
Anti-Aging Effects of Nanovesicles Derived from Human Tonsil-Derived Mesenchymal Stem Cells



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Isolation of Tonsil-Derived Mesenchymal Stem Cells
2.2. Production of Nanovesicles Derived from TMSCs
2.3. Western Blot
2.4. Cell Culture
2.5. Senescence-Associated β-Galactosidase Assay (SA-β-Galactosidase Assay)
2.6. Immunocytochemistry
2.7. Quantitative Real-Time Polymerase Chain Reaction (qPCR)
2.8. Statistical Analysis
3. Results
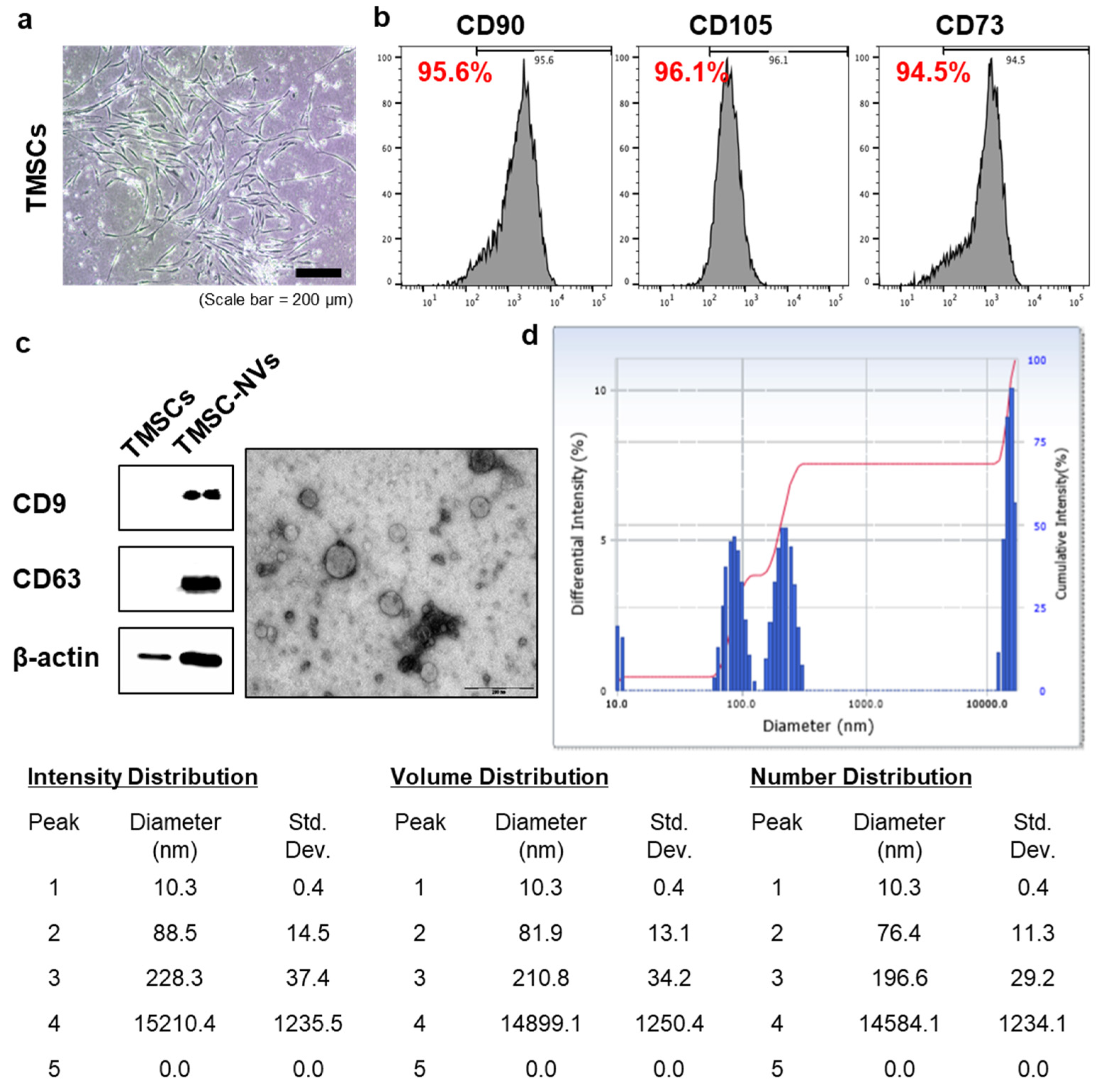
3.1. Production of Nanovesicles Derived from TMSC-NVs
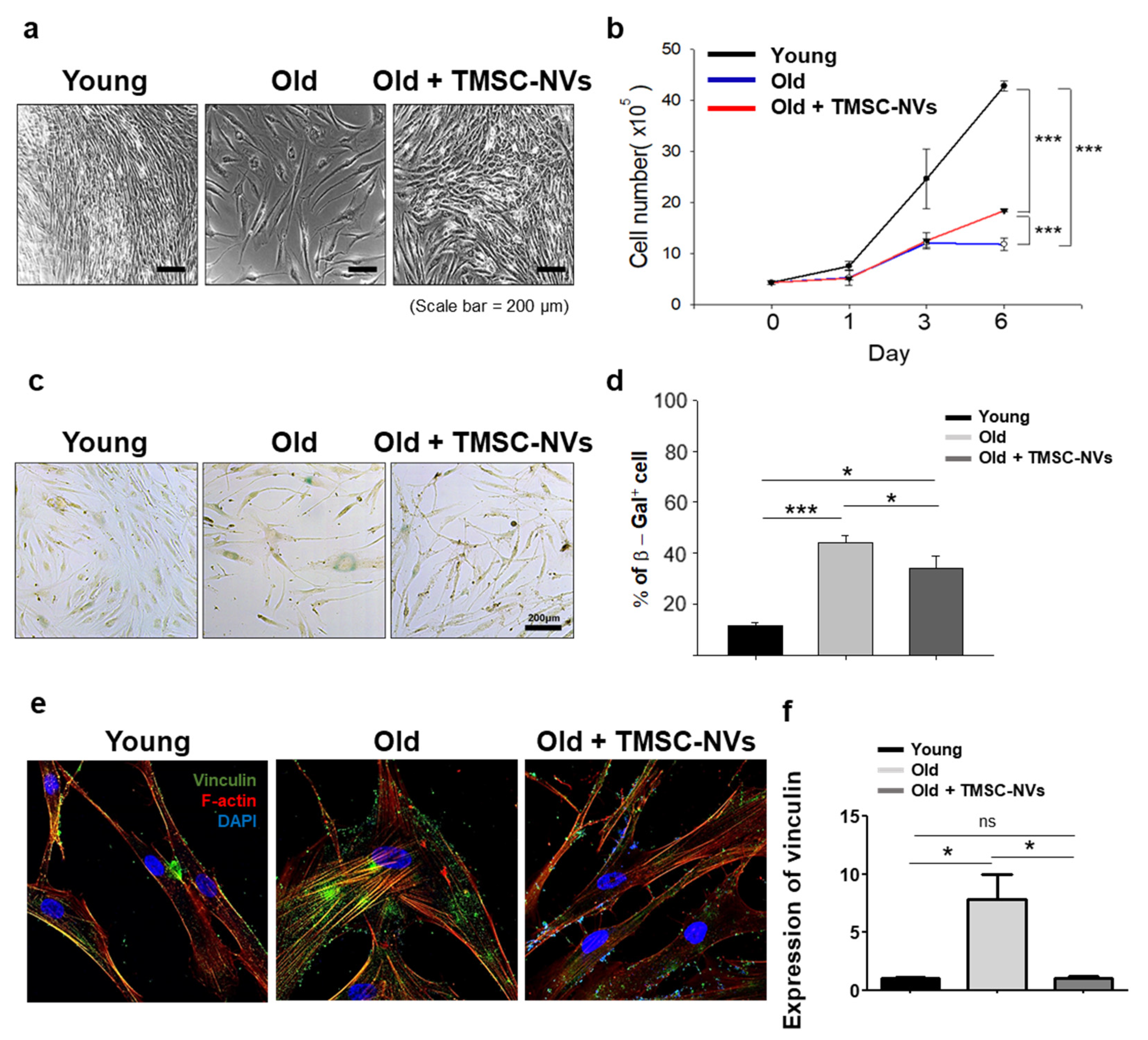
3.2. Anti-Aging Effect of TMSC-NVs in the Passage-Associated Senescence Model
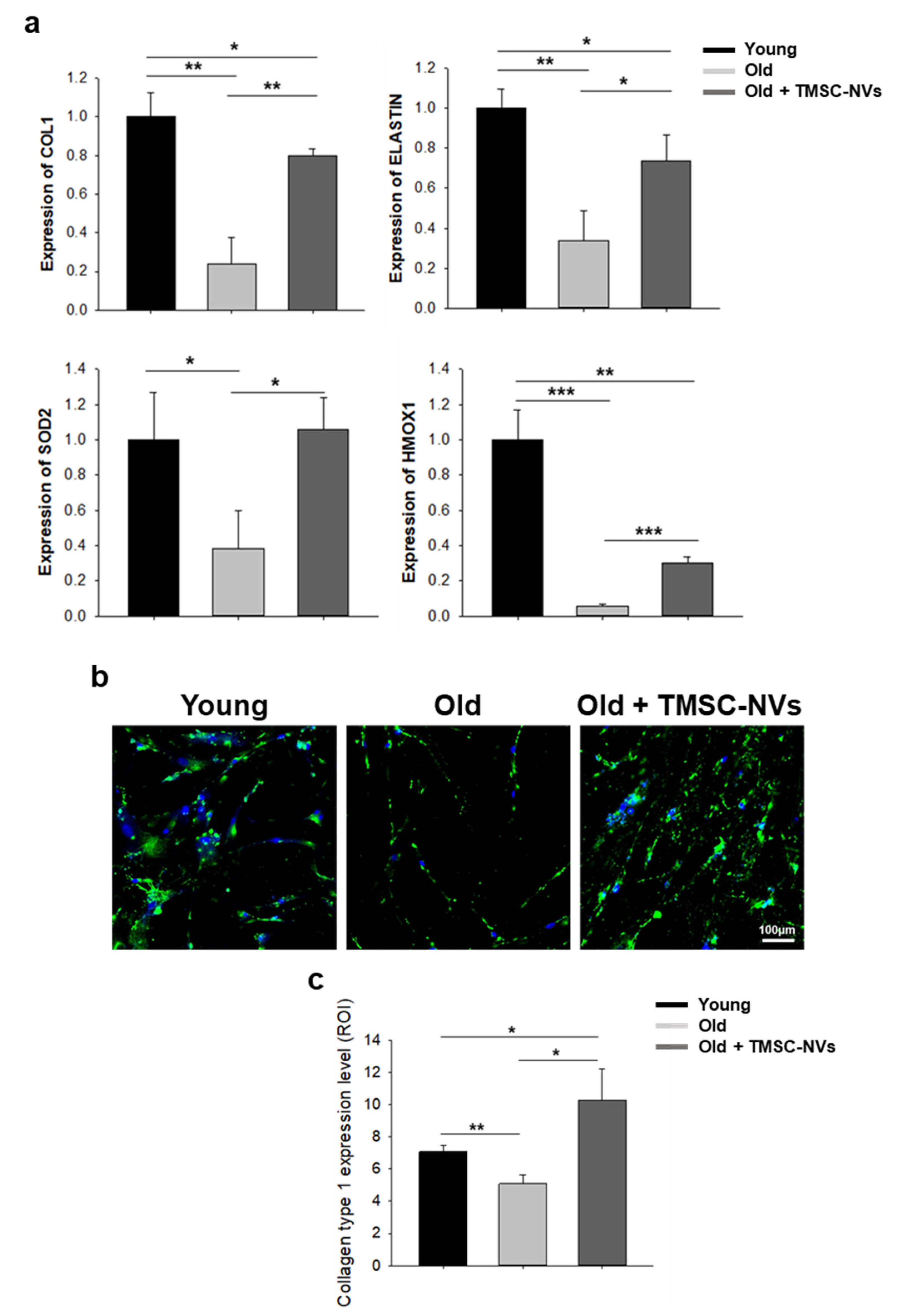
3.3. Regulation of Extracellular Matrix and Anti-Oxidant Gene by TMSC-NVs in the Passage-Associated Senescence Model
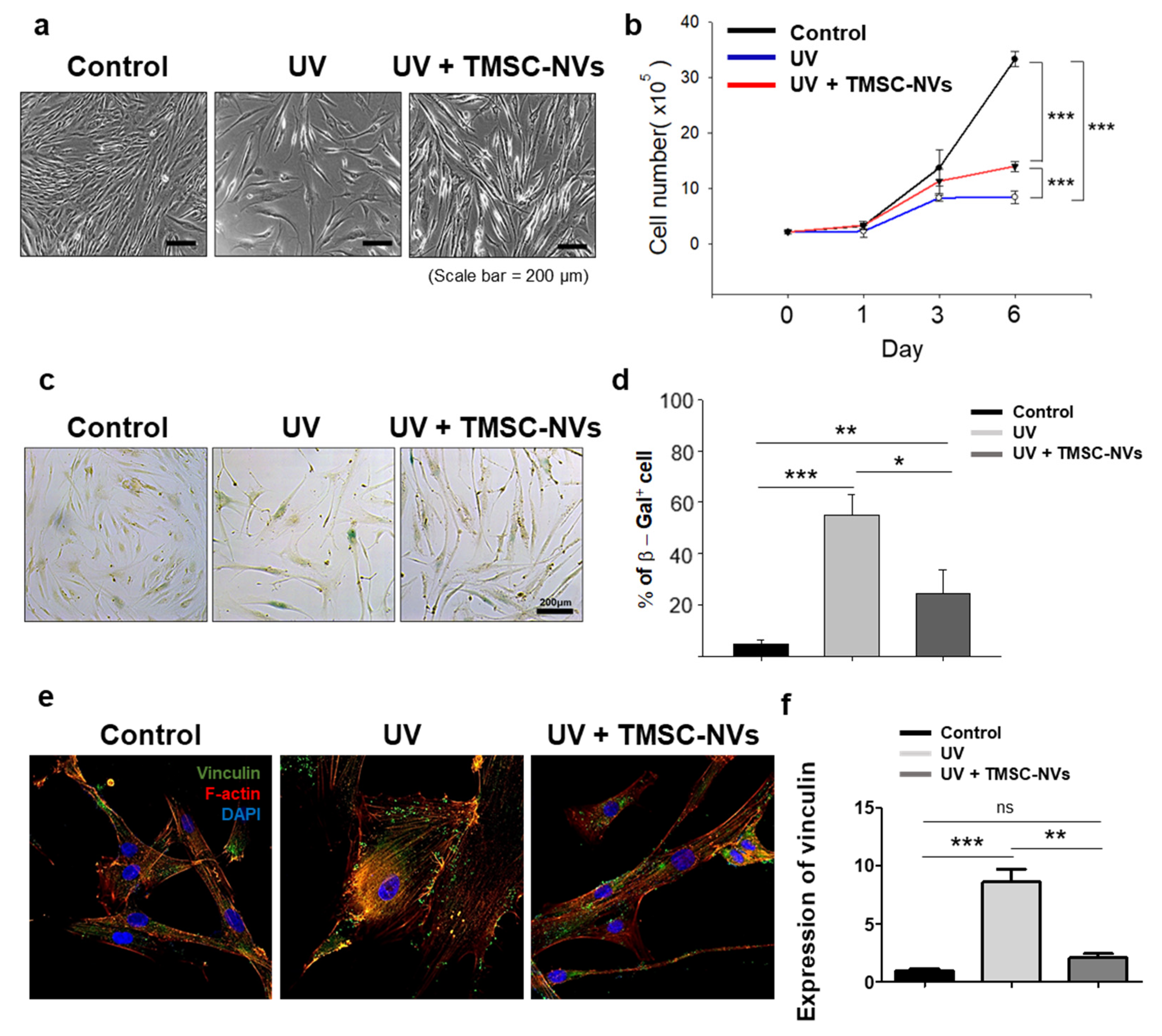
3.4. Anti-Aging Effect of TMSC-NV in the UV-Induced Senescence Model
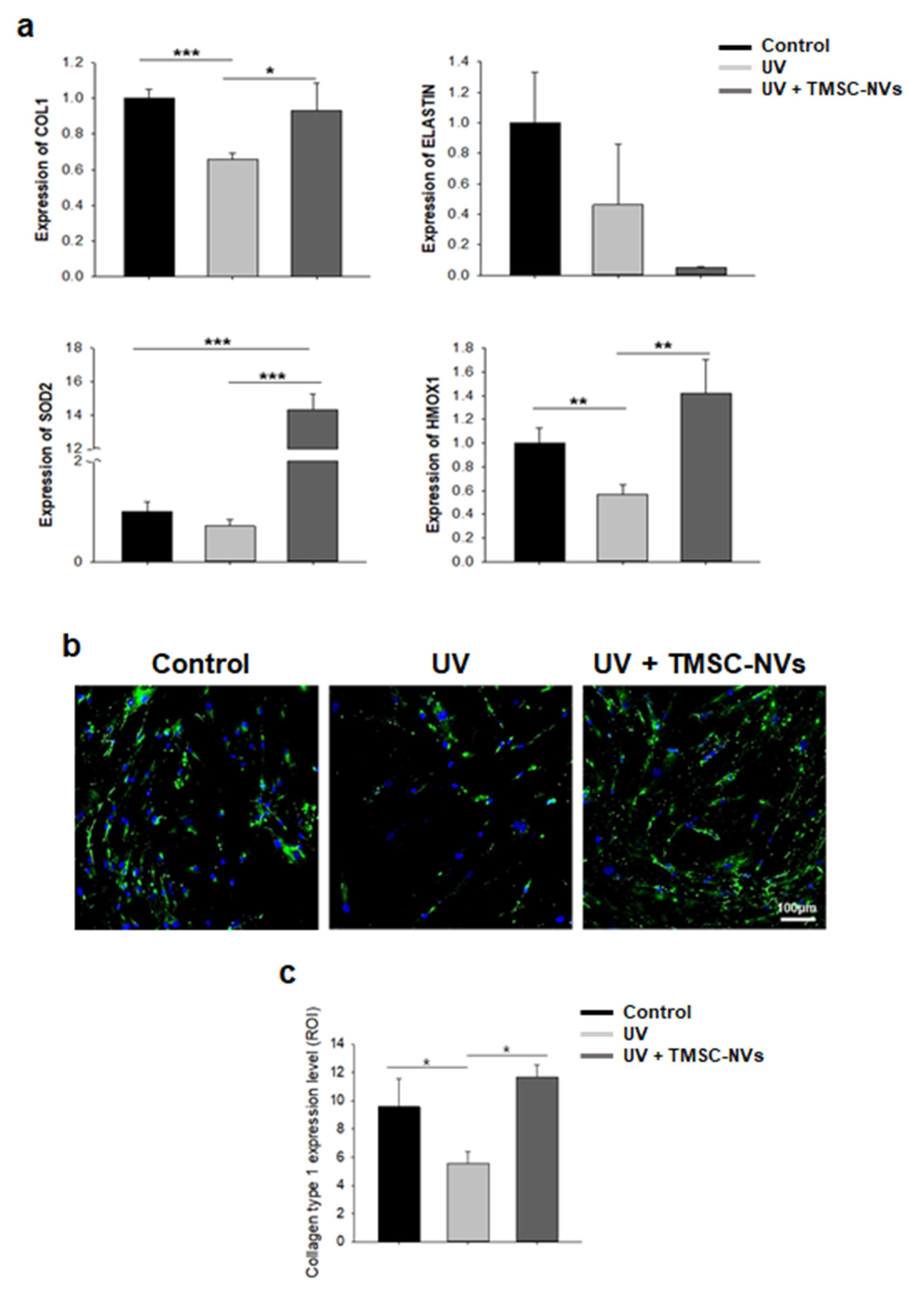
3.5. Regulation of Extracellular Matrix and Anti-Oxidant Gene by Treatment with TMSC-NV in the UV-Induced Senescence Model
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| DMEM | Dulbecco’s modified Eagle’s medium |
| ECM | Extracellular matrix |
| FBS | Fetal bovine serum |
| HDFs | Human dermal fibroblasts |
| PBS | Phosphate-buffered saline |
| SA-β-galactosidase | Senescence-associated beta galactosidase |
| TMSCs | Human tonsil-derived mesenchymal stem cells |
| TMSC-NVs | Nanovesicles derived from human tonsil-derived mesenchymal stem cells |
| UV | Ultraviolet B |
References
- Herranz, N.; Gil, J. Mechanisms and functions of cellular senescence. J. Clin. Investig. 2018, 128, 1238–1246. [Google Scholar] [CrossRef] [PubMed]
- Nishio, K.; Inoue, A. Senescence-associated alterations of cytoskeleton: Extraordinary production of vimentin that anchors cytoplasmic p53 in senescent human fibroblasts. Histochem. Cell Biol. 2005, 123, 263–273. [Google Scholar] [CrossRef] [PubMed]
- Moujaber, O.; Fishbein, F.; Omran, N.; Liang, Y.; Colmegna, I.; Presley, J.F.; Stochaj, U. Cellular senescence is associated with reorganization of the microtubule cytoskeleton. Cell. Mol. Life Sci. 2019, 76, 1169–1183. [Google Scholar] [CrossRef] [PubMed]
- Bu, H.; Wedel, S.; Cavinato, M.; Jansen-Dürr, P. MicroRNA regulation of oxidative stress-induced cellular senescence. Oxidative Med. Cell. Longev. 2017, 2017. [Google Scholar] [CrossRef]
- Chen, J.-H.; Ozanne, S.E.; Hales, C.N. Methods of cellular senescence induction using oxidative stress. In Biological Aging; Springer: Berlin/Heidelberg, Germany, 2007; pp. 179–189. [Google Scholar]
- Lee, S.; Jeong, S.-Y.; Lim, W.-C.; Kim, S.; Park, Y.-Y.; Sun, X.; Youle, R.J.; Cho, H. Mitochondrial fission and fusion mediators, hFis1 and OPA1, modulate cellular senescence. J. Biol. Chem. 2007, 282, 22977–22983. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vasileiou, P.V.; Evangelou, K.; Vlasis, K.; Fildisis, G.; Panayiotidis, M.I.; Chronopoulos, E.; Passias, P.-G.; Kouloukoussa, M.; Gorgoulis, V.G.; Havaki, S. Mitochondrial homeostasis and cellular senescence. Cells 2019, 8, 686. [Google Scholar] [CrossRef] [Green Version]
- Yang, Y.; Li, S. Dandelion extracts protect human skin fibroblasts from UVB damage and cellular senescence. Oxidative Med. Cell. Longev. 2015, 2015. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Helenius, M.; Mäkeläinen, L.; Salminen, A. Attenuation of NF-κB signaling response to UVB light during cellular senescence. Exp. Cell Res. 1999, 248, 194–202. [Google Scholar] [CrossRef]
- Hessvik, N.P.; Llorente, A. Current knowledge on exosome biogenesis and release. Cell Mol. Life Sci. 2018, 75, 193–208. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lai, R.C.; Yeo, R.W.Y.; Tan, K.H.; Lim, S.K. Exosomes for drug delivery—A novel application for the mesenchymal stem cell. Biotechnol. Adv. 2013, 31, 543–551. [Google Scholar] [CrossRef]
- Shao, H.; Chung, J.; Lee, K.; Balaj, L.; Min, C.; Carter, B.S.; Hochberg, F.H.; Breakefield, X.O.; Lee, H.; Weissleder, R. Chip-based analysis of exosomal mRNA mediating drug resistance in glioblastoma. Nat. Commun. 2015, 6, 1–9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Valadi, H.; Ekström, K.; Bossios, A.; Sjöstrand, M.; Lee, J.J.; Lötvall, J.O. Exosome-mediated transfer of mRNAs and microRNAs is a novel mechanism of genetic exchange between cells. Nat. Cell Biol. 2007, 9, 654–659. [Google Scholar] [CrossRef] [Green Version]
- Yuan, D.; Zhao, Y.; Banks, W.A.; Bullock, K.M.; Haney, M.; Batrakova, E.; Kabanov, A.V. Macrophage exosomes as natural nanocarriers for protein delivery to inflamed brain. Biomaterials 2017, 142, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Choi, E.W.; Seo, M.K.; Woo, E.Y.; Kim, S.H.; Park, E.J.; Kim, S. Exosomes from human adipose-derived stem cells promote proliferation and migration of skin fibroblasts. Exp. Derm. 2018, 27, 1170–1172. [Google Scholar] [CrossRef]
- Kim, S.; Lee, S.K.; Kim, H.; Kim, T.M. Exosomes Secreted from Induced Pluripotent Stem Cell-Derived Mesenchymal Stem Cells Accelerate Skin Cell Proliferation. Int. J. Mol. Sci. 2018, 19, 3119. [Google Scholar] [CrossRef] [Green Version]
- Zhang, B.; Wang, M.; Gong, A.; Zhang, X.; Wu, X.; Zhu, Y.; Shi, H.; Wu, L.; Zhu, W.; Qian, H.; et al. HucMSC-Exosome Mediated-Wnt4 Signaling Is Required for Cutaneous Wound Healing. Stem Cells 2015, 33, 2158–2168. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.J.; Yoo, S.M.; Park, H.H.; Lim, H.J.; Kim, Y.L.; Lee, S.; Seo, K.W.; Kang, K.S. Exosomes derived from human umbilical cord blood mesenchymal stem cells stimulates rejuvenation of human skin. Biochem. Biophys. Res. Commun. 2017, 493, 1102–1108. [Google Scholar] [CrossRef]
- Li, X.; Corbett, A.L.; Taatizadeh, E.; Tasnim, N.; Little, J.P.; Garnis, C.; Daugaard, M.; Guns, E.; Hoorfar, M.; Li, I.T.S. Challenges and opportunities in exosome research-Perspectives from biology, engineering, and cancer therapy. APL Bioeng. 2019, 3, 011503. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaneti, L.; Bronshtein, T.; Malkah Dayan, N.; Kovregina, I.; Letko Khait, N.; Lupu-Haber, Y.; Fliman, M.; Schoen, B.W.; Kaneti, G.; Machluf, M. Nanoghosts as a Novel Natural Nonviral Gene Delivery Platform Safely Targeting Multiple Cancers. Nano Lett. 2016, 16, 1574–1582. [Google Scholar] [CrossRef]
- Ou, Y.-H.; Zou, S.; Goh, W.J.; Wang, J.-W.; Wacker, M.; Czarny, B.; Pastorin, G. Cell-Derived Nanovesicles as Exosome-Mimetics for Drug Delivery Purposes: Uses and Recommendations. In Bio-Carrier Vectors; Springer: Berlin/Heidelberg, Germany, 2021; pp. 147–170. [Google Scholar]
- Oieni, J.; Lolli, A.; D’Atri, D.; Kops, N.; Yayon, A.; van Osch, G.J.; Machluf, M. Nano-ghosts: Novel biomimetic nano-vesicles for the delivery of antisense oligonucleotides. J. Control. Release 2021, 333, 28–40. [Google Scholar] [CrossRef]
- Yang, J.; Zhang, X.; Liu, C.; Wang, Z.; Deng, L.; Feng, C.; Tao, W.; Xu, X.; Cui, W. Biologically modified nanoparticles as theranostic bionanomaterials. Prog. Mater. Sci. 2021, 118, 100768. [Google Scholar] [CrossRef]
- Kumar, P.; Bose, P.P. Macrophage ghost entrapped amphotericin B: A novel delivery strategy towards experimental visceral leishmaniasis. Drug Deliv. Transl. Res. 2019, 9, 249–259. [Google Scholar] [CrossRef]
- Bose, R.J.; Lee, S.-H.; Park, H. Biofunctionalized nanoparticles: An emerging drug delivery platform for various disease treatments. Drug Discov. Today 2016, 21, 1303–1312. [Google Scholar] [CrossRef]
- Hwang, J.; Zheng, M.; Wiraja, C.; Cui, M.; Yang, L.; Xu, C. Reprogramming of macrophages with macrophage cell membrane-derived nanoghosts. Nanoscale Adv. 2020, 2, 5254–5262. [Google Scholar] [CrossRef]
- Bose, R.J.; Kim, B.J.; Arai, Y.; Han, I.-b.; Moon, J.J.; Paulmurugan, R.; Park, H.; Lee, S.-H. Bioengineered stem cell membrane functionalized nanocarriers for therapeutic targeting of severe hindlimb ischemia. Biomaterials 2018, 185, 360–370. [Google Scholar] [CrossRef] [PubMed]
- Krishnamurthy, S.; Gnanasammandhan, M.K.; Xie, C.; Huang, K.; Cui, M.Y.; Chan, J.M. Monocyte cell membrane-derived nanoghosts for targeted cancer therapy. Nanoscale 2016, 8, 6981–6985. [Google Scholar] [CrossRef] [Green Version]
- Toledano Furman, N.E.; Lupu-Haber, Y.; Bronshtein, T.; Kaneti, L.; Letko, N.; Weinstein, E.; Baruch, L.; Machluf, M. Reconstructed Stem Cell Nanoghosts: A Natural Tumor Targeting Platform. Nano Lett. 2013, 13, 3248–3255. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.; Lee, C.; Shin, Y.; Wang, S.; Han, J.; Kim, M.; Kim, J.M.; Shin, S.C.; Lee, B.J.; Kim, T.J.; et al. sEVs from tonsil-derived mesenchymal stromal cells alleviate activation of hepatic stellate cells and liver fibrosis through miR-486-5p. Mol. Ther. 2021, 29, 1471–1486. [Google Scholar] [CrossRef]
- Park, D.J.; Park, J.E.; Kong, T.H.; Seo, Y.A.-O. Alteration of payload in extracellular vesicles by crosstalk with mesenchymal stem cells from different origin. J. Nanobiotechnol. 2021, 19, 1–20. [Google Scholar] [CrossRef]
- Kurz, D.J.; Decary, S.; Hong, Y.; Erusalimsky, J.D. Senescence-associated (beta)-galactosidase reflects an increase in lysosomal mass during replicative ageing of human endothelial cells. J. Cell Sci. 2000. [Google Scholar] [CrossRef]
- Yoon, J.K.; Kim, D.H.; Kang, M.L.; Jang, H.K.; Park, H.J.; Lee, J.B.; Yi, S.W.; Kim, H.S.; Baek, S.; Park, D.B.; et al. Anti-Atherogenic Effect of Stem Cell Nanovesicles Targeting Disturbed Flow Sites. Small 2020, 16, e2000012. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ying, W.; Riopel, M.; Bandyopadhyay, G.; Dong, Y.; Birmingham, A.; Seo, J.B.; Ofrecio, J.M.; Wollam, J.; Hernandez-Carretero, A.; Fu, W.; et al. Adipose Tissue Macrophage-Derived Exosomal miRNAs Can Modulate In Vivo and In Vitro Insulin Sensitivity. Cell 2017, 171, 372–384.e312. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xin, H.; Li, Y.; Liu, Z.; Wang, X.; Shang, X.; Cui, Y.; Zhang, Z.G.; Chopp, M. MiR-133b Promotes Neural Plasticity and Functional Recovery After Treatment of Stroke with Multipotent Mesenchymal Stromal Cells in Rats Via Transfer of Exosome-Enriched Extracellular Particles. Stem Cells 2013, 31, 2737–2746. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lafferty-Whyte, K.; Cairney, C.J.; Jamieson, N.B.; Oien, K.A.; Keith, W.N. Pathway analysis of senescence-associated miRNA targets reveals common processes to different senescence induction mechanisms. Biochim. Biophys. Acta 2009, 1792, 341–352. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pascal, T.; Debacq-Chainiaux, F.; Chretien, A.; Bastin, C.; Dabee, A.F.; Bertholet, V.; Remacle, J.; Toussaint, O. Comparison of replicative senescence and stress-induced premature senescence combining differential display and low-density DNA arrays. FEBS Lett. 2005, 579, 3651–3659. [Google Scholar] [CrossRef] [PubMed] [Green Version]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, D.; Lee, Y.; Park, K.; Park, D.; Lee, W.J.; Roh, T.S.; Cho, H.; Baek, W. Anti-Aging Effects of Nanovesicles Derived from Human Tonsil-Derived Mesenchymal Stem Cells. Appl. Sci. 2021, 11, 5780. https://doi.org/10.3390/app11135780
Kim D, Lee Y, Park K, Park D, Lee WJ, Roh TS, Cho H, Baek W. Anti-Aging Effects of Nanovesicles Derived from Human Tonsil-Derived Mesenchymal Stem Cells. Applied Sciences. 2021; 11(13):5780. https://doi.org/10.3390/app11135780
Chicago/Turabian StyleKim, Dohyun, Youngdae Lee, Kwangsook Park, Danbi Park, Won Jai Lee, Tai Suk Roh, Hyungju Cho, and Wooyeol Baek. 2021. "Anti-Aging Effects of Nanovesicles Derived from Human Tonsil-Derived Mesenchymal Stem Cells" Applied Sciences 11, no. 13: 5780. https://doi.org/10.3390/app11135780
APA StyleKim, D., Lee, Y., Park, K., Park, D., Lee, W. J., Roh, T. S., Cho, H., & Baek, W. (2021). Anti-Aging Effects of Nanovesicles Derived from Human Tonsil-Derived Mesenchymal Stem Cells. Applied Sciences, 11(13), 5780. https://doi.org/10.3390/app11135780