Lab-Scale Optimization of Aurantiochytrium sp. Culture Medium for Improved Growth and DHA Production

 ,
,  ,
,  , and
, and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Thraustochytrid Strain
2.2. Experimental Trials
2.2.1. Effect of Basal Culture Medium and Trace Metals Solution
2.2.2. C Source Optimization
2.2.3. N Source Optimization
2.2.4. Non-Optimized vs. Optimized Conditions
2.3. Growth Assessment
2.4. pH Control
2.5. Determination of Glucose Concentration
2.6. Biochemical Composition
2.6.1. Proximate Composition
Determination of Total Lipid Content
Determination of Ash Content
Determination of Protein and Carbohydrate Contents
2.6.2. Fatty Acid Profile
2.7. Statistical Analysis
3. Results
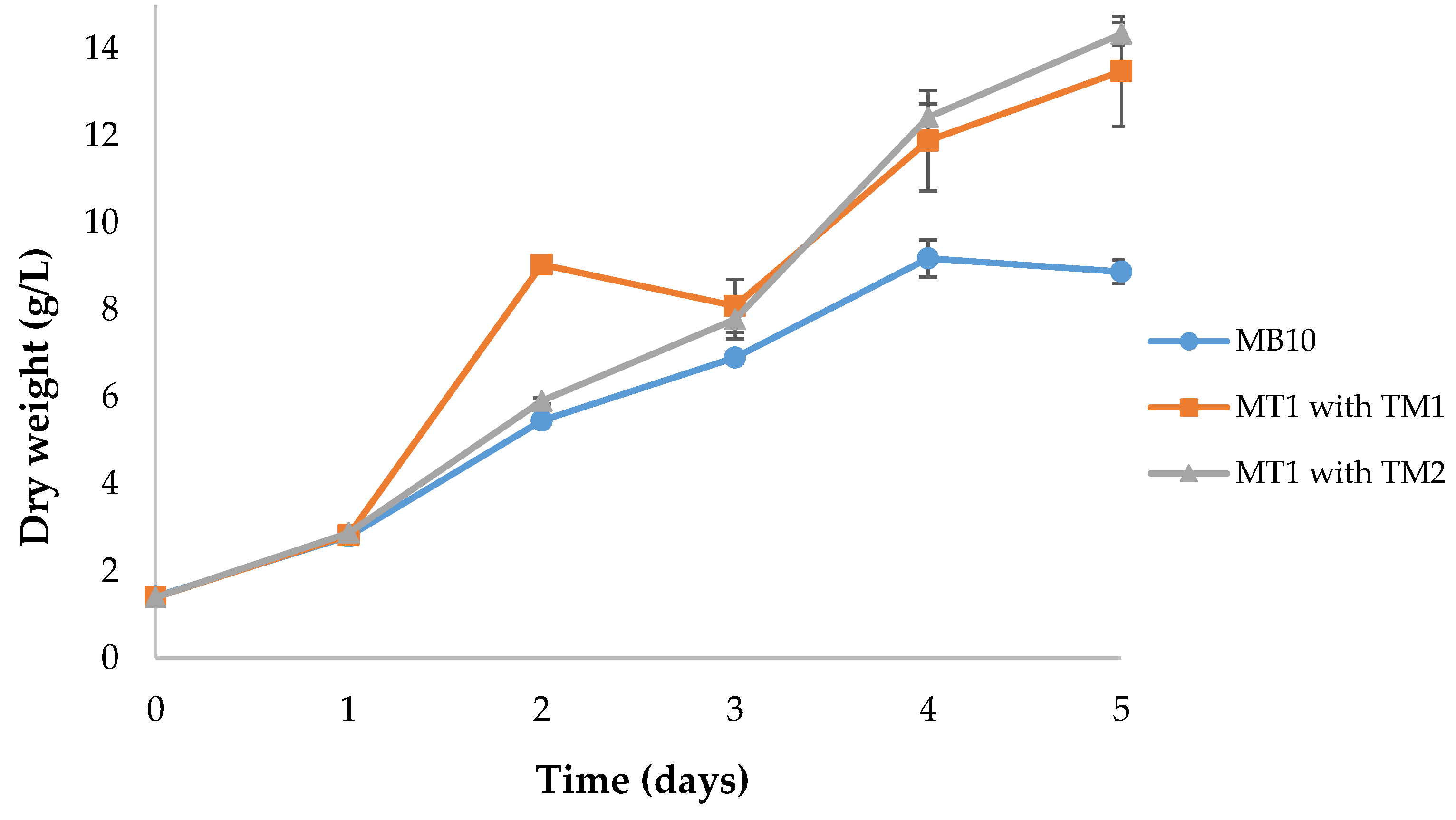
3.1. Effect of Different Medium and Trace Metals Composition on Growth
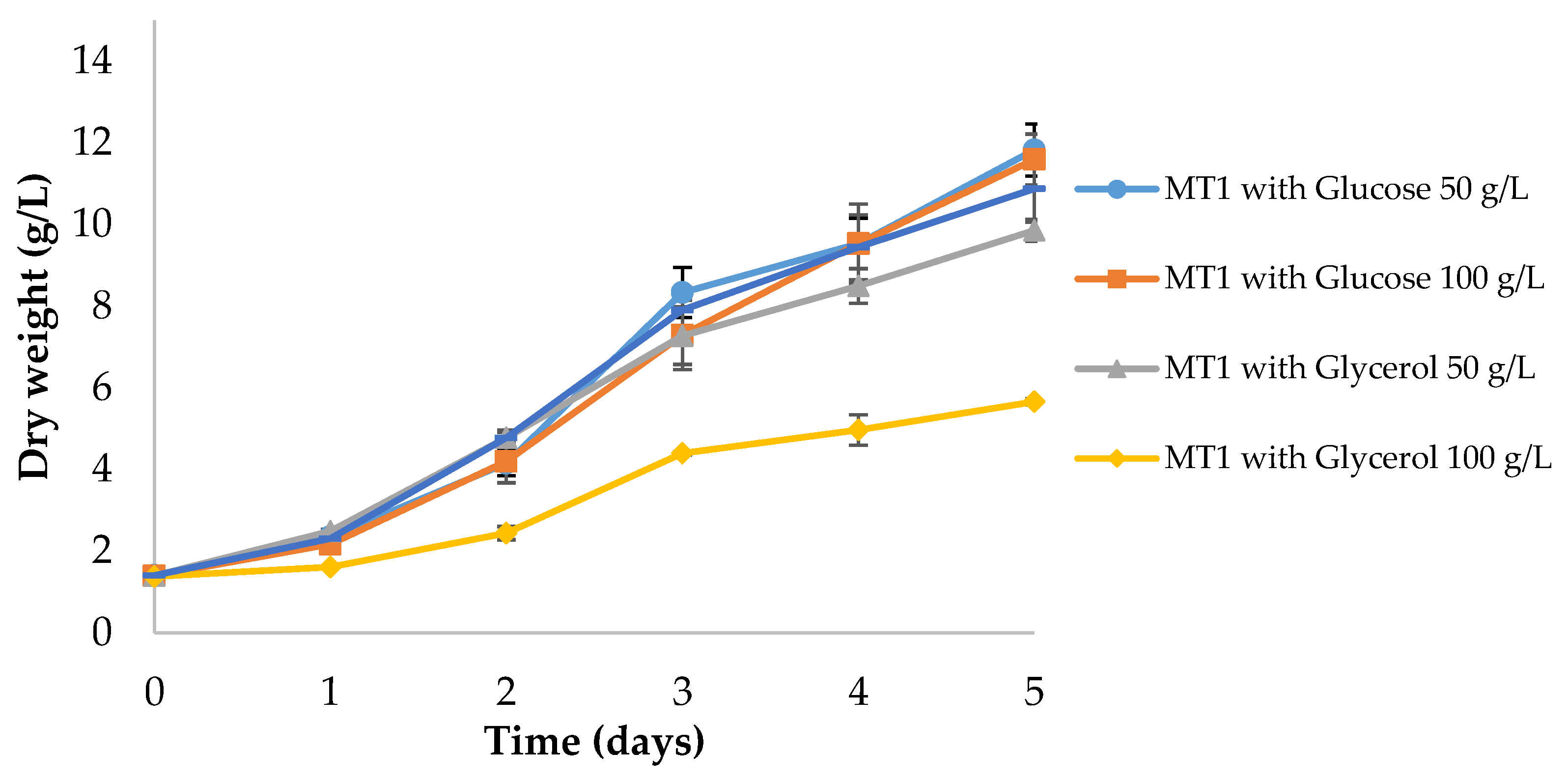
3.2. Carbon Source Optimization
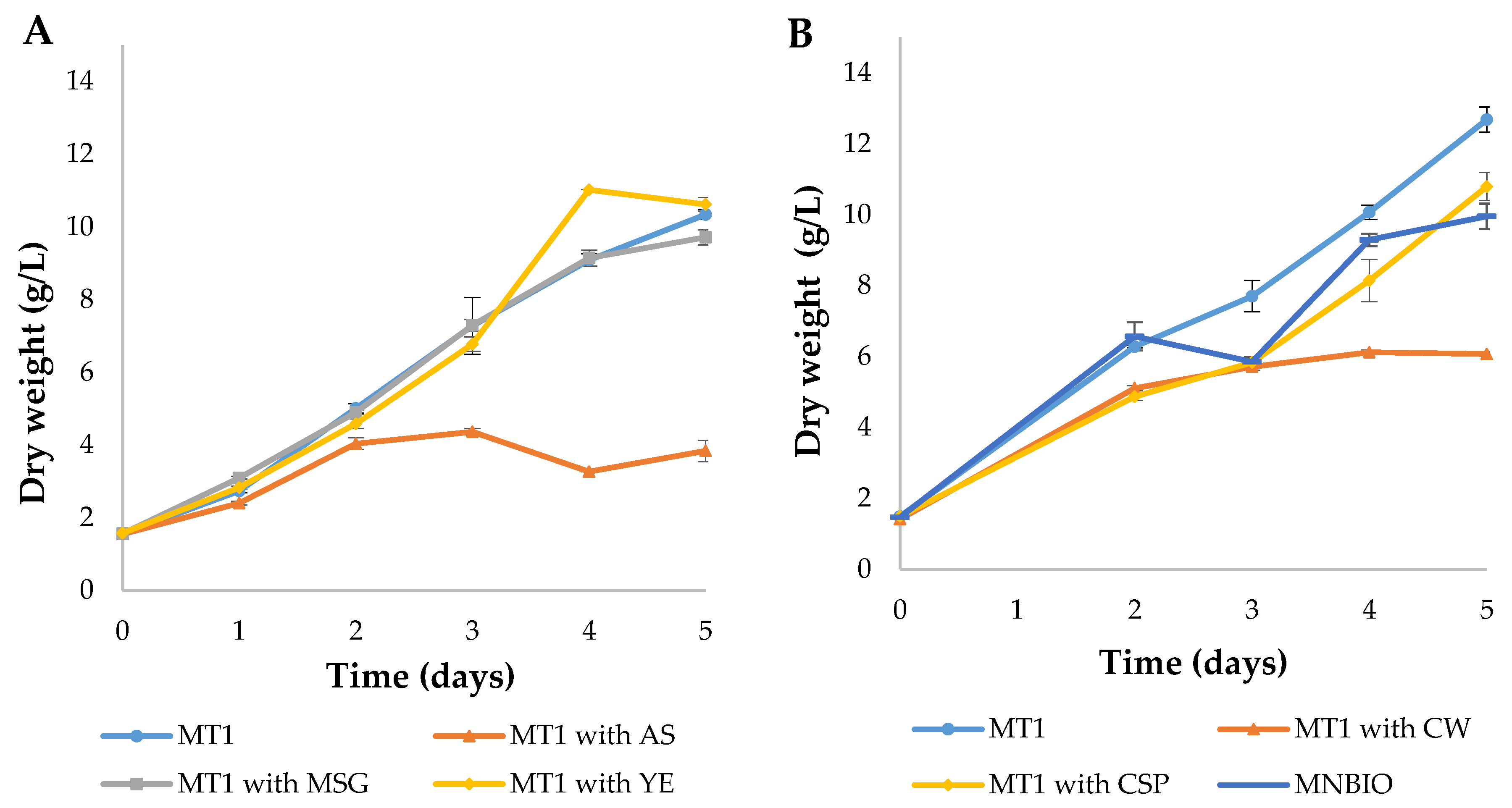
3.3. N Source Optimization
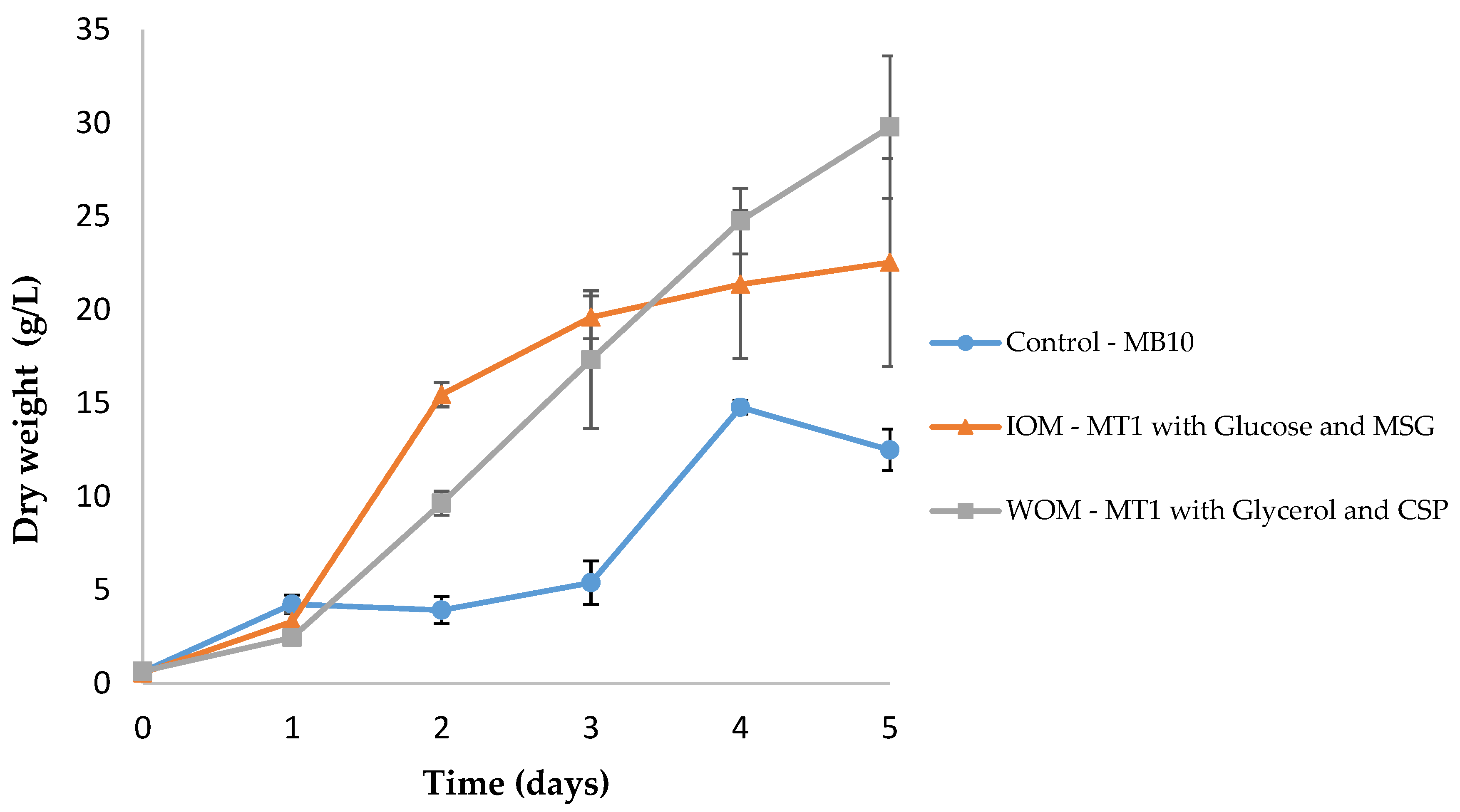
3.4. Non-Optimized vs. Optimized Conditions
3.5. Biochemical Composition of Biomass
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Ruggiero, M.A.; Gordon, D.P.; Orrell, T.M.; Bailly, N.; Bourgoin, T.; Brusca, R.C.; Cavalier-Smith, T.; Guiry, M.; Kirk, P.M. A higher level classification of all living organisms. PLoS ONE 2015, 10, 54. [Google Scholar]
- Jakobsen, A.N.; Aasen, I.M.; Josefsen, K.D.; Strøm, A.R. Accumulation of docosahexaenoic acid-rich lipid in thraustochytrid Aurantiochytrium sp. strain T66: Effects of N and P starvation and O2 limitation. Appl. Microbiol. Biotechnol. 2008, 80, 297. [Google Scholar] [CrossRef] [PubMed]
- Leyland, B.; Leu, S.; Boussiba, S. Are thraustochytrids algae? Fungal Biol. 2017, 121, 835–840. [Google Scholar] [CrossRef] [PubMed]
- Marchan, L.F.; Chang, K.J.L.; Nichols, P.D.; Mitchell, W.J.; Polglase, J.L.; Gutierrez, T. Taxonomy, ecology and biotechnological applications of thraustochytrids: A review. Biotechnol. Adv. 2018, 36, 26–46. [Google Scholar] [CrossRef]
- Aasen, I.M.; Ertesvag, H.; Heggeset, T.M.B.; Liu, B.; Brautaset, T.; Vadstein, O.; Ellingsen, T.E. Thraustochytrids as production organisms for docosahexaenoic acid (DHA), squalene, and carotenoids. Appl. Microbiol. Biotechnol. 2016, 100, 4309–4321. [Google Scholar] [CrossRef]
- Guo, D.S.; Ji, X.J.; Ren, L.J.; Li, G.L.; Yin, F.W.; Huang, H. Development of a real-time bioprocess monitoring method for docosahexaenoic acid production by Schizochytrium sp. Bioresour. Technol. 2016, 216, 422–427. [Google Scholar] [CrossRef] [Green Version]
- Gupta, A.; Barrow, C.J.; Puri, M. Omega-3 biotechnology: Thraustochytrids as a novel source of omega-3 oils. Biotechnol. Adv. 2012, 30, 1733–1745. [Google Scholar] [CrossRef]
- Ren, L.J.; Ji, X.J.; Huang, H.; Qu, L.A.; Feng, Y.; Tong, Q.Q.; Ouyang, P.K. Development of a stepwise aeration control strategy for efficient docosahexaenoic acid production by Schizochytrium sp. Appl. Microbiol. Biotechnol. 2010, 87, 1649–1656. [Google Scholar] [CrossRef]
- Richardson, A.J.; Burton, J.R.; Sewell, R.P.; Spreckelsen, T.F.; Montgomery, P. Docosahexaenoic acid for reading, cognition and behavior in children aged 7–9 years: A randomized, controlled trial (The DOLAB Study). PLoS ONE 2012, 7, 14. [Google Scholar] [CrossRef]
- Ren, L.J.; Sun, L.N.; Zhuang, X.Y.; Qu, L.; Ji, X.J.; Huang, H. Regulation of docosahexaenoic acid production by Schizochytrium sp.: Effect of nitrogen addition. Bioprocess Biosyst. Eng. 2014, 37, 865–872. [Google Scholar] [CrossRef]
- Ling, X.P.; Guo, J.; Liu, X.T.; Zhang, X.; Wang, N.; Lu, Y.H.; Ng, I.S. Impact of carbon and nitrogen feeding strategy on high production of biomass and docosahexaenoic acid (DHA) by Schizochytrium sp. LU310. Bioresour. Technol. 2015, 184, 139–147. [Google Scholar] [CrossRef] [PubMed]
- Allaire, J.; Harris, W.S.; Vors, C.; Charest, A.; Marin, J.; Jackson, K.H.; Tchernofd, A.; Couturea, P.; Lamarchea, B. Supplementation with high-dose docosahexaenoic acid increases the Omega-3 Index more than high-dose eicosapentaenoic acid. Prostaglandins Leukot. Essent. Fatty Acids 2017, 120, 8–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hadley, K.B.; Bauer, J.; Milgram, N.W. The oil-rich alga Schizochytrium sp. as a dietary source of docosahexaenoic acid improves shape discrimination learning associated with visual processing in a canine model of senescence. Prostaglandins Leukot. Essent. Fatty Acids 2017, 118, 10–18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ren, L.J.; Sun, X.M.; Ji, X.J.; Chen, S.L.; Guo, D.S.; Huang, H. Enhancement of docosahexaenoic acid synthesis by manipulation of antioxidant capacity and prevention of oxidative damage in Schizochytrium sp. Bioresour. Technol. 2017, 223, 141–148. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chang, J.P.C.; Su, K.P.; Mondelli, V.; Pariante, C.M. Omega-3 polyunsaturated fatty acids in youths with attention deficit hyperactivity disorder: A systematic review and meta- analysis of clinical trials and biological studies. Neuropsychopharmacology 2018, 43, 534–545. [Google Scholar] [CrossRef] [Green Version]
- Hoang, L.A.T.; Nguyen, H.C.; Le, T.T.; Hoang, T.H.Q.; Pham, V.N.; Hoang, M.H.T.; Ngo, H.T.T.; Hong, D.D. Different fermentation strategies by Schizochytrium mangrovei strain pq6 to produce feedstock for exploitation of squalene and omega-3 fatty acids. J. Phycol. 2018, 54, 550–556. [Google Scholar] [CrossRef]
- Guo, D.S.; Ji, X.J.; Ren, L.J.; Li, G.L.; Sun, X.M.; Chen, K.Q.; Gao, S.; Huang, H. Development of a scale-up strategy for fermentative production of docosahexaenoic acid by Schizochytrium sp. Chem. Eng. Sci. 2018, 176, 600–608. [Google Scholar] [CrossRef]
- Da Silva, P.S.; Mediano, M.F.F.; da Silva, G.M.S.; de Brito, P.D.; Cardoso, C.S.D.; de Almeida, C.F.; Sangenis, L.H.C.; Pinheiro, R.O.; Hasslocher-Moreno, A.M.; Brasil, P.E.A.A.; et al. Omega-3 supplementation on inflammatory markers in patients with chronic Chagas cardiomyopathy: A randomized clinical study. Nutr. J. 2017, 16, 9. [Google Scholar] [CrossRef]
- Sparkes, C.; Gibson, R.; Sinclair, A.; Else, P.L.; Meyer, B.J. Effect of low dose docosahexaenoic acid-rich fish oil on plasma lipids and lipoproteins in pre-menopausal women: A dose-response randomized placebo-controlled trial. Nutrients 2018, 10, 1460. [Google Scholar] [CrossRef] [Green Version]
- Sun, X.M.; Ren, L.J.; Bi, Z.Q.; Ji, X.J.; Zhao, Q.Y.; Jiang, L.; Huang, H. Development of a cooperative two-factor adaptive-evolution method to enhance lipid production and prevent lipid peroxidation in Schizochytrium sp. Biotechnol. Biofuels 2018, 11, 16. [Google Scholar] [CrossRef] [Green Version]
- Wu, S.T.; Yu, S.T.; Lin, L.P. Effect of culture conditions on docosahexaenoic acid production by Schizochytrium sp. S31. Process Biochem. 2005, 40, 3103–3108. [Google Scholar] [CrossRef]
- Allaire, J.; Couture, P.; Leclerc, M.; Charest, A.; Marin, J.; Lepine, M.C.; Talbot, D.; Tchernof, A.; Lamarche, B. A randomized, crossover, head-to-head comparison of eicosapentaenoic acid and docosahexaenoic acid supplementation to reduce inflammation markers in men and women: The Comparing EPA to DHA (ComparED) Study. Am. J. Clin. Nutr. 2016, 104, 280–287. [Google Scholar] [CrossRef] [PubMed]
- Calder, P.C. Omega-3 fatty acids and inflammatory processes: From molecules to man. Biochem. Soc. Trans. 2017, 45, 1105–1115. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qu, L.; Ren, L.J.; Huang, H. Scale-up of docosahexaenoic acid production in fed-batch fermentation by Schizochytrium sp based on volumetric oxygen-transfer coefficient. Biochem. Eng. J. 2013, 77, 82–87. [Google Scholar] [CrossRef]
- Li, J.; Liu, R.J.; Chang, G.F.; Li, X.Y.; Chang, M.; Liu, Y.F.; Jin, Q.; Wang, X. A strategy for the highly efficient production of docosahexaenoic acid by Aurantiochytrium limacinum SR21 using glucose and glycerol as the mixed carbon sources. Bioresour. Technol. 2015, 177, 51–57. [Google Scholar] [CrossRef]
- Yin, F.W.; Guo, D.S.; Ren, L.J.; Ji, X.J.; Huang, H. Development of a method for the valorization of fermentation wastewater and algal-residue extract in docosahexaenoic acid production by Schizochytrium sp. Bioresour. Technol. 2018, 266, 482–487. [Google Scholar] [CrossRef]
- Ganuza, E.; Anderson, A.J.; Ratledge, C. High-cell-density cultivation of Schizochytrium sp. in an ammonium/pH-auxostat fed-batch system. Biotechnol. Lett. 2008, 30, 1559–1564. [Google Scholar] [CrossRef]
- Wang, K.; Sun, T.; Cui, J.Y.; Liu, L.S.; Bi, Y.Q.; Pei, G.S.; Chen, L.; Zhang, W. Screening of chemical modulators for lipid accumulation in Schizochytrium sp. S31. Bioresour. Technol. 2018, 260, 124–129. [Google Scholar] [CrossRef] [Green Version]
- Chi, Z.Y.; Pyle, D.; Wen, Z.Y.; Frear, C.; Chen, S.L. A laboratory study of producing docosahexaenoic acid from biodiesel-waste glycerol by microalgal fermentation. Process Biochem. 2007, 42, 1537–1545. [Google Scholar] [CrossRef]
- Chen, W.; Zhou, P.P.; Zhu, Y.M.; Xie, C.; Ma, L.; Wang, X.P.; Bao, Z.; Yu, L. Improvement in the docosahexaenoic acid production of Schizochytrium sp. S056 by replacement of sea salt. Bioprocess Biosyst. Eng. 2016, 39, 315–321. [Google Scholar] [CrossRef]
- Patil, K.P.; Gogate, P.R. Improved synthesis of docosahexaenoic acid (DHA) using Schizochytrium limacinum SR21 and sustainable media. Chem. Eng. J. 2015, 268, 187–196. [Google Scholar] [CrossRef]
- Chen, G.Q.; Fan, K.W.; Lu, F.P.; Li, Q.A.; Aki, T.; Chen, F.; Jiang, Y. Optimization of nitrogen source for enhanced production of squalene from thraustochytrid Aurantiochytrium sp. New Biotechnol. 2010, 27, 382–389. [Google Scholar] [CrossRef] [PubMed]
- Guo, D.S.; Ji, X.J.; Ren, L.J.; Yin, F.W.; Sun, X.M.; Huang, H.; Zhen, G. Development of a multi-stage continuous fermentation strategy for docosahexaenoic acid production by Schizochytrium sp. Bioresour. Technol. 2018, 269, 32–39. [Google Scholar] [CrossRef] [PubMed]
- Bailey, R.B.; DiMasi, D.; Hansen, J.M.; Mirrasoul, P.J.; Ruecker, C.M.; Veeder, G.T., III; Kaneko, T.; Barclay, W.R. Enhanced Production of Lipids Containing Polyenoic Fatty Acid by Very High Density Cultures of Eukaryotic Microbes in Fermentors. U.S. Patent No. 7,732,170, 7 February 2008. [Google Scholar]
- Chang, G.F.; Gao, N.S.; Tian, G.W.; Wu, Q.H.; Chang, M.; Wang, X.G. Improvement of docosahexaenoic acid production on glycerol by Schizochytrium sp. S31 with constantly high oxygen transfer coefficient. Bioresour. Technol. 2013, 142, 400–406. [Google Scholar] [CrossRef]
- Bligh, E.G.; Dyer, W.J. A rapid method of total lipid extractionand purification. Can. J. Biochem. Physiol. 1959, 37, 911–917. [Google Scholar] [CrossRef] [Green Version]
- Pereira, H.; Barreira, L.; Mozes, A.; Florindo, C.; Polo, C.; Duarte, C.V.; Custódio, L.; Varela, J. Microplatebased high throughput screening procedure for the isolation of lipid-rich marine microalgae. Biotechnol. Biofuels 2011, 4, 12. [Google Scholar] [CrossRef] [Green Version]
- Merrill, A.L.; Watt, B.K. Energy value of foods: Basis and derivation. Agriculture Handbook; US Department of Agriculture, Forest Service: Washington, DC, USA, 1973; p. 74.
- Lepage, G.; Roy, C.C. Improved recovery of fatty-acid through direct transesterification without prior extraction or purification. J. Lipid Res. 1984, 25, 1391–1396. [Google Scholar]
- Pereira, H.; Barreira, L.; Figueiredo, F.; Custodio, L.; Vizetto-Duarte, C.; Polo, C.; Resek, E.; Engelen, A.; Varela, J. Polyunsaturated fatty acids of marine macroalgae: Potential for nutritional and pharmaceutical applications. Mar. Drugs 2012, 10, 1920–1935. [Google Scholar] [CrossRef] [Green Version]
- Bindea, M.; Rusu, B.; Rusu, A.; Trif, M.; Leopold, L.F.; Dulf, F.; Vodnar, D.C. Valorification of crude glycerol for pure fractions of docosahexaenoic acid and β-carotene production by using Schizochytrium limacinum and Blakeslea trispora. Microb. Cell Factories 2018, 17, 97. [Google Scholar] [CrossRef]
- Huang, T.Y.; Lu, W.C.; Chu, I.M. A fermentation strategy for producing docosahexaenoic acid in Aurantiochytrium limacinum SR21 and increasing C22:6 proportions in total fatty acid. Bioresour. Technol. 2012, 123, 8–14. [Google Scholar] [CrossRef]
- Yu, X.J.; Yu, Z.Q.; Liu, Y.L.; Sun, J.; Zheng, J.Y.; Wang, Z. Utilization of high-fructose corn syrup for biomass production containing high levels of docosahexaenoic acid by a newly isolated Aurantiochytrium sp. YLH70. Appl. Biochem. Biotechnol. 2015, 177, 1229–1240. [Google Scholar] [CrossRef] [PubMed]
- Yokochi, T.; Honda, D.; Higashihara, T.; Nakahara, T. Optimization of docosahexaenoic acid production by Schizochytrium limacinum SR21. Appl. Microbiol. Biotechnol. 1998, 49, 72–76. [Google Scholar] [CrossRef]
- Yokoyama, R.; Honda, D. Taxonomic rearrangement of the genus Schizochytrium sensu lato based on morphology, chemotaxonomic characteristics, and 18S rRNA gene phylogeny (Thraustochytriaceae, Labyrinthulomycetes): Emendation for Schizochytrium and erection of Aurantiochytrium and Oblongichytrium gen. nov. Mycoscience 2007, 48, 199–211. [Google Scholar]
- Taoka, Y.; Nagano, N.; Okita, Y.; Izumida, H.; Sugimoto, S.; Hayashi, M. Influences of culture temperature on the growth, lipid content and fatty acid composition of Aurantiochytrium sp. strain mh0186. Mar. Biotechnol. 2009, 11, 368–374. [Google Scholar] [CrossRef]
- Nakazawa, A.; Matsuura, H.; Kose, R.; Ito, K.; Ueda, M.; Honda, D.; Inouyea, I.; Kayaa, K.; Watanabe, M.M. Optimization of biomass and fatty acid production by Aurantiochytrium sp. strain 4W-1b. Innov. Res. Algal Biomass 2012, 15, 27–33. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Condition | Base Medium | N Source | C Source |
|---|---|---|---|
| Control | MB10 | Ammonium sulfate and Glutamate | Glucose |
| IOM | MT1 | Glutamate | Glucose |
| WOM | MT1 | Corn steep powder | Glycerol |
| Medium | Proteins (%) | Lipids (%) | Carbohydrates (%) | Ashes (%) |
|---|---|---|---|---|
| MB10 | 23.57 ± 0.65 a | 22.78 ± 0.20 a | 28.13 ± 0.69 a | 25.52 ± 2.46 a |
| IOM | 14.02 ± 0.04 c | 14.74 ± 0.15 c | 59.05 ± 3.47 b | 12.19 ± 0.05 b |
| WOM | 20.55 ± 0.33 b | 31.14 ± 3.41 b | 32.99 ± 2.71 a | 15.31 ± 0.36 b |
| FAME | MB10 | IOM | WOM |
|---|---|---|---|
| C14:0 | 14.53 ± 0.52 | 8.22 ± 4.14 | 9.40 ± 0.49 |
| C15:0 | 0.58 ± 0.05 | 1.71 ± 1.71 | 2.96 ± 0.18 |
| C16:1 | 2.73 ± 1.07 | 2.20 ± 0.91 | 3.12 ± 0.10 |
| C16:0 | 72.26 ± 0.27 | 64.21 ± 8.31 | 46.32 ± 1.22 |
| C17:0 | 0.04 ± 0.04 | 0.33 ± 0.33 | 1.16 ± 0.14 |
| C18:0 | 0.24 ± 0.20 | 0.55 ± 0.55 | 1.68 ± 0.58 |
| C22:5 (DPA) | 0.19 ± 0.11 | 5.74 ± 3.93 | 5.98 ± 5.98 |
| C22:6n-3 (DHA) | 10.12 ± 2.13 | 17.05 ± 11.10 | 29.66 ± 3.28 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Trovão, M.; Pereira, H.; Costa, M.; Machado, A.; Barros, A.; Soares, M.; Carvalho, B.; Silva, J.T.; Varela, J.; Silva, J. Lab-Scale Optimization of Aurantiochytrium sp. Culture Medium for Improved Growth and DHA Production. Appl. Sci. 2020, 10, 2500. https://doi.org/10.3390/app10072500
Trovão M, Pereira H, Costa M, Machado A, Barros A, Soares M, Carvalho B, Silva JT, Varela J, Silva J. Lab-Scale Optimization of Aurantiochytrium sp. Culture Medium for Improved Growth and DHA Production. Applied Sciences. 2020; 10(7):2500. https://doi.org/10.3390/app10072500
Chicago/Turabian StyleTrovão, Mafalda, Hugo Pereira, Margarida Costa, Adriana Machado, Ana Barros, Maria Soares, Bernardo Carvalho, Joana Teles Silva, João Varela, and Joana Silva. 2020. "Lab-Scale Optimization of Aurantiochytrium sp. Culture Medium for Improved Growth and DHA Production" Applied Sciences 10, no. 7: 2500. https://doi.org/10.3390/app10072500
APA StyleTrovão, M., Pereira, H., Costa, M., Machado, A., Barros, A., Soares, M., Carvalho, B., Silva, J. T., Varela, J., & Silva, J. (2020). Lab-Scale Optimization of Aurantiochytrium sp. Culture Medium for Improved Growth and DHA Production. Applied Sciences, 10(7), 2500. https://doi.org/10.3390/app10072500