The Antimicrobial Effect of Radiant Catalytic Ionization on the Bacterial Attachment and Biofilm Formation by Selected Foodborne Pathogens under Refrigeration Conditions

 , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Preparation of Bacterial Suspension
2.3. Assessment of the Biofilm Formation by the Tested Strains
2.4. Contamination of Meat and Glass Coupons
2.5. Experimental Design
- R—bacterial count reduction
- I—initial number of bacteria on glass coupons after contact with the contaminated meat [log CFU × cm−2]
- B—the number of bacteria recovered in a given experimental variant for the control (K+; -RCI) and tested samples (+RCI) [log CFU × cm−2]
- R—bacterial count reduction
- (-RCI)—number of bacteria determined on glass plates in the control variant without the RCI technology [log CFU × cm−2]
- (+RCI)—number of bacteria recovered after RCI technology [log CFU × cm−2]
- E—RCI absolute effectiveness coefficient
- R+RCI—level of bacterial reduction as a result of RCI application
- R-RCI—level of bacterial reduction without RCI application
2.6. Statistical Analysis
3. Results
3.1. Biofilm Formation
3.2. Recovery Ratio of Bacteria from Contaminated Samples
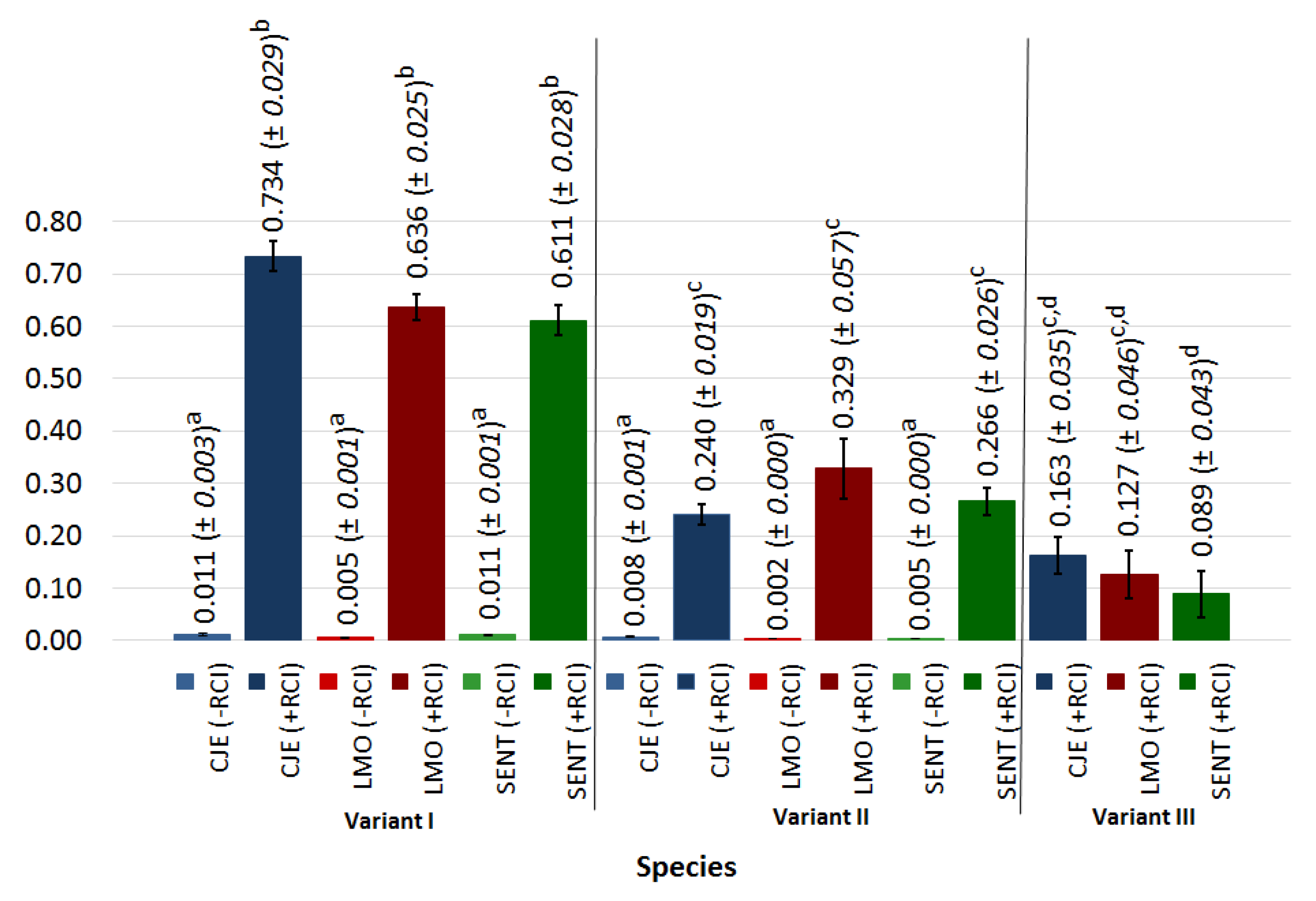
3.3. Assessment of the Effectiveness of Surface Disinfection Using Radiant Catalytic Ionization during Bacterial Attachment (Experimental Variant I)
3.4. Assessment of Effectiveness of Surface Disinfection Using Radiant Catalytic Ionization during Biofilm Maturation (Experimental Variant II)
3.5. Assessment of Effectiveness of Surface Disinfection Using Radiant Catalytic Ionization during Biofilm Formation (Experimental Variant III)
4. Discussion
Author Contributions
Funding
Conflicts of Interest
References
- EFSA. The European Union One Health 2018 Zoonoses Report. Available online: https://www.efsa.europa.eu/en/efsajournal/pub/5926 (accessed on 21 December 2019).
- Desai, A.N.; Anyoha, A.; Madoff, L.C.; Lassmann, B. Changing epidemiology of Listeria monocytogenes outbreaks, sporadic cases, and recalls globally: A review of ProMED reports from 1996 to 2018. Int. J. Infect. Dis. 2019, 84, 48–53. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smith, A.M.; Tau, N.P.; Smouse, S.L.; Allam, M.; Ismail, A.; Ramalwa, N.R.; Disenyeng, B.; Ngomane, M.; Thomas, J. Outbreak of Listeria monocytogenes in South Africa, 2017-2018: Laboratory Activities and Experiences Associated with Whole-Genome Sequencing Analysis of Isolates. Foodborne Pathog. Dis. 2019, 16, 524–530. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Das, A. Listeriosis in Australia—January to July 2018. Glob. Biosecurity 2019, 1, 150–158. [Google Scholar] [CrossRef]
- WHO. Available online: https://www.who.int/csr/don/16-september-2019-listeriosis-spain/en/ (accessed on 21 December 2019).
- Food Safety News. Available online: https://www.foodsafetynews.com/2019/10/german-meat-plant-linked-to-listeria-outbreak-stops-production/ (accessed on 21 December 2019).
- Carpentier, B.; Cerf, O. Review--Persistence of Listeria monocytogenes in food industry equipment and premises. Int. J. Food Microbiol. 2011, 31, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Skowron, K.; Wiktorczyk, N.; Kwiecińska-Piróg, J.; Sękowska, A.; Wałecka-Zacharska, E.; Gospodarek-Komkowska, E. Elimination of Klebsiella pneumoniae NDM from the air and selected surfaces in hospital using radiant catalytic ionization. Lett. Appl. Microbiol. 2019, 69, 333–338. [Google Scholar] [CrossRef] [PubMed]
- Saraiva, C.; García Díez, J.; Fontes, M.; Esteves, A. Modeling the Behavior of Listeria monocytogenes in Meat. List. Monocytogenes 2018, 25. [Google Scholar] [CrossRef] [Green Version]
- Zuber, I.; Lakicevic, B.; Pietzka, A.; Milanov, D.; Djordjevic, V.; Karabasil, N.; Teodorovic, V.; Ruppitsch, W.; Dimitrijevic, M. Molecular characterization of Listeria monocytogenes isolates from a small-scale meat processor in Montenegro, 2011–2014. Food Microbiol. 2019, 79, 116–122. [Google Scholar] [CrossRef]
- Russo, P.; Hadjilouka, A.; Beneduce, L.; Capozzi, V.; Paramithiotis, S.; Drosinos, E.H.; Spano, G. Effect of different conditions on Listeria monocytogenes biofilm formation and removal. Czech. J. Food Sci. 2018, 36, 208–214. [Google Scholar] [CrossRef] [Green Version]
- Manijeh, M.A.; Mohammad, J.M.; Roha, K.K. Biofilm Formation by Salmonella enteritidis on Food Contact Surfaces. J. Biol. Sci. 2008, 8, 502–505. [Google Scholar]
- Hue, O.; Le Bouquin, S.; Laisney, M.J.; Allain, V.; Lalande, F.; Petetin, I.; Rouxel, S.; Quesne, S.; Gloaguen, P.Y.; Picherot, M.; et al. Prevalence of and risk factors for Campylobacter spp. contamination of broiler chicken carcasses at the slaughterhouse. Food Microbiol. 2010, 27, 992–999. [Google Scholar] [CrossRef]
- Connerton, I.F.; Connerton, P.L. Foodborne Diseases, 3rd ed.; Elsevier Science Publishing Co Inc.: San Diego, CA, USA, 2017; pp. 209–221. [Google Scholar]
- Igwaran, A.; Okoh, A.I. Human campylobacteriosis: A public health concern of global importance. Heliyon 2019, 5, e02814. [Google Scholar] [CrossRef] [PubMed]
- Jensen, L.K.; Bjarnsholt, T.; Kragh, K.N.; Aalbæk, B.; Henriksen, N.L.; Blirup, S.A.; Pankoke, K.; Petersen, A.; Jensen, H.E. In vivo gentamicin susceptibility test for prevention of bacterial biofilms in bone tissue and on implants. Antimicrob Agents Chemother. 2019, 63, e01889-18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stewart, P.S. Animicrobial tolorance in biofilms. Microbiol. Spectr. 2015, 3, MB-0010-2014. [Google Scholar] [CrossRef] [Green Version]
- Wu, H.; Moser, C.; Wang, H.Z.; Hoiby, N.; Song, Z.J. Strategies for combating bacterial biofilm infections. Int. J. Oral Sci. 2015, 7, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Pearce, P.; Song, B.; Skinner, D.J.; Mok, R.; Hartmann, R.; Singh, P.K.; Jeckel, H.; Oishi, J.; Drescher, K.; Dunkel, J. Flow-induced symmetry breaking in growing bacterial biofilms. Phys. Rev. Lett. 2019, 20, 258101. [Google Scholar] [CrossRef]
- Flemming, H.C.; Wingender, J.; Szewzyk, U.; Steinberg, P.; Rice, S.A.; Kjelleberg, S. Biofilms: An emergent form of bacterial life. Nat. Rev. Microbiol. 2016, 14, 563–575. [Google Scholar] [CrossRef]
- Giaouris, E.; Heir, E.; Hébraud, M.; Chorianopoulos, N.; Langsrud, S.; Møretrø, T.; Habimana, O.; Desvaux, M.; Renier, S.; Nychas, G.J. Attachment and biofilm formation by foodborne bacteria in meat processing environments: Causes, implications, role of bacterial interactions and control by alternative novel methods. Meat Sci. 2014, 97, 298–309. [Google Scholar] [CrossRef]
- Rupel, K.; Zupin, L.; Ottaviani, G.; Iris Bertani, I.; Martinelli, V.; Porrelli, D.; Vodret, S.; Roman Vuerich, R.; Daniel Passos da Silva, D.; Bussani, R.; et al. Blue laser light inhibits biofilm formation in vitro and in vivo by inducing oxidative stress. NPJ Biofilms Microbiomes 2019, 29, 1–11. [Google Scholar] [CrossRef]
- Diehl, J.F. Food irradiation-Past, present and future. Rad. Phys. Chem. 2002, 63, 211–215. [Google Scholar] [CrossRef]
- Lado, B.H.; Yousef, A.E. Alternative food-preservation technologies: Efficacy and mechanisms. Microbes Infect 2002, 4, 433–440. [Google Scholar] [CrossRef]
- Ortega, M.T.; Franken, L.J.; Hatesohl, P.R.; Marsden, J.L. Efficacy of ecoquest radiant catalytic ionization cell and breeze at ozone generator at reducing microbial populations on stainless steel surfaces. J. Rapid Methods Autom Microbiol. 2007, 15, 359–368. [Google Scholar] [CrossRef]
- Skowron, K.; Grudlewska, K.; Krawczyk, A.; Gospodarek- Komkowska, E. The effectiveness of radiant catalytic ionization in inactivation of Listeria monocytogenes planktonic and biofilm cells from food and food contact surfaces as a method of food preservation. J. Appl. Microbiol. 2018, 124, 1493–1505. [Google Scholar] [CrossRef] [PubMed]
- Skowron, K.; Grudlewska, K.; Kwiecińska-Piróg, J.; Gryń, G.; Śrutek, M.; Gospodarek-Komkowska, E. Efficacy of radiant catalytic ionization to reduce bacterial populations in air and on different surfaces. Sci. Total Environ. 2018, 610–611, 111–120. [Google Scholar] [CrossRef] [PubMed]
- Mannozzi, J.T.; Filbert, V.J.; Mackay, W.J.; Fulford, D.E.; Steele, C.W. Evaluation of radiant catalytic ionization in reducing Escherichia coli, Listeria innocua and Salmonella typhimurium on represantative food contact surfaces. Texas J. Sci. 2018, 70, 6. [Google Scholar] [CrossRef]
- Vatansever, F.; de Melo, W.C.; Avci, P.; Vecchio, D.; Sadasivam, M.; Gupta, A.; Chandran, R.; Karimi, M.; Parizotto, N.A.; Yin, R.; et al. Antimicrobial strategies centered around reactive oxygen species--bactericidal antibiotics, photodynamic therapy, and beyond. FEMS Microbiol. Rev. 2013, 37, 955–989. [Google Scholar] [CrossRef] [Green Version]
- Dryden, M. Reactive oxygen species: A novel antimicrobial. Int. J. Antimicrob Agents 2018, 51, 299–303. [Google Scholar] [CrossRef]
- Soni, K.A.; Nannapaneni, R. Removal of Listeria monocytogenes biofilms with bacteriophage P100. J. Food Prot. 2010, 73, 1519–1524. [Google Scholar] [CrossRef]
- Niemira, B.A. Irradiation sensitivity of planktonic and biofilm-associated Escherichia coli O157:H7 isolates is influenced by culture conditions. Appl. Environ. Microbiol. 2007, 73, 3239–3244. [Google Scholar] [CrossRef] [Green Version]
- Vidács, A.; Kerekes, E.; Rajkó, R.; Petkovits, T.; Alharbi, N.S.; Khaled, J.M.; Vágvölgyi, C.; Krisch, J. Optimization of essential oil-based natural disinfectants against Listeria monocytogenes and Escherichia coli biofilms formed on polypropylene surfaces. J. Mol. Liq. 2018, 255, 257–262. [Google Scholar] [CrossRef]
- Higgins, S.E.; Wolfenden, A.D.; Bielke, L.R.; Pixley, C.M.; Torres-Rodriguez, A.; Vicente, J.L.; Bosseau, D.; Neighbor, N.; Hargis, B.M.; Tellez, G. Application of Ionized Reactive Oxygen Species for Disinfection of Carcasses, Table Eggs, and Fertile Eggs. J. Appl. Poult. Res. 2005, 14, 716–720. [Google Scholar] [CrossRef] [Green Version]
- Misra, N.N.; Jo, C. Applications of cold plasma technology for microbiological safety in meat industry. Trends Food Sci. Technol. 2017, 64, 74–86. [Google Scholar] [CrossRef]
- Yang, S.; Sadekuzzaman, M.; Ha Do, S. Reduction of Listeria monocytogenes on chicken breasts by combined treatment with UV-C light and bacteriophage ListShield. LWT Food Sci. Technol. 2017, 86, 193–200. [Google Scholar] [CrossRef]
- Ritter, A.C.; Santi, L.; Vannini, L.; Beys-da-Silva, W.O.; Gozzi, G.; Yates, J.; Ragni, L.; Brandelli, A. Comparative proteomic analysis of foodborne Salmonella Enteritidis SE86 subjected to cold plasma treatment. Food Microbiol. 2018, 76, 310–318. [Google Scholar] [CrossRef] [PubMed]
- Roh, S.H.; Lee, S.Y.; Park, H.H.; Lee, E.S.; Min, S.C. Effects of the treatment parameters on the efficacy of the inactivation of Salmonella contaminating boiled chicken breast by in-package atmospheric cold plasma treatment. Int. J. Food Microbiol. 2019, 293, 24–33. [Google Scholar] [CrossRef] [PubMed]
- Saini, J.K.; Marsden, J.L.; Getty, K.J.; Fung, D.Y. Advanced oxidation technology with photohydroionization as a surface treatment for controlling Listeria monocytogenes on stainless steel surfaces and ready-to-eat cheese and turkey. Foodborne Pathog. Dis. 2014, 11, 295–300. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; Kalchayanand, N.; Belk, K.E.; Wheeler, T.L. Photohydroionization Reduces Shiga Toxin-Producing Escherichia coli and Salmonella on fresh beef with minimal effects on meat quality. Meat Muscle Biol. 2019, 3, 105–115. [Google Scholar] [CrossRef] [Green Version]
- Chung, H.; Lee, N.; Jo, C.; Shin, D.; Byun, M. Use of gamma irradiation for inactivation of pathogens inoculated into Kimbab, steamed rice rolled by dried laver. Food Control 2007, 18, 108–112. [Google Scholar] [CrossRef]
- Osaili, T.M.; Al-Nabulsi, A.A.; Aljaafreh, T.F. Use of gamma radiation for inactivating Salmonella spp., Escherichia coli O157:H7 and Listeria monocytogenes in tahini halva. Int. J. Food Microbiol. 2018, 2, 20–25. [Google Scholar] [CrossRef]
- Kang, M.; Kim, H.-J.; Jayasena, D.D.; Bae, S.Y.; Yong, I.H.; Lee, M.; Jo, C. Effects of combined treatments of electron-beam irradiation and addition of Leek (Allium tuberosum) extract on reduction of pathogens in pork jerky. Foodborne Pathog. Dis. 2012, 9, 1083–1087. [Google Scholar] [CrossRef]
- Gabriel, A.A.; Vera, D.D.; Lazo, O.M.Y.; Azarcon, V.B.; De Ocampo, C.G.; Marasigan, J.C.; Sandel, G.T. Ultraviolet-C inactivation of Escherichia coli O157:H7, Listeria monocytogenes, Pseudomonas aeruginosa, and Salmonella enterica in liquid egg white. Food Control 2017, 73, 1303–1309. [Google Scholar] [CrossRef]
- Grinshpun, S.A.; Adhikari, A.; Honda, T.; Kim, K.Y.; Toivola, M.; Rao, K.S.; Reponen, T. Control of aerosol contaminants in indoor air: Combining the particle concentration reduction with microbial inactivation. Environ. Sci. Technol. 2007, 41, 606–612. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.; Kim, H.; Beuchat, L.R.; Ryu, J.H. Development of non-pathogenic bacterial biofilms on the surface of stainless steel which are inhibitory to Salmonella enterica. Food Microbiol. 2018, 69, 136–142. [Google Scholar] [CrossRef] [PubMed]
- Fagerlund, A.; Langsrud, S.; Heir, E.; Mikkelsen, M.I.; Møretrø, T. Biofilm Matrix Composition Affects the Susceptibility of Food Associated Staphylococci to Cleaning and Disinfection Agents. Front. Microbiol. 2016, 7, 856. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pang, X.Y.; Yang, Y.S.; Yuk, H.G. Biofilm formation and disinfectant resistance of Salmonella sp. in mono- and dual-species with Pseudomonas aeruginosa. J. Appl. Microbiol. 2017, 123, 651–660. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez-Melcón, C.; Riesco-Peláez, F.; García-Fernández, C.; Alonso-Calleja, C.; Capita, R. Susceptibility of Listeria monocytogenes planktonic cultures and biofilms to sodium hypochlorite and benzalkonium chloride. Food Microbiol. 2019, 82, 533–540. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Wen, Y. The role of bacterial biofilm in persistent infections and control strategies. Int. J. Oral Sci. 2011, 3, 66–73. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sommers, C.H. Irradiation of minimally processed meats. In Microbial Safety of Minimally Processed Foods; Novak, J.S., Sapers, G.M., Juneja, V.K., Eds.; CRC Press: Boca Raton, FL, USA, 2003; pp. 301–318. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Experimental Variant | Strain | Coefficient of Absolute RCI Efficiency (E) | Mean Value of Coefficient of Absolute RCI Efficiency (E) |
|---|---|---|---|
| I | CJE ATCC | 0.720 (± 0.081)* a | 0.723 (±0.323) a |
| CJE 1 | 0.725 (± 0.125) a | ||
| CJE 2 | 0.723 (± 1.188) a | ||
| LMO ATCC | 0.647 (± 0.089) a | 0.629 (± 0.181) a | |
| LMO 1 | 0.622 (± 0.155) a | ||
| LMO 2 | 0.620 (± 0.217) a | ||
| SENT ATCC | 0.612 (± 0.121) a | 0.600 (± 0.076) a | |
| SENT 1 | 0.605 (± 0.061) a | ||
| SENT 2 | 0.584 (± 0.110) a,b | ||
| II | CJE ATCC | 0.375 (± 0.041) c | 0.232 (± 0.011) b |
| CJE 1 | 0.173 (± 0.014) d | ||
| CJE 2 | 0.150 (± 0.030)d | ||
| LMO ATCC | 0.266 (± 0.040) c.d | 0.329 (± 0.061) b | |
| LMO 1 | 0.306 (± 0.084) c | ||
| LMO 2 | 0.409 (± 0.110) b,c | ||
| SENT ATCC | 0.250 (± 0.051) c,d | 0.261 (± 0.024) b | |
| SENT 1 | 0.231 (± 0.022) c.d | ||
| SENT 2 | 0.302 (± 0.058) c | ||
| III | CJE ATCC | 0.155 (± 0.017) d | 0.162 (± 0.005) c |
| CJE 1 | 0.162 (± 0.008) d | ||
| CJE 2 | 0.171 (± 0.039) d | ||
| LMO ATCC | 0.119 (± 0.016) d,e | 0.087 (± 0.022) c | |
| LMO 1 | 0.049 (± 0.009) e | ||
| LMO 2 | 0.092 (± 0.014) e | ||
| SENT ATCC | 0.083 (± 0.038) e | 0.088 (± 0.028) c | |
| SENT 1 | 0.102 (± 0.015) d,e | ||
| SENT 2 | 0.079 (± 0.025) e |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Skowron, K.; Skowron, K.J.; Bauza-Kaszewska, J.; Wałecka-Zacharska, E.; Kwiecińska-Piróg, J.; Grudlewska-Buda, K.; Wiktorczyk, N.; Gospodarek-Komkowska, E. The Antimicrobial Effect of Radiant Catalytic Ionization on the Bacterial Attachment and Biofilm Formation by Selected Foodborne Pathogens under Refrigeration Conditions. Appl. Sci. 2020, 10, 1364. https://doi.org/10.3390/app10041364
Skowron K, Skowron KJ, Bauza-Kaszewska J, Wałecka-Zacharska E, Kwiecińska-Piróg J, Grudlewska-Buda K, Wiktorczyk N, Gospodarek-Komkowska E. The Antimicrobial Effect of Radiant Catalytic Ionization on the Bacterial Attachment and Biofilm Formation by Selected Foodborne Pathogens under Refrigeration Conditions. Applied Sciences. 2020; 10(4):1364. https://doi.org/10.3390/app10041364
Chicago/Turabian StyleSkowron, Krzysztof, Karolina Jadwiga Skowron, Justyna Bauza-Kaszewska, Ewa Wałecka-Zacharska, Joanna Kwiecińska-Piróg, Katarzyna Grudlewska-Buda, Natalia Wiktorczyk, and Eugenia Gospodarek-Komkowska. 2020. "The Antimicrobial Effect of Radiant Catalytic Ionization on the Bacterial Attachment and Biofilm Formation by Selected Foodborne Pathogens under Refrigeration Conditions" Applied Sciences 10, no. 4: 1364. https://doi.org/10.3390/app10041364
APA StyleSkowron, K., Skowron, K. J., Bauza-Kaszewska, J., Wałecka-Zacharska, E., Kwiecińska-Piróg, J., Grudlewska-Buda, K., Wiktorczyk, N., & Gospodarek-Komkowska, E. (2020). The Antimicrobial Effect of Radiant Catalytic Ionization on the Bacterial Attachment and Biofilm Formation by Selected Foodborne Pathogens under Refrigeration Conditions. Applied Sciences, 10(4), 1364. https://doi.org/10.3390/app10041364