
Metabolic Reaction Network-Based Model Predictive Control of Bioprocesses

 ,
, 
 ,
,
Abstract
:1. Introduction
2. Methods
2.1. Metabolic Reaction Network-Based Modeling
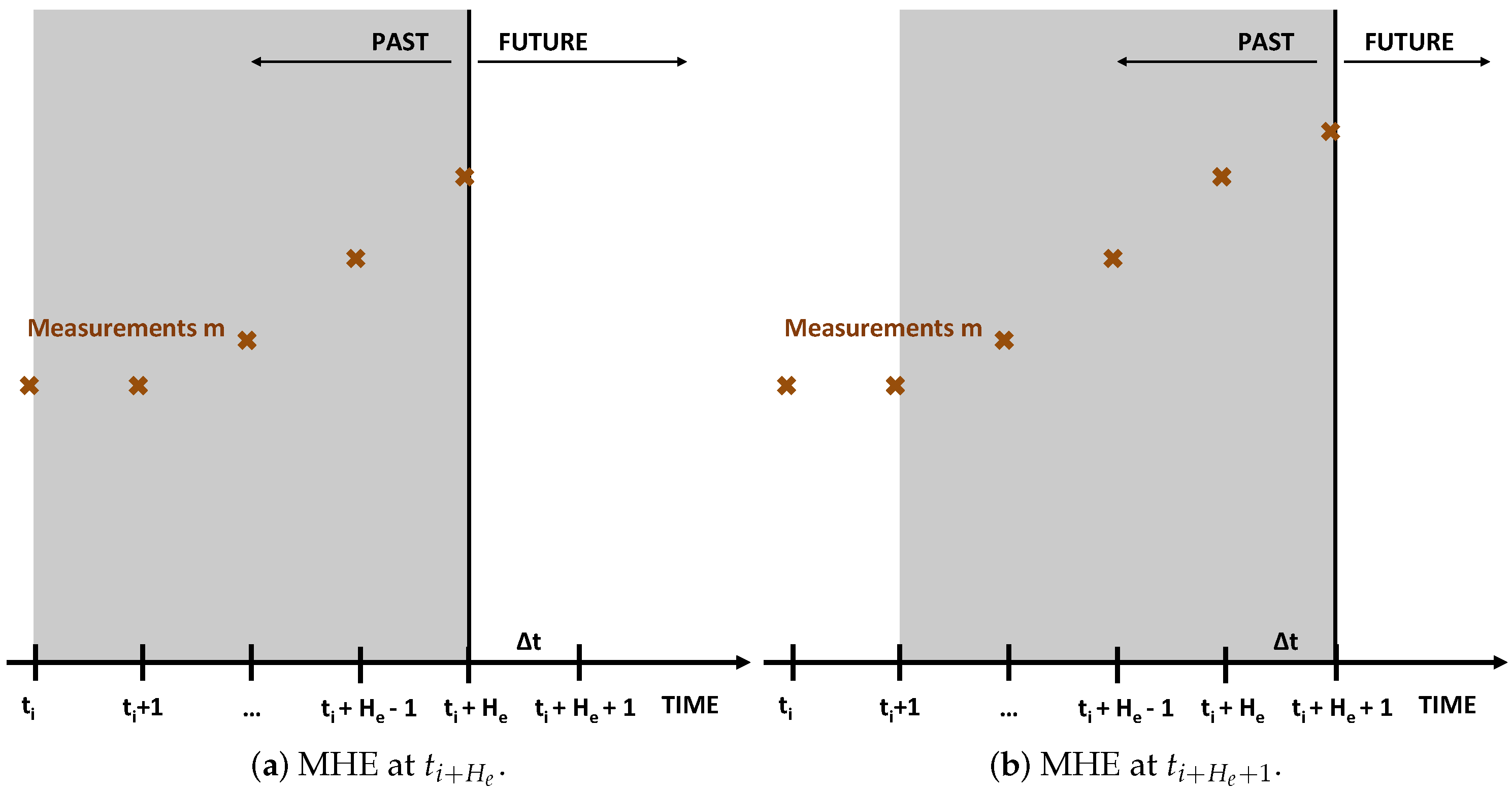
2.2. Moving Horizon Estimation (MHE)
2.3. Unscented Kalman Filter (UKF)
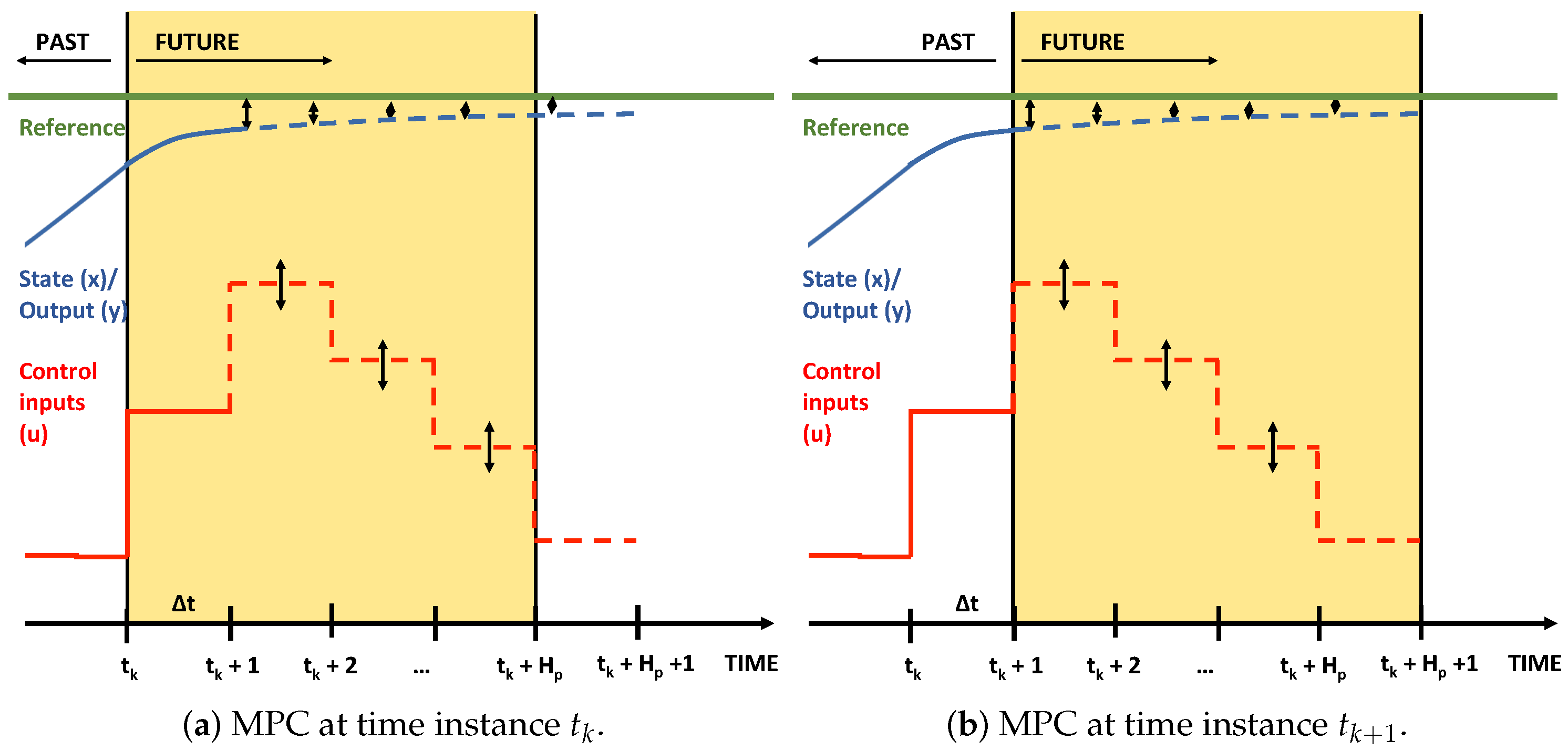
2.4. Model Predictive Control (MPC)
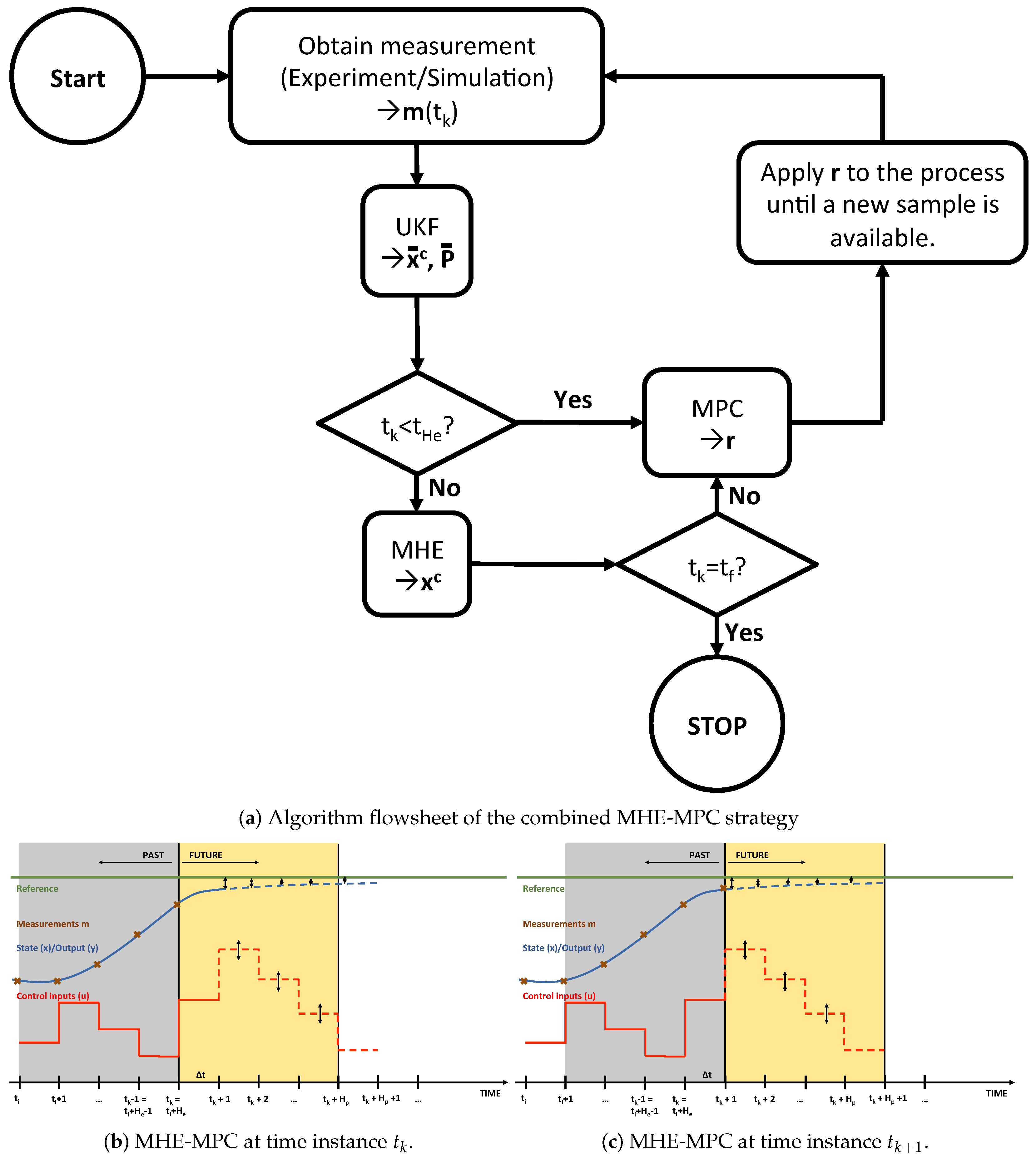
2.5. Combined MHE/MPC Strategy
3. Results and Discussion
3.1. Case Study Description
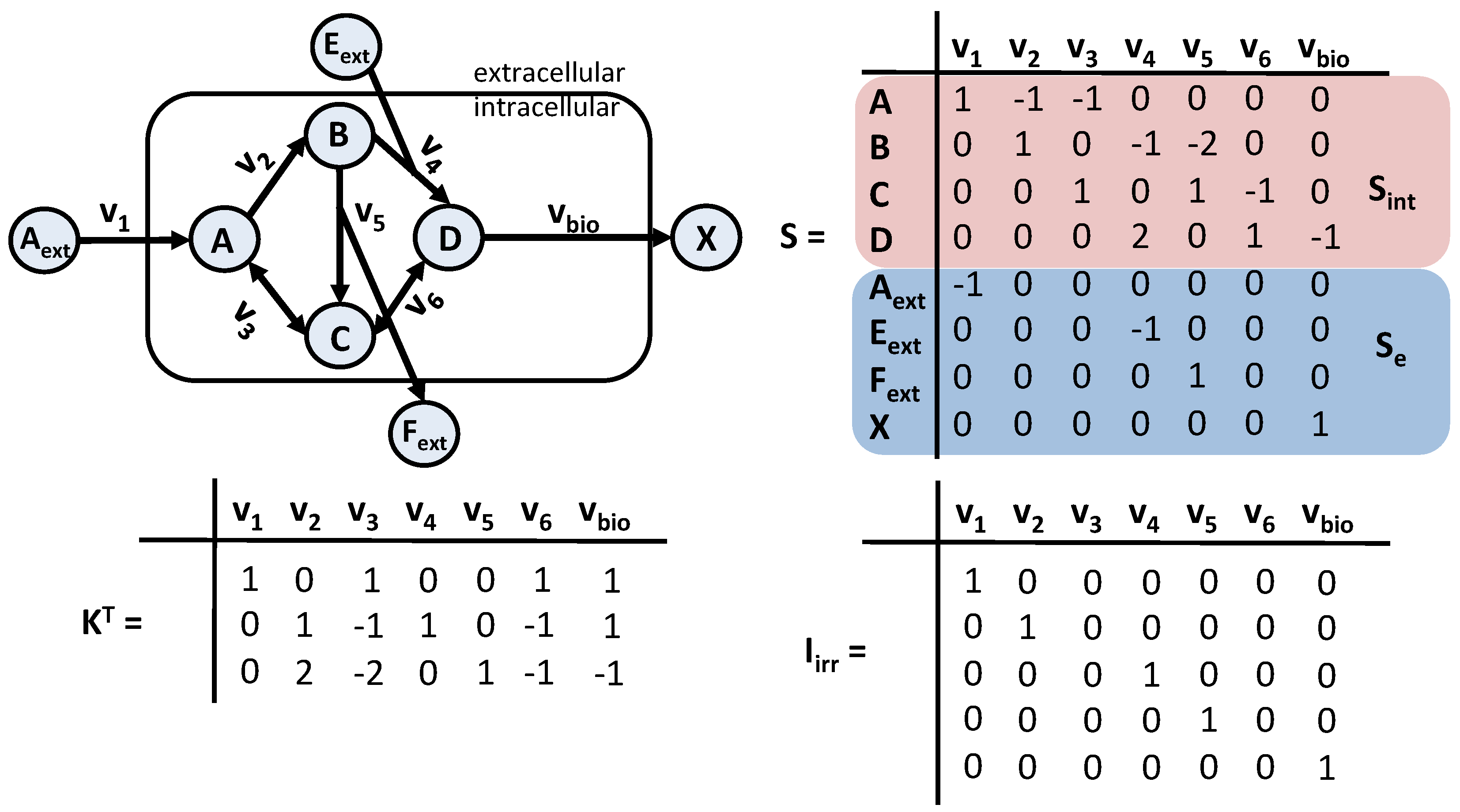
3.1.1. Metabolic Reaction Network
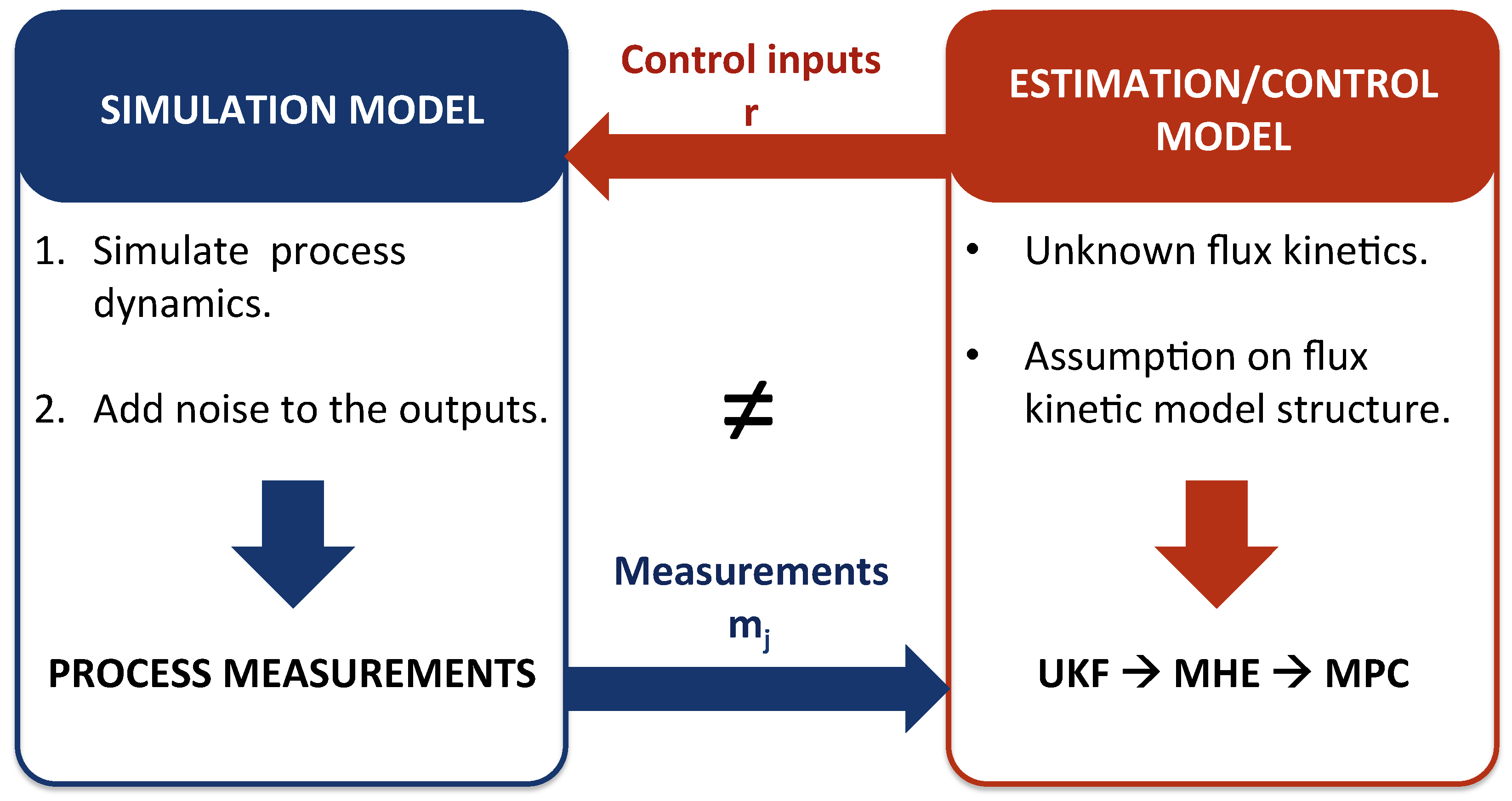
3.1.2. Simulation Flux Model
3.1.3. Estimation/Control Flux Model
3.2. Numerical Results
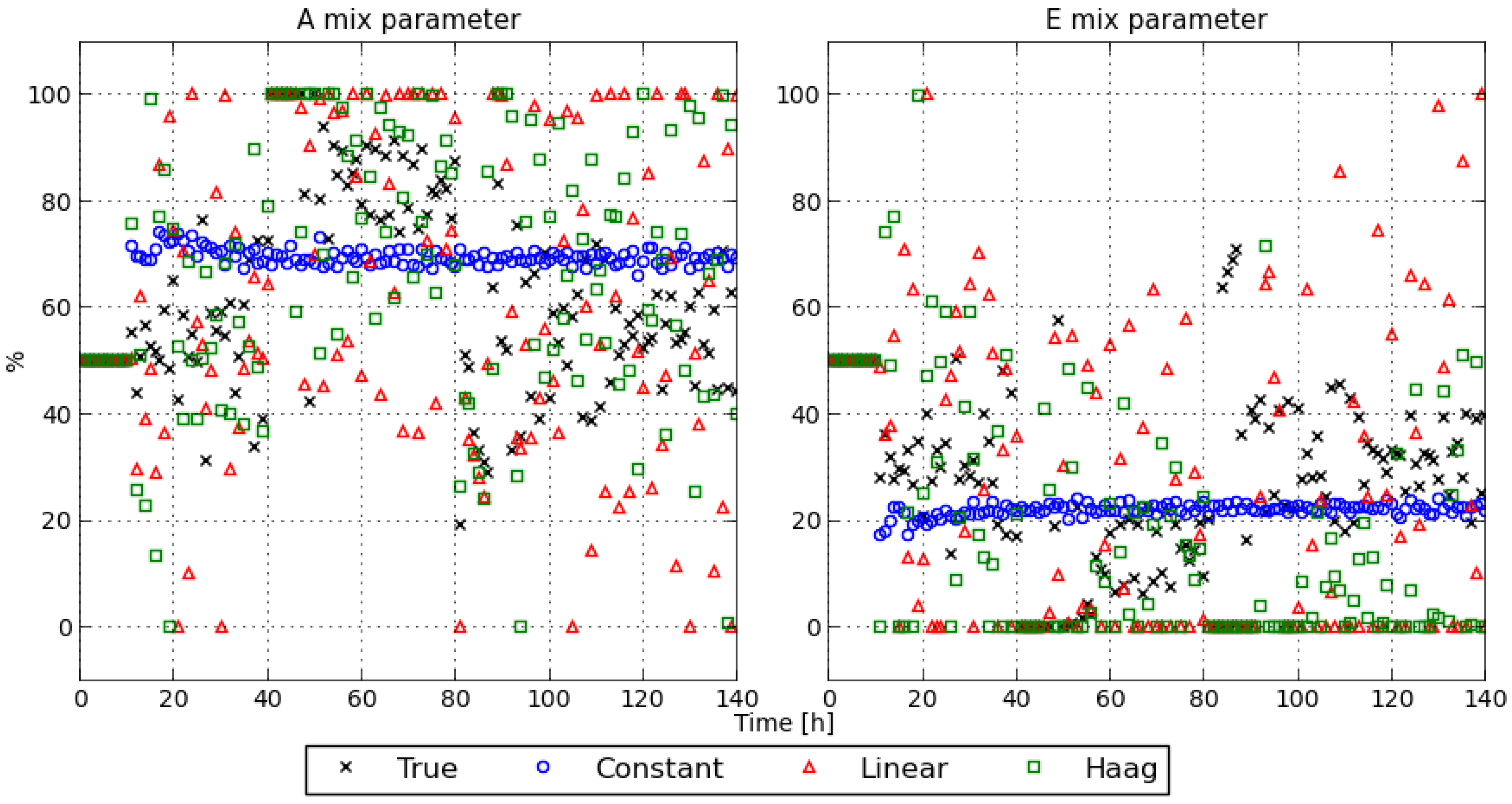
3.2.1. State Estimates
3.2.2. MPC Tracking Performance
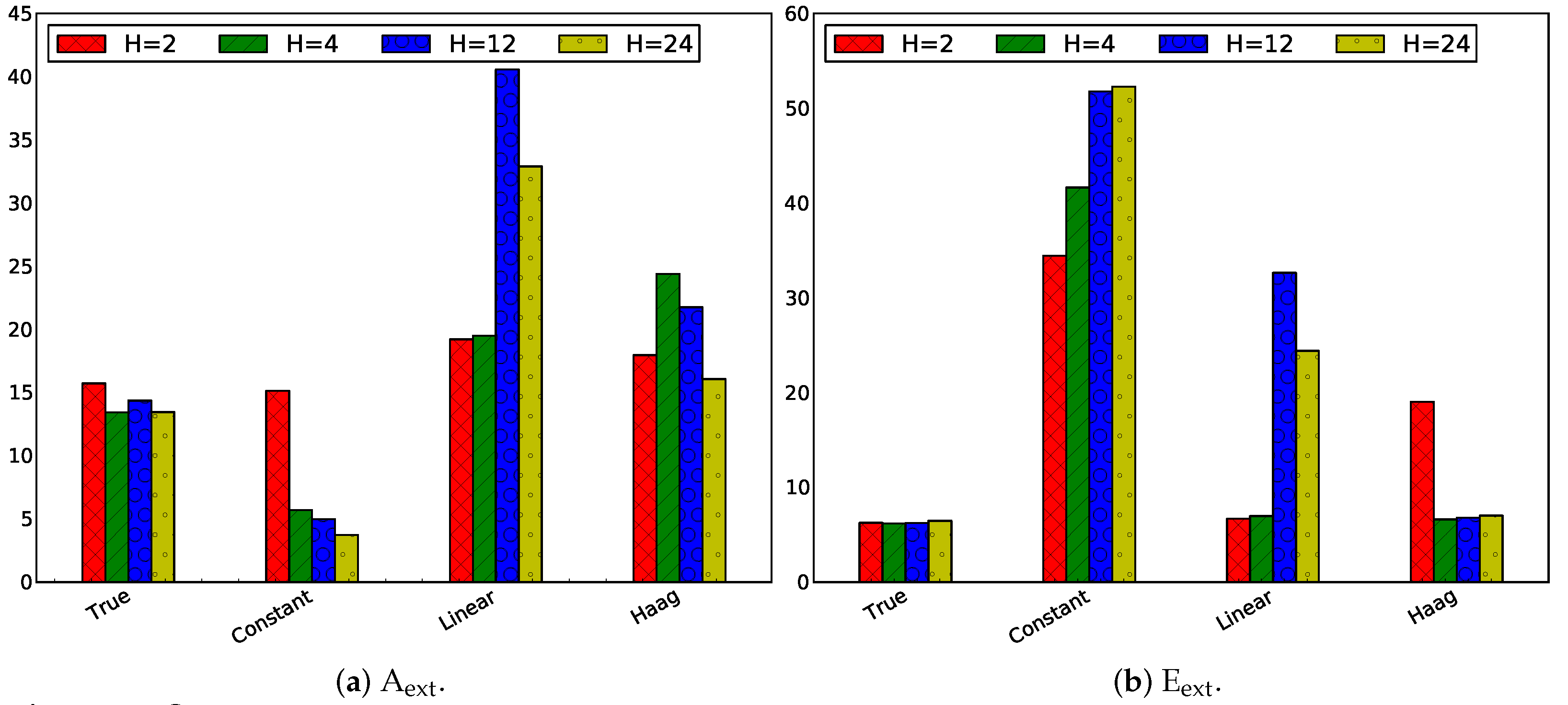
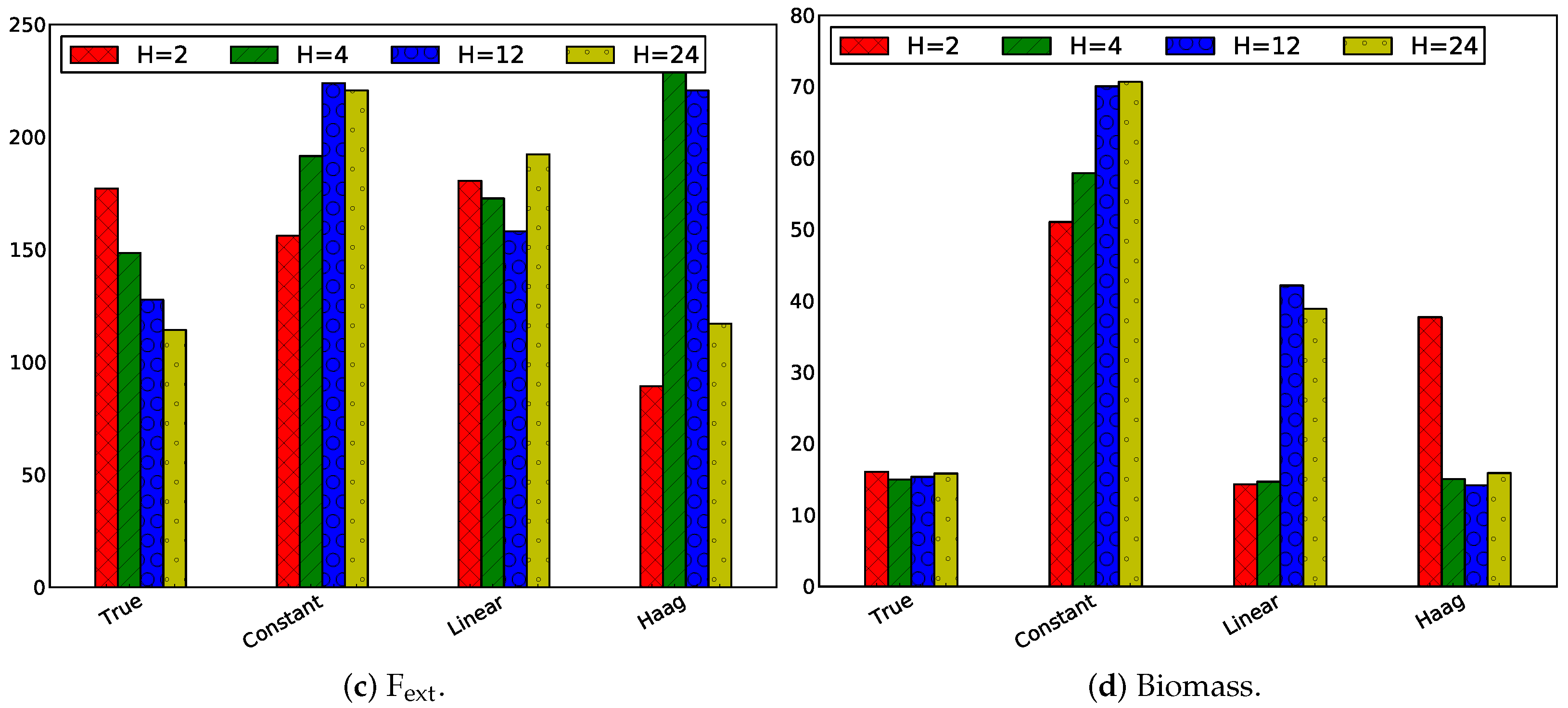
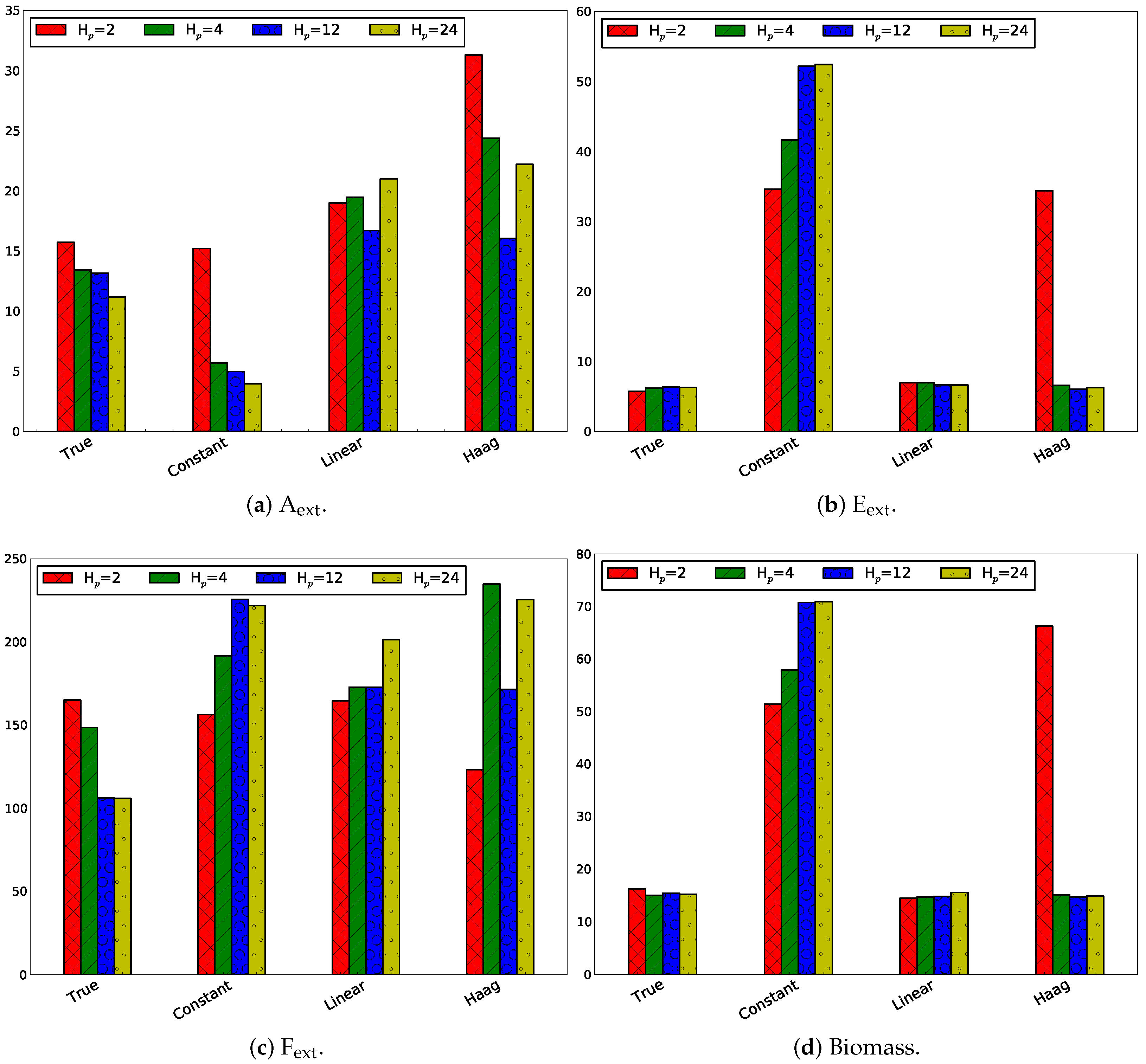
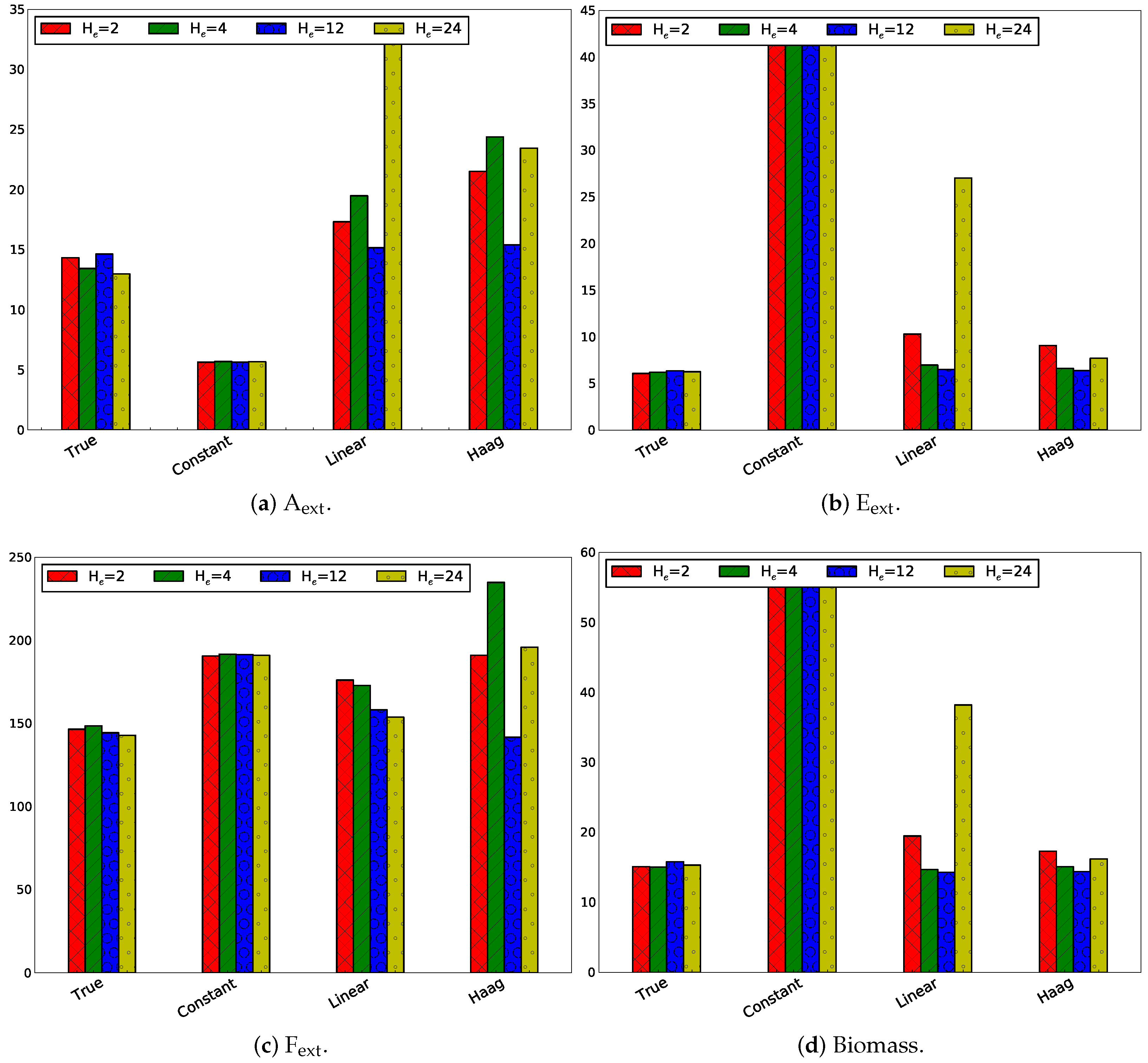
3.2.3. Influence of Horizon Length
Same Horizon and Estimation Horizon
Influence of Only Changing the Prediction Horizon
Influence of Only Changing the Estimation Horizon
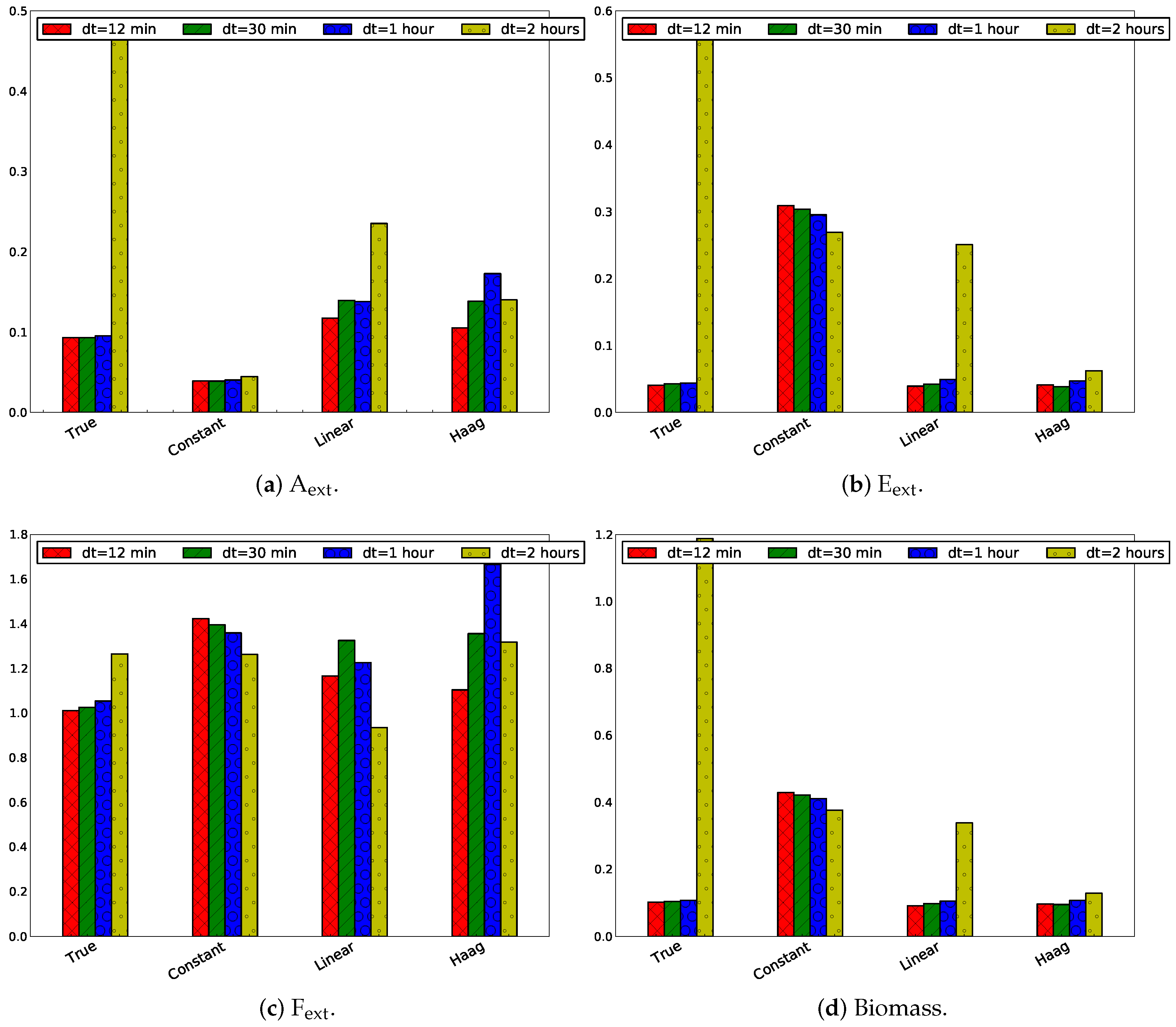
3.2.4. Influence of Sampling Frequency
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
Abbreviations
| CSTR | Continuous stirred tank reactor |
| DMFA | Dynamic metabolic flux analysis |
| MFA | Metabolic flux analysis |
| MHE | Moving horizon estimation |
| MPC | Model predictive control |
References
- Szallasi, Z.; Stelling, J.; Periwal, V. System Modeling in Cell Biology; MIT Press: Cambridge, MA, USA, 2006. [Google Scholar]
- Llaneras, F.; Picó, J. Stoichiometric modelling of cell metabolism. J. Biosci. Bioeng. 2008, 105, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Chang, L.; Liu, X.; Henson, M.A. Nonlinear model predictive control of fed-batch fermentations using dynamic flux balance models. J. Process Control 2016, 42, 137–149. [Google Scholar] [CrossRef]
- Leighty, R.W.; Antoniewicz, M.R. Dynamic metabolic flux analysis (DMFA): A framework for determining fluxes at metabolic non-steady state. Metab. Eng. 2011, 13, 745–755. [Google Scholar] [CrossRef] [PubMed]
- Martínez, V.S.; Buchsteiner, M.; Gray, P.; Nielsen, L.K.; Quek, L.E. Dynamic metabolic flux analysis using B-splines to study the effects of temperature shift on CHO cell metabolism. Metab. Eng. Commun. 2015, 2, 46–57. [Google Scholar] [CrossRef] [PubMed]
- De Sousa, S.F.; Bastin, G.; Jolicoeur, M.; Wouwer, A.V. Dynamic metabolic flux analysis using a convex analysis approach: Application to hybridoma cell cultures in perfusion. Biotechnol. Bioeng. 2015, 113, 1102–1112. [Google Scholar] [CrossRef] [PubMed]
- Vercammen, D.; Logist, F.; Van Impe, J. Online moving horizon estimation of fluxes in metabolic reaction networks. J. Process Control 2016, 37, 1–20. [Google Scholar] [CrossRef] [Green Version]
- Liu, P.; Hua, Y.; Zhang, W.; Xie, T.; Zhuang, Y.; Xia, J.; Noorman, H. A new strategy for dynamic metabolic flux estimation by integrating transient metabolome data into genome-scale metabolic models. Bioprocess Biosyst. Eng. 2021. [Google Scholar] [CrossRef]
- Kalman, R. A New Approach to Linear Filtering and Prediction Problems. Trans. ASME-J. Basic Eng. 1960, 82, 35–45. [Google Scholar] [CrossRef] [Green Version]
- Becerra, V.; Roberts, P.; Griffiths, G. Applying the extended Kalman filter to systems described by nonlinear differential-algebraic equations. Control Eng. Pract. 2001, 9, 267–281. [Google Scholar] [CrossRef]
- Julier, S.; Uhlmann, J. A General Method for Approximating Nonlinear Transformations of Probability Distributions; Technical Report; Robotics Research Group, Department of Engineering Science, University of Oxford: Oxford, UK, 1996. [Google Scholar]
- Robertson, D.G.; Lee, J.H.; Rawlings, J.B. A moving horizon-based approach for least-squares estimation. AIChE J. 1996, 42, 2209–2224. [Google Scholar] [CrossRef]
- Rao, C.V.; Rawlings, J.B. Constrained process monitoring: Moving-horizon approach. AIChE J. 2002, 48, 97–109. [Google Scholar] [CrossRef]
- Kühl, P.; Diehl, M.; Kraus, T.; Schlöder, J.P.; Bock, H.G. A real-time algorithm for moving horizon state and parameter estimation. Comput. Chem. Eng. 2011, 35, 71–83. [Google Scholar] [CrossRef]
- Vercammen, D.; Logist, F.; Van Impe, J. Dynamic estimation of specific fluxes in metabolic networks using non-linear dynamic optimization. BMC Syst. Biol. 2014, 8, 1–22. [Google Scholar] [CrossRef] [PubMed]
- Qu, C.C.; Hahn, J. Computation of arrival cost for moving horizon estimation via unscented Kalman filtering. J. Process Control 2009, 19, 358–363. [Google Scholar] [CrossRef]
- Maußner, J.; Freund, H. Optimization under uncertainty in chemical engineering: Comparative evaluation of unscented transformation methods and cubature rules. Chem. Eng. Sci. 2018, 183, 329–345. [Google Scholar] [CrossRef]
- Rawlings, J. Tutorial overview of model predictive control. IEEE Control Syst. Mag. 2000, 20, 38–52. [Google Scholar]
- Diehl, M.; Bock, H.; Schlöder, J.; Findeisen, R.; Nagy, Z.; Allgöwer, F. Real-time optimization and nonlinear model predictive control of processes governed by differential-algebraic equations. J. Process Control 2002, 12, 577–585. [Google Scholar] [CrossRef]
- Allgöwer, F.; Findeisen, R.; Ebenbauer, C. Nonlinear Model Predictive Control. In UNESCO Encyclopedia of Life Support Systems (EOLSS); EOLSS Publishers Co., Ltd.: Paris, France, 2003; Volume XI. [Google Scholar]
- Bhonsale, S.; Telen, D.; Vercammen, D.; Vallerio, M.; Hufkens, J.; Nimmegeers, P.; Logist, F.; Impe, J.V. Pomodoro: A Novel Toolkit for Dynamic (MultiObjective) Optimization, and Model Based Control and Estimation. IFAC-PapersOnLine 2018, 51, 719–724. [Google Scholar] [CrossRef]
- Andersson, J.; Akesson, J.; Diehl, M. CasADi—A symbolic package for automatic differentiation and optimal control. In Proceedings of the 6th International Conference on Automatic Differentiation, Fort Collins, CO, USA, 23–27 July 2012. [Google Scholar]
- Biegler, L.T. An overview of simultaneous strategies for dynamic optimization. Chem. Eng. Process. Process Intensif. 2007, 46, 1043–1053. [Google Scholar] [CrossRef]
- Wächter, A.; Biegler, L.T. On the implementation of an interior-point filter line-search algorithm for large-scale nonlinear programming. Math. Program. 2006, 106, 25–57. [Google Scholar] [CrossRef]
- Haag, J.E.; Vande Wouwer, A.; Remy, M. A general model of reaction kinetics in biological systems. Bioprocess Biosyst. Eng. 2005, 27, 303–309. [Google Scholar] [CrossRef]
- Duarte, N.C.; Becker, S.A.; Jamshidi, N.; Thiele, I.; Mo, M.L.; Vo, T.D.; Srivas, R.; Palsson, B.O. Global reconstruction of the human metabolic network based on genomic and bibliomic data. Proc. Natl. Acad. Sci. USA 2007, 104, 1777–1782. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leenders, J.; Grootveld, M.; Percival, B.; Gibson, M.; Casanova, F.; Wilson, P.B. Benchtop Low-Frequency 60 MHz NMR Analysis of Urine: A Comparative Metabolomics Investigation. Metabolites 2020, 10, 155. [Google Scholar] [CrossRef] [Green Version]
- Schinn, S.M.; Morrison, C.; Wei, W.; Zhang, L.; Lewis, N.E. A genome-scale metabolic network model and machine learning predict amino acid concentrations in Chinese Hamster Ovary cell cultures. Biotechnol. Bioeng. 2021, 118, 2118–2123. [Google Scholar] [CrossRef] [PubMed]














{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Model Parameter | Numerical Value | |
|---|---|---|
| F | L/h | |
| V | L | |
| mol/L |
| True | Constant | Linear | Haag | |
|---|---|---|---|---|
| A | 13.44 | 5.69 | 19.48 | 24.38 |
| E | 148.52 | 191.61 | 172.80 | 234.86 |
| F | 14.99 | 57.87 | 14.66 | 15.06 |
| Biomass | 6.19 | 41.64 | 6.96 | 6.62 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nimmegeers, P.; Vercammen, D.; Bhonsale, S.; Logist, F.; Van Impe , J. Metabolic Reaction Network-Based Model Predictive Control of Bioprocesses. Appl. Sci. 2021, 11, 9532. https://doi.org/10.3390/app11209532
Nimmegeers P, Vercammen D, Bhonsale S, Logist F, Van Impe J. Metabolic Reaction Network-Based Model Predictive Control of Bioprocesses. Applied Sciences. 2021; 11(20):9532. https://doi.org/10.3390/app11209532
Chicago/Turabian StyleNimmegeers, Philippe, Dominique Vercammen, Satyajeet Bhonsale, Filip Logist, and Jan Van Impe . 2021. "Metabolic Reaction Network-Based Model Predictive Control of Bioprocesses" Applied Sciences 11, no. 20: 9532. https://doi.org/10.3390/app11209532
APA StyleNimmegeers, P., Vercammen, D., Bhonsale, S., Logist, F., & Van Impe , J. (2021). Metabolic Reaction Network-Based Model Predictive Control of Bioprocesses. Applied Sciences, 11(20), 9532. https://doi.org/10.3390/app11209532