Isolation of Circulating Tumor Cells from Glioblastoma Patients by Direct Immunomagnetic Targeting

 , and
, and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Lines
2.2. Patients
2.3. Immunocytostaining
2.4. Flowcytometry
2.5. Immunomagnetic Cell Isolation
2.6. Immunocytostaining of Circculating Tumor Cells
3. Results
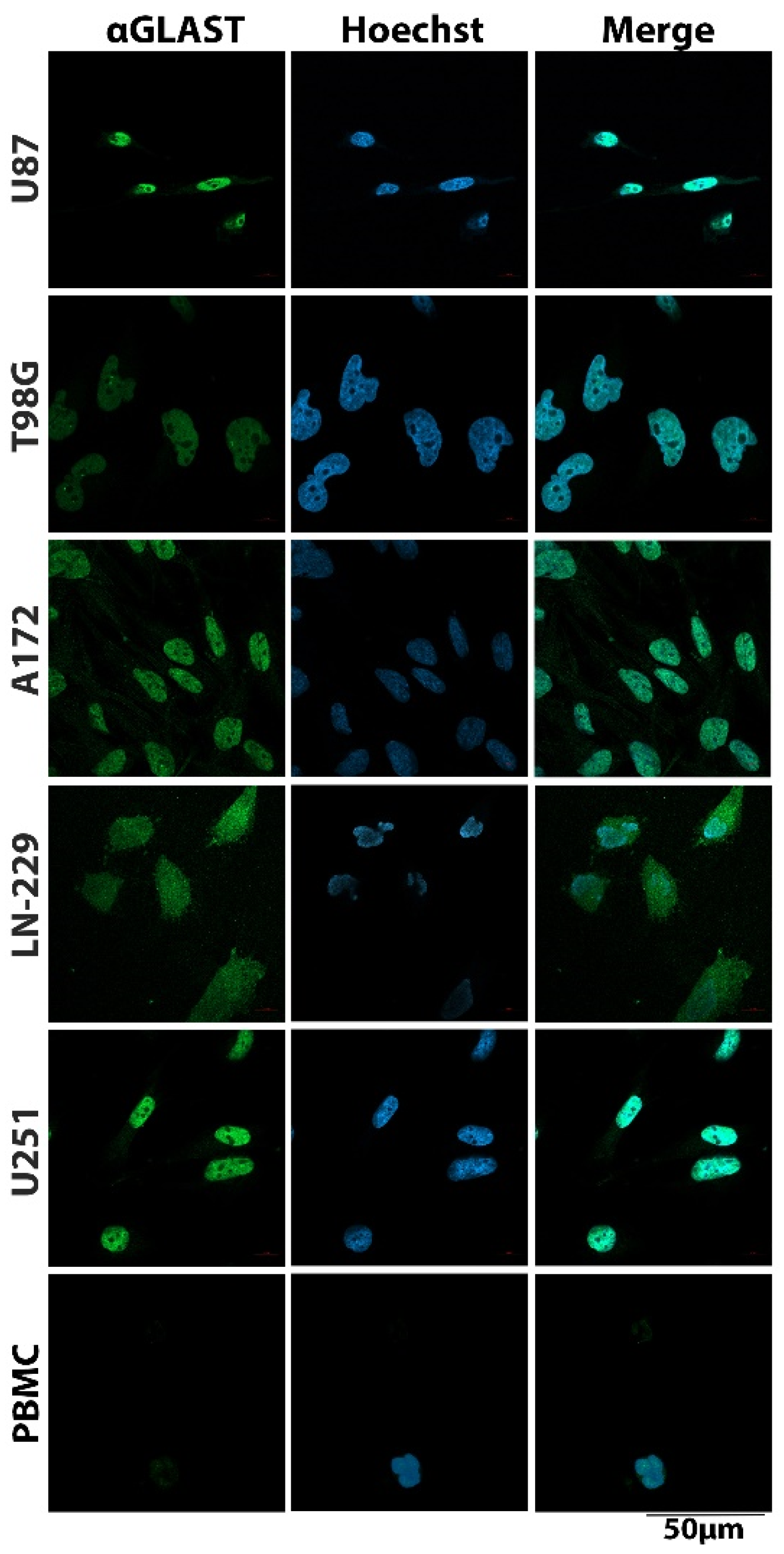
3.1. Cell Surface Protein Expression on Glioblastoma Cells
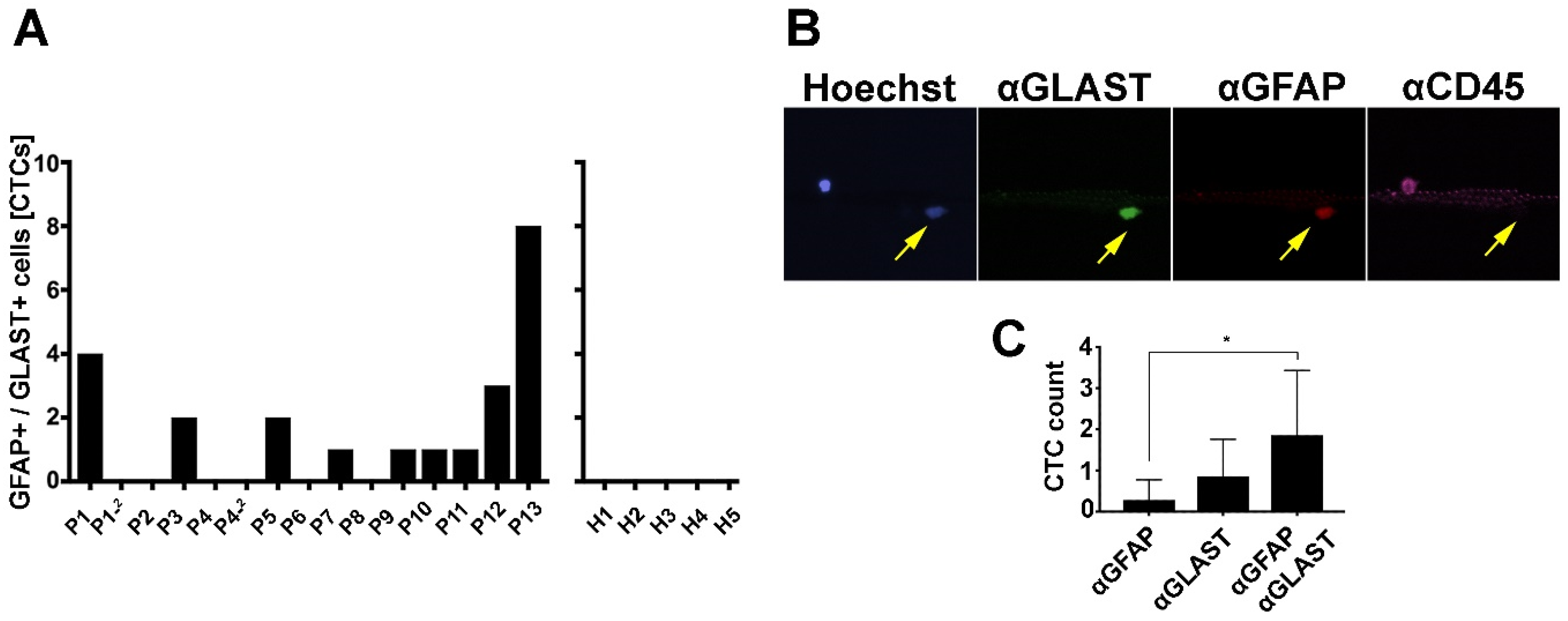
3.2. GBM-CTC Identification Markers
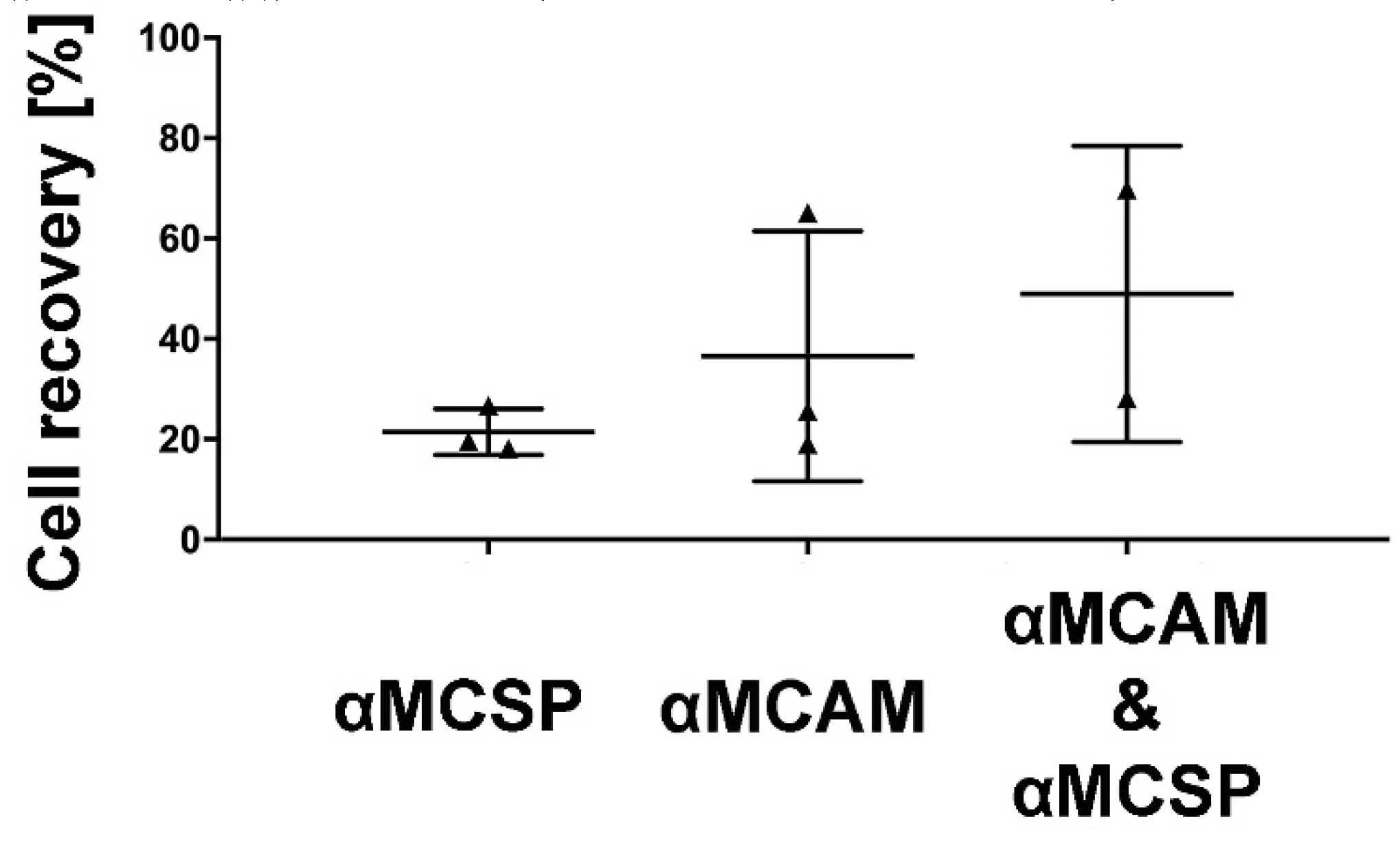
3.3. Immunomagnetic Capture of GBM Cells
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Ostrom, Q.T.; Gittleman, H.; Liao, P.; Rouse, C.; Chen, Y.; Dowling, J.; Wolinsky, Y.; Kruchko, C.; Barnholtz-Sloan, J. CBTRUS statistical report: Primary brain and central nervous system tumors diagnosed in the United States in 2007–2011. Neuro-Oncology 2014, 16, iv1–iv63. [Google Scholar] [CrossRef] [PubMed]
- Omuro, A.; DeAngelis, L.M. Glioblastoma and other malignant gliomas: A clinical review. JAMA 2013, 310, 1842–1850. [Google Scholar] [CrossRef] [PubMed]
- de Bono, J.S.; Scher, H.I.; Montgomery, R.B.; Parker, C.; Miller, M.C.; Tissing, H.; Doyle, G.V.; Terstappen, L.W.; Pienta, K.J.; Raghavan, D. Circulating tumor cells predict survival benefit from treatment in metastatic castration-resistant prostate cancer. Clin. Cancer Res. 2008, 14, 6302–6309. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alva, A.; Friedlander, T.; Clark, M.; Huebner, T.; Daignault, S.; Hussain, M.; Lee, C.; Hafez, K.; Hollenbeck, B.; Weizer, A. Circulating tumor cells as potential biomarkers in bladder cancer. J. Urol. 2015, 194, 790–798. [Google Scholar] [CrossRef] [PubMed]
- Hayes, D.F.; Cristofanilli, M.; Budd, G.T.; Ellis, M.J.; Stopeck, A.; Miller, M.C.; Matera, J.; Allard, W.J.; Doyle, G.V.; Terstappen, L.W. Circulating tumor cells at each follow-up time point during therapy of metastatic breast cancer patients predict progression-free and overall survival. Clin. Cancer Res. 2006, 12, 4218–4224. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Becker, T.M.; Caixeiro, N.J.; Lim, S.H.; Tognela, A.; Kienzle, N.; Scott, K.F.; Spring, K.J.; Souza, P. New frontiers in circulating tumor cell analysis: A reference guide for biomolecular profiling toward translational clinical use. Int. J. Cancer 2014, 134, 2523–2533. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Best, M.G.; Sol, N.; Zijl, S.; Reijneveld, J.C.; Wesseling, P.; Wurdinger, T. Liquid biopsies in patients with diffuse glioma. Acta Neuropathol. 2015, 129, 849–865. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Müller, C.; Holtschmidt, J.; Auer, M.; Heitzer, E.; Lamszus, K.; Schulte, A.; Matschke, J.; Langer-Freitag, S.; Gasch, C.; Stoupiec, M. Hematogenous dissemination of glioblastoma multiforme. Sci. Transl. Med. 2014, 6, ra101–ra247. [Google Scholar] [CrossRef] [PubMed]
- MacArthur, K.M.; Kao, G.D.; Chandrasekaran, S.; Alonso-Basanta, M.; Chapman, C.; Lustig, R.A.; Wileyto, E.P.; Hahn, S.M.; Dorsey, J.F. Detection of brain tumor cells in the peripheral blood by a telomerase promoter-based assay. Cancer Res. 2014, 74, 2152–2159. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sullivan, J.P.; Nahed, B.V.; Madden, M.W.; Oliveira, S.M.; Springer, S.; Bhere, D.; Chi, A.S.; Wakimoto, H.; Rothenberg, S.M.; Sequist, L.V. Brain tumor cells in circulation are enriched for mesenchymal gene expression. Cancer Discov. 2014, 4, 1299–1309. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gao, F.; Cui, Y.; Jiang, H.; Sui, D.; Wang, Y.; Jiang, Z.; Zhao, J.; Lin, S. Circulating tumor cell is a common property of brain glioma and promotes the monitoring system. Oncotarget 2016, 7, 71330. [Google Scholar] [CrossRef] [PubMed]
- Schiffer, D.; Giordana, M.T.; Germano, I.; Mauro, A. Anaplasia and heterogeneity of GFAP expression in gliomas. Tumori 1986, 72, 163–170. [Google Scholar] [CrossRef] [PubMed]
- Krol, I.; Castro-Giner, F.; Maurer, M.; Gkountela, S.; Szczerba, B.M.; Scherrer, R.; Coleman, N.; Carreira, S.; Bachmann, F.; Anderson, S.; et al. Detection of circulating tumour cell clusters in human glioblastoma. Br. J. Cancer 2018, 119, 487–491. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yaar, M.; Park, H.Y. Melanocytes: A window into the nervous system. J. Investig. Dermatol. 2012, 132, 835–845. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Po, J.W.; Ma, Y.; Balakrishna, B.; Brungs, D.; Azimi, F.; de Souza, P.; Becker, T.M. Immunomagnetic isolation of circulating melanoma cells and detection of PD-L1 status. PLoS ONE 2019, 14, e0211866. [Google Scholar] [CrossRef] [PubMed]
- Gray, E.S.; Reid, A.L.; Bowyer, S.; Calapre, L.; Siew, K.; Pearce, R.; Cowell, L.; Frank, M.H.; Millward, M.; Ziman, M. Circulating Melanoma Cell Subpopulations: Their Heterogeneity and Differential Responses to Treatment. J. Investig. Dermatol. 2015, 135, 2040–2048. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bang-Christensen, S.R.; Pedersen, R.S.; Pereira, M.A.; Clausen, T.M.; Loppke, C.; Sand, N.T.; Ahrens, T.D.; Jorgensen, A.M.; Lim, Y.C.; Goksoyr, L.; et al. Capture and Detection of Circulating Glioma Cells Using the Recombinant VAR2CSA Malaria Protein. Cells 2019, 8, 998. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ye, Z.-C.; Rothstein, J.D.; Sontheimer, H. Compromised glutamate transport in human glioma cells: Reduction–mislocalization of sodium-dependent glutamate transporters and enhanced activity of cystine–glutamate exchange. J. Neurosci. 1999, 19, 10767–10777. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Restrepo, A.; Smith, C.A.; Agnihotri, S.; Shekarforoush, M.; Kongkham, P.N.; Seol, H.J.; Northcott, P.; Rutka, J.T. Epigenetic regulation of glial fibrillary acidic protein by DNA methylation in human malignant gliomas. Neuro-Oncology 2011, 13, 42–50. [Google Scholar] [CrossRef] [PubMed] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Patient | Gender | Age # | Grade | Histology | Tumour Location | ECOG 1 | ECOG 2 |
|---|---|---|---|---|---|---|---|
| 1 | M | 61 | 4 | GBM | Left tempoparietal region | 1 | 0 |
| 2 | M | 47 | 4 | GBM | Right parietal lobe | 0 | 1 |
| 3 | M | 32 | 4 | GBM | Left frontal lobe | 0 | 1 |
| 4 | F | 30 | 4 | GSM * | Right frontoparietal region | 0 | 0 |
| 5 | F | 47 | 4 | GBM | Right temporal lobe | 1 | 0 |
| 6 | M | 75 | 4 | GBM | Cerebellum | 1 | 1 |
| 7 | M | 36 | 4 | GBM | Right parietal lobe | 0 | 1 |
| 8 | M | 33 | 4 | GBM | Right frontal lobe | 0 | 0 |
| 9 | M | 68 | 4 | GBM | Right temporal lobe | 1 | 2 |
| 10 | M | 60 | 4 | GBM | Left frontal lobe | 2 | 2 |
| 11 | F | 50 | 4 | GBM | Bifrontal region | 0 | 3 |
| 12 | F | 71 | 4 | GBM | Left parietal lobe | 1 | 2 |
| 13 | M | 83 | 4 | GBM | Right frontal lobe | 1 | 1 |
| Detectable [% of Cells] | ||||||
|---|---|---|---|---|---|---|
| Antigen | A172 | LN-229 | T98G | U87 | U251 | PBMCs |
| MCAM | 51.0 ± 3.6 | 94.6 ± 1.7 | 64.1 ± 5.4 | 67.8 ± 4.9 | 42.7 ± 0.1 | - |
| MCSP | 80.4 ± 4.1 | 92.1 ± 2.6 | - | 89.3 ± 13.7 | - | - |
| N-Cadherin | 70.2 ± 13.8 | 55.3 ± 0.6 | 17.1 ± 0.6 | - | 94.3 ± 3.3 | - |
| EGFR | 96.8 ± 0.3 | 78.1 ± 1.3 | 29.2 ± 0.4 | 11.2 ± 3.2 | 34.5 ± 8.3 | - |
| CD271 | 5.9 ± 0.1 | 57.3 ± 0.4 | 8.6 ± 10.5 | - | - | - |
| A2B5 | - | - | - | - | n/d | n/d |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lynch, D.; Powter, B.; Po, J.W.; Cooper, A.; Garrett, C.; Koh, E.-S.; Sheridan, M.; van Gelder, J.; Darwish, B.; Mckechnie, S.; et al. Isolation of Circulating Tumor Cells from Glioblastoma Patients by Direct Immunomagnetic Targeting. Appl. Sci. 2020, 10, 3338. https://doi.org/10.3390/app10093338
Lynch D, Powter B, Po JW, Cooper A, Garrett C, Koh E-S, Sheridan M, van Gelder J, Darwish B, Mckechnie S, et al. Isolation of Circulating Tumor Cells from Glioblastoma Patients by Direct Immunomagnetic Targeting. Applied Sciences. 2020; 10(9):3338. https://doi.org/10.3390/app10093338
Chicago/Turabian StyleLynch, David, Branka Powter, Joseph William Po, Adam Cooper, Celine Garrett, Eng-Siew Koh, Mark Sheridan, James van Gelder, Balsam Darwish, Simon Mckechnie, and et al. 2020. "Isolation of Circulating Tumor Cells from Glioblastoma Patients by Direct Immunomagnetic Targeting" Applied Sciences 10, no. 9: 3338. https://doi.org/10.3390/app10093338
APA StyleLynch, D., Powter, B., Po, J. W., Cooper, A., Garrett, C., Koh, E. -S., Sheridan, M., van Gelder, J., Darwish, B., Mckechnie, S., Bazina, R., Jaeger, M., Roberts, T. L., de Souza, P., & Becker, T. M. (2020). Isolation of Circulating Tumor Cells from Glioblastoma Patients by Direct Immunomagnetic Targeting. Applied Sciences, 10(9), 3338. https://doi.org/10.3390/app10093338