
The Long-Term Consequences of Forest Fires on the Carbon Fluxes of a Tropical Forest in Africa


Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Region Mt. Kilimanjaro
2.2. The Forest Model FORMIND
2.3. Review of Fire Models
2.3.1. Fire Model by Drossel and Schwabl
2.3.2. Fire Model by Green
2.3.3. Fire Model by Busing
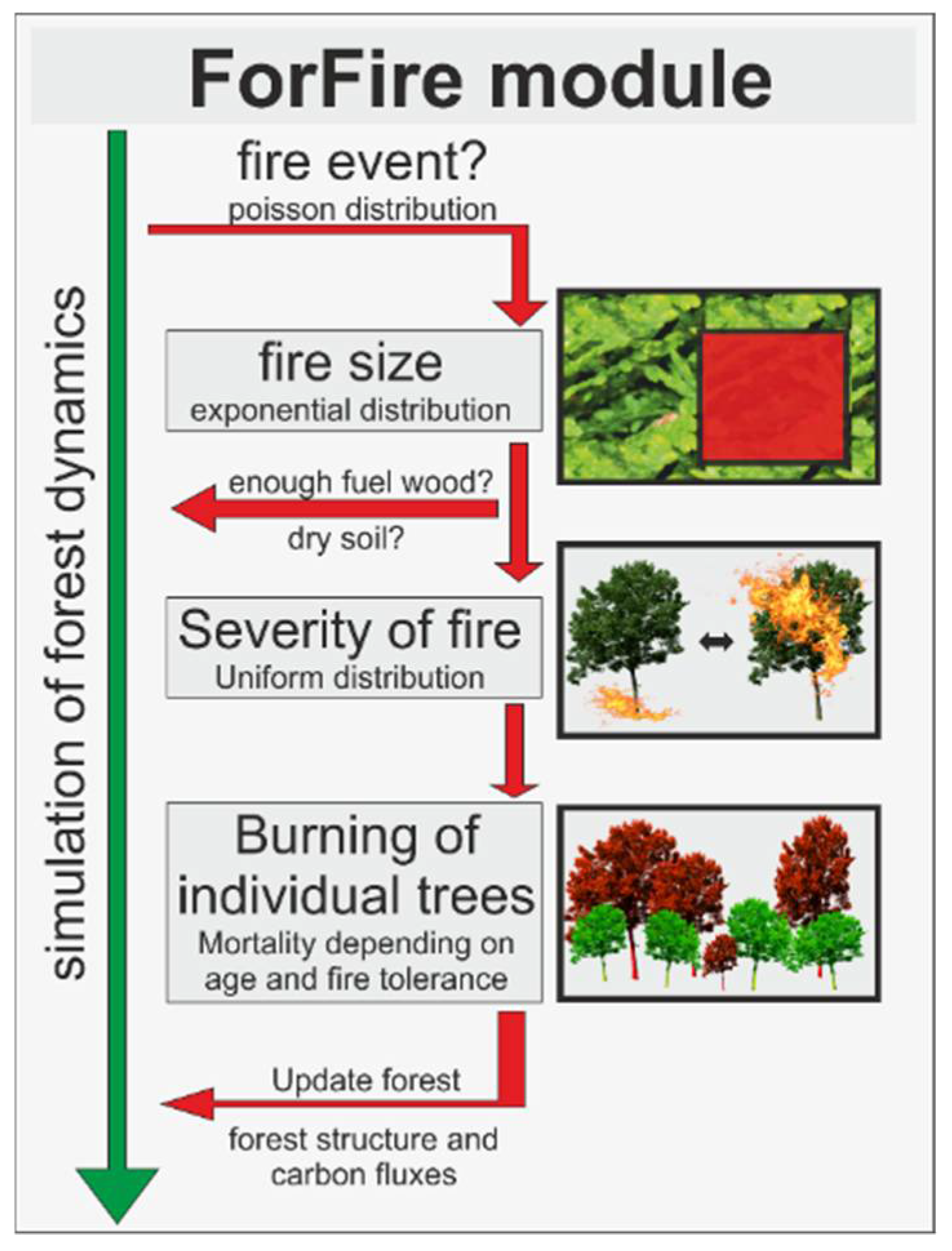
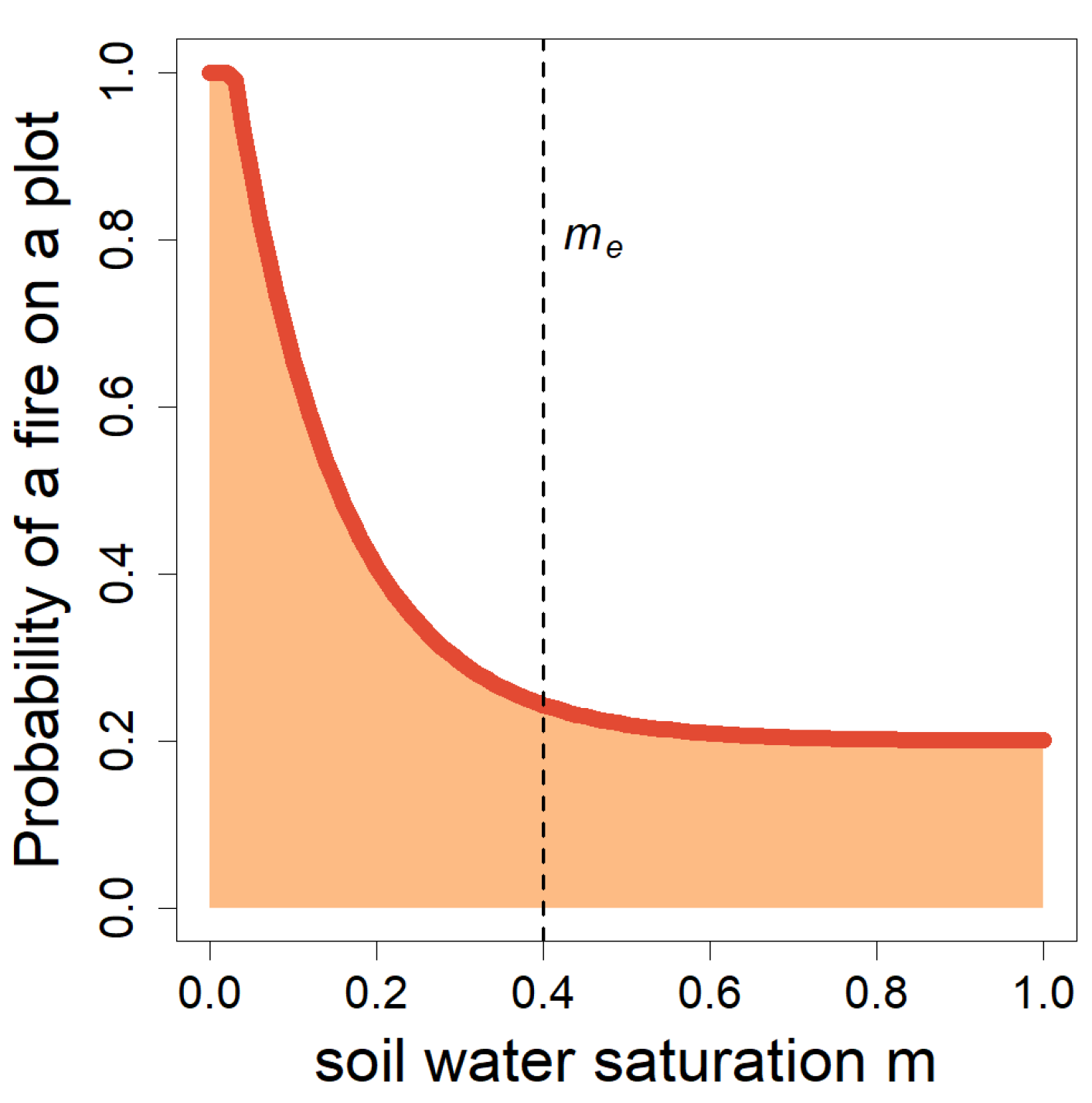
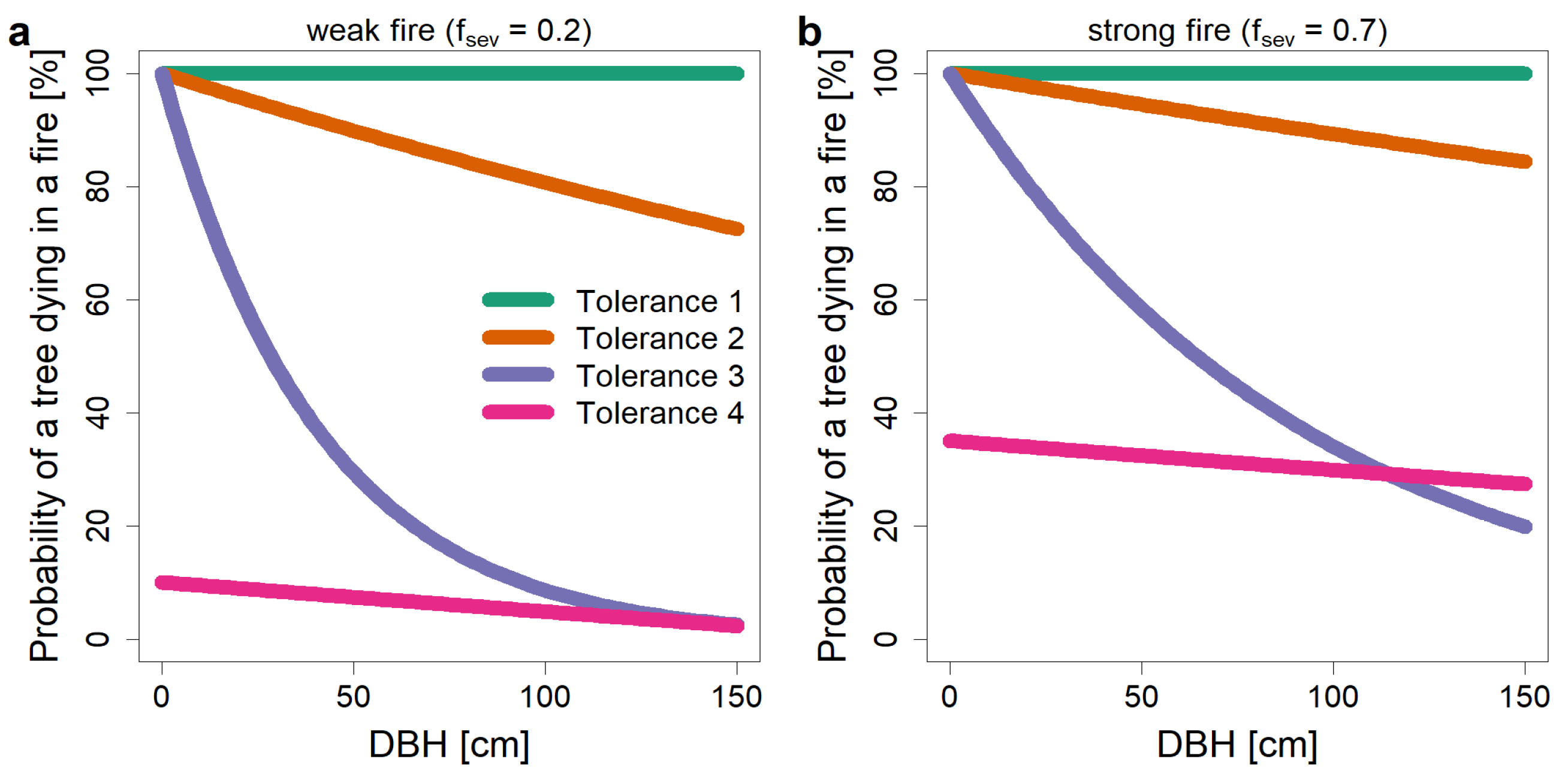
2.4. The New Fire Model ForFire
2.5. Simulation Settings
3. Results
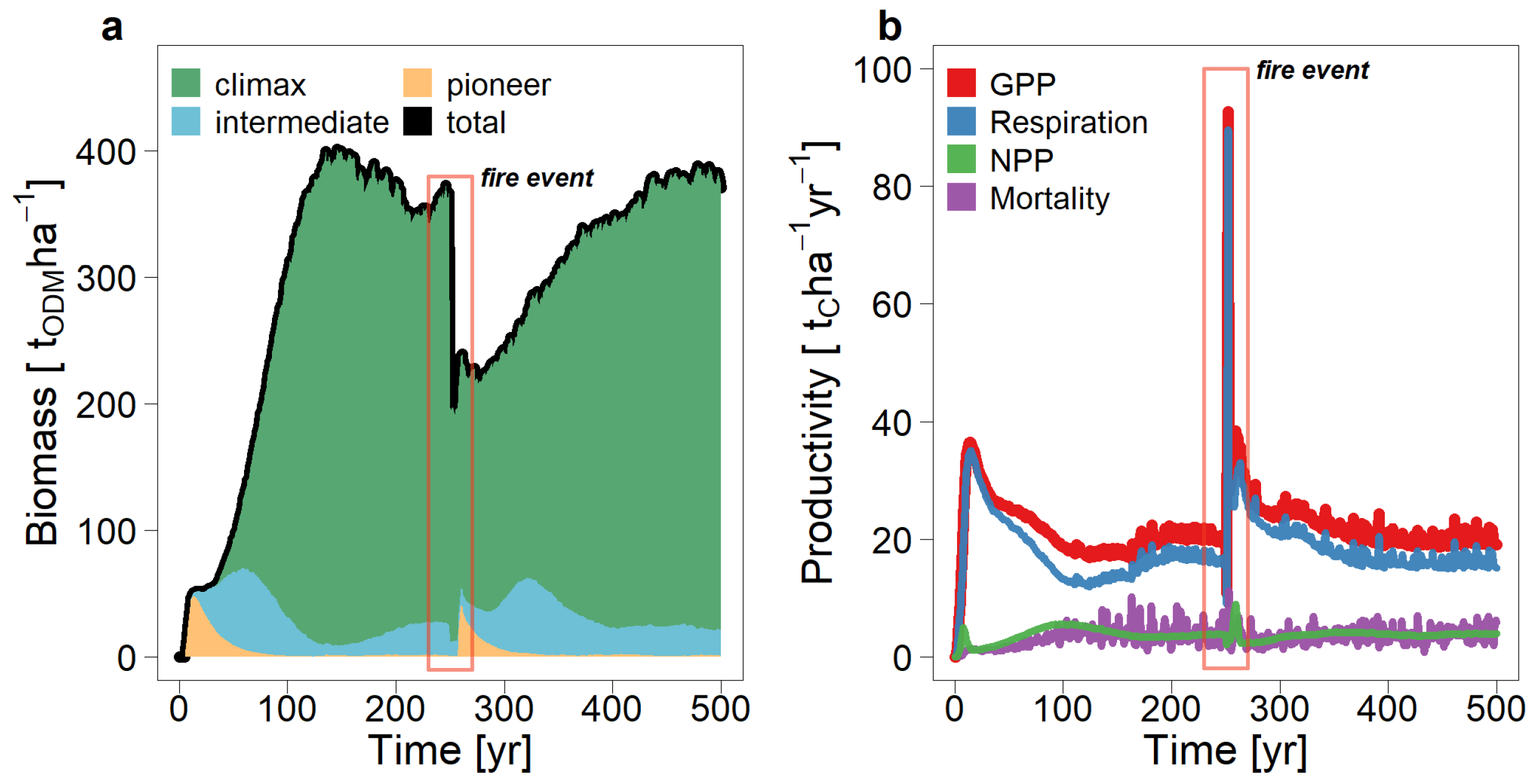
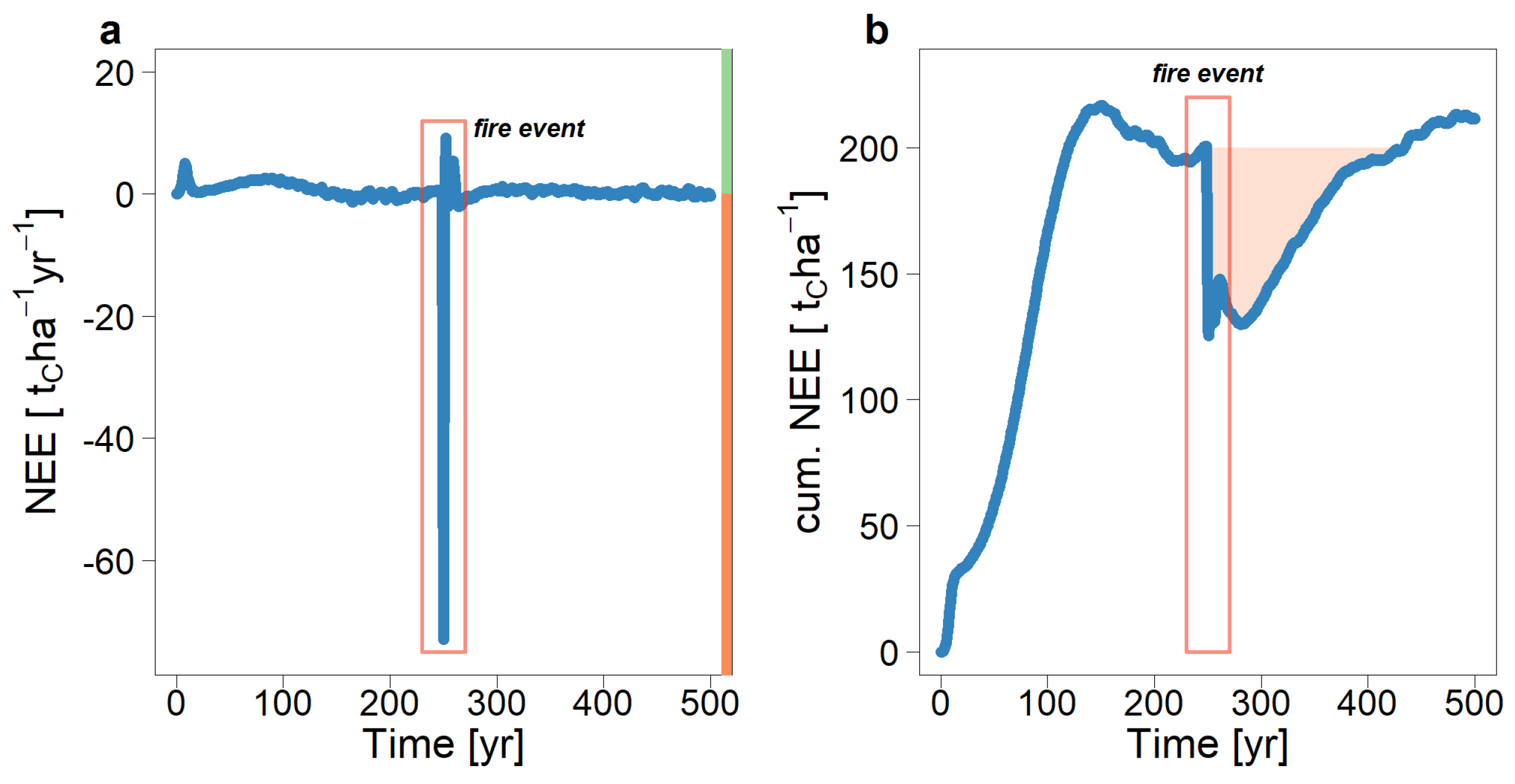
3.1. The Consequences of a Fire Event on the Carbon Fluxes in a Tropical Forest
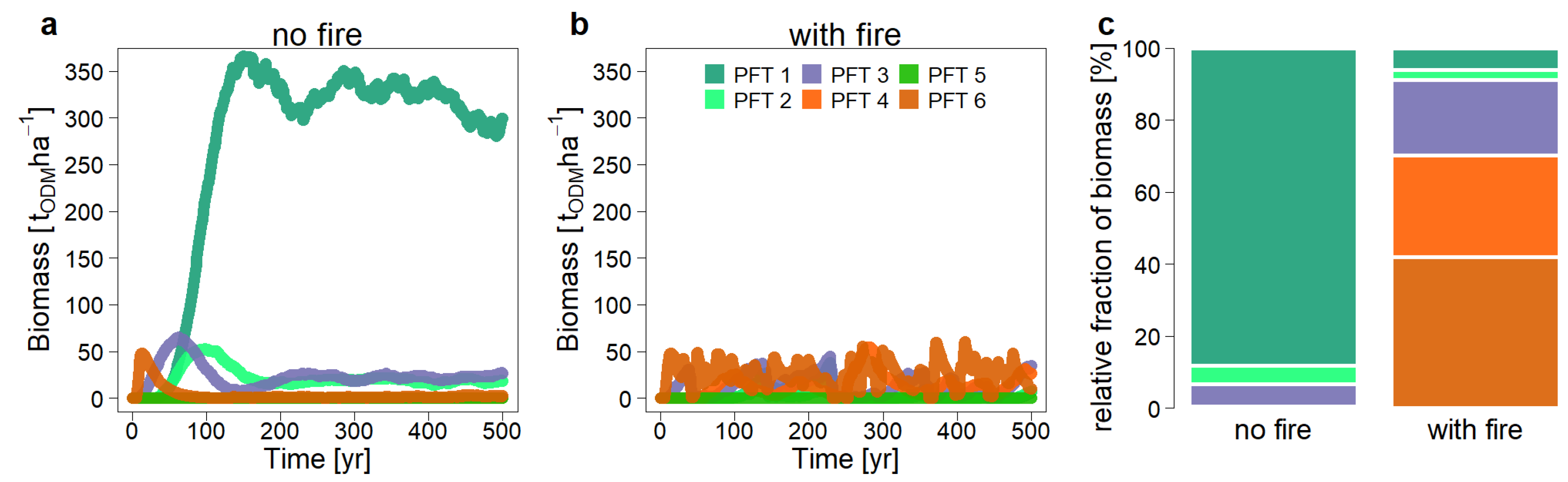
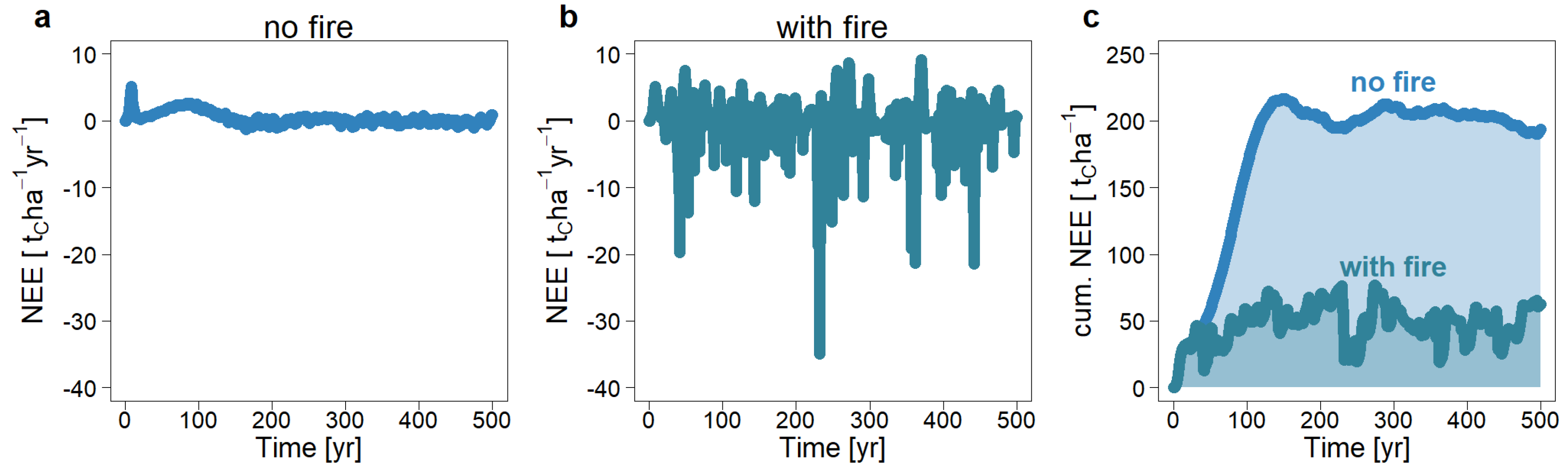
3.2. The Influence of Recurrent Fire Events on Forest Dynamics
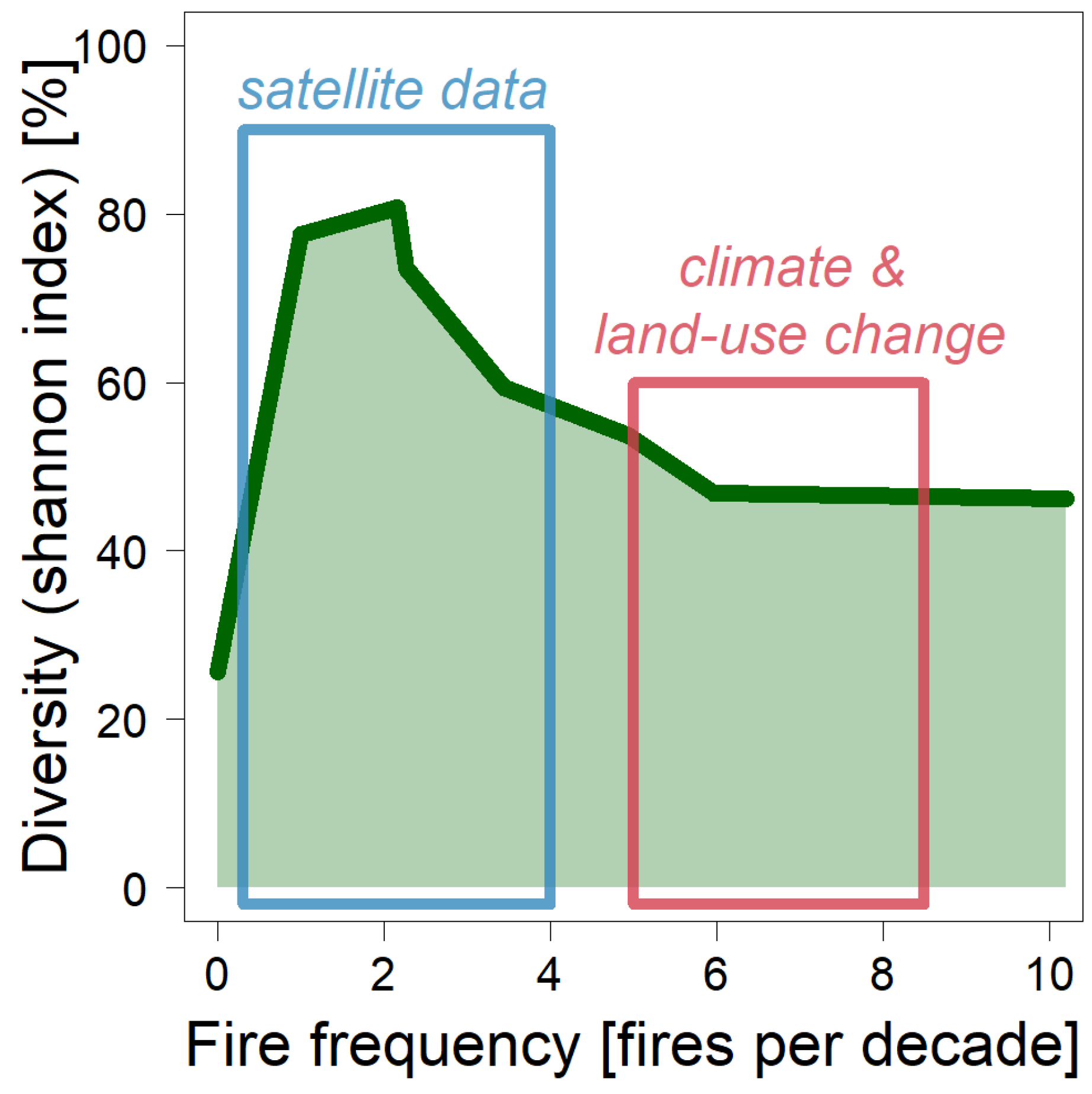
4. Discussion
5. Conclusions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A
Species List of the Study Area at Mt. Kilimanjaro
| Species | Max. Height (m) | Shade Tolerance | PFT | Fire Tolerance |
| Agarista salicifolia | 20 | intol | 4 | 4 |
| Alangium chinense | 25 | intol | 4 | 1 |
| Albizia gummifera | 30 | tol | 2 | 1 |
| Aningeria adolfi friedericii | 50 | tol | 1 | 1 |
| Aphloia theiformis | 15 | med | 3 | 1 |
| Bersama abyssinica | 20 | med | 3 | 1 |
| Canthium oligocarpum ssp captum | 10 | tol | 5 | 1 |
| Casearia battiscombei | 30 | tol | 1 | 1 |
| Celtis durandii | 25 | med | 3 | 1 |
| Clausena anisata | 20 | med | 3 | 1 |
| Cornus volkensii | 30 | med | 3 | 1 |
| Cyathea manniana | 15 | intol | 6 | 1 |
| Dracaena afromontana | 10 | tol | 5 | 1 |
| Dracaena laxissima | 5 | tol | 5 | 1 |
| Eckebergia capensis | 25 | tol | 2 | 1 |
| Embelia schimperi | 30 | tol | 2 | 1 |
| Entandrophragma excelsum | 70 | tol | 1 | 1 |
| Erica excelsa | 28 | intol | 4 | 4 |
| Erythrococca polyandra | 10 | tol | 5 | 1 |
| Ficus sur | 25 | med | 3 | 1 |
| Galiniera saxifraga | 15 | tol | 5 | 1 |
| Garcinia tansaniensis | 40 | tol | 1 | 1 |
| Garcinia volkensii | 20 | tol | 2 | 1 |
| Hagenia abyssinica | 25 | intol | 4 | 3 |
| Hallea rubrostipulata | 33 | med | 3 | 1 |
| Heinsenia diervilleoides | 25 | tol | 2 | 1 |
| Ilex mitis | 30 | tol | 2 | 1 |
| Lasianthus kilinandscharicus | 10 | tol | 5 | 1 |
| Lepidotrichilia volkensii | 16 | med | 3 | 1 |
| Leptonychia usambarensis | 15 | tol | 5 | 1 |
| Macaranga capensis var kilimandscharica | 30 | med | 3 | 1 |
| Maesa lanceolata | 20 | intol | 4 | 1 |
| Maytenus acuminata | 15 | tol | 5 | 1 |
| Morella salicifolia | 15 | intol | 4 | 4 |
| Myrica salicifolia | 15 | intol | 4 | 4 |
| Newtonia buchananii | 40 | tol | 1 | 1 |
| Ocotea usambarensis | 45 | tol | 1 | 2 |
| Olinia rochetiana | 25 | med | 3 | 1 |
| Pauridiantha paucinervis | 10 | tol | 5 | 1 |
| Pavetta abyssinica | 15 | tol | 5 | 1 |
| Peddiea fischeri | 15 | tol | 5 | 1 |
| Pittosporum spec lanatum | 10 | med | 3 | 1 |
| Podocarpus latifolius | 35 | tol | 1 | 1 |
| Polyscias fulva | 25 | intol | 4 | 1 |
| Polyscias albersiana | 15 | intol | 4 | 1 |
| Psychotria cyathicalyx | 15 | tol | 5 | 1 |
| Rapanea melanophloeos | 30 | med | 3 | 1 |
| Rawsonia lucida | 25 | tol | 2 | 1 |
| Rothmannia urcelliformis | 10 | tol | 2 | 1 |
| Schefflera myriantha | 30 | tol | 2 | 1 |
| Schefflera volkensii | 25 | tol | 2 | 1 |
| Strombosia scheffleri | 35 | tol | 1 | 1 |
| Syzygium guineense | 30 | tol | 2 | 1 |
| Tabernaemontana stapfiana | 25 | med | 3 | 1 |
| Teclea nobilis | 20 | tol | 2 | 1 |
| Xymalos monospora | 25 | tol | 2 | 1 |
References
- Ciais, P.; Sabine, C.; Bala, G.; Bopp, L.; Brovkin, V.; Canadell, J.; Chhabra, A.; DeFries, R.; Galloway, J.; Heimann, M.; et al. Carbon and other biogeochemical cycles. In Climate Change 2013: The Physical Science Basis. Contribution of Working Group I to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change; Stocker, T.F., Qin, D., Plattner, G.-K., Tignor, M., Allen, S.K., Boschung, I., Nauels, A., Xia, Y., Bex, V., Midgley, P.M., Eds.; Cambridge University Press: Cambridge, UK; New York, NY, USA, 2013; pp. 465–570. [Google Scholar] [CrossRef] [Green Version]
- Friedlingstein, P.; O’Sullivan, M.; Jones, M.W.; Andrew, R.M.; Hauck, J.; Olsen, A.; Peters, G.P.; Peters, W.; Pongratz, J.; Sitch, S.; et al. Global Carbon Budget 2020. Earth Syst. Sci. Data 2020, 12, 3269–3340. [Google Scholar] [CrossRef]
- Harris, N.L.; Gibbs, D.A.; Baccini, A.; Birdsey, R.A.; de Bruin, S.; Farina, M.; Fatoyinbo, L.; Hansen, M.C.; Herold, M.; Houghton, R.A.; et al. Global maps of twenty-first century forest carbon fluxes. Nat. Clim. Chang. 2021, 11, 234–240. [Google Scholar] [CrossRef]
- Houghton, R.A.; Hall, F.; Goetz, S.J. Importance of biomass in the global carbon cycle. J. Geophys. Res. Biogeosci. 2009, 114. [Google Scholar] [CrossRef]
- Pan, Y.D.; Birdsey, R.A.; Fang, J.Y.; Houghton, R.; Kauppi, P.E.; Kurz, W.A.; Phillips, O.L.; Shvidenko, A.; Lewis, S.L.; Canadell, J.G.; et al. A large and persistent carbon sink in the world’s forests. Science 2011, 333, 988–993. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pugh, T.A.M.; Arneth, A.; Kautz, M.; Poulter, B.; Smith, B. Important role of forest disturbances in the global biomass turnover and carbon sinks. Nat. Geosci. 2019, 12, 730–735. [Google Scholar] [CrossRef]
- Cochrane, M.A.; Alencar, A.; Schulze, M.D.; Souza, C.M.; Nepstad, D.C.; Lefebvre, P.; Davidson, E.A. Positive feedbacks in the fire dynamic of closed canopy tropical forests. Science 1999, 284, 1832–1835. [Google Scholar] [CrossRef] [PubMed]
- Nepstad, D.C.; Verissimo, A.; Alencar, A.; Nobre, C.; Lima, E.; Lefebvre, P.; Schlesinger, P.; Potter, C.; Moutinho, P.; Mendoza, E.; et al. Large-scale impoverishment of Amazonian forests by logging and fire. Nature 1999, 398, 505–508. [Google Scholar] [CrossRef]
- Andela, N.; Morton, D.C.; Giglio, L.; Chen, Y.; van der Werf, G.R.; Kasibhatla, P.S.; DeFries, R.S.; Collatz, G.J.; Hantson, S.; Kloster, S.; et al. A human-driven decline in global burned area. Science 2017, 356, 1356–1362. [Google Scholar] [CrossRef] [Green Version]
- Hansen, M.C.; Potapov, P.V.; Moore, R.; Hancher, M.; Turubanova, S.A.; Tyukavina, A.; Thau, D.; Stehman, S.V.; Goetz, S.J.; Loveland, T.R.; et al. High-resolution global maps of 21st-century forest cover change. Science 2013, 342, 850–853. [Google Scholar] [CrossRef] [Green Version]
- Brando, P.M.; Silverio, D.; Maracahipes-Santos, L.; Oliveira-Santos, C.; Levick, S.R.; Coe, M.T.; Migliavacca, M.; Balch, J.K.; Macedo, M.N.; Nepstad, D.C.; et al. Prolonged tropical forest degradation due to compounding disturbances: Implications for CO2 and H2O fluxes. Glob. Chang. Biol. 2019, 25, 2855–2868. [Google Scholar] [CrossRef] [PubMed]
- Morton, D.C.; DeFries, R.S.; Randerson, J.T.; Giglio, L.; Schroeder, W.; van der Werf, G.R. Agricultural intensification increases deforestation fire activity in Amazonia. Glob. Chang. Biol. 2008, 14, 2262–2275. [Google Scholar] [CrossRef] [Green Version]
- van der Werf, G.R.; Randerson, J.T.; Giglio, L.; Collatz, G.J.; Mu, M.; Kasibhatla, P.S.; Morton, D.C.; DeFries, R.S.; Jin, Y.; van Leeuwen, T.T. Global fire emissions and the contribution of deforestation, savanna, forest, agricultural, and peat fires (1997–2009). Atmos. Chem. Phys. 2010, 10, 11707–11735. [Google Scholar] [CrossRef] [Green Version]
- Cochrane, M.A. Fire science for rainforests. Nature 2003, 421, 913–919. [Google Scholar] [CrossRef] [PubMed]
- Hemp, A. The impact of fire on diversity, structure, and composition of the vegetation on Mt. Kilimanjaro. In Land Use Change and Mountain Biodiversity; Spehn, E.M., Liberman, M., Körner, C., Eds.; CRC Press: Boca Raton, FL, USA, 2006; pp. 51–68. [Google Scholar]
- Simon, M.; Plummer, S.; Fierens, F.; Hoelzemann, J.J.; Arino, O. Burnt area detection at global scale using ATSR-2: The GLOBSCAR products and their qualification. J. Geophys. Res. Atmos. 2004, 109, D14S02. [Google Scholar] [CrossRef]
- Hirschberger, P. Wälder in Flammen. Ursachen und Folgen der Weltweiten Waldbrände; WWF Deutschland: Berlin, Germany, 2012; p. 90. [Google Scholar]
- Hemp, A. Climate change-driven forest fires marginalize the impact of ice cap wasting on Kilimanjaro. Glob. Chang. Biol. 2005, 11, 1013–1023. [Google Scholar] [CrossRef]
- Whelan, R.J. The Ecology of Fire; Cambridge University Press: Cambridge, UK, 1995. [Google Scholar] [CrossRef]
- Axmacher, J.C.; Scheuermann, L.; Schrumpf, M.; Lyaruu, H.V.M.; Fiedler, K.; Müller-Hohenstein, K. Effects of fire on the diversity of geometrid moths on Mt. Kilimanjaro. In Land Use Change and Mountain Biodiversity; Spehn, E.M., Liberman, M., Körner, C., Eds.; CRC Press: Boca Raton, FL, USA, 2006; pp. 69–76. [Google Scholar]
- Thonicke, K.; Venevsky, S.; Sitch, S.; Cramer, W. The role of fire disturbance for global vegetation dynamics: Coupling fire into a Dynamic Global Vegetation Model. Glob. Ecol. Biogeogr. 2001, 10, 661–677. [Google Scholar] [CrossRef] [Green Version]
- Zinck, R.; Grimm, V. More realistic than anticipated: A classical forest-fire model from statistical physics captures real fire shapes. Open Ecol. J. 2008, 1, 8–13. [Google Scholar] [CrossRef] [Green Version]
- Gardner, R.H.; Romme, W.H.; Turner, M.G. Predicting forest fire effects at landscape scales. In Spatial Modeling of Forest LANDSCAPE Change. Approaches and Applications; Mladenoff, D.J., Baker, W.L., Eds.; Cambridge University Press: Cambridge, UK, 1999; pp. 163–185. [Google Scholar]
- Keane, R.E.; Cary, G.J.; Davies, I.D.; Flannigan, M.D.; Gardner, R.H.; Lavorel, S.; Lenihan, J.M.; Li, C.; Rupp, T.S. A classification of landscape fire succession models: Spatial simulations of fire and vegetation dynamics. Ecol. Model. 2004, 179, 3–27. [Google Scholar] [CrossRef]
- Bugmann, H. A review of forest gap models. Clim. Chang. 2001, 51, 259–305. [Google Scholar] [CrossRef]
- Shugart, H.H.; Wang, B.; Fischer, R.; Ma, J.; Fang, J.; Yan, X.; Huth, A.; Armstrong, A.H. Gap models and their individual-based relatives in the assessment of the consequences of global change. Environ. Res. Lett. 2018, 13, 033001. [Google Scholar] [CrossRef] [Green Version]
- Fischer, R.; Bohn, F.; de Paula, M.D.; Dislich, C.; Groeneveld, J.; Gutiérrez, A.G.; Kazmierczak, M.; Knapp, N.; Lehmann, S.; Paulick, S.; et al. Lessons learned from applying a forest gap model to understand ecosystem and carbon dynamics of complex tropical forests. Ecol. Model. 2016, 326, 124–133. [Google Scholar] [CrossRef] [Green Version]
- Köhler, P.; Huth, A. Simulating growth dynamics in a South-East Asian rainforest threatened by recruitment shortage and tree harvesting. Clim. Chang. 2004, 67, 95–117. [Google Scholar] [CrossRef]
- Hemp, A. Continuum or zonation? Altitudinal gradients in the forest vegetation of Mt. Kilimanjaro. Plant Ecol. 2006, 184, 27–42. [Google Scholar] [CrossRef]
- Peters, M.K.; Hemp, A.; Appelhans, T.; Behler, C.; Classen, A.T.; Detsch, F.; Ensslin, A.; Ferger, S.W.; Frederiksen, S.B.; Gebert, F.; et al. Predictors of elevational biodiversity gradients change from single taxa to the multi-taxa community level. Nat. Commun. 2016, 7, 13736. [Google Scholar] [CrossRef] [Green Version]
- Ensslin, A.; Rutten, G.; Pommer, U.; Zimmermann, R.; Hemp, A.; Fischer, M. Effects of elevation and land use on the biomass of trees, shrubs and herbs at Mount Kilimanjaro. Ecosphere 2015, 6, art45. [Google Scholar] [CrossRef] [Green Version]
- Fischer, R.; Ensslin, A.; Rutten, G.; Fischer, M.; Schellenberger Costa, D.; Kleyer, M.; Hemp, A.; Paulick, S.; Huth, A. Simulating carbon stocks and fluxes of an African tropical montane forest with an individual-based forest model. PLoS ONE 2015, 10, e0123300. [Google Scholar] [CrossRef] [Green Version]
- Fischer, R.; Rödig, E.; Huth, A. Consequences of a reduced number of plant functional types for the simulation of forest productivity. Forests 2018, 9, 460. [Google Scholar] [CrossRef] [Green Version]
- Paulick, S.; Dislich, C.; Homeier, J.; Fischer, R.; Huth, A. The carbon fluxes in different successional stages: Modelling the dynamics of tropical montane forests in South Ecuador. For. Ecosyst. 2017, 4. [Google Scholar] [CrossRef] [Green Version]
- Armstrong, A.H.; Huth, A.; Osmanoglu, B.; Sun, G.; Ranson, K.J.; Fischer, R. A multi-scaled analysis of forest structure using individual-based modeling in a costa rican rainforest. Ecol. Model. 2020, 433, 109226. [Google Scholar] [CrossRef]
- Hiltner, U.; Huth, A.; Hérault, B.; Holtmann, A.; Bräuning, A.; Fischer, R. Climate change alters the ability of neotropical forests to provide timber and sequester carbon. For. Ecol. Manag. 2021, 492, 119166. [Google Scholar] [CrossRef]
- Kammesheidt, L.; Köhler, P.; Huth, A. Sustainable timber harvesting in Venezuela: A modelling approach. J. Appl. Ecol. 2001, 38, 756–770. [Google Scholar] [CrossRef] [Green Version]
- Huth, A.; Drechsler, M.; Köhler, P. Multicriteria evaluation of simulated logging scenarios in a tropical rain forest. J. Environ. Manag. 2004, 71, 321–333. [Google Scholar] [CrossRef] [PubMed]
- Drossel, B.; Schwabl, F. Self-organized critical forest-fire model. Phys. Rev. Lett. 1992, 69, 1629–1632. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Green, D.G. Simulated effects of fire, dispersal and spatial pattern on competition within forest mosaics. Vegetatio 1989, 82, 139–153. [Google Scholar] [CrossRef]
- Bugmann, H.K.M.; Solomon, A.M. The use of a European forest model in North America: A study of ecosystem response to climate gradients. J. Biogeogr. 1995, 22, 477–484. [Google Scholar] [CrossRef]
- Bugmann, H.K.M. A simplified forest model to study species composition along climate gradients. Ecology 1996, 77, 2055–2074. [Google Scholar] [CrossRef]
- Busing, R.T.; Solomon, A.M. Modeling the Effects of Fire Frequency and Severity on Forests in the Northwestern United States; Scientific Investigations Report 2006-5061; USGS: Reston, Virginia, 2007; p. 16.
- Busing, R.T.; Solomon, A.M.; McKane, R.B.; Burdick, C.A. Forest dynamics in oregon landscapes: Evaluation and application of an individual-based model. Ecol. Appl. 2007, 17, 1967–1981. [Google Scholar] [CrossRef]
- Schultz, J. Die Ökozonen der Erde-Die Ökologische Gliederung der Geosphäre, 1st ed.; Eugen Ulmer: Stuttgart, Germany, 1988; p. 488. [Google Scholar]
- Thonicke, K.; Spessa, A.; Prentice, I.C.; Harrison, S.P.; Dong, L.; Carmona-Moreno, C. The influence of vegetation, fire spread and fire behaviour on biomass burning and trace gas emissions: Results from a process-based model. Biogeosciences 2010, 7, 1991–2011. [Google Scholar] [CrossRef] [Green Version]
- Fischer, R.; Armstrong, A.; Shugart, H.H.; Huth, A. Simulating the impacts of reduced rainfall on carbon stocks and net ecosystem exchange in a tropical forest. Environ. Model. Softw. 2014, 52, 200–206. [Google Scholar] [CrossRef]
- Dale, V.H.; Hemstrom, M.A. CLIMACS: A Computer Model of Forest Stand Development for Western Oregon and Washington; PNW-327; United States Department of Agriculture: Washington, DC, USA, 1984. [Google Scholar]
- Archibald, S.; Roy, D.P. Identifying individual fires from satellite-derived burned area data. In Proceedings of the IEEE International Geoscience and Remote Sensing Symposium (IGARSS), Cape Town, South Africa, 12–17 July 2009; pp. 160–163. [Google Scholar]
- Alrutz, M. Auswirkungen von Feuer auf den Tropischen Regenwald am Kilimandscharo, Tansania. Eine Modell-Studie; Universität Leipzig, Fakultät für Physik und Geowissenschaften, Institut für Geographie: Leipzig, Germany, 2013. [Google Scholar]
- Goldammer, J.G. Feuer in Waldökosystemen der Tropen und Subtropen; Birkhäuser: Basel, Switzerland; Berlin, Germany, 1993. [Google Scholar]
- Bond, W.J.; van Wilgen, B.W. Fire and Plants; Chapman & Hall: London, UK, 1996; Volume 14. [Google Scholar]
- Hemp, A. Vegetation of Kilimanjaro: Hidden endemics and missing bamboo. Afr. J. Ecol. 2006, 44, 305–328. [Google Scholar] [CrossRef]
- Hemp, A. Climate change and its impact on the forests of Kilimanjaro. Afr. J. Ecol. 2009, 47, 3–10. [Google Scholar] [CrossRef]
- Tutul, E.; Uddin, M.; Rahman, M.; Hassan, M. Angiospermic flora of Runctia sal forest, Bangladesh. II. Magnoliopsida (Dicots). Bangladesh J. Plant Taxon. 2010, 17, 33–53. [Google Scholar] [CrossRef]
- Rasolonandrasana, B.P.N.; Goodman, S.M. The influence of fire on mountain sclerophyllous forests and their small-mammal communities in Madagascar. In Land Use Change and Mountain Biodiversity; Spehn, E.M., Liberman, M., Körner, C., Eds.; CRC Press/Taylor & Francis: Boca Raton, FL, USA, 2006. [Google Scholar] [CrossRef] [Green Version]
- Bowman, D.M.J.S.; Balch, J.K.; Artaxo, P.; Bond, W.J.; Carlson, J.M.; Cochrane, M.A.; D’Antonio, C.M.; DeFries, R.S.; Doyle, J.C.; Harrison, S.P.; et al. Fire in the Earth System. Science 2009, 324, 481–484. [Google Scholar] [CrossRef] [PubMed]
- Pfeiffer, M.; Spessa, A.; Kaplan, J.O. A model for global biomass burning in preindustrial time: LPJ-LMfire (v1.0). Geosci. Model Dev. 2013, 6, 643–685. [Google Scholar] [CrossRef] [Green Version]
- Andersen, A.N.; Cook, G.D.; Corbett, L.K.; Douglas, M.M.; Eager, R.W.; Russell-Smith, J.; Setterfield, S.A.; Williams, R.J.; Woinarski, J.C.Z. Fire frequency and biodiversity conservation in Australian tropical savannas: Implications from the Kapalga fire experiment. Austral Ecol. 2005, 30, 155–167. [Google Scholar] [CrossRef]
- Aragón, R.; Cristóbal, L.; Carilla, J. Fire, plant species richness, and aerial biomass distribution in mountain grasslands of northwest Argentina. In Land Use Change and Mountain Biodiversity; Spehn, E.M., Liberman, M., Körner, C., Eds.; CRC Press/Taylor & Francis: Boca Raton, FL, USA, 2006; pp. 89–99. [Google Scholar] [CrossRef] [Green Version]
- Vogl, R.J. Effects of fire on grasslands. In Fire and Ecosystems; Kozlowski, T.T., Ahlgren, C.E., Eds.; Academic Press: New York, NY, USA, 1974; pp. 139–194. [Google Scholar] [CrossRef]
- Cochrane, M.A.; Schulze, M.D. Fire as a recurrent event in tropical forests of the eastern Amazon: Effects on forest structure, biomass, and species composition. Biotropica 1999, 31, 2–16. [Google Scholar] [CrossRef]
- Kauffman, J.B.; Cummings, D.L.; Ward, D.E.; Babbitt, R. Fire in the Brazilian Amazon: 1. Biomass, nutrient pools, and losses in slashed primary forests. Oecologia 1995, 104, 397–408. [Google Scholar] [CrossRef] [PubMed]
- Oliveras, I.; Malhi, Y.; Salinas, N.; Huaman, V.; Urquiaga-Flores, E.; Kala-Mamani, J.; Quintano-Loaiza, J.A.; Cuba-Torres, I.; Lizarraga-Morales, N.; Roman-Cuesta, R.M. Changes in forest structure and composition after fire in tropical montane cloud forests near the Andean treeline. Plant Ecol. Divers. 2014, 7, 329–340. [Google Scholar] [CrossRef]
- Peterson, D.W.; Reich, P.B. Fire frequency and tree canopy structure influence plant species diversity in a forest-grassland ecotone. Plant Ecol. 2008, 194, 5–16. [Google Scholar] [CrossRef]
- Connell, J.H. Diversity in tropical rain forests and coral reefs. Science 1978, 199, 1302–1310. [Google Scholar] [CrossRef] [PubMed] [Green Version]














{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter | Description | Parameter Value for This Study |
|---|---|---|
| Fire frequency F | mean time between two fire events | 4 years |
| Fire size S | average size of a fire (here: 2 ha) in relation to the total size of the simulated forest (here: 9ha) | 20% |
| Fire severity fsev | indicator (0–1) for the strength and type of fire (light ground fire or strong crown fire) | 0.55 |
| PFT | Maximum Height [m] | Shade Tolerance | Exemplary Tree Species | Fire Tolerance |
|---|---|---|---|---|
| 1 | >33 | shade tolerant | Strombosia scheffleri | 2 |
| 2 | 16–33 | shade tolerant | Heinsenia diervilleoides | 1 |
| 3 | 16–33 | medium shade tolerant | Ficus sur | 1 |
| 4 | 16–33 | shade intolerant (pioneer) | Polyscias albersiana | 4 |
| 5 | <16 | shade tolerant | Leptonychia usambarensis | 2 |
| 6 | <16 | shade intolerant (pioneer) | Cyathea manniana | 2 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fischer, R. The Long-Term Consequences of Forest Fires on the Carbon Fluxes of a Tropical Forest in Africa. Appl. Sci. 2021, 11, 4696. https://doi.org/10.3390/app11104696
Fischer R. The Long-Term Consequences of Forest Fires on the Carbon Fluxes of a Tropical Forest in Africa. Applied Sciences. 2021; 11(10):4696. https://doi.org/10.3390/app11104696
Chicago/Turabian StyleFischer, Rico. 2021. "The Long-Term Consequences of Forest Fires on the Carbon Fluxes of a Tropical Forest in Africa" Applied Sciences 11, no. 10: 4696. https://doi.org/10.3390/app11104696
APA StyleFischer, R. (2021). The Long-Term Consequences of Forest Fires on the Carbon Fluxes of a Tropical Forest in Africa. Applied Sciences, 11(10), 4696. https://doi.org/10.3390/app11104696