
Diversity and Seasonal Dynamics of Airborne Fungi in Nerja Cave, Spain

 ,
, 
 , and
, and 
Abstract
:1. Introduction
2. Materials and Methods
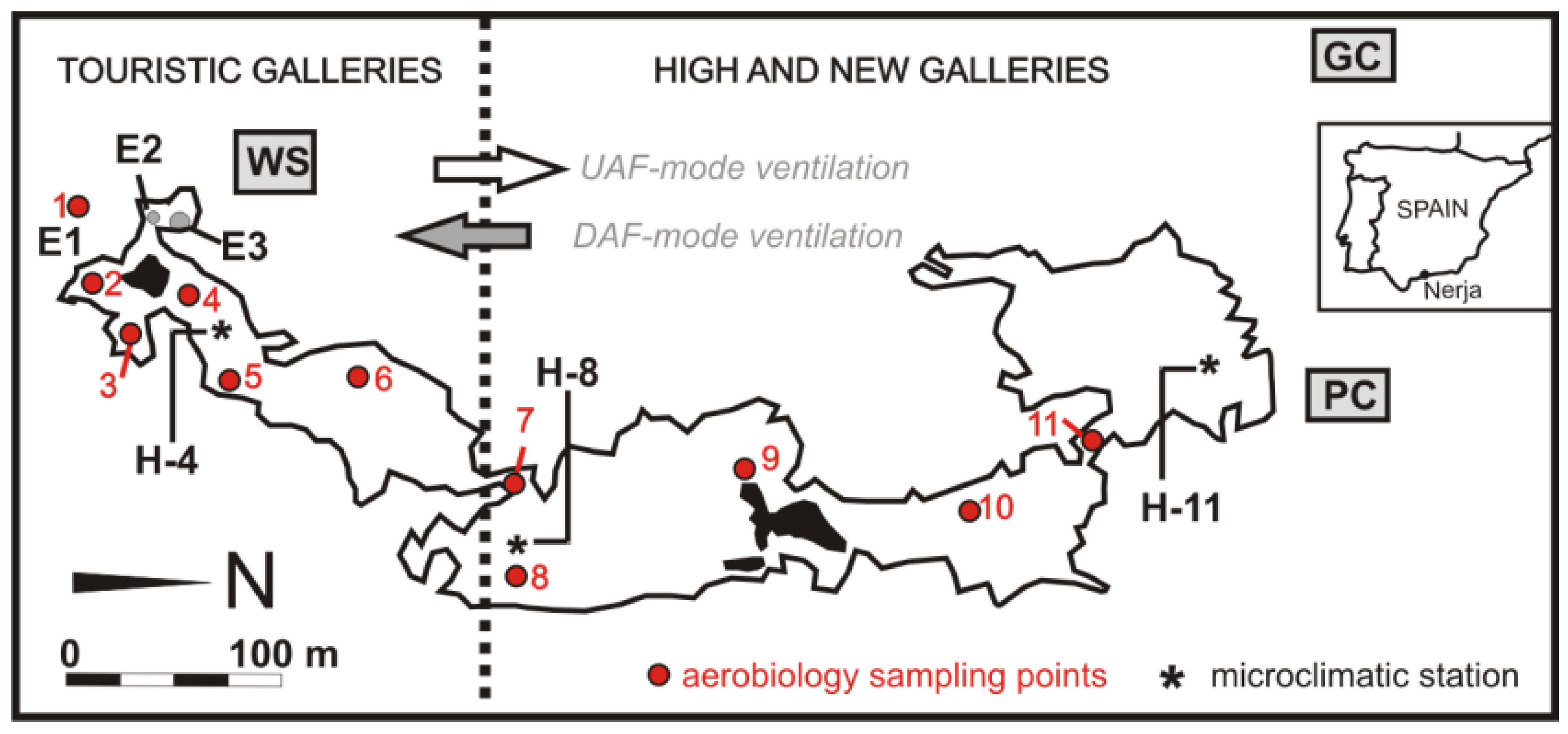
2.1. Site Description
2.2. Microclimatology
2.3. Aerobiology
3. Results
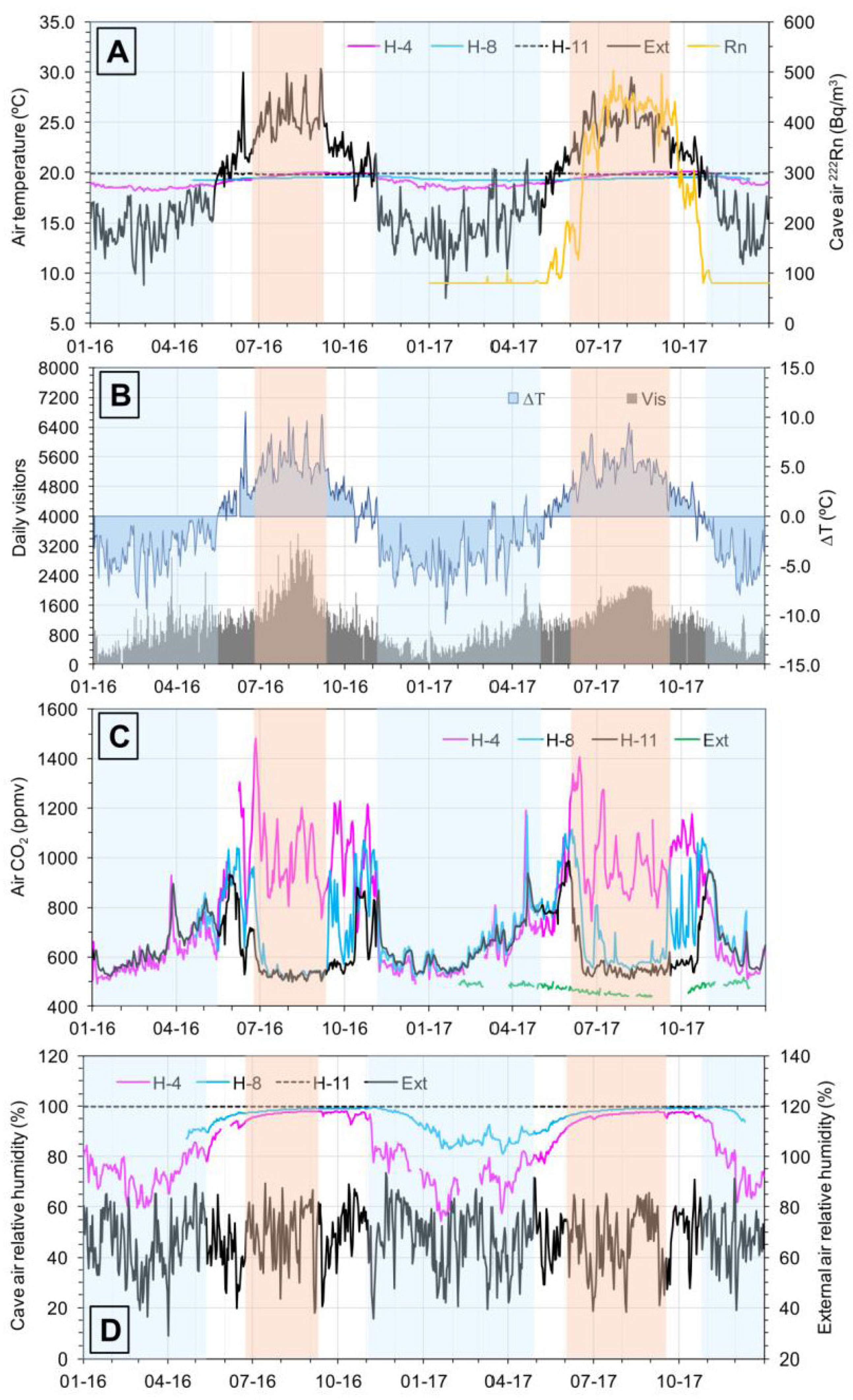
3.1. Microclimatology
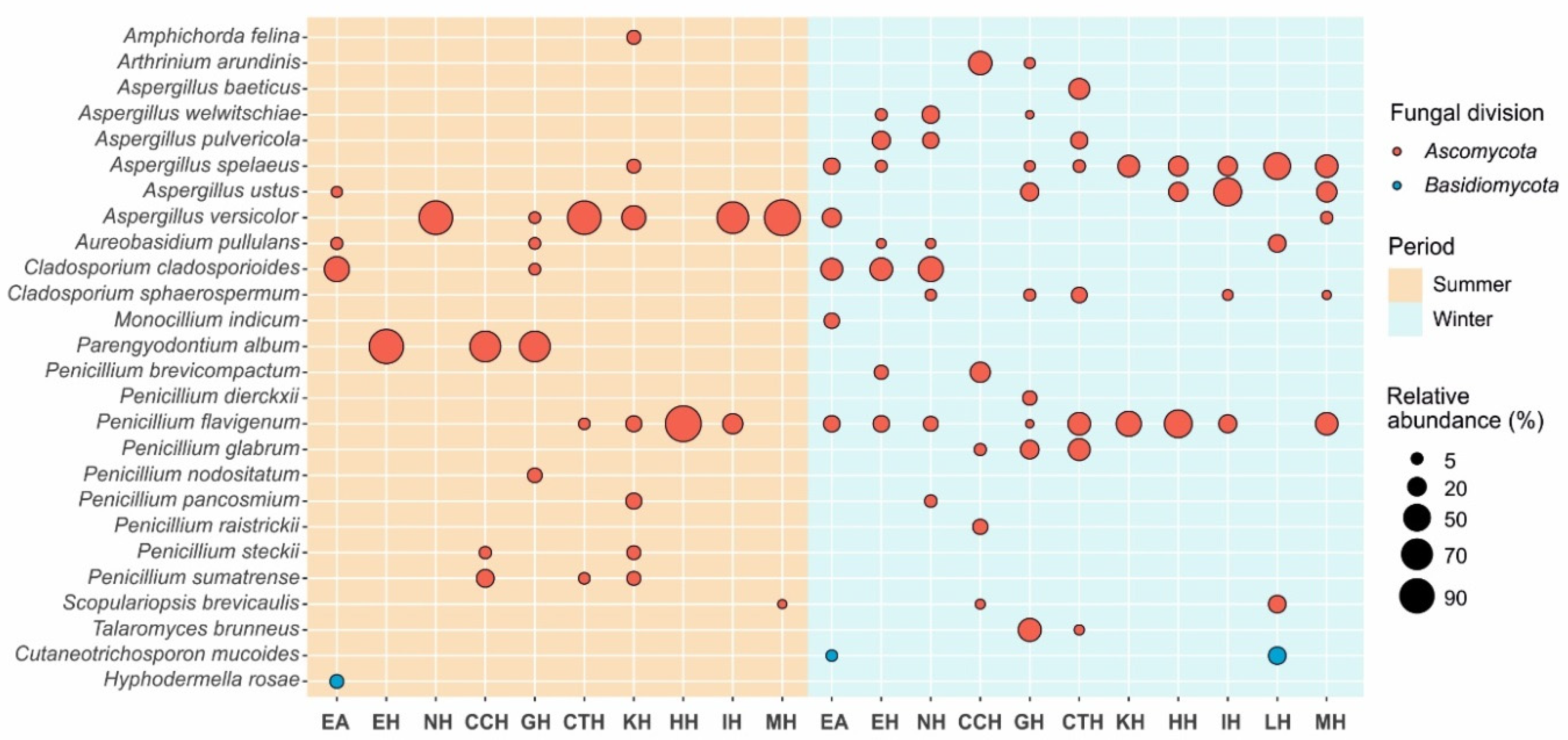
3.2. Aerobiology
3.2.1. Summer Campaign, 9 July 2016
3.2.2. Winter Campaign, 12 January 2017
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Adetutu, E.M.; Thorpe, K.; Bourne, S.; Cao, X.; Shahsavari, E.; Kirby, G.; Ball, A.S. Phylogenetic diversity of fungal communities in areas accessible and not accessible to tourists in Naracoorte Caves. Mycologia 2011, 103, 959–968. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martin-Sanchez, P.M.; Nováková, A.; Bastian, F.; Alabouvette, C.; Saiz-Jimenez, C. Use of biocides for the control of fungal outbreaks in subterranean environments: The case of the Lascaux Cave in France. Environ. Sci. Technol. 2012, 46, 3762–3770. [Google Scholar] [CrossRef]
- Di Piazza, S.; Isaia, M.; Vizzini, A.; Badino, G.; Voyron, S.; Zotti, M. First mycological assessment in hydrothermal caves of Monte Kronio (Sicily, southern Italy). Webbia. J. Plant Taxon. Geogr. 2017, 72, 277–285. [Google Scholar] [CrossRef]
- Man, B.; Wang, H.; Yun, Y.; Xiang, X.; Wang, R.; Duan, Y.; Cheng, X. Diversity of fungal communities in Heshang Cave of Central China revealed by mycobiome-sequencing. Front. Microbiol. 2018, 9, 1400. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ogórek, R.; Kozak, B.; Visnovská, Z.; Tancinová, D. Phenotypic and genotypic diversity of airborne fungal spores in Demänovská Ice Cave (Low Tatras, Slovakia). Aerobiologia 2018, 34, 13–28. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pusz, W.; Król, M.; Zwijacz-Kozica, T. Airborne fungi as indicators of ecosystem disturbance: An example from selected Tatra Mountains caves (Poland). Aerobiologia 2018, 34, 111–118. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, W.; Ma, X.; Ma, Y.; Mao, L.; Wu, F.; Ma, X.; An, L.; Feng, H. Seasonal dynamics of airborne fungi in different caves of the Mogao Grottoes, Dunhuang. China. Int. Biodeter. Biodegr. 2010, 64, 461–466. [Google Scholar] [CrossRef]
- Porca, E.; Jurado, V.; Martin-Sanchez, P.M.; Hermosin, B.; Bastian, F.; Alabouvette, C.; Saiz-Jimenez, C. Aerobiology: An ecological indicator for early detection and control of fungal outbreaks in caves. Ecol. Indic. 2011, 11, 1594–1598. [Google Scholar] [CrossRef]
- Fernandez-Cortes, A.; Cuezva, S.; Sanchez-Moral, S.; Porca, E.; Jurado, V.; Saiz-Jimenez, C. Detection of human-induced environmental disturbances in a show cave. Environ. Sci. Pollut. Res. 2011, 18, 1037–1045. [Google Scholar] [CrossRef] [Green Version]
- Martin-Sanchez, P.M.; Jurado, V.; Porca, E.; Bastian, F.; Lacanette, D.; Alabouvette, C.; Saiz-Jimenez, C. Airborne microorganisms in Lascaux Cave (France). Int. J. Speleol. 2014, 43, 295–303. [Google Scholar] [CrossRef]
- Carrasco, F.; Vadillo, I.; Liñan, C.; Andreo, B.; Durán, J.J. Control of environmental parameters for management and conservation of Nerja Cave (Malaga, Spain). Acta Carsologica 2002, 31, 105–122. [Google Scholar] [CrossRef]
- Liñán, C.; Vadillo, I.; Carrasco, F. Carbon dioxide concentration in air within the Nerja Cave (Malaga, Andalusia, Spain). Int. J. Speleol. 2008, 37, 99–106. [Google Scholar] [CrossRef]
- Liñán, C.; Ojeda, L.; Benavente, J.; Del Rosal, Y.; Vadillo, I.; Carrasco, F. Coupling air temperature records and gravimetric data to interpret ventilation patterns in a Mediterranean karstic system (Nerja-Pintada caves, southern Spain). Sci. Total Environ. 2020, 730, 139147. [Google Scholar] [CrossRef] [PubMed]
- Benavente, J.; Vadillo, I.; Liñan, C.; Carrasco, F.; Soler, A. Ventilation effects in a karstic show cave and in its vadose environment, Nerja, Southern Spain. Carbonates Evapotites 2011, 26, 11–17. [Google Scholar] [CrossRef]
- Dueñas, C.; Fernández, M.C.; Cañete, S.; Pérez, M.; Gordo, E. Seasonal variations of radon and the radiation exposure levels in Nerja cave, Spain. Radiat. Meas. 2011, 46, 1181–1186. [Google Scholar] [CrossRef]
- Del Rosal, Y.; Liñan, C.; Hernandez-Marine, M. The conservation of the Nerja Cave: Preserving anthropogenic impact in a tourist cave. In The Conservation of Subterranean Cultural Heritage; Saiz-Jimenez, C., Ed.; CRC Press/Balkema: Leiden, The Netherlands, 2014; pp. 193–206. [Google Scholar]
- Ojeda, L.; Vadillo, I.; Etiope, G.; Benavente, J.; Liñán, C.; Del Rosal, Y.; Tapia, S.T.; Moríñigo, M.A.; Carrasco, F. Methane sources and sinks in karst systems: The Nerja cave and its vadose environment (Spain). Geochim. Cosmochim. Acta 2019, 259, 302–315. [Google Scholar] [CrossRef]
- Garcia-Anton, E.; Cuezva, S.; Jurado, V.; Porca, E.; Miller, A.Z.; Fernandez-Cortes, A.; Saiz-Jimenez, C.; Sanchez-Moral, S. Combining stable isotope (δ13C) of trace gases and aerobiological data to monitor the entry and dispersion of microorganisms in caves. Environ. Sci. Pollut. Res. 2014, 21, 473–484. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sanchez-Moral, S.; Jurado, V.; Fernandez-Cortes, A.; Cuezva, S.; Martin-Pozas, T.; Gonzalez-Pimentel, J.L.; Ontañon, R.; Saiz-Jimenez, C. Environment-driven control of the dispersal of fungi in subterranean ecosystems: The case of La Garma Cave (northern Spain). Int. Microbiol. 2021, in press. [Google Scholar]
- Ruíz, J.; Marín-Girón, F.; Ojeda, F.; Marón-Olalla, F.; Berros, J.; Marín-Olalla, E. Estudio macroscópico “in situ” y microscópico-ecológico de pequeñas zonas de flora verde (algas verdes y verdeazuladas) del interior de la Cueva de Nerja. In Investigación Biológica y Edafológica de la Cueva de Nerja (Málaga); Marín, F., Carrasco, F., Eds.; Trabajos sobre la Cueva de Nerja: Málaga, Spain, 1992; Volume 2, pp. 113–125. [Google Scholar]
- Del Rosal, Y.; Hernández-Mariné, M.; Roldán, M. Phototrophic microorganisms in the tourist cave of Nerja. In Science, Technology and Cultural Heritage; Rogerio-Candelera, M.A., Ed.; CRC Press/Balkema: Leiden, The Netherlands, 2014; pp. 229–234. [Google Scholar]
- Del Rosal Padial, Y.; Jurado Lobo, V.; Hernández Mariné, M.; Roldán Molina, M.; Sáiz Jiménez, C. Biofilms en cuevas turísticas: La Cueva de Nerja y la Cueva del Tesoro. In El Karst y el Hombre: Las Cuevas como Patrimonio Mundial; Andreo, B., Durán, J.J., Eds.; Asociación de Cuevas Turísticas Españolas: Madrid, Spain, 2016; pp. 103–114. [Google Scholar]
- Jurado, V.; Del Rosal, Y.; Hernández-Mariné, M.; Galocha Zapata, I.; Domínguez-Moñino, I.; Rogerio Candelera, M.A.; Sáiz-Jiménez, C. Biología molecular de los biofilms fotosintéticos de la Cueva de Nerja. In Análisis, Impacto y Evolución de los Biofilms Fotosintéticos en Espeleotemas. El caso de la Cueva de Nerja (Málaga, España); Del Rosal Padial, Y., Ed.; Fundación Pública de Servicios Cueva de Nerja: Nerja, Spain, 2017; pp. 229–270. [Google Scholar]
- Jurado, V.; Del Rosal, Y.; Gonzalez-Pimentel, J.L.; Hermosin, B.; Saiz-Jimenez, C. Biological control of phototrophic biofilms in a show cave: The case of Nerja Cave. Appl. Sci. 2020, 10, 3448. [Google Scholar] [CrossRef]
- Del Rosal, Y.; Martínez-Manzanares, E.; Marín, F.; Liñán, C.; Simón, M.D. Análisis de la calidad microbiológica aérea en el interior de la Cueva de Nerja. In Cuevas Turísticas: Aportación al Desarrollo Sostenible; Durán, J.J., Robledo, P.A., Vázquez, J., Eds.; Instituto Geológico y Minero de España: Madrid, Spain, 2007; pp. 181–190. [Google Scholar]
- Docampo, S.; Trigo, M.M.; Recio, M.; Melgar, M.; Garcia-Sanchez, J.; Calderon-Ezquerro, M.C.; Cabezudo, B. High incidence of Aspergillus and Penicillium spores in the atmosphere of the cave of Nerja (Malaga, southern Spain). Aerobiologia 2010, 26, 89–98. [Google Scholar] [CrossRef]
- Docampo, S.; Trigo, M.M.; Recio, M.; Melgar, M.; García-Sánchez, J.; Cabezudo, B. Fungal spores content of the atmosphere of the Cave of Nerja (southern Spain): Diversity and origin. Sci. Total Environ. 2011, 409, 835–843. [Google Scholar] [CrossRef]
- Liñán, C.; Del Rosal, Y.; Carrasco, F.; Vadillo, I.; Benavente, J.; Ojeda, L. Highlighting the importance of transitional ventilation regimes in the management of Mediterranean show caves (Nerja-Pintada system, southern Spain). Sci. Total Environ. 2018, 631–632, 1268–1278. [Google Scholar] [CrossRef] [PubMed]
- Docampo, S.; Recio, M.; Trigo, M.M.; Cabezudo, B. Risk of pollen allergy in Nerja (southern Spain): A pollen calendar. Aerobiologia 2007, 23, 189–199. [Google Scholar] [CrossRef]
- Dominguez-Moñino, I.; Jurado, V.; Rogerio-Candelera, M.A.; Hermosin, B.; Saiz-Jimenez, C. Airborne fungi in show caves from Southern Spain. Appl. Sci. 2021, 11, 5027. [Google Scholar] [CrossRef]
- Grbic, M.L.; Vukojevic, J.; Simic, G.S.; Krizmanic, J.; Stupar, M. Biofilm forming cyanobacteria, algae and fungi on two historic monuments in Belgrade, Serbia. Arch. Biol. Sci. 2010, 62, 625–631. [Google Scholar] [CrossRef]
- Pfendler, S.; Borderie, F.; Bousta, F.; Alaoui-Sosse, L.; Alaoui-Sosse, B.; Aleya, L. Comparison of biocides, allelopathic substances and UV-C as treatments for biofilm proliferation on heritage monuments. J. Cult. Herit. 2018, 33, 117–124. [Google Scholar] [CrossRef]
- De Hoog, G.S. The genera Beauveria, Isaria, Tritirachium and Acrodontium gen. nov. Stud. Mycol. 1972, 1, 1–41. [Google Scholar]
- Novakova, A.; Hubka, V.; Saiz-Jimenez, C. Microscopic fungi isolated from cave air and sediments in the Nerja Cave-preliminary results. In The Conservation of Subterranean Cultural Heritage; Saiz-Jimenez, C., Ed.; CRC Press/Balkema: Leiden, The Netherlands, 2014; pp. 239–246. [Google Scholar]
- Jurado, V.; Sanchez-Moral, S.; Saiz-Jimenez, C. Entomogenous fungi and the conservation of the cultural heritage: A review. Int. Biodeter. Biodegr. 2008, 62, 325–330. [Google Scholar] [CrossRef] [Green Version]
- Leplat, J.; François, A.; Bousta, F. Parengyodontium album, a frequently reported fungal species in the cultural heritage environment. Fungal Biol. Rev. 2020, 34, 126–135. [Google Scholar] [CrossRef]
- Bastian, F.; Alabouvette, C.; Saiz-Jimenez, C. The impact of arthropods on fungal community structure in Lascaux Cave. J. Appl. Microbiol. 2009, 106, 1456–1462. [Google Scholar] [CrossRef] [PubMed]
- Jurado, V.; Porca, E.; Cuezva, S.; Fernandez-Cortes, A.; Sanchez-Moral, S.; Saiz-Jimenez, C. Fungal outbreak in a show cave. Sci. Total Environ. 2010, 408, 3632–3638. [Google Scholar] [CrossRef] [Green Version]
- Porca Belío, E. Aerobiología: Mecanismos de Dispersión de los Microorganismos en Cuevas Turísticas. Ph.D. Thesis, Facultad de Biología, Universidad de Sevilla, Seville, Spain, 2011. [Google Scholar]
- Saarela, M.; Alakomi, H.-L.; Suihko, M.-L.; Maunuksela, L.; Raaska, L.; Mattila-Sandholm, T. Heterotrophic microorganisms in air and biofilm samples from Roman catacombs, with special emphasis on actinobacteria and fungi. Int. Biodeter. Biodegr. 2004, 54, 27–37. [Google Scholar] [CrossRef]
- Simonovicová, A.; Gódyová, M.; Sevc, J. Airborne and soil microfungi as contaminants of stone in a hypogean cemetery. Int. Biodeter. Biodegr. 2004, 54, 7–11. [Google Scholar] [CrossRef]
- Nugari, M.P.; Pietrini, A.M.; Caneva, G.; Imperi, F.; Visca, P. Biodeterioration of mural paintings in a rocky habitat: The Crypt of the Original Sin (Matera, Italy). Int. Biodeter. Biodegr. 2009, 63, 705–711. [Google Scholar] [CrossRef]
- Mang, S.M.; Scrano, L.; Camele, I. Preliminary studies on fungal contamination of two rupestrian churches from Matera (Southern Italy). Sustainability 2020, 12, 6988. [Google Scholar] [CrossRef]
- Saiz-Jimenez, C.; Samson, R.A. Microorganisms and environmental pollution as deteriorating agents of the frescoes of ‘‘Santa María de la Rábida’’, Huelva, Spain. In Sixth Triennial Meeting ICOM; Paper 81/15/5; Committee for Conservation: Ottawa, ON, Canada, 1981; p. 14. [Google Scholar]
- Gorbushina, A.A.; Petersen, K. Distribution of microorganisms on ancient wall paintings as related to associated faunal elements. Int. Biodeter. Biodegr. 2000, 46, 277–284. [Google Scholar] [CrossRef]
- Berner, M.; Wanner, G.; Lubitz, W. A comparative study of the fungal flora present in medieval wall paintings in the chapel of the castle Herberstein and in the parish church of St. Georgen in Styria, Austria. Int. Biodeter. Biodegr. 2004, 40, 53–61. [Google Scholar] [CrossRef]
- Barranco, P.; Tinaut, A.; Baena, M. Entomofauna cavernícola de Andalucía. In El Karst de Andalucía. Geoespeleología, Bioespeleología y Presencia Humana; Calaforra Chordi, J.M., Berrocal Pérez, J.A., Eds.; Consejería de Medio Ambiente, Junta de Andalucía: Sevilla, Spain, 2008; pp. 75–85. [Google Scholar]
- Gunde-Cimerman, N.; Ramos, J.; Plemenitas, A. Halotolerant and halophilic fungi. Mycol. Res. 2009, 113, 1231–1241. [Google Scholar] [CrossRef]
- Nováková, A. Microscopic fungi in caves of the National Park Slovak Karst. Phytopedon 2004, 3, 26–31. [Google Scholar]
- Nováková, A. Microscopic fungi isolated from the Domica Cave system (Slovak Karst National Park, Slovakia). A review. Int. J. Speleol. 2009, 38, 71–82. [Google Scholar] [CrossRef] [Green Version]
- Domínguez-Moñino, I.; Jurado, V.; Hermosín, B.; Sáiz-Jiménez, C. Aerobiología de cuevas andaluzas. In Las Cuevas Turísticas como Activos Económicos: Conservación e Innovación; Durán, J.J., Robledo, P.A., Eds.; Asociación de Cuevas Turísticas Españolas: Madrid, Spain, 2012; pp. 299–307. [Google Scholar]
- Taylor, E.L.S.; Resende-Stoianoff, M.A.A.; Lopes Ferreira, R. Mycological study for a management plan of a neotropical show cave (Brazil). Int. J. Speleol. 2013, 42, 267–277. [Google Scholar] [CrossRef] [Green Version]
- Mitova, M.M.; Iliev, M.; Nováková, A.; Gorbushina, A.A.; Groudeva, V.I.; Martin-Sanchez, P.M. Diversity and biocide susceptibility of fungal assemblages dwelling in the Art Gallery of Magura Cave, Bulgaria. Int. J. Speleol. 2017, 46, 67–80. [Google Scholar] [CrossRef] [Green Version]
- Mammola, S.; Di Piazza, S.; Zotti, M.; Isaia, M. Human-induced alterations of the cave mycobiota in an Alpine show cave (ltaly, SW-Alps). Acta Carsologica 2017, 46, 111–123. [Google Scholar] [CrossRef] [Green Version]
- Marvanová, L.; Kalouskova, V.; Hanuláková, D.; Scháněl, L. Microscopic fungi in the Zbrašov aragonite caves. Czech Mycol. 1992, 46, 243–250. [Google Scholar]
- Novakova, A.; Jurado, V.; Saiz-Jimenez, C. Are fungi a real threat for the conservation of Altamira Cave? In The Conservation of Subterranean Cultural Heritage; Saiz-Jimenez, C., Ed.; CRC Press/Balkema: Leiden, The Netherlands, 2014; pp. 223–228. [Google Scholar]
- Hubka, V.; Nováková, A.; Peterson, S.W.; Frisvad, J.C.; Sklenář, F.; Matsuzawa, T.; Kubátová, A.; Kolarík, M. A reappraisal of Aspergillus section Nidulantes with descriptions of two new sterigmatocystin-producing species. Plant Syst. Evol. 2016, 302, 1267–1299. [Google Scholar] [CrossRef]
- Nováková, A.; Hubka, V.; Saiz-Jimenez, C.; Kolarik, M. Aspergillus baeticus sp. nov. and Aspergillus thesauricus sp. nov.: Two new species in section Usti originating from Spanish caves. Int. J. Syst. Evol. Microbiol. 2012, 62, 2778–2785. [Google Scholar] [CrossRef]
- Nováková, A.; Kubátová, A.; Sklenář, F.; Hubka, V. Microscopic fungi on cadavers and skeletons from cave and mine environments. Czech Mycol. 2018, 70, 101–121. [Google Scholar] [CrossRef]
- Vanderwolf, K.J.; Malloch, D.; McAlpine, D.F.; Forbes, G.J. A world review of fungi, yeasts, and slime molds in caves. Int. J. Speleol. 2013, 42, 77–96. [Google Scholar] [CrossRef]
- Popović, S.; Simić, G.S.; Stupar, M.; Unković, N.; Predojević, D.; Jovanović, J.; Grbić, M.L. Cyanobacteria, algae and microfungi present in biofilm from Božana Cave (Serbia). Int. J. Speleol. 2015, 44, 141–149. [Google Scholar] [CrossRef] [Green Version]
- Kozlova, E.V.; Mazina, S.E. Biodiversity of fungi in the photic and aphotic zones of Montenegro caves. Aerobiologia 2020, 36, 589–604. [Google Scholar] [CrossRef]
- Pusz, W.; Ogórek, R.; Knapik, R.; Kozak, B.; Bujak, H. The occurrence of fungi in the recently discovered Jarkowicka Cave in the Karkonosze Mts. (Poland). Geomicrobiol. J. 2015, 32, 59–67. [Google Scholar] [CrossRef]
- Nováková, A.; Hubka, V.; Valinova, S.; Kolarik, M.; Hillebrand-Voiculescu, A.M. Cultivable microscopic fungi from an underground chemosynthesis-based ecosystem: A preliminary study. Folia Microbiol. 2018, 63, 43–55. [Google Scholar] [CrossRef]
- Tavares, D.G.; Barbosa, B.V.L.; Ferreira, R.L.; Duarte, W.F.; Cardoso, P.G. Antioxidant activity and phenolic compounds of the extract from pigmentproducing fungi isolated from Brazilian caves. Biocatal. Agric. Biotechnol. 2018, 16, 148–154. [Google Scholar] [CrossRef]
- Zhang, Z.F.; Zhou, S.Y.; Eurwilaichitr, L.; Ingsriswang, S.; Raza, M.; Chen, Q.; Zhao, P.; Liu, F.; Cai, L. Culturable mycobiota from Karst caves in China II, with descriptions of 33 new species. Fungal Divers. 2021, 106, 29–136. [Google Scholar] [CrossRef]
- Madsen, A.M.; Larsen, S.T.; Koponen, I.K.; Kling, K.I.; Barooni, A.; Karottki, D.G.; Tendal, K.; Wolkoffa, P. Generation and characterization of indoor fungal aerosols for inhalation studies. Appl. Environ. Microbiol. 2016, 82, 2479–2493. [Google Scholar] [CrossRef] [Green Version]
- Samson, R.A.; Yilmaz, N.; Houbraken, J.; Spierenburg, H.; Seifert, K.A.; Peterson, S.W.; Varga, J.; Frisvad, J.C. Phylogeny and nomenclature of the genus Talaromyces and taxa accommodated in Penicillium subgenus Biverticillium. Stud. Mycol. 2011, 70, 159–183. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shibata, S.; Udagawa, S. Metabolic products of fungi. XIX. Isolation of rugulosin from Penicillium brunneum Udagawa. Chem. Pharm. Bull. 1963, 11, 402–403. [Google Scholar] [CrossRef] [Green Version]
- Paula, C.C.P.; Montoya, Q.V.; Meirelles, L.A.; Sanchez, C.F.; Rodrigues, A.; Seleghim, M.H.R. High cellulolytic activities in filamentous fungi isolated from an extreme oligotrophic subterranean environment (Catão cave) in Brazil. An. Acad. Bras. Cienc. 2019, 91, e20180583. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, J.-R.; Cai, L.; Liu, F. Oligotrophic fungi from a carbonate cave, with three new species of Cephalotrichum. Mycology 2017, 8, 164–177. [Google Scholar] [CrossRef] [Green Version]
- Razafinarivo, J.; Jany, J.-L.; Crous, P.W.; Looten, R.; Gaydou, V.; Barbier, G.; Mounier, J.; Vasseur, V. Cladosporium lebrasiae, a new fungal species isolated from milk bread rolls in France. Fungal Biol. 2016, 120, 1017–1029. [Google Scholar] [CrossRef]
- Zalar, P.; de Hoog, G.S.; Schroers, H.-J.; Crous, P.W.; Groenewald, J.Z.; Gunde-Cimerman, N. Phylogeny and ecology of the ubiquitous saprobe Cladosporium sphaerospermum, with descriptions of seven new species from hypersaline environments. Stud. Mycol. 2007, 58, 157–183. [Google Scholar] [CrossRef]
- Manamgoda, D.S.; Rossman, A.Y.; Castlebury, L.A.; Crous, P.W.; Madrid, H.; Chukeatirote, E.; Hyde, K.D. The genus Bipolaris. Stud. Mycol. 2014, 79, 221–288. [Google Scholar] [CrossRef] [Green Version]
- Wang, M.; Tan, X.-M.; Liu, F.; Cai, L. Eight new Arthrinium species from China. MycoKeys 2018, 34, 1–24. [Google Scholar] [CrossRef] [Green Version]
- Saiz-Jimenez, C. Cave Conservation: A Microbiologist’s Perspective. In Cave Microbiomes: A Novel Resource for Drug Discovery; Cheepham, N., Ed.; SpringerBrief in Microbiology 1: New York, NY, USA, 2013; pp. 69–84. [Google Scholar]
- Zhang, Z.F.; Liu, F.; Zhou, X.; Liu, X.Z.; Liu, S.J.; Cai, L. Culturable mycobiota from Karst caves in China, with descriptions of 20 new species. Persoonia 2017, 39, 1–31. [Google Scholar] [CrossRef]
- Belyagoubi, L.; Belyagoubi-Benhammou, N.; Jurado, V.; Dupont, J.; Lacoste, S.; Djebbah, F.; Ounadjela, F.Z.; Benaissa, S.; Habi, S.; Abdelouahi, D.E.; et al. Antimicrobial activities of culturable microorganisms (actinomycetes and fungi) isolated from Chaabe Cave, Algeria. Int. J. Speleol. 2018, 47, 189–199. [Google Scholar] [CrossRef] [Green Version]
- Woudenberg, J.H.C.; Meijer, M.; Houbraken, J.; Samson, R.A. Scopulariopsis and scopulariopsis-like species from indoor environments. Stud. Mycol. 2017, 88, 1–35. [Google Scholar] [CrossRef]
- Larcher, G.; Bouchara, J.P.; Pailley, P.; Montfort, D.; Beguin, H.; De Biève, C.; Chabasse, D. Fungal biota associated with bats in Western France. J. Med. Mycol. 2003, 13, 29–34. [Google Scholar]
- Vaughan-Martini, A.; Angelini, P.; Zacchi, L. The influence of human and animal visitation on the yeast ecology of three Italian caverns. Ann. Microbiol. 2000, 50, 133–140. [Google Scholar]
- Sugita, T.; Kikuchi, K.; Makimura, K.; Urata, K.; Someya, T.; Kamei, K.; Niimi, M.; Uehara, Y. Trichosporon species isolated from guano samples obtained from bat-inshabited caves in Japan. Appl. Environ. Microbiol. 2005, 71, 7626–7629. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gómez de Ana, S.; Torres-Rodríguez, J.M.; Ramírez, E.A.; Belmonte-Soler, J. Seasonal distribution of Alternaria, Aspergillus, Cladosporium and Penicillium species isolated in homes of fungal allergic patients. J. Investig. Allergol. Clin. Immunol. 2006, 16, 357–363. [Google Scholar]
- Hurst, C.J. Understanding Terrestrial Microbial Communities; Springer: Cham, Switzerland, 2019. [Google Scholar]
- Davolos, D.; Pietrangeli, B.; Persiani, A.M.; Maggi, O. Victoriomyces antarcticus gen. nov., sp. nov., a distinct evolutionary lineage of the Cephalothecaceae (Ascomycota) based on sequence-based phylogeny and morphology. Int. J. Syst. Evol. Microbiol. 2019, 69, 1099–1110. [Google Scholar] [CrossRef] [PubMed]
- Mapook, A.; Hyde, K.D.; McKenzie, E.H.C.; Jones, E.B.G.; Bhat, D.J.; Jeewon, R.; Stadler, M.; Samarakoon, M.C.; Malaithong, M.; Tanunchai, B.; et al. Taxonomic and phylogenetic contributions to fungi associated with the invasive weed Chromolaena odorata (Siam weed). Fungal Divers. 2020, 101, 1–175. [Google Scholar] [CrossRef]
- Sandoval-Denis, M.; Gené, J.; Sutton, D.A.; Cano-Lira, J.F.; de Hoog, G.S.; Decock, C.A.; Wiederhold, N.P.; Guarro, J. Redefining Microascus, Scopulariopsis and allied genera. Persoonia 2016, 36, 1–36. [Google Scholar] [CrossRef] [Green Version]
- Melo, A.G.; Souza, P.N.C.; Maia, N.C.; Thomas, A.B.; Silva, L.B.R.; Batista, L.R.; Ferreira, R.L.; Cardoso, P.G. Screening and identification of tannase-producing fungi isolated from Brazilian caves. African J. Microbiol. Res. 2013, 7, 483–487. [Google Scholar]
- Cunha, A.O.B.; Bezerra, J.D.P.; Oliveira, T.G.L.; Barbier, E.; Bernard, E.; Machado, A.R.; Souza-Motta, C.M. Living in the dark: Bat caves as hotspots of fungal diversity. PLoS ONE 2020, 15, e0243494. [Google Scholar] [CrossRef] [PubMed]
- Trovão, J.; Mesquita, N.; Paiva, D.S.; Carvalho, H.P.; Avelar, L.; Portugal, A. Can arthropods act as vectors of fungal dispersion in heritage collections? A case study on the archive of the University of Coimbra, Portugal. Int. Biodeter. Biodegr. 2013, 79, 49–55. [Google Scholar] [CrossRef]
- Jaber, S.; Mercier, A.; Knio, K.; Brum, S.; Kambris, Z. Isolation of fungi from dead arthropods and identification of a new mosquito natural pathogen. Parasite Vector 2016, 9, 491. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yun, T.-S.; Park, S.Y.; Yu, J.; Hwang, Y.; Hong, K.-J. Isolation and identification of fungal species from the insect pest Tribolium castaneum in rice processing complexes in Korea. Plant Pathol. J. 2018, 34, 356–366. [Google Scholar] [CrossRef]
- Jenoh, E.M.; de Villiers, E.P.; de Villiers, S.M.; Okoth, S.; Jefwa, J.; Kioko, E.; Kaimenyi, D.; Hendrickx, M.; Dahdouh-Guebas, F.; Koedam, N. Infestation mechanisms of two woodborer species in the mangrove Sonneratia alba J. Smith in Kenya and co-occurring endophytic fungi. PLoS ONE 2019, 14, e0221285. [Google Scholar] [CrossRef]
- Hubka, V.; Nováková, A.; Kolarík, M.; Jurjević, A.; Peterson, S.W. Revision of Aspergillus section Flavipedes: Seven new species and proposal of section Jani sect. nov. Mycologia 2015, 107, 169–208. [Google Scholar] [CrossRef]
- Dominguez-Moñino, I.; Jurado, V.; Hermosin, B.; Saiz-Jimenez, C. Paracoccus cavernae sp. nov., isolated from a show cave. Int. J. Syst. Evol. Microbiol. 2016, 66, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Dominguez-Moñino, I.; Jurado, V.; Gonzalez-Pimentel, J.L.; Miller, A.Z.; Hermosin, B.; Saiz-Jimenez, C. Bacillus onubensis sp. nov. isolated from the air of two Andalusian caves. Syst. Appl. Microbiol. 2018, 41, 167–172. [Google Scholar] [CrossRef] [PubMed] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| WS | H-4 | H-8 | H-11 | WS | H-4 | H-8 | H-11 | |
|---|---|---|---|---|---|---|---|---|
| Air Temperature (°C) | Relative Humidity (%) | |||||||
| n | 731 | 727 | 600 | 731 | 731 | 688 | 600 | 731 |
| max | 30.34 | 20.08 | 19.66 | 19.92 | 93.68 | 98.11 | 99.77 | 100.00 |
| min | 7.27 | 18.15 | 19.18 | 19.75 | 28.92 | 54.40 | 81.11 | 100.00 |
| m | 18.93 | 19.21 | 19.40 | 19.86 | 67.08 | 84.49 | 95.21 | 100.00 |
| s | 4.80 | 0.60 | 0.13 | 0.04 | 11.69 | 12.27 | 4.96 | 0.00 |
| v (%) | 25.33 | 3.10 | 0.69 | 0.19 | 17.43 | 14.53 | 5.21 | 0.00 |
| range | 22.88 | 1.93 | 0.48 | 0.16 | 65.00 | 43.71 | 18.66 | 0.00 |
| WS | H-4 | H-8 | H-11 | WS | H-4 | H-8 | H-11 | |
| Air CO2 (ppmv) | Air 222Rn (Bq/m3) | |||||||
| n | 223 | 688 | 600 | 731 | nd | 1379 | nd | nd |
| max | 518 | 1483 | 1170 | 986 | nd | 622 | nd | nd |
| min | 436 | 489 | 512 | 499 | nd | 80 | nd | nd |
| m | 474 | 797 | 699 | 633 | nd | 213 | nd | nd |
| s | 19 | 226 | 158 | 111 | nd | 161 | nd | nd |
| v (%) | 4 | 28 | 23 | 18 | nd | 76 | nd | nd |
| range | 83 | 994 | 657 | 487 | nd | 542 | nd | nd |
| Sampling | CFU m−3 (SD) * | Identification (% Similarity) | Abundance (%) |
|---|---|---|---|
| Entrance Hall (EH) | 170 (40) | Cladosporium lebrasiae (100%) | 6.45 |
| Parengyodontium album (100%) | 90.32 | ||
| Penicillium citreosulfuratum (100%) | 3.23 | ||
| Nativity Hall (NH) | 80 (20) | Alternaria tenuissima (100%) | 6.67 |
| Aspergillus versicolor (100%) | 86.67 | ||
| Penicillium citreosulfuratum (100%) | 6.67 | ||
| Cascade Hall (CCH) | 90 (20) | Parengyodontium album (100%) | 70.59 |
| Phoma sp. (99%) | 5.88 | ||
| Penicillium sumatrense (100%) | 17.65 | ||
| Penicillium steckii (100%) | 5.88 | ||
| Ghosts Hall (GH) | 100 (0) | Aspergillus versicolor (100%) | 5.00 |
| Aureobasidium pullulans (100%) | 5.00 | ||
| Bipolaris cynodontis (100%) | 5.00 | ||
| Cladosporium cladosporioides (100%) | 5.00 | ||
| Parengyodontium album (100%) | 70.00 | ||
| Penicillium nodositatum (99%) | 10.00 | ||
| Cataclysm Hall (CTH) | 110 (20) | Aspergillus versicolor (100%) | 85.71 |
| Penicillium citreosulfuratum (100%) | 4.76 | ||
| Penicillium flavigenum (100%) | 4.76 | ||
| Penicillium sumatrense (100%) | 4.76 | ||
| Kitchen Hall (KH) | 120 (10) | Aspergillus spelaeus (100%) | 8.70 |
| Aspergillus versicolor (100%) | 39.13 | ||
| Amphichorda felina (100%) | 8.70 | ||
| Penicillium flavigenum (100%) | 13.04 | ||
| Penicillium pancosmium (100%) | 13.04 | ||
| Penicillium steckii (100%) | 8.70 | ||
| Penicillium sumatrense (100%) | 8.70 | ||
| Heracles Hall (HH) | 10 (0) | Penicillium flavigenum (100%) | 100.00 |
| Immensity Hall (IH) | 20 (0) | Aspergillus versicolor (100%) | 75.00 |
| Penicillium flavigenum (100%) | 25.00 | ||
| Lance Hall (LH) | 0 | No detected | - |
| Exterior air (EA) | 390 (60) | Tremateia chromolaenae (99%) | 5.56 |
| Victoriomyces antarcticus (100%) | 2.78 | ||
| Alternaria tenuissima (100%) | 1.39 | ||
| Aspergillus tubingensis (100%) | 1.39 | ||
| Aspergillus melleus (100%) | 5.56 | ||
| Aspergillus ustus (100%) | 4.17 | ||
| Aureobasidium pullulans (100%) | 5.56 | ||
| Botrytis cinerea (100%) | 2.78 | ||
| Cladosporium cladosporioides (100%) | 43.06 | ||
| Hyphodermella rosae (100%) | 8.33 | ||
| Microascus croci (99%) | 5.56 | ||
| Penicillium olsonii (99%) | 5.56 | ||
| Purpureocillium lilacinum (100%) | 8.33 |
| Sampling | CFU m−3 (SD) * | Identification (% Similarity) | Abundance (%) |
|---|---|---|---|
| Entrance Hall (EH) | 430 (10) | Aspergillus welwitschiae (99%) | 5.13 |
| Aspergillus pulvericola (99%) | 19.23 | ||
| Aspergillus spelaeus (99%) | 5.13 | ||
| Aureobasidium pullulans (100%) | 2.56 | ||
| Botrytis cinerea (100%) | 1.28 | ||
| Cladosporium cladosporioides (100%) | 33.33 | ||
| Penicillium brevicompactum (100%) | 8.97 | ||
| Aspergillus europaeus (100%) | 2.56 | ||
| Penicillium flavigenum (100%) | 14.10 | ||
| Penicillium rubens (100%) | 7.69 | ||
| Nativity Hall (NH) | 360 (30) | Aspergillus welwitschiae (99%) | 16.67 |
| Aspergillus pulvericola (99%) | 13.64 | ||
| Aspergillus ochraceus (100%) | 1.52 | ||
| Aureobasidium pullulans (100%) | 3.03 | ||
| Botrytis cinerea (100%) | 1.52 | ||
| Cladosporium cladosporioides (100%) | 42.42 | ||
| Cladosporium sphaerospermum (100%) | 4.55 | ||
| Penicillium flavigenum (100%) | 10.61 | ||
| Penicillium pancosmium (99%) | 6.06 | ||
| Cascade Hall (CCH) | 190 (30) | Arthrinium arundinis (99%) | 36.11 |
| Penicillium olsonii (99%) | 5.56 | ||
| Penicillium sumatrense (98%) | 13.89 | ||
| Penicillium glabrum (100%) | 5.56 | ||
| Penicillium raistrickii (99%) | 11.11 | ||
| Penicillium brevicompactum (100%) | 25.00 | ||
| Scopulariopsis brevicaulis (100%) | 2.78 | ||
| Ghosts Hall (GH) | 420 (40) | Arthrinium arundinis (99%) | 4.00 |
| Aspergillus welwitschiae (99%) | 1.33 | ||
| Aspergillus spelaeus (99%) | 4.00 | ||
| Aspergillus ustus (100%) | 18.67 | ||
| Cladosporium sphaerospermum (100%) | 5.33 | ||
| Penicillium glabrum (100%) | 20.00 | ||
| Penicillium dierckxii (100%) | 9.33 | ||
| Penicillium flavigenum (100%) | 1.33 | ||
| Aspergillus oryzae (100%) | 1.33 | ||
| Talaromyces brunneus (100%) | 34.67 | ||
| Cataclysm Hall (CTH) | 180 (60) | Aspergillus spelaeus (99%) | 6.06 |
| Aspergillus pulvericola (99%) | 15.15 | ||
| Cladosporium sphaerospermum (100%) | 12.12 | ||
| Penicillium flavigenum (100%) | 33.33 | ||
| Penicillium glabrum (100%) | 30.30 | ||
| Talaromyces brunneus (100%) | 3.03 | ||
| Kitchen Hall (KH) | 2170 (130) | Aspergillus baeticus (100%) | 26.91 |
| Aspergillus spelaeus (99%) | 30.18 | ||
| Penicillium flavigenum (100%) | 42.91 | ||
| Heracles Hall (HH) | 1330 (40) | Aspergillus spelaeus (99%) | 23.50 |
| Aspergillus ustus (100%) | 21.50 | ||
| Penicillium flavigenum (100%) | 55.00 | ||
| Immensity Hall (IH) | 140 (20) | Aspergillus ustus (100%) | 55.56 |
| Aspergillus spelaeus (99%) | 22.22 | ||
| Cladosporium sphaerospermum (100%) | 3.70 | ||
| Penicillium flavigenum (100%) | 18.52 | ||
| Lance Hall (LH) | 30 (10) | Aspergillus spelaeus (99%) | 50.00 |
| Aureobasidium pullulans (100%) | 16.67 | ||
| Cutaneotrichosporon mucoides (100%) | 16.67 | ||
| Scopulariopsis brevicaulis (100%) | 16.67 | ||
| Mountain Hall (MH) | 280 (0) | Aspergillus ustus (100%) | 25.00 |
| Aspergillus spelaeus (99%) | 32.69 | ||
| Aspergillus versicolor (100%) | 5.77 | ||
| Cladosporium sphaerospermum (100%) | 1.92 | ||
| Penicillium flavigenum (100%) | 32.69 | ||
| Scopulariopsis brevicaulis (100%) | 1.92 | ||
| Exterior air (EA) | 230 (20) | Alternaria tenuissima (100%) | 2.33 |
| Aspergillus spelaeus (99%) | 13.95 | ||
| Aspergillus versicolor (100%) | 20.93 | ||
| Botrytis cinerea (100%) | 2.33 | ||
| Cladosporium cladosporioides (100%) | 30.23 | ||
| Cutaneotrichosporon mucoides (100%) | 4.65 | ||
| Monocillium indicum (99%) | 11.63 | ||
| Penicillium flavigenum (100%) | 13.95 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jurado, V.; Del Rosal, Y.; Liñan, C.; Martin-Pozas, T.; Gonzalez-Pimentel, J.L.; Saiz-Jimenez, C. Diversity and Seasonal Dynamics of Airborne Fungi in Nerja Cave, Spain. Appl. Sci. 2021, 11, 6236. https://doi.org/10.3390/app11136236
Jurado V, Del Rosal Y, Liñan C, Martin-Pozas T, Gonzalez-Pimentel JL, Saiz-Jimenez C. Diversity and Seasonal Dynamics of Airborne Fungi in Nerja Cave, Spain. Applied Sciences. 2021; 11(13):6236. https://doi.org/10.3390/app11136236
Chicago/Turabian StyleJurado, Valme, Yolanda Del Rosal, Cristina Liñan, Tamara Martin-Pozas, Jose Luis Gonzalez-Pimentel, and Cesareo Saiz-Jimenez. 2021. "Diversity and Seasonal Dynamics of Airborne Fungi in Nerja Cave, Spain" Applied Sciences 11, no. 13: 6236. https://doi.org/10.3390/app11136236
APA StyleJurado, V., Del Rosal, Y., Liñan, C., Martin-Pozas, T., Gonzalez-Pimentel, J. L., & Saiz-Jimenez, C. (2021). Diversity and Seasonal Dynamics of Airborne Fungi in Nerja Cave, Spain. Applied Sciences, 11(13), 6236. https://doi.org/10.3390/app11136236