
Environmentally Relevant Concentrations of Triclosan Induce Cyto-Genotoxicity and Biochemical Alterations in the Hatchlings of Labeo rohita

 ,
, 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals
2.2. Test Organisms
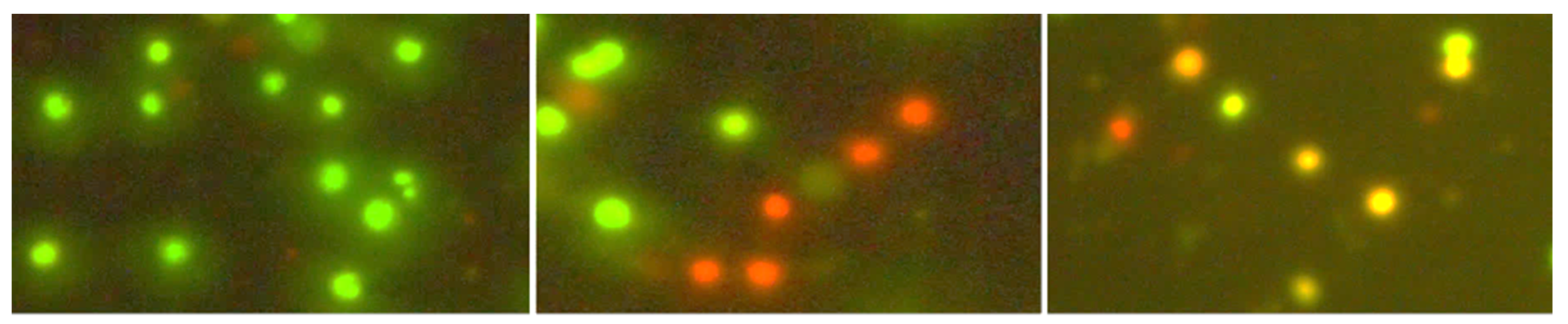
2.3. Cell Viability Test
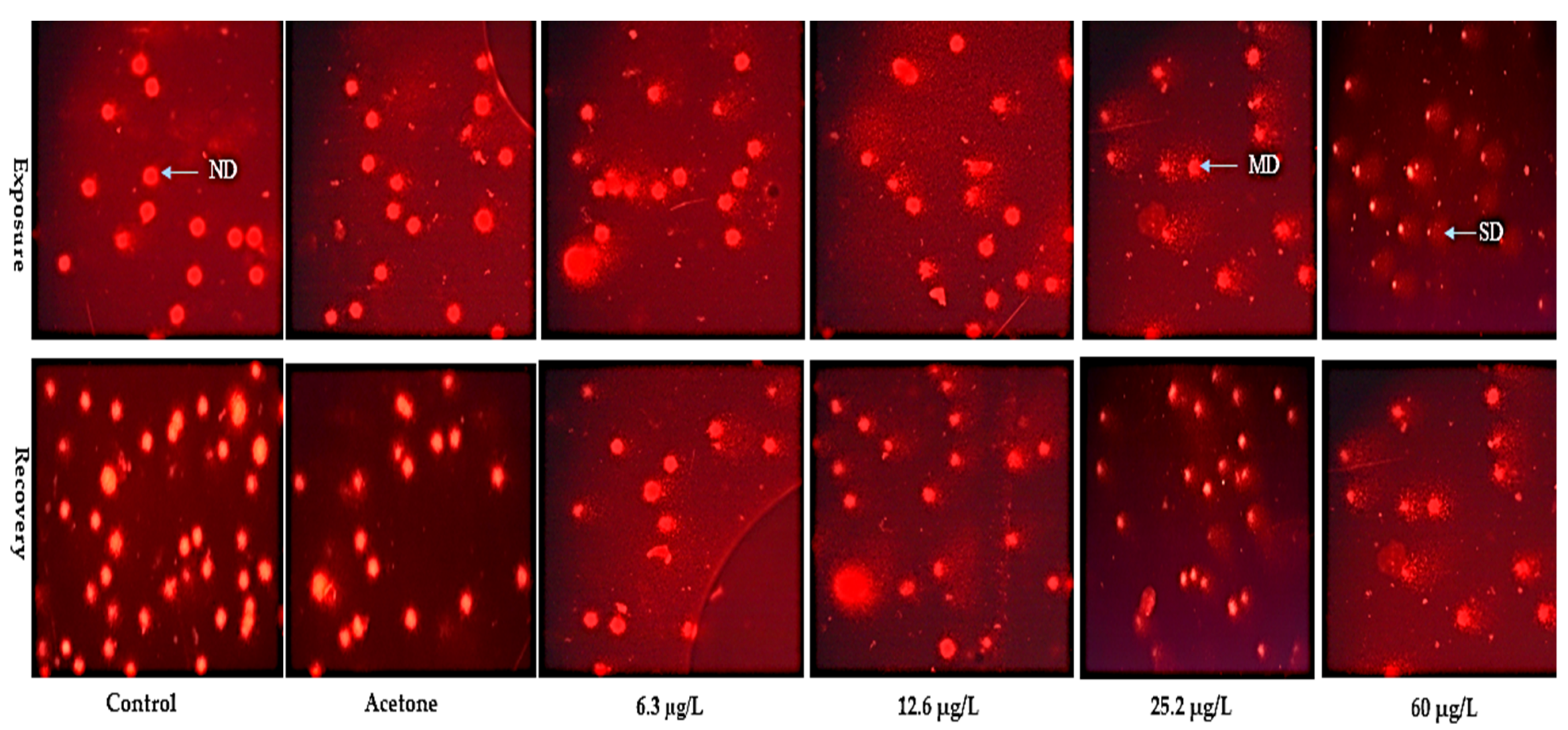
2.4. Micronucleus Assay
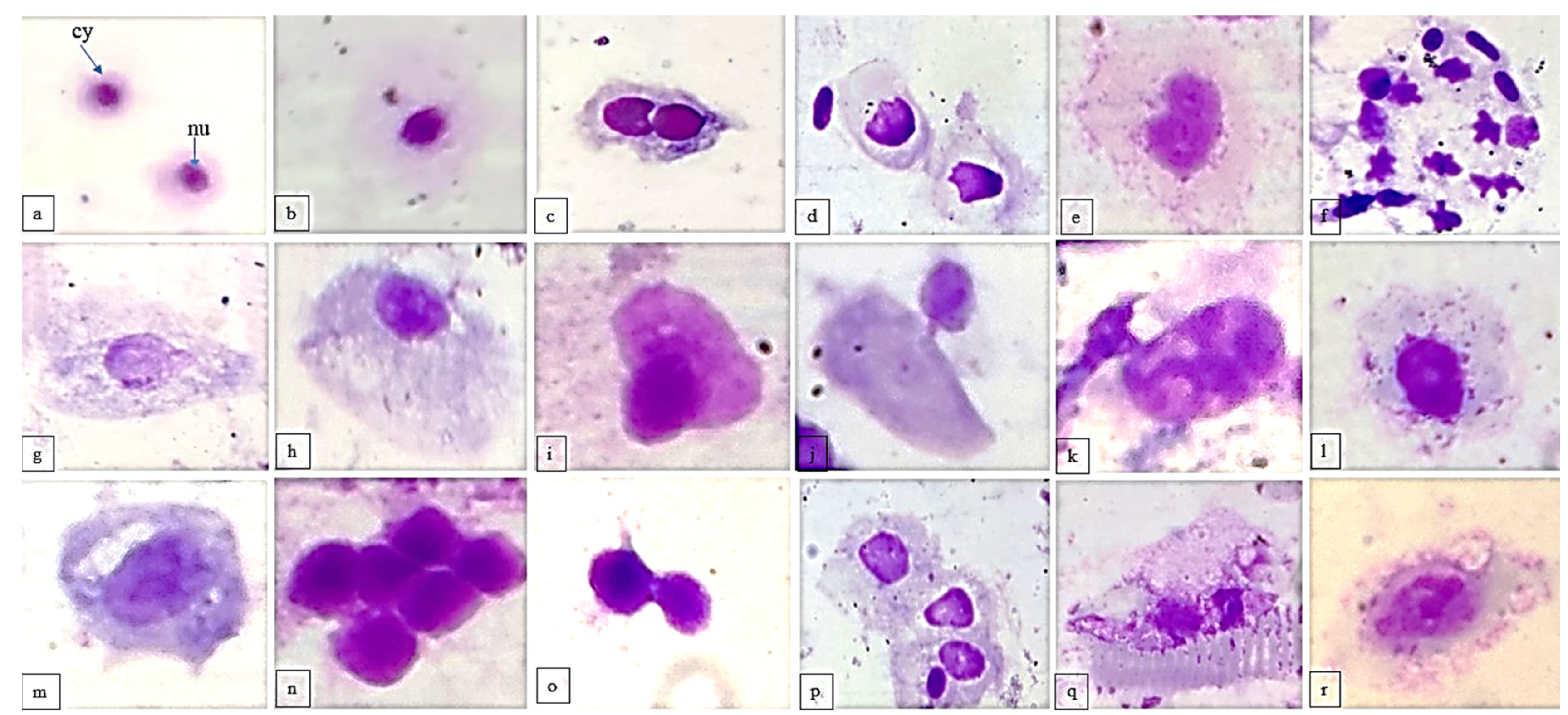
2.5. Comet Assay
2.6. Biochemical Changes
2.7. Statistical Analysis
3. Results
3.1. Cytotoxicity
3.2. Micronucleus Assay
3.3. Comet Assay
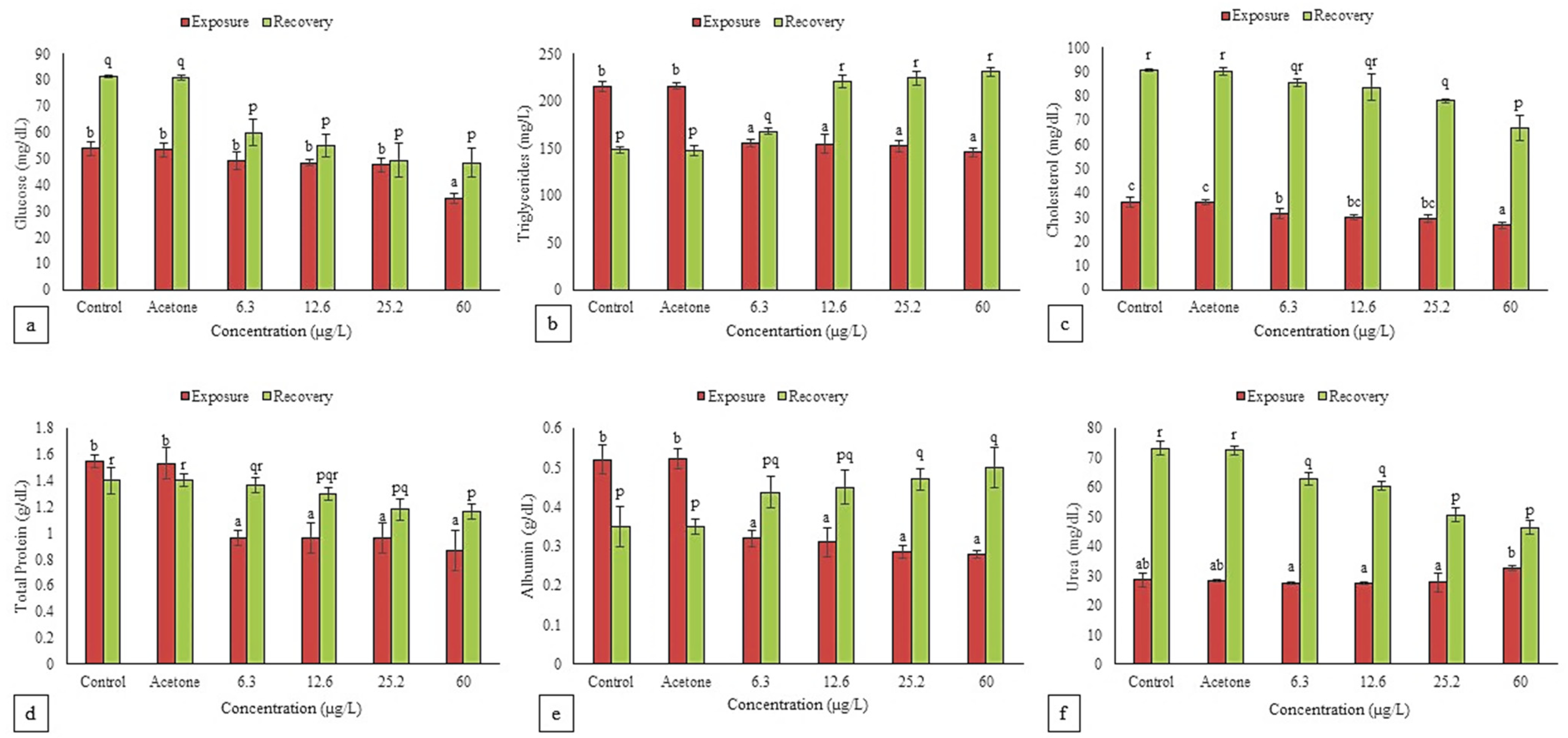
3.4. Biochemical Changes
3.4.1. Glucose
3.4.2. Cholesterol and Triglycerides
3.4.3. Total Protein and Albumin
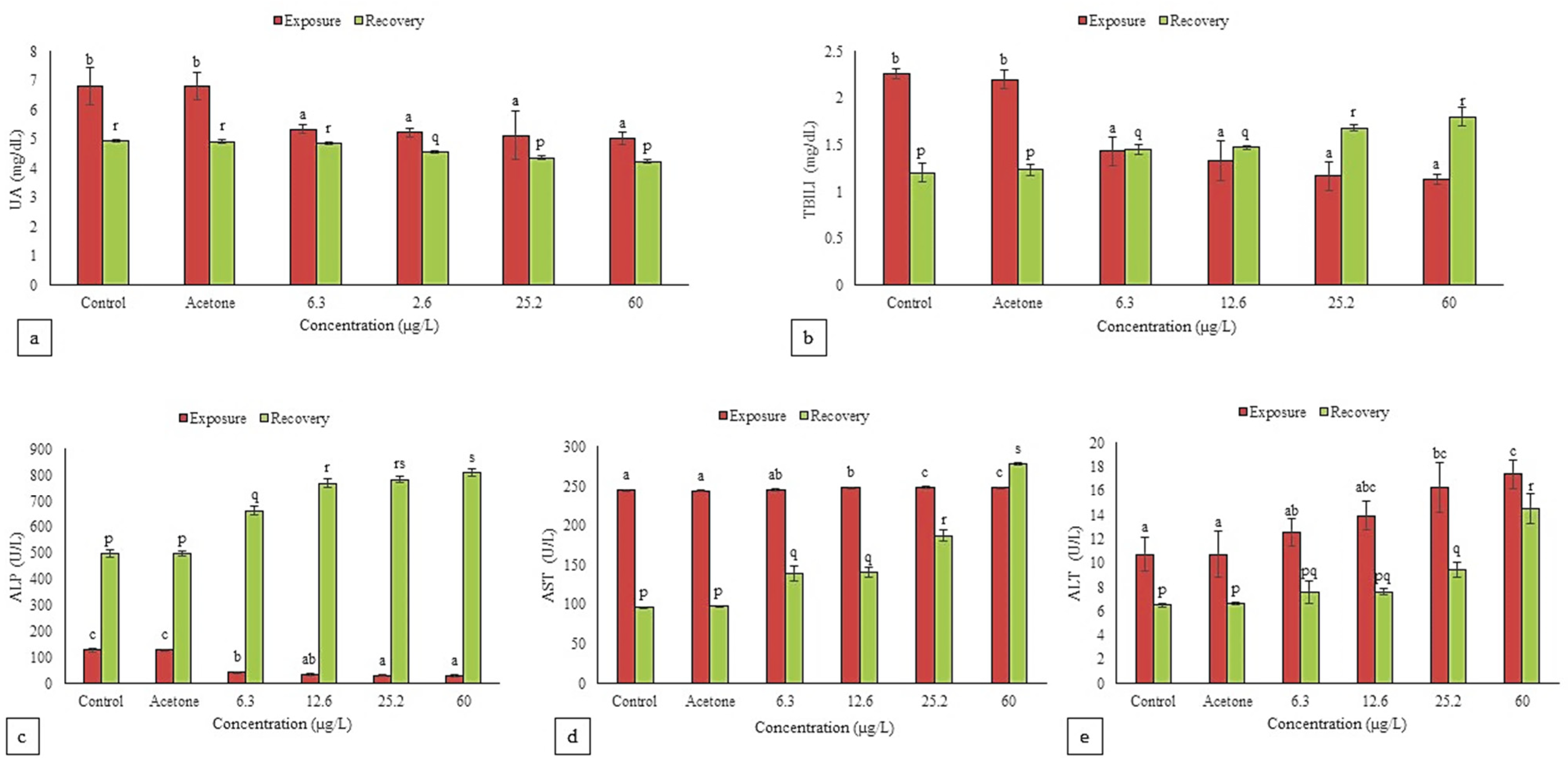
3.4.4. Urea, Uric Acid and Total Bilirubin
3.4.5. ALP, AST and ALT
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Dann, A.B.; Hontela, A. Triclosan: Environmental exposure, toxicity and mechanisms of action. J. Appl. Toxicol. 2011, 31, 285–311. [Google Scholar] [CrossRef]
- Kasprzyk-Hordern, B.; Dinsdale, R.M.; Guwy, A.J. The removal of pharmaceuticals, personal care products, endocrine disruptors and illicit drugs during wastewater treatment and its impact on the quality of receiving waters. Water Res. 2009, 43, 363–380. [Google Scholar] [CrossRef]
- Dhillon, G.S.; Kaur, S.; Pulicharla, R.; Brar, S.K.; Cledón, M.; Verma, M.; Surampalli, R.Y. Triclosan: Current status, occurrence, environmental risks and bioaccumulation potential. Int. J. Environ. Res. Public Health 2015, 12, 5657–5684. [Google Scholar] [CrossRef]
- Ramaswamy, B.R.; Shanmugam, G.; Velu, G.; Rengarajan, B.; Larsson, D.G.J. GC-MS analysis and ecotoxicological risk assessment of triclosan, carbamazepine and parabens in Indian rivers. J. Hazard. Mater. 2011, 186, 1586–1593. [Google Scholar] [CrossRef]
- Nag, S.K.; Das Sarkar, S.; Manna, S.K. Triclosan–an antibacterial compound in water, sediment and fish of River Gomti, India. Int. J. Environ. Health Res. 2018, 28, 461–470. [Google Scholar] [CrossRef]
- Das, S.; Subir, S.; Nag, K.; Kumari, K.; Saha, K.; Bandyopadhyay, S. Occurrence and Safety Evaluation of Antimicrobial Compounds Triclosan and Triclocarban in Water and Fishes of the Multitrophic Niche of River Torsa, India. Arch. Environ. Contam. Toxicol. 2020, 79, 488–499. [Google Scholar] [CrossRef]
- Dar, O.I.; Sharma, S.; Singh, K.; Kaur, A. Teratogenicity and accumulation of triclosan in the early life stages of four food fish during the bioassay. Ecotoxicol. Environ. Saf. 2019, 176, 346–354. [Google Scholar] [CrossRef]
- Sahu, V.K.; Karmakar, S.; Kumar, S.; Shukla, S.P.; Kumar, K. Triclosan toxicity alters behavioral and hematological parameters and vital antioxidant and neurological enzymes in Pangasianodon hypophthalmus (Sauvage, 1878). Aquat. Toxicol. 2018, 202, 145–152. [Google Scholar] [CrossRef]
- Hemalatha, D.; Nataraj, B.; Rangasamy, B.; Shobana, C.; Ramesh, M. DNA damage and physiological responses in an Indian major carp Labeo rohita exposed to an antimicrobial agent triclosan. Fish Physiol. Biochem. 2019, 45, 1463–1484. [Google Scholar] [CrossRef]
- Rauty, S.A.; Angus, R.A. Triclosan has endocrine-disrupting effects in male western mosquitofish, gambusia affinis. Environ. Toxicol. Chem. 2010, 29, 1287–1291. [Google Scholar] [CrossRef]
- Dar, O.I.; Sharma, S.; Singh, K.; Sharma, A.; Bhardwaj, R.; Kaur, A. Biomarkers for the toxicity of sublethal concentrations of triclosan to the early life stages of carps. Sci. Rep. 2020, 10, 17322. [Google Scholar] [CrossRef]
- Capkin, E.; Ozcelep, T.; Kayis, S.; Altinok, I. Antimicrobial agents, triclosan, chloroxylenol, methylisothiazolinone and borax, used in cleaning had genotoxic and histopathologic effects on rainbow trout. Chemosphere 2017, 182, 720–729. [Google Scholar] [CrossRef]
- Sharma, S.; Dar, O.I.; Singh, K.; Thakur, S.; Kesavan, A.K.; Kaur, A. Genomic Markers for the Biological Responses of Triclosan Stressed Hatchlings of Labeo Rohita. ESPR 2021. [Google Scholar] [CrossRef]
- Schweizer, H.P. Triclosan: A widely used biocide and its link to antibiotics. FEMS Microbiol. Lett. 2001, 202, 1–7. [Google Scholar] [CrossRef]
- Binelli, A.; Cogni, D.; Parolini, M.; Riva, C.; Provini, A. In vivo experiments for the evaluation of genotoxic and cytotoxic effects of Triclosan in Zebra mussel hemocytes. Aquat. Toxicol. 2009, 91, 238–244. [Google Scholar] [CrossRef]
- Vijitha, C.K.; Asifa, K.P.; Chitra, K.C. Assessment of genotoxic and haematological consequence of triclosan in the Assessment of genotoxic and haematological consequence of triclosan in the fish, Oreochromis niloticus (Linnaeus, 1758). Int. J. Appl. Res. 2017, 3, 101–109. [Google Scholar]
- Parenti, C.C.; Ghilardi, A.; Della Torre, C.; Mandelli, M.; Magni, S.; Del Giacco, L.; Binelli, A. Environmental concentrations of triclosan activate cellular defence mechanism and generate cytotoxicity on zebrafish (Danio rerio) embryos. Sci. Total Environ. 2019, 650, 1752–1758. [Google Scholar] [CrossRef]
- Paul, T.; Shukla, S.P.; Kumar, K.; Poojary, N.; Kumar, S. Effect of temperature on triclosan toxicity in Pangasianodon hypophthalmus (Sauvage, 1878): Hematology, biochemistry and genotoxicity evaluation. Sci. Total Environ. 2019, 668, 104–114. [Google Scholar] [CrossRef]
- Paul, T.; Kumar, S.; Shukla, S.P.; Pal, P.; Kumar, K.; Poojary, N.; Biswal, A.; Mishra, A. A multi-biomarker approach using integrated biomarker response to assess the effect of pH on triclosan toxicity in Pangasianodon hypophthalmus (Sauvage, 1878). Environ. Pollut. 2020, 260, 114001. [Google Scholar] [CrossRef]
- Gyimah, E.; Dong, X.; Qiu, W.; Zhang, Z.; Xu, H. Sublethal concentrations of triclosan elicited oxidative stress, DNA damage, and histological alterations in the liver and brain of adult zebrafish. Environ. Sci. Pollut. Res. 2020, 27, 17329–17338. [Google Scholar] [CrossRef]
- Agrahari, S.; Pandey, K.C.; Gopal, K. Biochemical alteration induced by monocrotophos in the blood plasma of fish, Channa punctatus (Bloch). Pestic. Biochem. Physiol. 2007, 88, 268–272. [Google Scholar] [CrossRef]
- Prokić, M.D.; Gavrilović, B.R.; Radovanović, T.B.; Gavrić, J.P.; Petrović, T.G.; Despotović, S.G.; Faggio, C. Studying microplastics: Lessons from evaluated literature on animal model organisms and experimental approaches. J. Hazard. Mater. 2021, 414, 125476. [Google Scholar] [CrossRef]
- Sinha, R.; Jindal, R.; Faggio, C. Protective effect of emblica officinalis in cyprinus carpio against hepatotoxicity induced by malachite green: Ultrastructural and molecular analysis. Appl. Sci. 2021, 11, 3507. [Google Scholar] [CrossRef]
- Vajargah, M.F.; Mohsenpour, R.; Yalsuyi, A.M.; Galangash, M.M.; Faggio, C. Evaluation of Histopathological Effect of Roach (Rutilus rutilus caspicus) in Exposure to Sub-Lethal Concentrations of Abamectin. Water. Air. Soil Pollut. 2021, 232, 1–8. [Google Scholar] [CrossRef]
- Canli, E.G.; Dogan, A.; Canli, M. Serum biomarker levels alter following nanoparticle (Al2O3, CuO, TiO2) exposures in freshwater fish (Oreochromis niloticus). Environ. Toxicol. Pharmacol. 2018, 62, 181–187. [Google Scholar] [CrossRef]
- Sethuraman, S.; Kumar, T.R.; Vivekananthan, T. Triclosan Induced Alterations of Biochemical Parameters in Zebra Fish Brachydaniorerio (Ham.) and their Productive Role by Dietary Supplementation of Garlic Extract and L-Ascorbic acid (Vitamin C). Asian J. Pharmacol. Toxicol. 2015, 03, 5–12. [Google Scholar]
- Kola, R.K.; Mohd, A.R.; Yalavarthy, P.D. Research Article Effect of Triclosan on Total Protein Content in Fresh Water Fish, Channa Punctatus. Int. J. Recent Sci. Res. 2015, 6, 4641–4643. [Google Scholar]
- Hemalatha, D.; Rangasamy, B.; Nataraj, B.; Ramesh, M. Assessment of triclosan impact on enzymatic biomarkers in an Indian major carp, Catla catla. J. Basic Appl. Zool. 2019, 80, 1–8. [Google Scholar] [CrossRef]
- Sharma, S.; Dar, O.I.; Singh, K.; Kaur, A.; Faggio, C. Triclosan elicited biochemical and transcriptomic alterations in Labeo rohita larvae. Environ. Toxicol. Pharmacol. 2021, 88, 103748. [Google Scholar] [CrossRef]
- Cahova, J.; Blahova, J.; Plhalova, L.; Svobodova, Z.; Faggio, C. Do Single-Component and Mixtures Selected Organic UV Filters Induce Embryotoxic Effects in Zebrafish (Danio rerio)? Water 2021, 13, 2203. [Google Scholar] [CrossRef]
- Er, A.; Uçkan, F.; Rivers, D.B.; Sak, O. Cytotoxic effects of parasitism and application of venom from the endoparasitoid Pimpla turionellae on hemocytes of the host Galleria mellonella. J. Appl. Entomol. 2011, 135, 225–236. [Google Scholar] [CrossRef]
- Sharma, M.; Chadha, P. Widely used non-ionic surfactant 4-nonylphenol: Showing genotoxic effects in various tissues of Channa punctatus. Environ. Sci. Pollut. Res. 2017, 24, 11331–11339. [Google Scholar] [CrossRef]
- Singh, N.P.; McCoy, M.T.; Tice, R.R.; Schneider, E.L. A simple technique for quantitation of low levels of DNA damage in individual cells. Exp. Cell Res. 1988, 175, 184–191. [Google Scholar] [CrossRef] [Green Version]
- Freitas, R.; Silvestro, S.; Coppola, F.; Meucci, V.; Battaglia, F.; Intorre, L.; Soares, A.M.V.M.; Pretti, C.; Faggio, C. Combined effects of salinity changes and salicylic acid exposure in Mytilus galloprovincialis. Sci. Total Environ. 2020, 715, 136804. [Google Scholar] [CrossRef]
- Aranami, K.; Readman, J.W. Photolytic degradation of triclosan in freshwater and seawater. Chemosphere 2007, 66, 1052–1056. [Google Scholar] [CrossRef]
- Martin, S.J.; Green, D.R.; Cotter, T.G. Dicing with death: Dissecting the components of the apoptosis machinery. Trends Biochem. Sci. 1994, 19, 26–30. [Google Scholar] [CrossRef]
- Steinert, S.A. Contribution of apoptosis to observed DNA damage in mussel cells. Mar. Environ. Res. 1996, 42, 253–259. [Google Scholar] [CrossRef]
- Park, B.K.; Gonzales, E.L.T.; Yang, S.M.; Bang, M.; Choi, C.S.; Shin, C.Y. Effects of triclosan on neural stem cell viability and survival. Biomol. Ther. 2016, 24, 99–107. [Google Scholar] [CrossRef] [Green Version]
- Szychowski, K.A.; Sitarz, A.M.; Wojtowicz, A.K. Triclosan induces Fas receptor-dependent apoptosis in mouse neocortical neurons in vitro. Neuroscience 2015, 284, 192–201. [Google Scholar] [CrossRef]
- Guidony, N.S.; Scaini, J.L.R.; Oliveira, M.W.B.; Machado, K.S.; Bastos, C.; Escarrone, A.L.; Souza, M.M. ABC proteins activity and cytotoxicity in zebrafish hepatocytes exposed to triclosan. Environ. Pollut. 2020, 116368. [Google Scholar] [CrossRef]
- Pernot, E.; Hall, J.; Baatout, S.; Benotmane, M.A.; Blanchardon, E.; Bouffler, S.; El Saghire, H.; Gomolka, M.; Guertler, A.; Harms-Ringdahl, M.; et al. Ionizing radiation biomarkers for potential use in epidemiological studies. Mutat. Res.—Rev. Mutat. Res. 2012, 751, 258–286. [Google Scholar] [CrossRef]
- Sabharwal, R.; Verma, P.; Syed, M.A.; Sharma, T.; Subudhi, S.K.; Mohanty, S.; Gupta, S. Emergence of micronuclei as a genomic biomarker. Indian J. Med. Paediatr. Oncol. 2015, 36, 212–218. [Google Scholar] [CrossRef] [Green Version]
- Rodilla, V. Origin and evolution of binucleated cells and binucleated cells with micronuclei in cisplatin-treated CHO cultures. Mutat. Res. Toxicol. 1993, 300, 281–291. [Google Scholar] [CrossRef]
- Nwani, C.D.; Lakra, W.S.; Nagpure, N.S.; Kumar, R.; Kushwaha, B.; Srivastava, S.K. Mutagenic and genotoxic effects of carbosulfan in freshwater fish Channa punctatus (Bloch) using micronucleus assay and alkaline single-cell gel electrophoresis. Food Chem. Toxicol. 2010, 48, 202–208. [Google Scholar] [CrossRef]
- Emery, D. Assessing the Genotoxicity of Triclosan in Tadpoles of the American Bullfrog, Lithobates Catesbeianus. 2012, p. 10. Available online: https://doi.org/10.25772/H46E-PZ53 (accessed on 31 August 2021).
- Wang, F.; Xu, R.; Zheng, F.; Liu, H. Effects of triclosan on acute toxicity, genetic toxicity and oxidative stress in goldfish (Carassius auratus). Exp. Anim. 2018, 67, 219–227. [Google Scholar] [CrossRef] [Green Version]
- Jimoh, R.O. Sublethal and environmentally relevant concentrations of triclosan and triclocarban induce histological, genotoxic, and embryotoxic effects in Clarias gariepinus (Burchell, 1822). Environ. Sci. Pollut. Res. 2021, 28, 31071–31083. [Google Scholar] [CrossRef]
- Falisse, E.; Ducos, B.; Stockwell, P.A.; Morison, I.M.; Chatterjee, A.; Silvestre, F. DNA methylation and gene expression alterations in zebrafish early-life stages exposed to the antibacterial agent triclosan. Environ. Pollut. 2018, 243, 1867–1877. [Google Scholar] [CrossRef]
- Gao, L.; Yuan, T.; Cheng, P.; Bai, Q.; Zhou, C.; Ao, J.; Wang, W.; Zhang, H. Effects of triclosan and triclocarban on the growth inhibition, cell viability, genotoxicity and multixenobiotic resistance responses of Tetrahymena thermophila. Chemosphere 2015, 139, 434–440. [Google Scholar] [CrossRef]
- Lin, D.; Zhou, Q.; Xie, X.; Liu, Y. Potential biochemical and genetic toxicity of triclosan as an emerging pollutant on earthworms (Eisenia fetida). Chemosphere 2010, 81, 1328–1333. [Google Scholar] [CrossRef]
- Lin, D.; Li, Y.; Zhou, Q.; Xu, Y.; Wang, D. Effect of triclosan on reproduction, DNA damage and heat shock protein gene expression of the earthworm Eisenia fetida. Ecotoxicology 2014, 23, 1826–1832. [Google Scholar] [CrossRef]
- Jeon, J.; Lim, H.K.; Kannan, K.; Kim, S.D. Effect of perfluorooctanesulfonate on osmoregulation in marine fish, Sebastes schlegeli, under different salinities. Chemosphere 2010, 81, 228–234. [Google Scholar] [CrossRef]
- Bharti, S.; Rasool, F. Analysis of the biochemical and histopathological impact of a mild dose of commercial malathion on Channa punctatus (Bloch) fish. Toxicol. Rep. 2021, 8, 443–455. [Google Scholar] [CrossRef]
- Van Meer, G.; Voelker, D.R.; Feigenson, G.W. Membrane lipids: Where they are and how they behave. Nat. Rev. Mol. Cell Biol. 2008, 9, 112–124. [Google Scholar] [CrossRef]
- Hatami, M.; Banaee, M.; Nematdoost Haghi, B. Sub-lethal toxicity of chlorpyrifos alone and in combination with polyethylene glycol to common carp (Cyprinus carpio). Chemosphere 2019, 219, 981–988. [Google Scholar] [CrossRef]
- Chai, L.; Chen, A.; Luo, P.; Zhao, H.; Wang, H. Histopathological changes and lipid metabolism in the liver of Bufo gargarizans tadpoles exposed to Triclosan. Chemosphere 2017, 182, 255–266. [Google Scholar] [CrossRef]
- Santos, C.R.; Schulze, A. Lipid metabolism in cancer. FEBS J. 2012, 279, 2610–2623. [Google Scholar] [CrossRef]
- Yang, J.L.; Chen, H.C. Effects of gallium on common carp (Cyprinus carpio): Acute test, serum biochemistry, and erythrocyte morphology. Chemosphere 2003, 53, 877–882. [Google Scholar] [CrossRef]
- Banaee, M.; Akhlaghi, M.; Soltanian, S.; Sureda, A.; Gholamhosseini, A.; Rakhshaninejad, M. Combined effects of exposure to sub-lethal concentration of the insecticide chlorpyrifos and the herbicide glyphosate on the biochemical changes in the freshwater crayfish Pontastacus leptodactylus. Ecotoxicology 2020, 29, 1500–1515. [Google Scholar] [CrossRef]
- Banaee, M.; Nemadoost Haghi, B.; Tahery, S.; Shahafve, S.; Vaziriyan, M. Effects of Sub-Lethal Toxicity of Paraquat on Blood Biochemical Parameters of Common Carp, Cyprinus carpio (Linnaeus, 1758). Iran. J. Toxicol. 2016, 10, 1–5. [Google Scholar] [CrossRef] [Green Version]
- Onck, P. Protein Mechanics: From Amino Acid to Swimming Cells. Procedia IUTAM 2017, 20, 73–80. [Google Scholar] [CrossRef]
- Deepika, S.; Padmavathy, P.; Srinivasan, A.; Sugumar, G.; Jawahar, P. Effect of triclosan (TCS) on the protein content and associated histological changes on tilapia, Oreochromis mossambicus (Peters, 1852). Environ. Sci. Pollut. Res. 2021, 1–9. [Google Scholar] [CrossRef]
- Ao, J.; Gao, L.; Yuan, T.; Jiang, G. Interaction mechanisms between organic UV filters and bovine serum albumin as determined by comprehensive spectroscopy exploration and molecular docking. Chemosphere 2015, 119, 590–600. [Google Scholar] [CrossRef]
- Krishna, K.; Kumar, S.; Paul, T.; Pani, K.; Shukla, S.P. Triclosan induces immunosuppression and reduces survivability of striped cat fi sh Pangasianodon hypophthalmus during the challenge to a fi sh pathogenic bacterium Edwardsiella tarda. Environ. Res. 2020, 186, 109575. [Google Scholar] [CrossRef]
- Niemelä, O.; Alatalo, P. Biomarkers of alcohol consumption and related liver disease. Scand. J. Clin. Lab. Investig. 2010, 70, 305–312. [Google Scholar] [CrossRef]
- Miri, M.; Rahdari, A. Sub lethal effects of copper oxide (CuO) nanoparticles on blood parameters of common carp (Cyprinus Carpio). J. Biodivers. Environ. Sci. 2015, 6, 283–291. [Google Scholar]
- Hamed, H.S.; Abdel-Tawwab, M. Dietary pomegranate (Punica granatum) peel mitigated the adverse effects of silver nanoparticles on the performance, haemato-biochemical, antioxidant, and immune responses of Nile tilapia fingerlings. Aquaculture 2021, 540, 736742. [Google Scholar] [CrossRef]
- Dar, O.I.; Sharma, S.; Singh, K.; Sharma, A.; Bhardwaj, R.; Kaur, A. Biochemical markers for prolongation of the acute stress of triclosan in the early life stages of four food fishes. Chemosphere 2020, 247, 125914. [Google Scholar] [CrossRef]
- Giang, P.T.; Burkina, V.; Sakalli, S.; Schmidt-Posthaus, H.; Rasmussen, M.K.; Randak, T.; Grabic, R.; Grabicova, K.; Fedorova, G.; Koba, O.; et al. Effects of Multi-Component Mixtures from Sewage Treatment Plant Effluent on Common Carp (Cyprinus carpio) under Fully Realistic Condition. Environ. Manag. 2019, 63, 466–484. [Google Scholar] [CrossRef]
- Gallego, R.S.E.; Peñuela, G.A.; Martínez-López, E. Enzymatic activity changes in striped catfish Pseudoplatystoma magdaleniatum, induced by exposure to different concentrations of ibuprofen and triclosan. Chemosphere 2021, 271, 129399. [Google Scholar] [CrossRef]
- Elia, A.C.; Pacini, N.; Fioravanti, M.L.; Dörr, A.J.M.; Zaccaroni, A.; Parmeggiani, A.M.; Gustinelli, A.; Mordenti, O.; Abete, M.C.; Prearo, M. Assessment of detoxifying markers for florfenicol in Rainbow Trout Liver. J. Aquat. Anim. Health 2016, 28, 258–265. [Google Scholar] [CrossRef]
- Lakra, K.C.; Banerjee, T.K.; Lal, B. Coal mine effluent-induced metal bioaccumulation, biochemical, oxidative stress, metallothionein, and histopathological alterations in vital tissues of the catfish, Clarias batrachus. Environ. Sci. Pollut. Res. 2021, 28, 25300–25315. [Google Scholar] [CrossRef]
- Banaee, M.; Sureda, A.; Mirvaghefi, A.R.; Rafei, G.R. Effects of long-term silymarin oral supplementation on the blood biochemical profile of rainbow trout (Oncorhynchus mykiss). Fish Physiol. Biochem. 2011, 37, 885–896. [Google Scholar] [CrossRef]
- Banaee, M. Physiological Dysfunction in Fish After Insecticides Exposure. Insectic.-Dev. Safer More Eff. Technol. 2013, 103–142. [Google Scholar] [CrossRef] [Green Version]
- Adamu, K.M.; Kori-Siakpere, O. Effects of sublethal concentrations of tobacco (Nicotiana tobaccum) leaf dust on some biochemical parameters of Hybrid catfish (Clarias gariepinus and Heterobranchus bidorsalis). Braz. Arch. Biol. Technol. 2011, 54, 183–196. [Google Scholar] [CrossRef]
- Jee, J.H.; Masroor, F.; Kang, J.C. Responses of cypermethrin-induced stress in haematological parameters of Korean rockfish, Sebastes schlegeli (Hilgendorf). Aquac. Res. 2005, 36, 898–905. [Google Scholar] [CrossRef]
- Jyothi, B.; Narayan, G. Certain pesticide-induced carbohydrate metabolic disorders in the serum of freshwater fish Clarias batrachus (Linn.). Food Chem. Toxicol. 1999, 37, 417–421. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment | Viable Cells (%) | Necrotic Cells (%) | Apoptotic Cells (%) | |
|---|---|---|---|---|
| Control | E | 90.53 ± 0.11 d | 7.00 ± 0.53 a | 2.47 ± 0.42 a |
| R | 89.07 ± 0.64 r | 6.87 ± 0.42 p | 4.07 ± 0.23 p | |
| Acetone | E | 90.67 ± 0.90 d | 7.13 ± 0.50 a | 2.20 ± 0.40 a |
| R | 89.00 ± 0.40 r | 7.07 ± 0.30 p | 3.93 ± 0.42 p | |
| 6.3 µg/L | E | 86.07 ± 1.51 c | 9.40 ± 1.11 ab | 4.53 ± 0.50 b |
| R | 87.80 ± 0.35 r | 7.53 ± 0.50 pq | 4.67 ± 0.30 p | |
| 12.6 µg/L | E | 84.80 ± 0.60 c | 10.20 ± 0.40 b | 5.00 ± 0.53 b |
| R | 82.87 ± 1.94 q | 8.80 ± 1.44 pq | 8.33 ± 0.50 q | |
| 25.2 µg/L | E | 77.47 ± 1.92 b | 15.13 ± 1.40 c | 7.40 ± 0.53 c |
| R | 81.33 ± 2.05 q | 10.13 ± 1.55 q | 8.53 ± 0.50 q | |
| 60 µg/L | E | 74.40 ± 1.83 a | 17.60 ± 1.44 d | 8.00 ± 0.40 c |
| R | 77.80 ± 1.05 p | 13.13 ± 1.33 r | 9.07 ± 0.30 q | |
| Exposure | F | 78.77 ** | 57.49 ** | 74.47 ** |
| Recovery | F | 40.21 ** | 15.25 ** | 117.10 ** |
| Treatment | MN (%) | NCA (%) | |
|---|---|---|---|
| Control | E | 0.33 ± 0.11 a | 1.27 ± 0.30 a |
| R | 0.47 ± 0.11 p | 1.60 ± 0.20 p | |
| Acetone | E | 0.33 ± 0.11 a | 1.20 ± 0.20 a |
| R | 0.40 ± 0.20 p | 1.47 ± 0.30 p | |
| 6.3 µg/L | E | 0.80 ± 0.20 ab | 1.80 ± 0.20 ab |
| R | 1.27 ± 0.30 q | 3.53 ± 0.30 q | |
| 12.6 µg/L | E | 1.13 ± 0.30 bc | 2.67 ± 0.30 b |
| R | 1.93 ± 0.30 qr | 5.87 ± 0.50 r | |
| 25.2 µg/L | E | 1.53 ± 0.30 cd | 4.13 ± 0.50 c |
| R | 2.30 ± 0.30 r | 6.73 ± 0.42 r | |
| 60 µg/L | E | 2.13 ± 0.30 d | 5.67 ± 0.50 d |
| R | 3.46 ± 0.41 s | 9.27 ± 0.42 s | |
| Exposure | F | 26.00 ** | 74.13 ** |
| Recovery | F | 49.43 ** | 208 ** |
| Treatment | TL (µm) | TM | OTM | TDNA (%) | |
|---|---|---|---|---|---|
| Control | E | 22.36 ± 0.58 a | 1.82 ± 0.08 a | 3.53 ± 0.22 a | 7.26 ± 0.17 a |
| R | 21.71 ± 0.61 p | 1.90 ± 0.12 p | 3.39 ± 0.04 p | 7.68 ± 0.20 p | |
| Acetone | E | 23.16 ± 0.67 a | 1.90 ± 0.07 a | 3.76 ± 0.13 a | 7.39 ± 0.03 a |
| R | 21.68 ± 0.42 p | 1.85 ± 0.12 p | 3.43 ± 0.31 p | 7.56 ± 0.25 p | |
| 6.3 µg/L | E | 29.17 ± 0.34 b | 2.64 ± 0.13 b | 4.48 ± 0.09 b | 8.37 ± 0.10 b |
| R | 23.20 ± 0.56 pq | 1.91 ± 0.04 p | 3.44 ± 0.16 p | 7.77 ± 0.35 p | |
| 12.6 µg/L | E | 30.70 ± 1.14 b | 3.20 ± 0.11 c | 4.98 ± 0.14 c | 9.50 ± 0.31 c |
| R | 24.87 ± 1.11 qr | 2.37 ± 0.15 pq | 3.79 ± 0.31 p | 8.64 ± 0.42 p | |
| 25.2 µg/L | E | 35.36 ± 0.60 c | 4.18 ± 0.25 d | 6.22 ± 0.25 d | 11.14 ± 0.24 d |
| R | 26.82 ± 0.95 r | 2.60 ± 0.19 q | 4.05 ± 0.15 p | 8.77 ± 0.25 p | |
| 60 µg/L | E | 39.56 ± 1.71 d | 5.42 ± 0.17 e | 7.84 ± 0.16 e | 13.18 ± 0.58 e |
| R | 30.73 ± 0.92 s | 3.61 ± 0.44 r | 5.08 ± 0.65 q | 11.29 ± 0.95 q | |
| Exposure | F | 148.99 ** | 267.43 ** | 268.9 ** | 184.08 ** |
| Recovery | F | 57.30 ** | 30.34 ** | 11.21 ** | 25.70 ** |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sharma, S.; Iqbal Dar, O.; Andotra, M.; Sharma, S.; Kaur, A.; Faggio, C. Environmentally Relevant Concentrations of Triclosan Induce Cyto-Genotoxicity and Biochemical Alterations in the Hatchlings of Labeo rohita. Appl. Sci. 2021, 11, 10478. https://doi.org/10.3390/app112110478
Sharma S, Iqbal Dar O, Andotra M, Sharma S, Kaur A, Faggio C. Environmentally Relevant Concentrations of Triclosan Induce Cyto-Genotoxicity and Biochemical Alterations in the Hatchlings of Labeo rohita. Applied Sciences. 2021; 11(21):10478. https://doi.org/10.3390/app112110478
Chicago/Turabian StyleSharma, Sunil, Owias Iqbal Dar, Megha Andotra, Simran Sharma, Arvinder Kaur, and Caterina Faggio. 2021. "Environmentally Relevant Concentrations of Triclosan Induce Cyto-Genotoxicity and Biochemical Alterations in the Hatchlings of Labeo rohita" Applied Sciences 11, no. 21: 10478. https://doi.org/10.3390/app112110478
APA StyleSharma, S., Iqbal Dar, O., Andotra, M., Sharma, S., Kaur, A., & Faggio, C. (2021). Environmentally Relevant Concentrations of Triclosan Induce Cyto-Genotoxicity and Biochemical Alterations in the Hatchlings of Labeo rohita. Applied Sciences, 11(21), 10478. https://doi.org/10.3390/app112110478