
Heterologous Expression and Characterization of a Ferulic Acid Esterase from Aspergillus aculeatus with Potential Use in Sunflower Seed Processing



Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals and Substrates
2.2. Coding Sequence Analysis
2.3. Expression of Wild Type C (WTC) FAE
2.4. Production of reFAEs in E. coli BL21
2.5. Optimization of Induction Conditions
2.6. Separation and Purification of reFAEs
2.7. Activity Assay
2.8. Biochemical Characterization of reFAEs
2.9. Substrate Specificity
2.10. Application Trials
2.11. Statistical Analysis
3. Results and Discussion
3.1. Sequence Comparison to Known Esterases
3.2. Optimal Conditions for the Expression of Recombinant Vectors in E. coli
3.3. Purification of the reFAEs
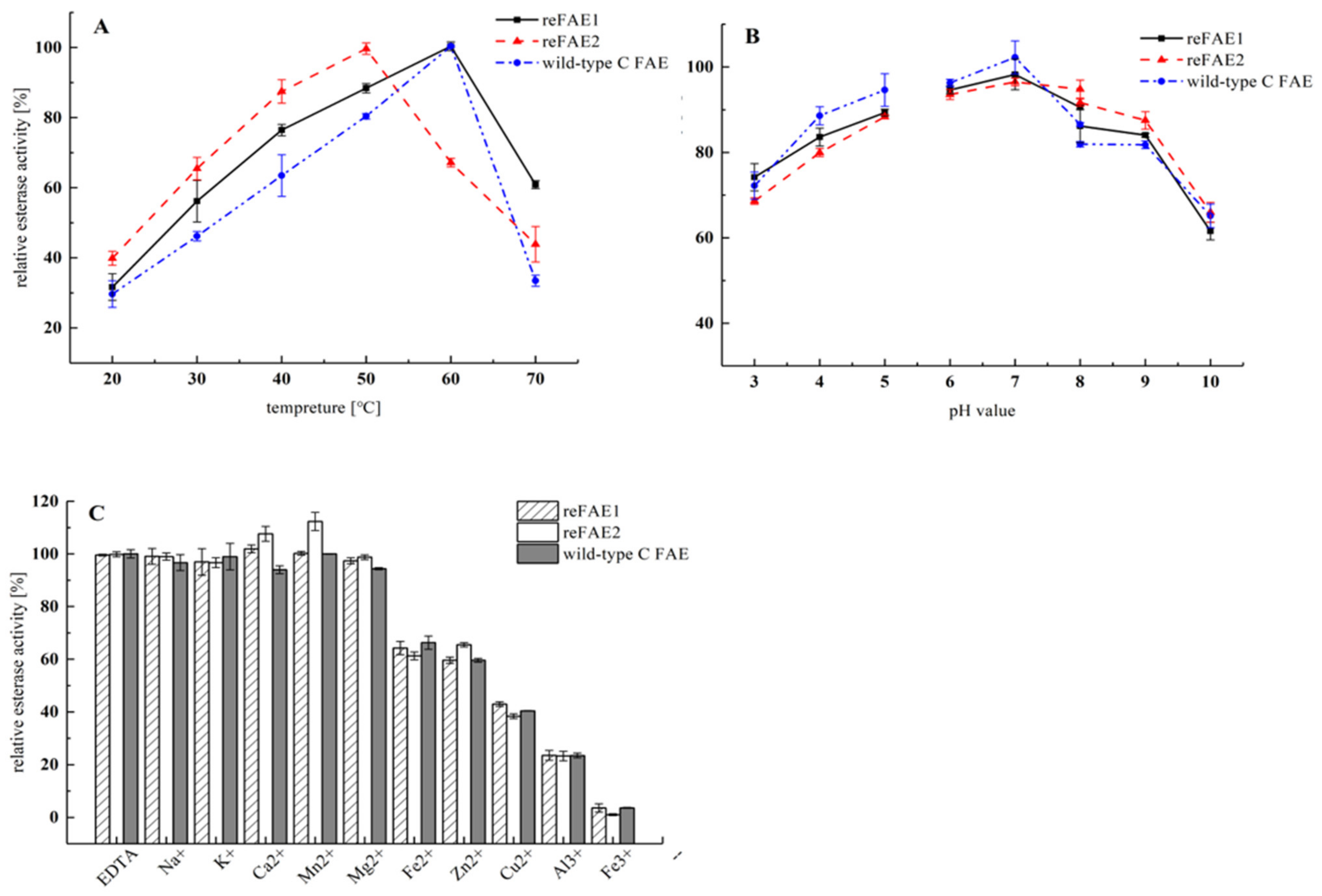
3.4. Biochemical Characterization of reFAEs
3.5. Substrate Profile and Kinetic Parameters
3.6. Application of reFAE2 in SSP Solution
4. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Wijayanti, I.; Ysa, Z.; Khotijah, L. The Evaluation of Sunflower Seed Meal as Protein Source in Lamb Ration. IOP Conf. Ser. Environ. Earth Sci. 2020, 478, 012028. [Google Scholar] [CrossRef]
- Liang, S.; Were, L.M. Chlorogenic acid induced colored reactions and their effect on carbonyls, phenolic content, and antioxidant capacity in sunflower butter cookies. LWT Food Sci. Technol. 2018, 87, 16–22. [Google Scholar] [CrossRef] [Green Version]
- Zardo, I.; de Espíndola Sobczyk, A.; Marczak, L.D.F.; Sarkis, J. Optimization of Ultrasound Assisted Extraction of Phenolic Compounds from Sunflower Seed Cake Using Response Surface Methodology. Waste Biomass Valoris. 2017, 10, 33–44. [Google Scholar] [CrossRef]
- Głowacka, R.; Górska, A.; Wirkowska-Wojdyła, M.; Wołosiak, R.; Majewska, E.; Derewiaka, D. The influence of brewing method on bioactive compounds residues in spent coffee grounds of different roasting degree and geographical origin. Int. J. Food Sci. Technol. 2019, 54, 3008–3014. [Google Scholar] [CrossRef]
- Moon, J.K.; Yoo, H.S.; Shibamoto, T. Role of roasting conditions in the level of chlorogenic acid content in coffee beans: Correlation with coffee acidity. J. Agric. Food Chem. 2009, 57, 5365–5369. [Google Scholar] [CrossRef]
- Tai, E.-S.; Hsieh, P.-C.; Sheu, S.-C. Effect of polygalacturonase and feruloyl esterase from Aspergillus tubingensis on demucilage and quality of coffee beans. Process. Biochem. 2014, 49, 1274–1280. [Google Scholar] [CrossRef]
- Butiuk, A.P.; Adachi, O.; Hours, R.A. Yerba mate as a novel inducer for fungal chlorogenate hydrolase production. Biocatal. Agric. Biotechnol. 2015, 4, 327–334. [Google Scholar] [CrossRef]
- De Luca, S.; Ciotoli, E.; Biancolillo, A.; Bucci, R.; Magri, A.D.; Marini, F. Simultaneous quantification of caffeine and chlorogenic acid in coffee green beans and varietal classification of the samples by HPLC-DAD coupled with chemometrics. Environ. Sci. Pollut. Res. Int. 2018, 25, 28748–28759. [Google Scholar] [CrossRef]
- Siebert, M.; Berger, R.G.; Pfeiffer, F. Hydrolysis of chlorogenic acid in apple juice using a p-coumaryl esterase of Rhizoctonia solani. J. Sci. Food Agric. 2019, 99, 6644–6648. [Google Scholar] [CrossRef] [Green Version]
- Wildermuth, S.R.; Young, E.E.; Were, L.M. Chlorogenic Acid Oxidation and Its Reaction with Sunflower Proteins to Form Green-Colored Complexes. Compr. Rev. Food Sci. Food Saf. 2016, 15, 829–843. [Google Scholar] [CrossRef]
- Zhang, W.; Liu, Y.; Hu, M.; Yang, R. Preparation of high-quality sunflower seed protein with a new chlorogenic acid hydrolase from Aspergillus niger. Biotechnol. Lett. 2019, 41, 565–574. [Google Scholar] [CrossRef]
- Duggett, N.; Abuoun, M.; Randall, L.; Horton, R.; Anjum, M.F. The importance of using whole genome sequencing and extended spectrum beta-lactamase selective media when monitoring antimicrobial resistance. Sci. Rep. 2020, 10, 19880. [Google Scholar] [CrossRef]
- Crepin, V.F.; Faulds, C.B.; Connerton, I.F. A non-modular type B feruloyl esterase from Neurospora crassa exhibits concentration-dependent substrate inhibition. Biochem. J. 2003, 370, 417–427. [Google Scholar] [CrossRef] [Green Version]
- Levasseur, A.; Benoit, I.; Asther, M.; Asther, M.; Record, E. Homologous expression of the feruloyl esterase B gene from Aspergillus niger and characterization of the recombinant enzyme. Protein Expres. Purif. 2004, 37, 126–133. [Google Scholar] [CrossRef]
- Koseki, T.; Handa, H.; Watanabe, Y.-Y.; Ohtsuka, M.; Shiono, Y. An unusual feruloyl esterase from Aspergillus oryzae: Two tryptophan residues play a crucial role for the activity. J. Mol. Catal. B Enzym. 2016, 133, S560–S568. [Google Scholar] [CrossRef]
- Yang, L.; Zhou, N.; Tian, Y. Purification, characterization, and biocatalytic potential of a novel dextranase from Chaetomium globosum. Biotechnol. Lett. 2018, 40, 1407–1418. [Google Scholar] [CrossRef]
- Song, J.; Kim, S.Y.; Kim, D.H.; Lee, Y.S.; Sim, J.S.; Hahn, B.S.; Lee, C.M. Characterization of an inhibitor-resistant endo-1,4-beta-mannanase from the gut microflora metagenome of Hermetia illucens. Biotechnol. Lett. 2018, 40, 1377–1387. [Google Scholar] [CrossRef]
- Dilokpimol, A.; Makela, M.R.; Aguilar-Pontes, M.V.; Benoit-Gelber, I.; Hilden, K.S.; de Vries, R.P. Diversity of fungal feruloyl esterases: Updated phylogenetic classification, properties, and industrial applications. Biotechnol. Biofuels 2016, 9, 231. [Google Scholar] [CrossRef] [Green Version]
- Durmus, D. CIELAB color space boundaries under theoretical spectra and 99 test color samples. Color Res. Appl. 2020, 45, 796–802. [Google Scholar] [CrossRef]
- Koseki, T.; Hori, A.; Seki, S.; Murayama, T.; Shiono, Y. Characterization of two distinct feruloyl esterases, AoFaeB and AoFaeC, from Aspergillus oryzae. Appl. Microbiol. Biotechnol. 2009, 83, 689–696. [Google Scholar] [CrossRef]
- Kroon, P.; Faulds, C.; Williamson, G. Purification and characterization of a novel esterase induced by growth of Aspergillus niger on sugar-beet pulp. Biotechnol. Appl. Biochem. 1996, 23, 255–262. [Google Scholar] [CrossRef] [PubMed]
- Galagan, J.E.; Calvo, S.E.; Cuomo, C.; Ma, L.J.; Wortman, J.R.; Batzoglou, S.; Lee, S.I.; Bastuerkmen, M.; Spevak, C.C.; Clutterbuck, J. Sequencing of Aspergillus nidulans and comparative analysis with A. fumigatus and A. oryzae. Nature 2005, 438, 1105–1115. [Google Scholar] [CrossRef] [PubMed]
- Moukouli, M.; Topakas, E.; Christakopoulos, P. Cloning, characterization and functional expression of an alkalitolerant type C feruloyl esterase from Fusarium oxysporum. Appl. Microbiol. Biotechnol. 2008, 79, 245–254. [Google Scholar] [CrossRef] [PubMed]
- Nieter, A.; Haase-Aschoff, P.; Kelle, S.; Linke, D.; Krings, U.; Popper, L.; Berger, R.G. A chlorogenic acid esterase with a unique substrate specificity from Ustilago maydis. Appl. Environ. Microbiol. 2015, 81, 1679–1688. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.; Zhang, R.; Ma, Z.; Wang, H.; Ng, T. A feruloyl esterase (FAE) characterized by relatively high thermostability from the edible mushroom Russula virescens. Appl. Biochem. Biotechnol. 2014, 172, 993–1003. [Google Scholar] [CrossRef]
- Benoit, I.; Danchin, E.G.; Bleichrodt, R.J.; de Vries, R.P. Biotechnological applications and potential of fungal feruloyl esterases based on prevalence, classification and biochemical diversity. Biotechnol. Lett. 2008, 30, 387–396. [Google Scholar] [CrossRef]
- Udatha, D.B.; Kouskoumvekaki, I.; Olsson, L.; Panagiotou, G. The interplay of descriptor-based computational analysis with pharmacophore modeling builds the basis for a novel classification scheme for feruloyl esterases. Biotechnol. Adv. 2011, 29, 94–110. [Google Scholar] [CrossRef] [Green Version]
- Wong, D.W.S.; Chan, V.J.; Liao, H. Metagenomic discovery of feruloyl esterases from rumen microflora. Appl. Microbiol. Biotechnol. 2019, 103, 8449–8457. [Google Scholar] [CrossRef]
- Uraji, M.; Tamura, H.; Mizohata, E.; Arima, J.; Wan, K.; Ogawa, K.; Inoue, T.; Hatanaka, T. Loop of Streptomyces Feruloyl Esterase Plays an Important Role in the Enzyme’s Catalyzing the Release of Ferulic Acid from Biomass. Appl. Environ. Microbiol. 2018, 84. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Esterases | Total Protein (mg) | Total Activity (U) | Specific Activity (U/g) |
|---|---|---|---|
| reFAE1 | 21.19 ± 0.03 b | 5.2238 ± 0.03 a | 246.37 ± 0.03 c |
| reFAE2 | 24.36 ± 0.08 b | 8.3055 ± 0.02 a | 340.95 ± 0.04 c |
| Substrate | Specific Activity (U/g) | |
|---|---|---|
| reFAE1 | reFAE2 | |
| MFA | 8.90 ± 0.01 a | 12.86 ± 0.28 b |
| MCA | 433.35 ± 0.68 a | 583.66 ± 1.03 b |
| MSA | 14.21 ± 0.08 b | 7.13 ± 0.05 a |
| MpCA | 258.97 ± 0.16 a | 294.58 ± 0.82 b |
| CGA | 246.37 ± 0.02 a | 340.95 ±0.04 b |
| Sample | L* | a* | b* | Appearance |
|---|---|---|---|---|
| A | 53.61 ± 1.32 a | −50.10 ± 0.66 a | −24.32 ± 0.27 a |  |
| B | 87.75 ± 1.24 b | 20.62 ± 0.72 b | 32.14 ± 0.32 b |  |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, W.; Wang, J.; Liu, Y.; Leng, J. Heterologous Expression and Characterization of a Ferulic Acid Esterase from Aspergillus aculeatus with Potential Use in Sunflower Seed Processing. Appl. Sci. 2021, 11, 4453. https://doi.org/10.3390/app11104453
Zhang W, Wang J, Liu Y, Leng J. Heterologous Expression and Characterization of a Ferulic Acid Esterase from Aspergillus aculeatus with Potential Use in Sunflower Seed Processing. Applied Sciences. 2021; 11(10):4453. https://doi.org/10.3390/app11104453
Chicago/Turabian StyleZhang, Wenbin, Jieyu Wang, Yuechen Liu, and Juncai Leng. 2021. "Heterologous Expression and Characterization of a Ferulic Acid Esterase from Aspergillus aculeatus with Potential Use in Sunflower Seed Processing" Applied Sciences 11, no. 10: 4453. https://doi.org/10.3390/app11104453
APA StyleZhang, W., Wang, J., Liu, Y., & Leng, J. (2021). Heterologous Expression and Characterization of a Ferulic Acid Esterase from Aspergillus aculeatus with Potential Use in Sunflower Seed Processing. Applied Sciences, 11(10), 4453. https://doi.org/10.3390/app11104453