–1114. [Google Scholar] [CrossRef]
- Birch, M.C.; Quinn, R.D.; Hahm, G.; Phillips, S.M.; Drennan, B.T.; Fife, A.J.; Pollack, A.J.; Beer, R.; Yu, X.; Garverick, S.; et al. Cricket-based robots. IEEE Robot. Autom. Mag. 2002, 9, 20–30. [Google Scholar] [CrossRef]
- Allen, T.J.; Quinn, R.D.; Bachmann, R.J.; Ritzmann, R.E. Abstracted biological principles applied with reduced actuation improve mobility of legged vehicles. In Proceedings of the 2003 IEEE/RSJ International Conference on Intelligent Robots and Systems, Las Vegas, NV, USA, 27–31 October 2003; Volume 2, pp. 1370–1375. [Google Scholar] [CrossRef]
- Lambrecht, B.G.; Horchler, A.D.; Quinn, R.D. A small, insect-inspired robot that runs and jumps. In Proceedings of the 2005 IEEE International Conference on Robotics and Automation, Barcelona, Spain, 18–22 April 2005; pp. 1240–1245. [Google Scholar] [CrossRef]
- Jayaram, K.; Full, R.J. Cockroaches traverse crevices, crawl rapidly in confined spaces, and inspire a soft, legged robot. Proc. Natl. Acad. Sci. USA 2016, 113, 950–957. [Google Scholar] [CrossRef] [Green Version]
- HAMR: Versatile Crawling Microrobot. Available online: https://wyss.harvard.edu/technology/versatile-ambulatory-microrobots/ (accessed on 20 August 2020).
- Chen, Y.; Doshi, N.; Goldberg, B.; Wang, H.; Wood, R.J. Controllable water surface to underwater transition through electrowetting in a hybrid terrestrial-aquatic microrobot. Nat. Commun. 2018, 9, 2495. [Google Scholar] [CrossRef]
- Sutton, G.P.; Burrows, M. The mechanics of azimuth control in jumping by froghopper insects. J. Exp. Biol. 2010, 213, 1406–1416. [Google Scholar] [CrossRef] [Green Version]
- Jung, G.P.; Cho, K.J. Froghopper-inspired direction-changing concept for miniature jumping robots. Bioinspir. Biomim. 2016, 11, 056015. [Google Scholar] [CrossRef]
- Scarfogliero, U.; Stefanini, C.; Dario, P. A bioinspired concept for high efficiency locomotion in micro robots: The jumping robot grillo. In Proceedings of the 2006 IEEE International Conference on Robotics and Automation, Orlando, FL, USA, 15–19 May 2006; pp. 4037–4042. [Google Scholar] [CrossRef] [Green Version]
- Scarfogliero, U.; Stefanini, C.; Dario, P. Design and Development of the Long-Jumping “Grillo” Mini Robot. In Proceedings of the 2007 IEEE International Conference on Robotics and Automation, Roma, Italy, 10–14 April 2007; pp. 467–472. [Google Scholar] [CrossRef]
- Li, F.; Bonsignori, G.; Scarfogliero, U.; Chen, D.; Stefanini, C.; Liu, W.; Dario, P.; Fu, X. Jumping mini-robot with bio-inspired legs. In Proceedings of the IEEE International Conference on Robotics and Biomimetics, Bangkok, Thailand, 22–25 February 2009; pp. 933–938. [Google Scholar] [CrossRef]
- Marsh, R.L.; John-Alder, H.B. Jumping performance of hylid frogs measured with high-speed cine film. J. Exp. Biol. 1994, 188, 131–141. [Google Scholar] [CrossRef]
- Azizi, E.; Roberts, T.J. Muscle performance during frog jumping: Influence of elasticity on muscle operating lengths. Proc. R. Soc. B Biol. Sci. 2010, 277, 1523–1530. [Google Scholar] [CrossRef] [Green Version]
- Astley, H.C.; Roberts, T.J. Evidence for a vertebrate catapult: Elastic energy storage in the plantaris tendon during frog jumping. Biol. Lett. 2011, 8, 386–389. [Google Scholar] [CrossRef] [Green Version]
- Astley, H.C.; Roberts, T.J. The mechanics of elastic loading and recoil in anuran jumping. J. Exp. Biol. 2014, 217, 4372–4378. [Google Scholar] [CrossRef] [Green Version]
- Wang, M.; Zang, X.Z.; Fan, J.Z.; Zhao, J. Biological jumping mechanism analysis and modeling for frog robot. J. Bionic Eng. 2008, 5, 181–188. [Google Scholar] [CrossRef]
- Zhong, J.; Luo, M.; Liu, X.; Fan, J.; Zhao, J. Frog-inspired jumping robot actuated by pneumatic muscle actuators. Adv. Mech. Eng. 2018, 10, 1687814018782303. [Google Scholar] [CrossRef] [Green Version]
- Fiorini, P.; Hayati, S.; Heverly, M.; Gensler, J. A hopping robot for planetary exploration. In Proceedings of the IEEE Aerospace Conference, Aspen, CO, USA, 7 March 1999; Volume 2, pp. 153–158. [Google Scholar] [CrossRef]
- Fiorini, P.; Burdick, J. The development of hopping capabilities for small robots. Auton. Robot. 2003, 14, 239–254. [Google Scholar] [CrossRef]
- Ahn, J.; Park, J.; Kim, K.S.; Kim, S. Frog-inspired jumping robot for overcoming high obstacles. In Proceedings of the 2013 IEEE International Symposium on Robotics, Seoul, South Korea, 24–26 October 2013; pp. 1–4. [Google Scholar] [CrossRef]
- Jung, G.P.; Casarez, C.S.; Jung, S.P.; Fearing, R.S.; Cho, K.J. An integrated jumping-crawling robot using height-adjustable jumping module. In Proceedings of the 2016 IEEE International Conference on Robotics and Automation, Stockholm, Sweden, 16–21 May 2016; pp. 4680–4685. [Google Scholar] [CrossRef]
- Yim, S.; Baek, S.M.; Jung, G.P.; Cho, K.J. An Omnidirectional Jumper with Expanded Movability via Steering, Self-Righting and Take-off Angle Adjustment. In Proceedings of the 2018 IEEE/RSJ International Conference on Intelligent Robots and Systems, Madrid, Spain, 1–5 October 2018; pp. 416–421. [Google Scholar] [CrossRef]
- Woodward, M.A.; Sitti, M. Design of a miniature integrated multi-modal jumping and gliding robot. In Proceedings of the 2011 IEEE/RSJ International Conference on Intelligent Robots and Systems, San Francisco, CA, USA, 25–30 September 2011; pp. 556–561. [Google Scholar] [CrossRef]
- Woodward, M.A.; Sitti, M. Multimo-bat: A biologically inspired integrated jumping–gliding robot. Int. J. Rob. Res. 2014, 33, 1511–1529. [Google Scholar] [CrossRef]
- Woodward, M.A.; Sitti, M. Morphological intelligence counters foot slipping in the desert locust and dynamic robots. Proc. Natl. Acad. Sci. USA 2018, 115, E8358–E8367. [Google Scholar] [CrossRef] [Green Version]
- Kim, H.; Woodward, M.A.; Sitti, M. Enhanced Non-Steady Gliding Performance of the MultiMo-Bat through Optimal Airfoil Configuration and Control Strategy. In Proceedings of the 2018 IEEE/RSJ International Conference on Intelligent Robots and Systems, Madrid, Spain, 1–5 October 2018; pp. 1382–1388. [Google Scholar] [CrossRef]
- Alexander, R.M. Tendon elasticity and muscle function. Comp. Biochem. Physiol. A Mol. Integr. Physiol. 2002, 133, 1001–1011. [Google Scholar] [CrossRef]
- Hall-Crags, E.C.B. An analysis of the jump of the lesser galago (Galago senegalensis). In Proceedings of the Zoological Society of London; Blackwell Publishing Ltd.: Oxford, UK, 1965; Volume 147, pp. 20–29. [Google Scholar] [CrossRef]
- Aerts, P. Vertical jumping in Galago senegalensis: The quest for an obligate mechanical power amplifier. Philos. Trans. R. Soc. Lond. B Biol. Sci. 1998, 353, 1607–1620. [Google Scholar] [CrossRef] [Green Version]
- Haldane, D.W.; Plecnik, M.; Yim, J.K.; Fearing, R.S. A power modulating leg mechanism for monopedal hopping. In Proceedings of the 2016 IEEE/RSJ International Conference on Intelligent Robots and Systems, Daejeon, Korea, 9–14 October 2016; pp. 4757–4764. [Google Scholar] [CrossRef]
- Haldane, D.W.; Plecnik, M.M.; Yim, J.K.; Fearing, R.S. Robotic vertical jumping agility via series-elastic power modulation. Sci. Robot. 2016, 1, eaag2048. [Google Scholar] [CrossRef] [Green Version]
- Plecnik, M.M.; Haldane, D.W.; Yim, J.K.; Fearing, R.S. Design exploration and kinematic tuning of a power modulating jumping monopod. J. Mech. Robot. 2017, 9, 011009. [Google Scholar] [CrossRef]
- Haldane, D.W.; Yim, J.K.; Fearing, R.S. Repetitive extreme-acceleration (14-g) spatial jumping with Salto-1P. In Proceedings of the 2017 IEEE/RSJ International Conference on Intelligent Robots and Systems, Vancouver, BC, Canada, 24–28 September 2017; pp. 3345–3351. [Google Scholar] [CrossRef]
- Yim, J.K.; Fearing, R.S. Precision Jumping Limits from Flight-phase Control in Salto-1P. In Proceedings of the 2018 IEEE/RSJ International Conference on Intelligent Robots and Systems, Madrid, Spain, 1–5 October 2018; pp. 2229–2236. [Google Scholar] [CrossRef]
- Why is Australia so Hopping Mad? Available online: https://www.australiangeographic.com.au/blogs/wild-journey/2016/11/why-do-australian-animals-hop/ (accessed on 20 August 2020).
- McGowan, C.P.; Collins, C.E. Why do mammals hop? Understanding the ecology, biomechanics and evolution of bipedal hopping. J. Exp. Biol. 2018, 221, jeb161661. [Google Scholar] [CrossRef] [Green Version]
- Dawson, T.J.; Taylor, C.R. Energetic cost of locomotion in kangaroos. Nature 1973, 246, 313. [Google Scholar] [CrossRef]
- Alexander, R.M.; Vernon, A. The mechanics of hopping by kangaroos (Macropodidae). J. Zool. 1975, 177, 265–303. [Google Scholar] [CrossRef]
- Bennett, M.B. Fast locomotion of some kangaroos. J. Zool. 1987, 212, 457–464. [Google Scholar] [CrossRef]
- Festo’s Newest Robot Is a Hopping Bionic Kangaroo. Available online: https://spectrum.ieee.org/automaton/robotics/robotics-hardware/festo-newest-robot-is-a-hopping-bionic-kangaroo (accessed on 20 August 2020).
- Liu, G.H.; Lin, H.Y.; Lin, H.Y.; Chen, S.T.; Lin, P.C. Design of a kangaroo robot with dynamic jogging locomotion. In Proceedings of the 2013 IEEE/SICE International Symposium on System Integration, Kobe, Japan, 15–17 December 2013; pp. 306–311. [Google Scholar] [CrossRef]
- Liu, G.H.; Lin, H.Y.; Lin, H.Y.; Chen, S.T.; Lin, P.C. A bio-inspired hopping kangaroo robot with an active tail. J. Bionic Eng. 2014, 11, 541–555. [Google Scholar] [CrossRef]
- Jun, B.R.; Kim, Y.J.; Jung, S. Design and control of jumping mechanism for a Kangaroo-inspired robot. In Proceedings of the 2016 6th IEEE International Conference on Biomedical Robotics and Biomechatronics, Singapore, 26–29 June 2016; pp. 436–440. [Google Scholar] [CrossRef]
- Spägele, T.; Kistner, A.; Gollhofer, A. Modelling, simulation and optimisation of a human vertical jump. J. Biomech. 1999, 32, 521–530. [Google Scholar] [CrossRef]
- Niiyama, R.; Nagakubo, A.; Kuniyoshi, Y. Mowgli: A bipedal jumping and landing robot with an artificial musculoskeletal system. In Proceedings of the 2007 IEEE International Conference on Robotics and Automation, Roma, Italy, 10–14 April 2007; pp. 2546–2551. [Google Scholar] [CrossRef]
- Niiyama, R.; Kuniyoshi, Y. A pneumatic biped with an artificial musculoskeletal system. In Proceedings of the 4th International Symposium on Adaptive Motion of Animals and Machines, Cleveland, OH, USA, 1–6 June 2008; pp. 80–81. [Google Scholar]
- Niiyama, R.; Kuniyoshi, Y. Design principle based on maximum output force profile for a musculoskeletal robot. Ind. Rob. Int. J. 2010, 37, 250–255. [Google Scholar] [CrossRef] [Green Version]
- Boston Dynamics’ Humanoid Robot Can Now Jump Upstairs Like a Parkour Pro. Available online: https://www.businessinsider.com/boston-dynamics-atlas-robot-do-parkour-2018-10?IR=T (accessed on 20 August 2020).
- Parkour Atlas. Available online: https://www.youtube.com/watch?v=LikxFZZO2sk (accessed on 20 August 2020).
- Hyon, S.H.; Mita, T. Development of a biologically inspired hopping robot—“Kenken”. In Proceedings of the 2002 IEEE International Conference on Robotics and Automation, Washington, DC, USA, 11–15 May 2002; Volume 4, pp. 3984–3991. [Google Scholar] [CrossRef]
- Hyon, S.H.; Emura, T.; Mita, T. Dynamics-based control of a one-legged hopping robot. Proc. Inst. Mech. Eng. Part I J. Syst. Control Eng. 2003, 217, 83–98. [Google Scholar] [CrossRef]
- Koh, J.S.; Yang, E.; Jung, G.P.; Jung, S.P.; Son, J.H.; Lee, S.I.; Jablonski, P.G.; Wood, R.J.; Kim, H.Y.; Cho, K.J. Jumping on water: Surface tension–dominated jumping of water striders and robotic insects. Science 2015, 349, 517–521. [Google Scholar] [CrossRef]
- Burrows, M.; Sutton, G.P. Pygmy mole crickets jump from water. Curr. Biol. 2012, 22, 990–991. [Google Scholar] [CrossRef] [Green Version]
- Burrows, M. Jumping from the surface of water by the long-legged fly Hydrophorus (Diptera, Dolichopodidae). J. Exp. Biol. 2013, 216, 1973–1981. [Google Scholar] [CrossRef] [Green Version]
- Shih, A.M.; Mendelson, L.; Techet, A.H. Archer fish jumping prey capture: Kinematics and hydrodynamics. J. Exp. Biol. 2017, 220, 1411–1422. [Google Scholar] [CrossRef] [Green Version]
- Martinez, M.M.; Full, R.J.; Koehl, M.A. Underwater punting by an intertidal crab: A novel gait revealed by the kinematics of pedestrian locomotion in air versus water. J. Exp. Biol. 1998, 201, 2609–2623. [Google Scholar]
- Nauwelaerts, S.; Scholliers, J.; Aerts, P. A functional analysis of how frogs jump out of water. Biol. J. Linn. Soc. 2004, 83, 413–420. [Google Scholar] [CrossRef]
- Bush, J.W.; Hu, D.L. Walking on Water: Biolocomotion at the Interface. Annu. Rev. Fluid Mech. 2006, 38, 339–369. [Google Scholar] [CrossRef]
- Glasheen, J.W.; McMahon, T.A. A hydrodynamic model of locomotion in the basilisk lizard. Nature 1996, 380, 340. [Google Scholar] [CrossRef]
- Hsieh, S.T.; Lauder, G.V. Running on water: Three-dimensional force generation by basilisk lizards. Proc. Natl. Acad. Sci. USA 2004, 101, 16784–16788. [Google Scholar] [CrossRef] [Green Version]
- Burrows, M.; Picker, M.D. Jumping mechanisms and performance of pygmy mole crickets (Orthoptera, Tridactylidae). J. Exp. Biol. 2010, 213, 2386–2398. [Google Scholar] [CrossRef] [Green Version]
- Dewenter, J.; Gerullis, P.; Hecker, A.; Schuster, S. Archerfish use their shooting technique to produce adaptive underwater jets. J. Exp. Biol. 2017, 220, 1019–1025. [Google Scholar] [CrossRef] [Green Version]
- Shih, A.M.; Techet, A.H. Characterization of the jumping behavior of archer fish, Toxotes Microlepis. In Proceedings of the 6th World Congress of Biomechanics (WCB 2010), Singapore, 1–6 August 2010. [Google Scholar] [CrossRef]
- Feng, X.Q.; Gao, X.; Wu, Z.; Jiang, L.; Zheng, Q.S. Superior water repellency of water strider legs with hierarchical structures: Experiments and analysis. Langmuir 2007, 23, 4892–4896. [Google Scholar] [CrossRef]
- Shi, F.; Niu, J.; Liu, J.; Liu, F.; Wang, Z.; Feng, X.Q.; Zhang, X. Towards understanding why a superhydrophobic coating is needed by water striders. Adv. Mater. 2007, 19, 2257–2261. [Google Scholar] [CrossRef]
- Song, Y.S.; Sitti, M. Surface-tension-driven biologically inspired water strider robots: Theory and experiments. IEEE Trans. Robot. 2007, 23, 578–589. [Google Scholar] [CrossRef]
- Shin, B.; Kim, H.Y.; Cho, K.J. Towards a biologically inspired small-scale water jumping robot. In Proceedings of the 2008 IEEE RAS & EMBS International Conference on Biomedical Robotics and Biomechatronics, Scottsdale, AZ, USA, 19–22 October 2008; pp. 127–131. [Google Scholar] [CrossRef]
- Zhang, X.; Zhao, J.; Zhu, Q.; Chen, N.; Zhang, M.; Pan, Q. Bioinspired aquatic microrobot capable of walking on water surface like a water strider. ACS Appl. Mater. Interfaces 2011, 3, 2630–2636. [Google Scholar] [CrossRef]
- Zhao, J.; Zhang, X.; Chen, N.; Pan, Q. Why superhydrophobicity is crucial for a water-jumping microrobot? Experimental and theoretical investigations. ACS Appl. Mater. Interfaces 2012, 4, 3706–3711. [Google Scholar] [CrossRef]
- Yan, J.; Yang, K.; Wang, T.; Zhang, X.; Zhao, J. A continuous jumping robot on water mimicking water striders. In Proceedings of the 2016 IEEE International Conference on Robotics and Automation, Stockholm, Sweden, 16–21 May 2016; pp. 4686–4691. [Google Scholar] [CrossRef]
- Jiang, F.; Zhao, J.; Kota, A.K.; Xi, N.; Mutka, M.W.; Xiao, L. A miniature water surface jumping robot. IEEE Robot. Autom. Lett. 2017, 2, 1272–1279. [Google Scholar] [CrossRef]
- Yu, J.; Su, Z.; Wu, Z.; Tan, M. Development of a fast-swimming dolphin robot capable of leaping. IEEE/ASME Trans. Mechatron. 2016, 21, 2307–2316. [Google Scholar] [CrossRef]
- How to Get a Robot to Jump Out of Water. Available online: https://www.electronicsweekly.com/news/research-news/get-robot-jump-water-2018-11/ (accessed on 20 August 2020).
- Yu, J.; Su, Z.; Wu, Z.; Tan, M. An integrative control method for bio-inspired dolphin leaping: Design and experiments. IEEE Trans. Ind. Electron. 2016, 63, 3108–3116. [Google Scholar] [CrossRef]
- Zufferey, R.; Ancel, A.O.; Farinha, A.; Siddall, R.; Kovac, M. Consecutive aquatic jump-gliding with water-reactive fuel. Sci. Robot. 2019, 4, eaax7330. [Google Scholar] [CrossRef]
- Siddall, R.; Kovac, M. Fast aquatic escape with a jet thruster. IEEE-ASME Trans. Mech. 2017, 22, 217–226. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.; Wang, H.; Helbling, E.F.; Jafferis, N.T.; Zufferey, R.; Ong, A.; Ma, K.; Gravish, N.; Chirarattananon, P.; Kovac, M.; et al. A biologically inspired, flapping-wing, hybrid aerial-aquatic microrobot. Sci. Robot. 2017, 2, eaao5619. [Google Scholar] [CrossRef] [Green Version]
- Siddall, R.; Kovač, M. Launching the AquaMAV: Bioinspired design for aerial-aquatic robotic platforms. Bioinspir. Biomim. 2014, 9, 031001. [Google Scholar] [CrossRef]
- Calisti, M.; Falotico, E.; Laschi, C. Hopping on uneven terrains with an underwater one-legged robot. IEEE Robot. Autom. Lett. 2016, 1, 461–468. [Google Scholar] [CrossRef]
- Calisti, M.; Laschi, C. Morphological and control criteria for self-stable underwater hopping. Bioinspir. Biomim. 2017, 13, 016001. [Google Scholar] [CrossRef] [Green Version]
- Romano, D.; Benelli, G.; Stefanini, C. Escape and surveillance asymmetries in locusts exposed to a Guinea fowl-mimicking robot predator. Sci. Rep. 2017, 7, 12825. [Google Scholar] [CrossRef] [Green Version]
- Romano, D.; Benelli, G.; Stefanini, C. Encoding lateralization of jump kinematics and eye use in a locust via bio-robotic artifacts. J. Exp. Biol. 2019, 222, jeb187427. [Google Scholar] [CrossRef] [Green Version]
- Sutton, G.P.; Burrows, M. The mechanics of elevation control in locust jumping. J. Comp. Physiol. A 2008, 194, 557–563. [Google Scholar] [CrossRef]
- Chai, H.; Li, J.; Ge, W. Gait Analysis on Bionic Kangaroo-hopping Robot Based on Adjustable Geared Five-linkage Mechanism. Robot 2009, 31. [Google Scholar] [CrossRef]
- Santer, R.D.; Yamawaki, Y.; Rind, F.C.; Simmons, P.J. Motor activity and trajectory control during escape jumping in the locust Locusta migratoria. J. Comp. Physiol. A 2005, 191, 965–975. [Google Scholar] [CrossRef]
- Simmons, P.J.; Rind, F.C.; Santer, R.D. Escapes with and without preparation: The neuroethology of visual startle in locusts. J. Insect Physiol. 2010, 56, 876–883. [Google Scholar] [CrossRef]
- Zhao, J.; Xu, J.; Gao, B.; Xi, N.; Cintron, F.J.; Mutka, M.W.; Xiao, L. MSU jumper: A single-motor-actuated miniature steerable jumping robot. IEEE Trans. Robot 2013, 29, 602–614. [Google Scholar] [CrossRef]
- Zhao, J.; Zhao, T.; Xi, N.; Mutka, M.W.; Xiao, L. MSU tailbot: Controlling aerial maneuver of a miniature-tailed jumping robot. IEEE/ASME Trans. Mechatron. 2015, 20, 2903–2914. [Google Scholar] [CrossRef]
- Bennet-Clark, H.C. The energetics of the jump of the locust Schistocerca gregaria. J. Exp. Biol. 1975, 63, 53–83. [Google Scholar] [CrossRef]
- Burrows, M.; Sutton, G.P. Locusts use a composite of resilin and hard cuticle as an energy store for jumping and kicking. J. Exp. Biol. 2012, 215, 3501–3512. [Google Scholar] [CrossRef] [Green Version]
- Frantsevich, L. Righting kinematics in beetles (Insecta: Coleoptera). Arthropod Struct. Dev. 2004, 33, 221–235. [Google Scholar] [CrossRef]
- Warrick, D.R.; Dial, K.P.; Biewener, A.A. Asymmetrical force production in the maneuvering flight of pigeons. Auk 1998, 115, 916–928. [Google Scholar] [CrossRef] [Green Version]
- Warrick, D.; Dial, K.P. Kinematic, aerodynamic and anatomical mechanisms in the slow, maneuvering flight of pigeons. J. Exp. Biol. 1998, 201, 655–672. [Google Scholar]
- Warrick, D.R.; Bundle, M.W.; Dial, K.P. Bird maneuvering flight: Blurred bodies, clear heads. Integr. Comp. Biol. 2002, 42, 141–148. [Google Scholar] [CrossRef]
- Robertson, R.M.; Johnson, A.G. Collision avoidance of flying locusts: Steering torques and behaviour. J. Exp. Biol. 1993, 183, 35–60. [Google Scholar]
- Karásek, M.; Muijres, F.T.; De Wagter, C.; Remes, B.D.; de Croon, G.C. A tailless aerial robotic flapper reveals that flies use torque coupling in rapid banked turns. Science 2018, 361, 1089–1094. [Google Scholar] [CrossRef] [Green Version]
- Luu, T.; Cheung, A.; Ball, D.; Srinivasan, M.V. Honeybee flight: A novel ‘streamlining’ response. J. Exp. Biol. 2011, 214, 2215–2225. [Google Scholar] [CrossRef] [Green Version]
- Zanker, J.M. On the mechanism of speed and altitude control in Drosophila melanogaster. Physiol. Entomol. 1988, 13, 351–361. [Google Scholar] [CrossRef]
- Weis-Fogh, T. Quick estimates of flight fitness in hovering animals, including novel mechanisms for lift production. J. Exp. Biol. 1973, 59, 169–230. [Google Scholar]
- Thomas, A.L. Why do birds have tails? The tail as a drag reducing flap, and trim control. J. Theor. Biol. 1996, 183, 247–253. [Google Scholar] [CrossRef]
- Thomas, A.L. On the aerodynamics of birds’ tails. Philos. Trans. R. Soc. Lond. B Biol. Sci. 1993, 340, 361–380. [Google Scholar] [CrossRef]
- Dyhr, J.P.; Cowan, N.J.; Colmenares, D.J.; Morgansen, K.A.; Daniel, T.L. Autostabilizing airframe articulation: Animal inspired air vehicle control. In Proceedings of the 2012 IEEE Conference on Decision and Control, Maui, HI, USA, 10–13 December 2012; pp. 3715–3720. [Google Scholar] [CrossRef] [Green Version]
- Baader, A. The posture of the abdomen during locust flight: Regulation by steering and ventilatory interneurones. J. Exp. Biol. 1990, 151, 109–131. [Google Scholar]
- Santer, R.D.; Simmons, P.J.; Rind, F.C. Gliding behaviour elicited by lateral looming stimuli in flying locusts. J. Comp. Physiol. A 2005, 191, 61–73. [Google Scholar] [CrossRef]
- Tucker, V.A. Gliding birds: The effect of variable wing span. J. Exp. Biol. 1987, 133, 33–58. [Google Scholar] [CrossRef] [Green Version]
- McGuire, J.A.; Dudley, R. The biology of gliding in flying lizards (genus Draco) and their fossil and extant analogs. Integr. Comp. Biol. 2011, 51, 983–990. [Google Scholar] [CrossRef] [Green Version]
- Flaherty, E.A.; Ben-David, M.; Smith, W.P. Quadrupedal locomotor performance in two species of arboreal squirrels: Predicting energy savings of gliding. J. Comp. Physiol. B 2010, 180, 1067–1078. [Google Scholar] [CrossRef]
- Byrnes, G.; Lim, N.T.L.; Spence, A.J. Take-off and landing kinetics of a free-ranging gliding mammal, the Malayan colugo (Galeopterus variegatus). Philos Trans. R Soc. Lond. B Biol. Sci. 2008, 275, 1007–1013. [Google Scholar] [CrossRef] [Green Version]
- Socha, J.J. Gliding Flight in Chrysopelea: Turning a Snake into a Wing. Integr. Comp. Biol. 2011, 51, 969–982. [Google Scholar] [CrossRef] [Green Version]
- Socha, J.J. Kinematics: Gliding flight in the paradise tree snake. Nature 2002, 418, 603. [Google Scholar] [CrossRef]
- Socha, J.J.; Jafari, F.; Munk, Y.; Byrnes, G. How animals glide: From trajectory to morphology. Can. J. Zool. 2015, 93, 901–924. [Google Scholar] [CrossRef]
- Bishop, K.L. Aerodynamic force generation, performance and control of body orientation during gliding in sugar gliders (Petaurus breviceps). J. Exp. Biol. 2007, 210, 2593–2606. [Google Scholar] [CrossRef] [Green Version]
- Paskins, K.E.; Bowyer, A.; Megill, W.M.; Scheibe, J.S. Take-off and landing forces and the evolution of controlled gliding in northern flying squirrels Glaucomys sabrinus. J. Exp. Biol. 2007, 210, 1413–1423. [Google Scholar] [CrossRef] [Green Version]
- Bahlman, J.W.; Swartz, S.M.; Riskin, D.K.; Breuer, K.S. Glide performance and aerodynamics of non-equilibrium glides in northern flying squirrels (Glaucomys sabrinus). J. R. Soc. Interface 2013, 10, 20120794. [Google Scholar] [CrossRef] [Green Version]
- Socha, J.J.; Miklasz, K.; Jafari, F.; Vlachos, P.P. Non-equilibrium trajectory dynamics and the kinematics of gliding in a flying snake. Bioinspir. Biomim. 2010, 5, 045002. [Google Scholar] [CrossRef] [Green Version]
- Yanoviak, S.P.; Munk, Y.; Kaspari, M.; Dudley, R. Aerial manoeuvrability in wingless gliding ants (Cephalotes atratus). Philos. Trans. R. Soc. Lond. B Biol. Sci. 2010, 277, 2199–2204. [Google Scholar] [CrossRef] [Green Version]
- Munk, Y.; Yanoviak, S.P.; Koehl, M.A.R.; Dudley, R. The descent of ant: Field-measured performance of gliding ants. J. Exp. Biol. 2015, 218, 1393–1401. [Google Scholar] [CrossRef] [Green Version]
- Zeng, Y.; Lam, K.; Chen, Y.; Gong, M.; Xu, Z.; Dudley, R. Biomechanics of aerial righting in wingless nymphal stick insects. Interface Focus 2017, 7, 20160075. [Google Scholar] [CrossRef]
- Sechzer, J.A.; Folstein, S.E.; Geiger, E.H.; Mervis, R.F.; Meehan, S.M. Development and maturation of postural reflexes in normal kittens. Exp. Neurol. 1984, 86, 493–505. [Google Scholar] [CrossRef]
- Diamond, J.M. Why cats have nine lives. Nature 1988, 332, 586–587. [Google Scholar] [CrossRef]
- Kane, T.R.; Scher, M.P. A dynamical explanation of the falling cat phenomenon. Int. J. Solids Struct. 1969, 5, 663–670. [Google Scholar] [CrossRef]
- Arabyan, A.; Tsai, D. A distributed control model for the air-righting reflex of a cat. Biol. Cybern. 1998, 79, 393–401. [Google Scholar] [CrossRef]
- Galli, J.R. Angular momentum conservation and the cat twist. Phys. Teach. 1995, 33, 404–407. [Google Scholar] [CrossRef]
- Kawano, F.; Yoneshima, H.; Ishihara, A.; Igarashi, M.; Ohira, Y. Hindlimb suspension inhibits air-righting due to altered recruitment of neck and back muscles in rats. Jpn. J. Physiol. 2004, 54, 229–242. [Google Scholar] [CrossRef] [Green Version]
- Masuda, K.; Yamaguchi, T. Abnormal air-righting reflex in striatal rats. Jpn. J. Physiol. 2000, 50, 163–166. [Google Scholar] [CrossRef]
- Schönfelder, J. The development of air-righting reflex in postantal growing rabbits. Behav. Brain Res. 1984, 11, 213–221. [Google Scholar] [CrossRef]
- Burrows, M.; Cullen, D.A.; Dorosenko, M.; Sutton, G.P. Mantises exchange angular momentum between three rotating body parts to jump precisely to targets. Curr. Biol. 2015, 25, 786–789. [Google Scholar] [CrossRef] [Green Version]
- Jusufi, A.; Goldman, D.I.; Revzen, S.; Full, R.J. Active tails enhance arboreal acrobatics in geckos. Proc. Natl. Acad. Sci. USA 2008, 105, 4215–4219. [Google Scholar] [CrossRef] [Green Version]
- Chang-Siu, E.; Libby, T.; Tomizuka, M.; Full, R.J. A lizard-inspired active tail enables rapid maneuvers and dynamic stabilization in a terrestrial robot. In Proceedings of the 2011 IEEE/RSJ International Conference on Intelligent Robots and Systems, San Francisco, CA, USA, 25–30 September 2011; pp. 1887–1894. [Google Scholar] [CrossRef]
- Libby, T.; Moore, T.Y.; Changsiu, E.; Li, D.; Cohen, D.; Jusufi, A.; Full, R.J. Tail-assisted pitch control in lizards, robots and dinosaurs. Nature 2012, 481, 181–184. [Google Scholar] [CrossRef]
- Gillis, G.B.; Bonvini, L.A.; Irschick, D.J. Losing stability: Tail loss and jumping in the arboreal lizard Anolis carolinensis. J. Exp. Biol. 2009, 212, 604–609. [Google Scholar] [CrossRef] [Green Version]
- Higham, T.E.; Davenport, M.S.; Jayne, B.C. Maneuvering in an arboreal habitat: The effects of turning angle on the locomotion of three sympatric ecomorphs of Anolis lizards. J. Exp. Biol. 2001, 204, 4141–4155. [Google Scholar] [CrossRef] [Green Version]
- Hickman, G.C. The mammalian tail: A review of functions. Mammal Rev. 1979, 9, 143–157. [Google Scholar] [CrossRef]
- O’Connor, S.M.; Dawson, T.J.; Kram, R.; Donelan, J.M. The kangaroo’s tail propels and powers pentapedal locomotion. Biol. Lett. 2014, 10, 559–578. [Google Scholar] [CrossRef] [Green Version]
- Kawata, K. A note on the function of the tail in the Macropodinae. Int. Zoo Yearb. 1971, 11, 23. [Google Scholar] [CrossRef]
- Moore, J.; Gutmann, A.; Craig, M.; McKinley, P. Exploring the role of the tail in bipedal hopping through computational evolution. In Artificial Life Conference Proceedings, 13 September 2013; MIT Press: Cambridge, MA, USA; pp. 11–18. [CrossRef]
- Patel, A.; Boje, E. On the conical motion of a two-degree-of-freedom tail inspired by the cheetah. IEEE Trans. Robot. 2015, 31, 1555–1560. [Google Scholar] [CrossRef]
- Patel, A.; Boje, E.; Fisher, C.; Louis, L.; Lane, E. Quasi-steady state aerodynamics of the cheetah tail. Biol. Open 2016, 5, 1072–1076. [Google Scholar] [CrossRef] [Green Version]
- Patel, A.; Braae, M. Rapid acceleration and braking: Inspirations from the cheetah’s tail. In Proceedings of the 2014 IEEE International Conference on Robotics and Automation, Hong Kong, China, 31 May–7 June 2014; pp. 793–799. [Google Scholar] [CrossRef]
- Patel, A.; Braae, M. Rapid turning at high-speed: Inspirations from the cheetah’s tail. In Proceedings of the 2013 IEEE/RSJ International Conference on Intelligent Robots and Systems, Hong Kong, China, 31 May–7 June 2014; pp. 5506–5511. [Google Scholar] [CrossRef]
- Briggs, R.; Lee, J.; Haberland, M.; Kim, S. Tails in biomimetic design: Analysis, simulation, and experiment. In Proceedings of the 2012 IEEE/RSJ International Conference on Intelligent Robots and Systems, Vilamoura, Portugal, 7–12 October 2012; pp. 1473–1480. [Google Scholar] [CrossRef]
- Jusufi, A.; Kawano, D.T.; Libby, T.; Full, R.J. Righting and turning in mid-air using appendage inertia: Reptile tails, analytical models and bio-inspired robots. Bioinspir. Biomim. 2010, 5, 045001. [Google Scholar] [CrossRef] [Green Version]
- Walker, C.; Vierck, C.J., Jr.; Ritz, L.A. Balance in the cat: Role of the tail and effects of sacrocaudal transection. Behav. Brain Res. 1998, 91, 41–47. [Google Scholar] [CrossRef]
- Dyhr, J.P.; Morgansen, K.A.; Daniel, T.L.; Cowan, N.J. Flexible strategies for flight control: An active role for the abdomen. J. Exp. Biol. 2013, 216, 1523–1536. [Google Scholar] [CrossRef] [Green Version]
- Jusufi, A.; Zeng, Y.; Full, R.J.; Dudley, R. Aerial Righting Reflexes in Flightless Animals. Integr. Comp. Biol. 2011, 51, 937–943. [Google Scholar] [CrossRef] [Green Version]
- Zeglin, G.J. Uniroo—A One Legged Dynamic Hopping Robot. Bachelor’s Thesis, Massachusetts Institute of Technology, Cambridge, MA, USA, 1991. [Google Scholar]
- Chang-Siu, E.; Libby, T.; Brown, M.; Full, R.J.; Tomizuka, M. A nonlinear feedback controller for aerial self-righting by a tailed robot. In Proceedings of the 2013 IEEE International Conference on Robotics and Automation, Karlsruhe, Germany, 6–10 May 2013; pp. 32–39. [Google Scholar] [CrossRef]
- De, A.; Koditschek, D.E. The Penn Jerboa: A platform for exploring parallel composition of templates. arXiv 2015, arXiv:1502.05347. [Google Scholar]
- Saab, W.; Rone, W.S.; Ben-Tzvi, P. Robotic tails: A state-of-the-art review. Robotica 2018, 36, 1263–1277. [Google Scholar] [CrossRef] [Green Version]
- Truong, T.N.; Phan, H.V.; Park, H.C. Design and demonstration of a bio-inspired flapping-wing-assisted jumping robot. Bioinspir. Biomim. 2019, 14, 036010. [Google Scholar] [CrossRef]
- Slow Motion Video of Vertical Jump with Synchronized Vertical Force Data. Available online: https://www.youtube.com/watch?v=qN3apht8zRs (accessed on 20 August 2020).
- Nauwelaerts, S.; Aerts, P. Take-off and landing forces in jumping frogs. J. Exp. Biol. 2006, 209, 66–77. [Google Scholar] [CrossRef] [Green Version]
- Alexander, R.M. Elastic Mechanisms in Animal Movement; Cambridge University Press: Cambridge, UK, 1988. [Google Scholar]
- Domokos, G.; Várkonyi, P.L. Geometry and self-righting of turtles. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2007, 275, 11–17. [Google Scholar] [CrossRef] [Green Version]
- Chen, D.; Zheng, W.; Huang, Y.; Shen, Q. The design and optimization of a hopping robot’s tipping mechanism. Chin. J. Mech. Eng. 2011, 47, 17–23. [Google Scholar] [CrossRef]
- Zhao, J.; Yang, R.; Xi, N.; Gao, B.; Fan, X.; Mutka, M.W.; Xiao, L. Development of a miniature self-stabilization jumping robot. In Proceedings of the 2009 IEEE/RSJ International Conference on Intelligent Robots and Systems, St. Louis, MO, USA, 10–15 October 2009; pp. 2217–2222. [Google Scholar] [CrossRef]
- Ho, T.; Lee, S. A novel design of a robot that can jump and roll with a single actuator. In Proceedings of the 2012 IEEE/RSJ International Conference on Intelligent Robots and Systems, Vilamoura, Portugal, 7–12 October 2012; pp. 908–913. [Google Scholar] [CrossRef]
- Armour, R.H. A Biologically Inspired Jumping and Rolling Robot. Ph.D. Thesis, University of Bath, Bath, UK, 2010. [Google Scholar]
- Kovac, M.; Savioz, G.; Zufferey, J.C.; Floreano, D.; Srinivasan, M.; Ellington, C. Towards the Self Deploying Microglider, Gliding Flight and Bioinspired Wing Folding Mechanism. In Proceedings of the International Symposium on Flying Insects and Robots, Ascona, Switzerland, 12–17 August 2007; pp. 61–62. [Google Scholar]
- Kovac, M.; Guignard, A.; Nicoud, J.D.; Zufferey, J.C.; Floreano, D. A 1.5 g SMA-actuated microglider looking for the light. In Proceedings of the 2007 IEEE International Conference on Robotics and Automation, Roma, Italy, 10–14 April 2007; pp. 367–372. [Google Scholar] [CrossRef] [Green Version]
- Kovač, M.; Zufferey, J.C.; Floreano, D. Towards a self-deploying and gliding robot. In Flying Insects and Robots; Springer: Berlin/Heidelberg, Germany, 2009; pp. 271–284. [Google Scholar] [CrossRef] [Green Version]
- Desbiens, A.L.; Pope, M.; Berg, F.; Teoh, Z.E.; Lee, J.; Cutkosky, M. Efficient jumpgliding: Theory and design considerations. In Proceedings of the 2013 IEEE International Conference on Robotics and Automation, Karlsruhe, Germany, 6–10 May 2013; pp. 4451–4458. [Google Scholar] [CrossRef]
- Zhang, Z.; Chen, D.; Chen, K. Analysis and comparison of three leg models for bionic locust robot based on landing buffering performance. Sci. China Technol. Sci. 2016, 59, 1413–1427. [Google Scholar] [CrossRef]
- Dupeyroux, J.; Serres, J.R.; Viollet, S. AntBot: A six-legged walking robot able to home like desert ants in outdoor environments. Sci. Robot. 2019, 4, eaau0307. [Google Scholar] [CrossRef] [Green Version]
- Chen, X.; Ghadirzadeh, A.; Folkesson, J.; Björkman, M.; Jensfelt, P. Deep reinforcement learning to acquire navigation skills for wheel-legged robots in complex environments. In Proceedings of the 2018 IEEE/RSJ International Conference on Intelligent Robots and Systems, Madrid, Spain, 1–5 October 2018; pp. 3110–3116. [Google Scholar] [CrossRef] [Green Version]
- Pfeiffer, M.; Shukla, S.; Turchetta, M.; Cadena, C.; Krause, A.; Siegwart, R.; Nieto, J. Reinforced imitation: Sample efficient deep reinforcement learning for mapless navigation by leveraging prior demonstrations. IEEE Robot. Autom. Mag. 2018, 3, 4423–4430. [Google Scholar] [CrossRef] [Green Version]
- Hwu, T.; Wang, A.Y.; Oros, N.; Krichmar, J.L. Adaptive robot path planning using a spiking neuron algorithm with axonal delays. IEEE Trans. Cogn. Dev. Syst. 2017, 10, 126–137. [Google Scholar] [CrossRef]
- Antonelo, E.A.; Schrauwen, B. On learning navigation behaviors for small mobile robots with reservoir computing architectures. IEEE Trans. Neur. Netw. Learn. Syst. 2014, 26, 763–780. [Google Scholar] [CrossRef]
- Wu, Q.; Lin, C.M.; Fang, W.; Chao, F.; Yang, L.; Shang, C.; Zhou, C. Self-organizing brain emotional learning controller network for intelligent control system of mobile robots. IEEE Access 2018, 6, 59096–59108. [Google Scholar] [CrossRef]
- Nichols, E.; McDaid, L.J.; Siddique, N. Biologically inspired SNN for robot control. IEEE Trans. Cybern. 2013, 43, 115–128. [Google Scholar] [CrossRef]




| Strategy | Animals | Body Length (mm) | Body Mass (mg) | Take-off Time (ms) t | Take-off Angle (°) | Take-off Velocity (m/s) v | Acceleration (m/s2) | a/g g = 9.81 | Energy (uJ) | Power (mW) p | Power/Muscle Mass (w/kg) | Ratio of Leg Length 16 | Leg Ratio 17 |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Catapult mechanism | Locust 1 [48,51,52,53] | 70 | 25–30 | 45 | 3.2 | 107 | 11 | 103 | 360–450 | 51,420–64,285 | * | * | |
| Flea 2 [54,55] | 1.8 ± 0.19 | 0.7 ± 0.16 | 1.4 ± 0.25 | 39 ± 5.7 | 1.3 ± 0.21 | 960 ± 233 | 98 ± 23.7 | 0.6 ± 0.28 | 0.43 ± 0.19 | 6000 ± 2100 | 1:1.3:1.9 | 154 | |
| Cockroach 3 [56,57] | 6.3–10 | 31 | 10.6 ± 1.0 | 40 | 1.5 ± 0.3 | 220 | 23 | 38 | 3.4 | 1100 | 1:1.2:2.4 | 150 | |
| Cricket 4 [58] | 700 ± 100 | 1.1 | * | * | * | * | 820 | 30 ± 20 | 3800 ± 2200 | * | * | ||
| Froghopper 5 [59,60] | 6.1 ± 0.08 | 12.3 ± 0.74 | 0.875 | 46.8 ± 2.0 | 4.7 | 5400 | 550 | 136 | 155 | 126,000 | 1:1:1.5 | 66 | |
| Leafhopper 6 [61,62,63] | 9.2 ± 0.33 | 19 ± 1.10 | 6.4 ± 0.21 | 34.3 ± 5.90 | 1.2 ± 0.13 | 188 | 19 | 14 | 2 | 1053 | 1:1.1:1.9 | 82 | |
| Shore bug 7 [64] | 3.5 ± 0.09 | 2.1 ± 0.09 | 3.9 ± 0.05 | 34 ± 0.9 | 1.3 ± 0.03 | 335 ± 9.4 | 34 ± 0.9 | 1.8 ± 0.1 | 0.5 ± 0.02 | 4500 | 1:1.2:1.8 | 89.6 | |
| Direct muscle actuation | Moth 8 [47] | 9.1 ± 0.6 | 5.4 ± 0.9 | 11.8 ± 1.1 | 44.4 ± 5.1 | 0.8 ± 0.1 | 71 | 7 | 1.9 | 0.2 | 299 | 1:1.4:2.1 | 132 |
| Caddisfly 9 [46] | 11.4 ± 0.3 | 38.1 ± 2.4 | 15.4 ± 0.8 | * | 0.9 ± 0.04 | 57 | 6 | 16.4 | 1 | 256 | 1:1.5:1.7 | 105 | |
| Mantis 10 [42] | 70.7 ± 4.0 | 939.2 ± 84.8 | 65.9 ± 2.7 | 23.1 ± 2.6 | 1.12 ± 0.03 | 17 | 1.73 | 598.07 | 8.94 | 63.44 | 1:0.7:1.0 | 66 | |
| Bush cricket 11 [65] | 23.2 ± 0.8 | 602 ± 42 | 32.6 ± 0.95 | 34 | 2.12 ± 0.33 | 65.03 | 6.63 | 1380 | 40 (67) | 664 (1113) | 1:1.2:4.6 | 158 | |
| Hydrostatic pressure | Jump spider 12 [66,67,68] | 6.1 ± 0.3 | 19.95 ± 4.71 | * | 11.4 ± 5.2 | 0.99 ± 0.13 | * | * | 10 ± 4 | * | * | * | * |
| Jump without legs | Click beetle 13 [69,70,71] | 10–12 | 40 | 0.64 | 85 | 2.4 | 3800 | 380 | 115 | 185,000 | 130 | * | * |
| Springtail 14 [45,72] | 2 | 2.1 | 6.0 | * | 1.4 | 981 | 98 | 0.001 | * | * | * | * | |
| Larva 15 [49,50] | 3 | * | * | * | 1 | 18,000 | * | * | * | * | * | * |
| Aspects | Strategies Used in Animals | Strategies Used in Robots |
|---|---|---|
| Take-off angle | Orientation of hindlegs. | (1) Changing some components (bar length, gear ratio) manually; (2) Adjusting the position of the counterpart; (3) Adjusting the body pitch angle using support bar. |
| Take-off direction | (1) Orientation of two fore legs; (2) Orientation of two hind legs. | (1) Rotating the whole robot around axis; (2) Discontinuous rotation the body around fixed tail; (3) Differential wheels. |
| Take-off velocity | Stored energy inside muscle and elastic components. | (1) More than one end points of energy storage component; (2) Changing the gas ratio and flow rate of flammable gas; (3) The deformation and initial shape of SMA; (4) External magnetic field and direction. |
| Take-off stability | (1) Ensuring that contact force passes through the centroid and counter of the stomach’s contraction; (2) Balancing tail. | (1) Balancing tail; (2) Zero-momentum by ensuring that contact force passes through the centroid during take-off phase. |
| Aerial righting | (1) Aerial righting by way of aerodynamic torque; (2) Aerial righting by way of inertia; (3) Using dragline silk. | (1) One/two-DOF balanced tail; (2) Gliding wings; (3) Propellers. |
| Landing buffer and resetting mechanism | (1) Compliant body structure helps absorb shock; (2) Landing softly with help of tendons in legs; (3) Resetting with help of legs. | (1) Active resetting using support legs; (2) Passive resetting assisted by flexible frame; (3) Gliding wings help landing; (4) Elastic buffering legs help absorb impact. |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mo, X.; Ge, W.; Miraglia, M.; Inglese, F.; Zhao, D.; Stefanini, C.; Romano, D. Jumping Locomotion Strategies: From Animals to Bioinspired Robots. Appl. Sci. 2020, 10, 8607. https://doi.org/10.3390/app10238607
Mo X, Ge W, Miraglia M, Inglese F, Zhao D, Stefanini C, Romano D. Jumping Locomotion Strategies: From Animals to Bioinspired Robots. Applied Sciences. 2020; 10(23):8607. https://doi.org/10.3390/app10238607
Chicago/Turabian StyleMo, Xiaojuan, Wenjie Ge, Marco Miraglia, Francesco Inglese, Donglai Zhao, Cesare Stefanini, and Donato Romano. 2020. "Jumping Locomotion Strategies: From Animals to Bioinspired Robots" Applied Sciences 10, no. 23: 8607. https://doi.org/10.3390/app10238607
APA StyleMo, X., Ge, W., Miraglia, M., Inglese, F., Zhao, D., Stefanini, C., & Romano, D. (2020). Jumping Locomotion Strategies: From Animals to Bioinspired Robots. Applied Sciences, 10(23), 8607. https://doi.org/10.3390/app10238607