
Chronic Effects of Diazinon® Exposures Using Integrated Biomarker Responses in Freshwater Walking Catfish, Clarias batrachus

 , , , ,
, , , , 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Organism
2.2. Test Chemical
2.3. Experimental Water
2.4. QA/QC (Quality Assurance/Quality Control) Procedure
2.5. Experimental Design of Chronic Toxicity Study
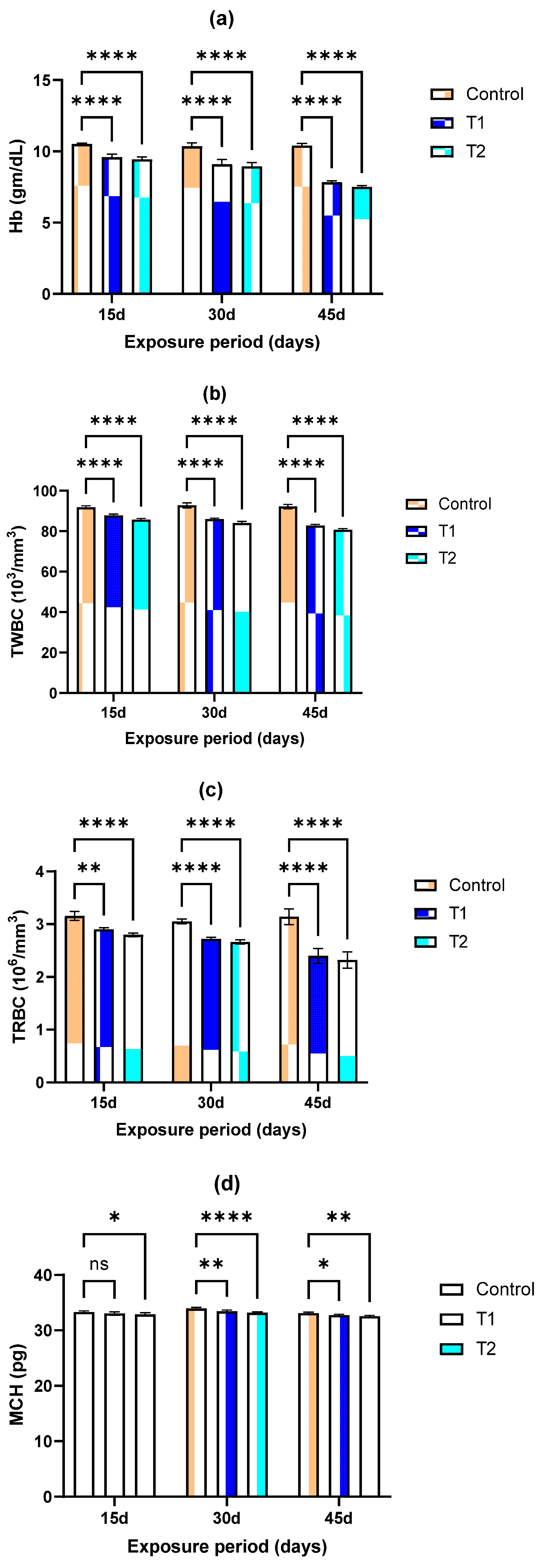
2.6. Hematological Profiles and Indexes
2.7. Growth Endpoints
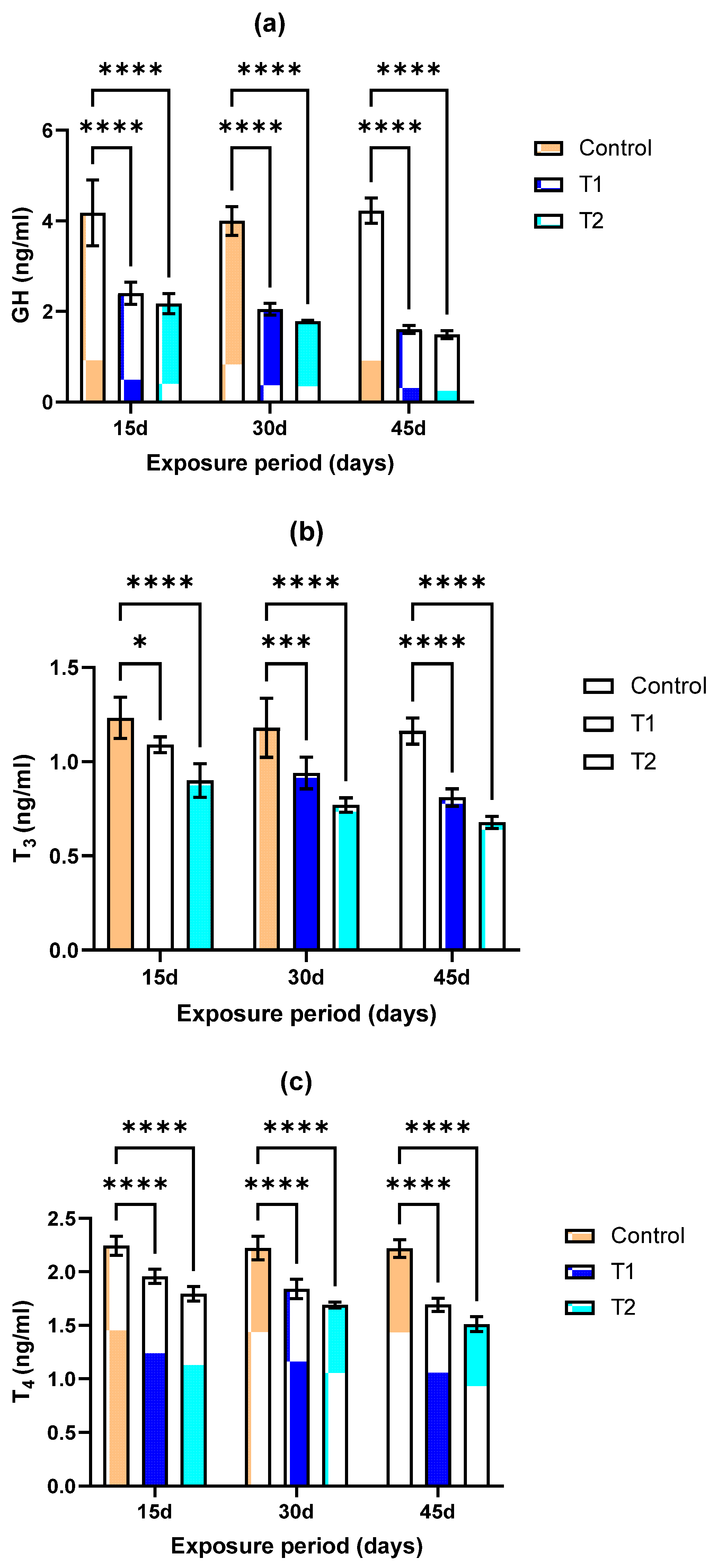
2.8. Endocrine (GH, T3, and T4) Endpoints
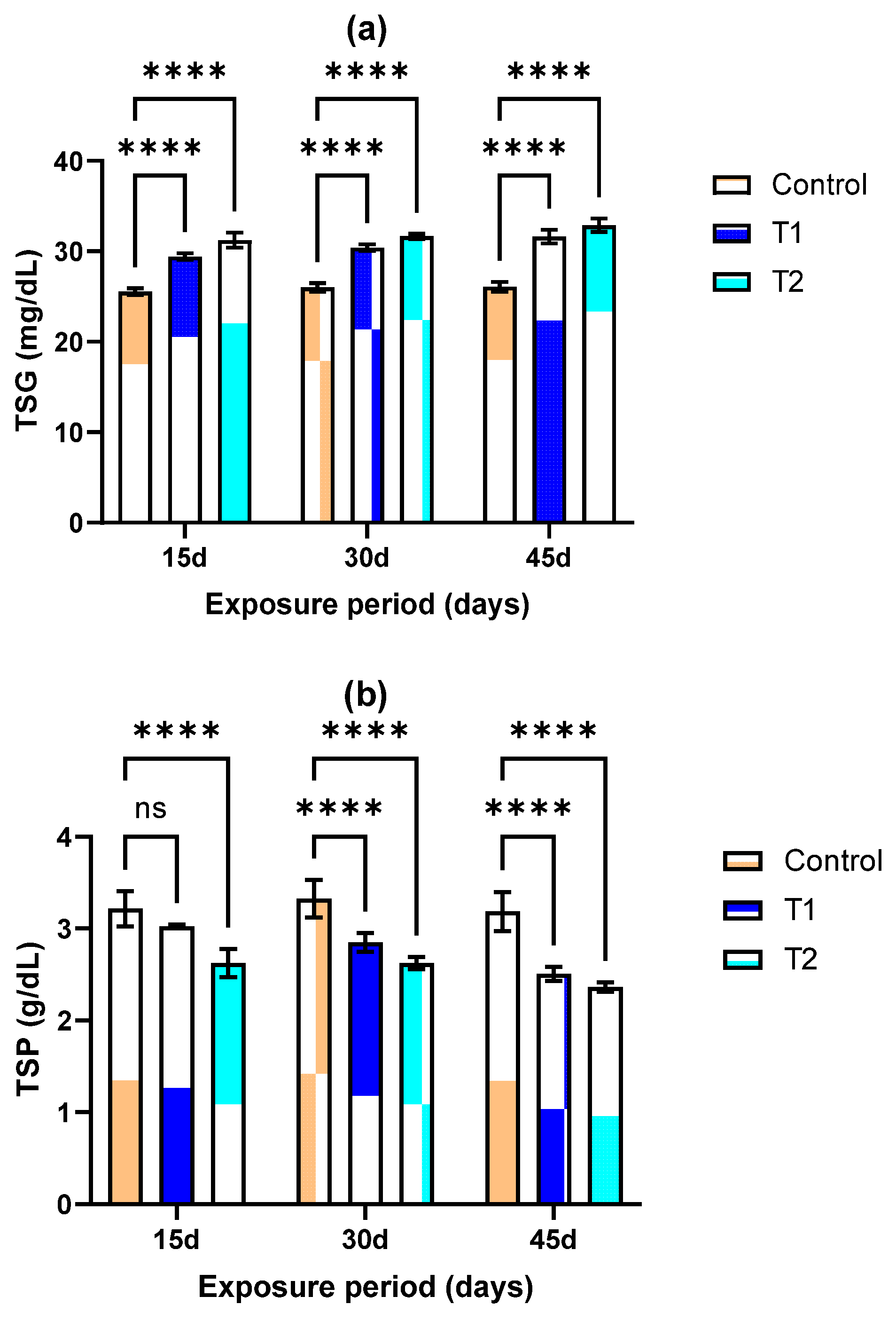
2.9. Biochemical Total Serum Protein (TSP), Total Serum Glucose (TSG)
2.10. Integrated Biomarker Response (IBR)
2.11. Biomarker Response Index (BRI)
3. Results and Discussion
3.1. Hematological Profiles
3.2. Growth Endpoints
3.3. Endocrine Endpoints (GH, T3, and T4)
3.4. Total Serum Protein (TSP) and Total SERUM Glucose (TSG)
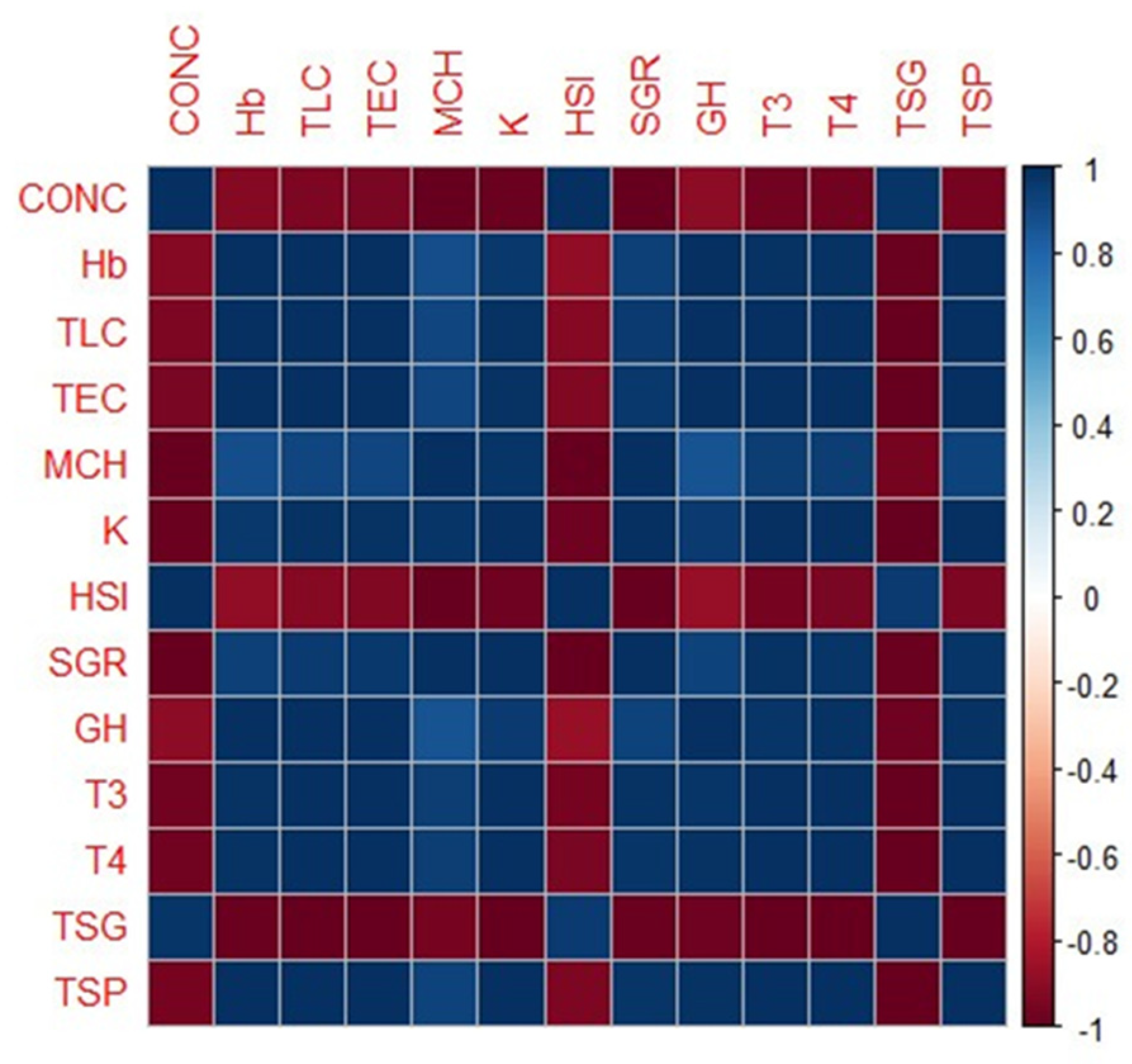
3.5. Correlation Analysis
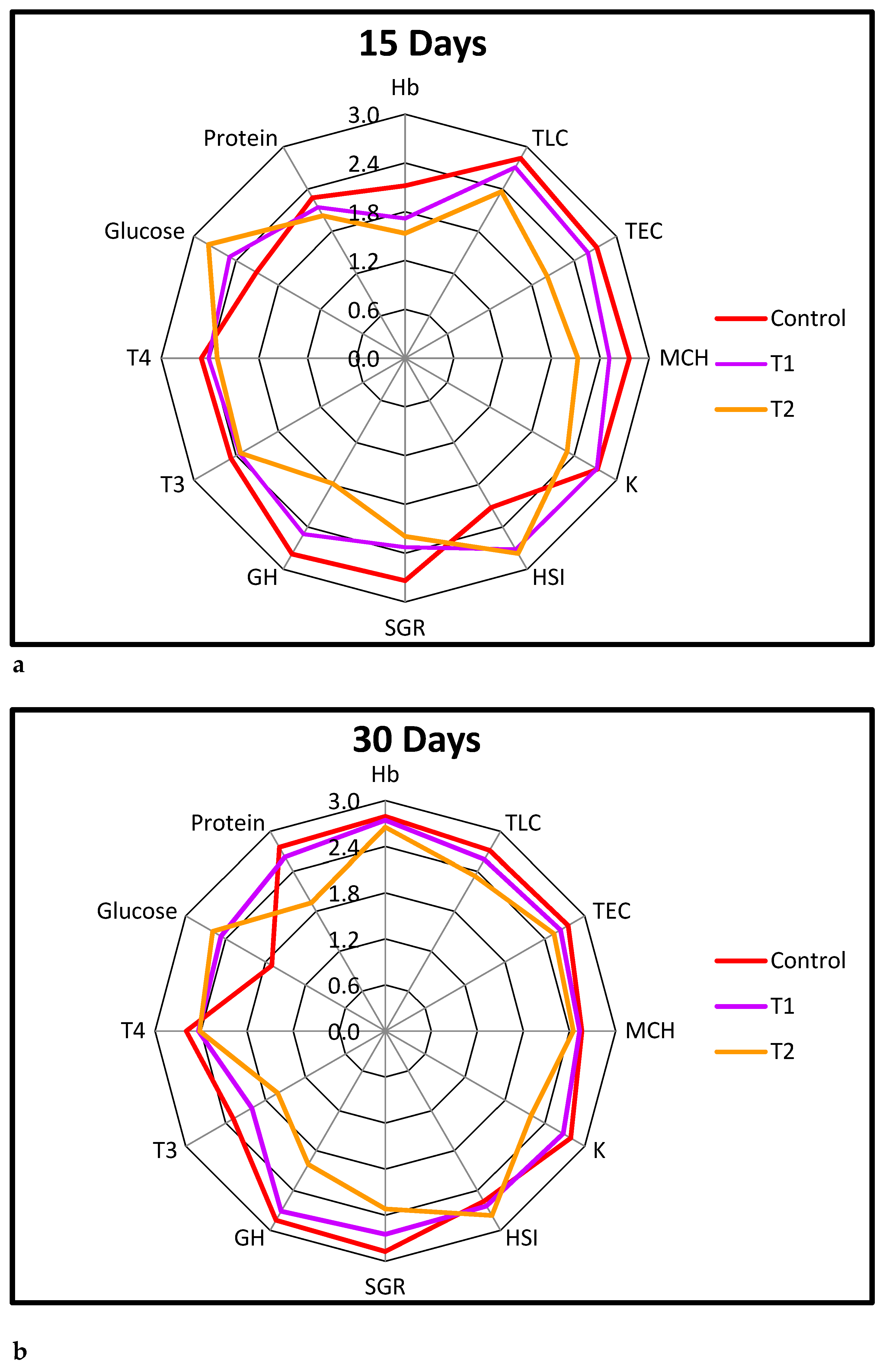
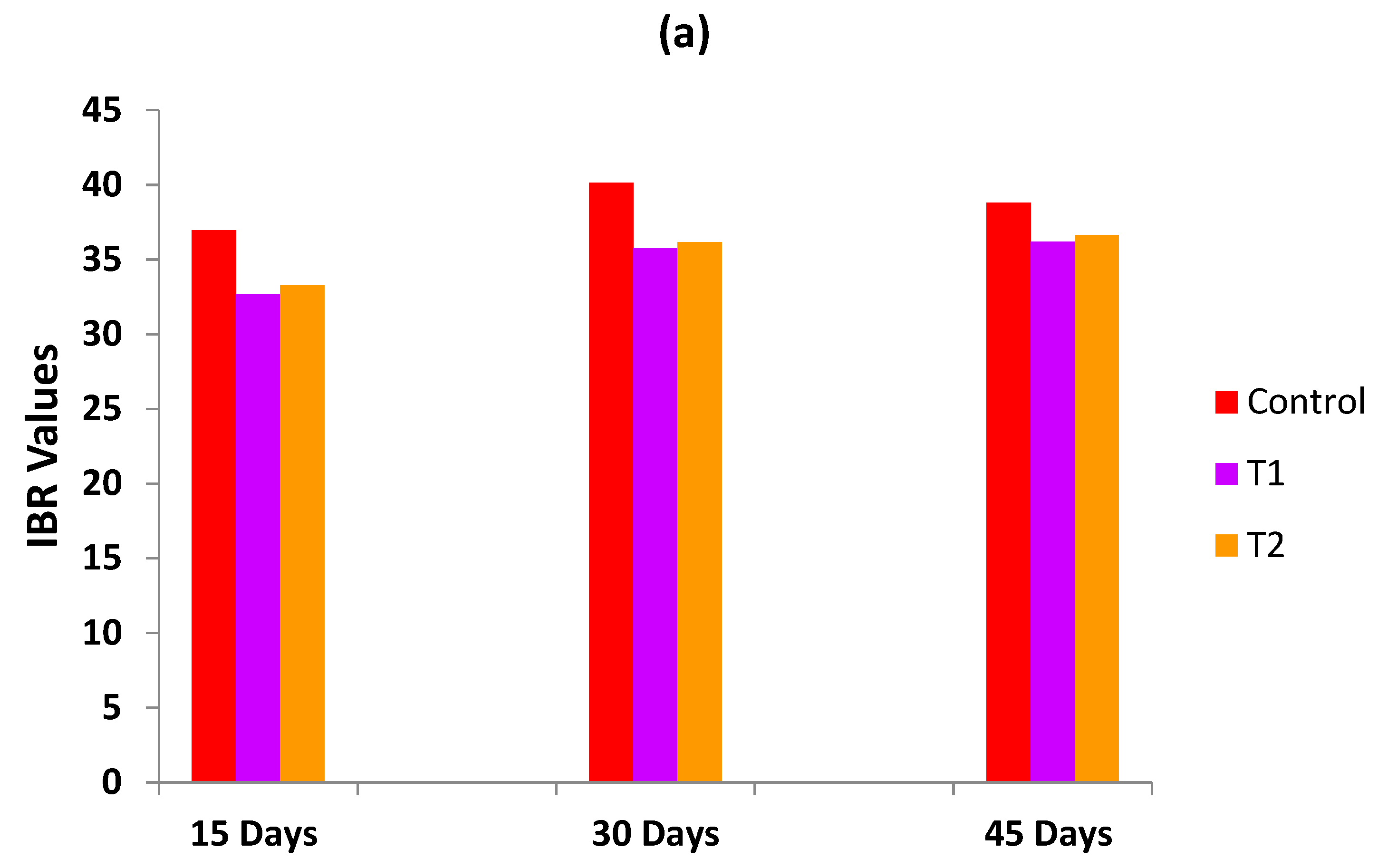
3.6. Integrated Biomarkers Response (IBR)
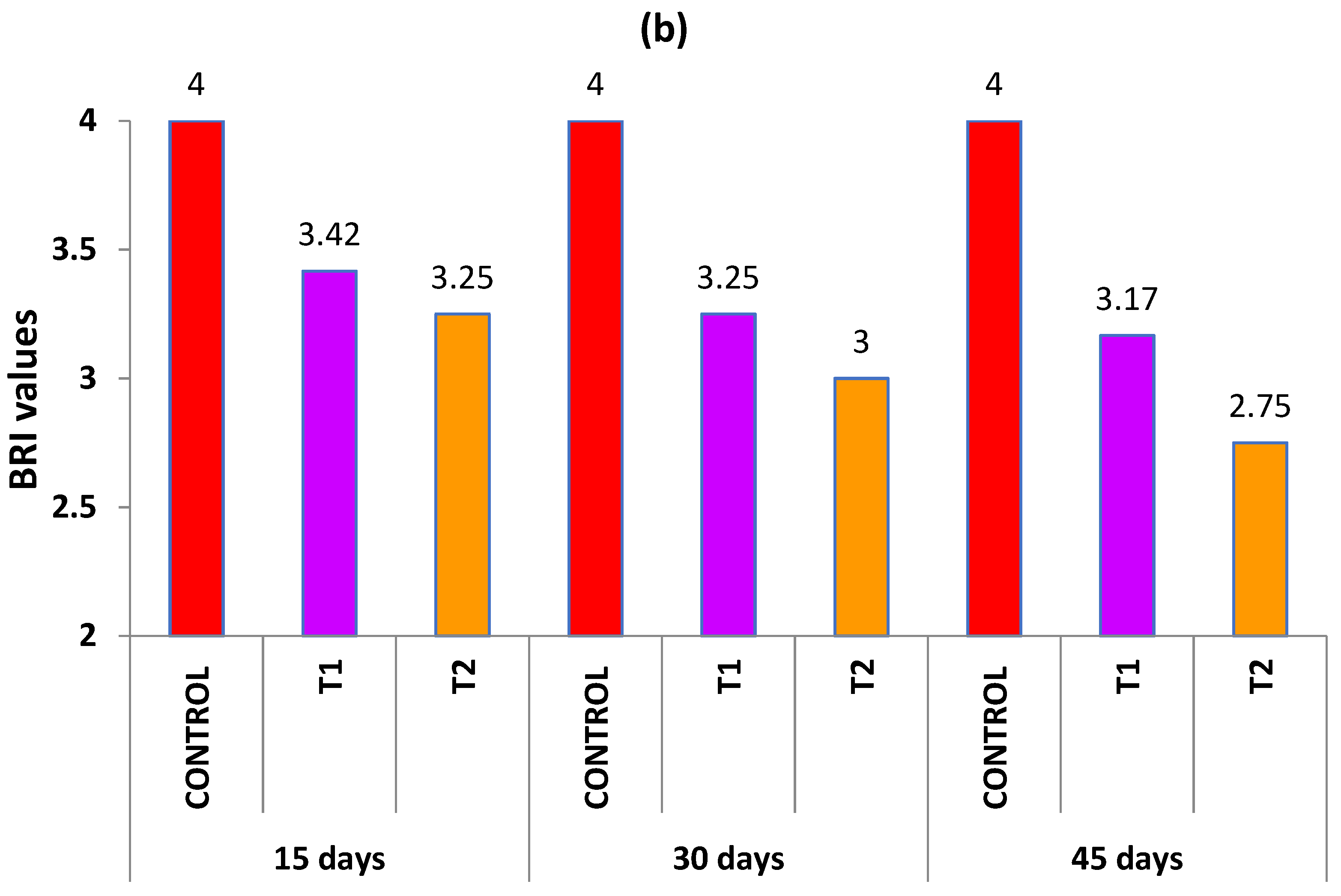
3.7. Biomarker Response Index (BRI)
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Saha, S.; Saha, N.C. Study on Acute Toxicity of Bifenthrin to (Clarias batrachus Linn.). Indian J. Ecol. 2021, 48, 545–548. [Google Scholar]
- Chukwuka, A.V.; Ogbeide, O. Riparian-Buffer Loss and Pesticide Incidence in Freshwater Matrices of Ikpoba River (Nigeria): Policy Recommendations for the Protection of Tropical River Basins. River Basin Manag. Sustain. Issues Plan. Strateg. 2021, 113. [Google Scholar] [CrossRef]
- Ogbeide, O.; Chukwuka, A.; Tongo, I.; Ezemonye, L. Relationship between geosorbent properties and field-based partition coefficients for pesticides in surface water and sediments of selected agrarian catchments: Implications for risk assessment. J. Environ. Manag. 2018, 217, 23–37. [Google Scholar] [CrossRef]
- Sarkar, S.; Gil, J.D.B.; Keeley, J.; Jansen, K. The Use of Pesticides in Developing Countries and Their Impact on Health and the Right to Food; European Union: Brussels, Belgium, 2021. [Google Scholar]
- Xie, J.; Hu, L.; Tang, J.; Wu, X.; Li, N.; Yuan, Y.; Yang, H.; Zhang, J.; Luo, S.; Chen, X. Ecological mechanisms underlying the sustainability of the agricultural heritage rice-fish coculture system. Proc. Natl. Acad. Sci. USA 2011, 108, E1381–E1387. [Google Scholar] [CrossRef] [Green Version]
- Saha, S.; Mukherjee, D.; Dhara, K.; Saha, N.C. Acute Toxicity Bioassay of a Pyrethroid Pesticide Bifenthrin to the Asian stinging Catfish, Heteropneustes Fossilis (Bloch). Curr. World Environ. 2021, 16, 250–258. [Google Scholar] [CrossRef]
- Owokotomo, A.I.; Ajayi, O.O.; Alabi, O.O.; Chukwuka, A.V. Watershed land use, surface water vulnerability and public health risks of two urban rivers, Ado-Ekiti, South-West Nigeria. SN Appl. Sci. 2020, 2, 1–21. [Google Scholar] [CrossRef]
- Barrett, K.; Jaward, F.M. A review of endosulfan, dichlorvos, diazinon, and diuron-pesticides used in Jamaica. Int. J. Environ. Health Res. 2012, 22, 481–499. [Google Scholar] [CrossRef]
- Stara, A.; Pagano, M.; Capillo, G.; Fabrello, J.; Sandova, M.; Vazzana, I.; Zuskova, E.; Velisek, J.; Matozzo, V.; Faggio, C. Assessing the effects of neonicotinoid insecticide on the bivalve mollusc Mytilus galloprovincialis. Sci. Total Environ. 2020, 700, 134914. [Google Scholar] [CrossRef] [PubMed]
- van der Werf, H. Assessing the impact of pesticides on the environment. Agric. Ecosyst. Environ. 1996, 60, 81–96. [Google Scholar] [CrossRef]
- Hodkovicova, N.; Enevova, V.; Cahova, J.; Blahova, J.; Siroka, Z.; Plhalova, L.; Doubkova, V.; Marsalek, P.; Franc, A.; Fiorino, E.; et al. Could the Musk Compound Tonalide Affect Physiological Functions and Act as an Endocrine Disruptor in Rainbow Trout? Physiol. Res. 2020, S595–S606. [Google Scholar] [CrossRef]
- Sula, E.; Aliko, V.; Barceló, D.; Faggio, C. Combined effects of moderate hypoxia, pesticides and PCBs upon crucian carp fish, Carassius carassius, from a freshwater lake in situ ecophysiological approach. Aquat. Toxicol. 2020, 228, 105644. [Google Scholar] [CrossRef] [PubMed]
- Sula, E.; Aliko, V.; Marku, E.; Nuro, A.; Faggio, C. Evaluation of kidney histopathological alterations in Crucian Carp, Carassius carassius, from a pesticide and PCB-contaminated freshwater ecosystem, using light microscopy and organ index mathematical model. Int. J. Aquat. Biol. 2020, 8, 154–165. [Google Scholar]
- Sharma, S.; Dar, O.I.; Singh, K.; Kaur, A.; Faggio, C. Triclosan elicited biochemical and transcriptomic alterations in Labeo rohita larvae. Environ. Toxicol. Pharmacol. 2021, 88, 103748. [Google Scholar] [CrossRef] [PubMed]
- Yalsuyi, A.M.; Vajargah, M.F.; Hajimoradloo, A.; Galangash, M.M.; Prokić, M.D.; Faggio, C. Evaluation of Behavioral Changes and Tissue Damages in Common Carp (Cyprinus carpio) after Exposure to the Herbicide Glyphosate. Veter. Sci. 2021, 8, 218. [Google Scholar] [CrossRef]
- Dhara, K.; Saha, S.; Saha, N.C. Sensitivity of Common Carp, Cyprinus carpio (Linnaeus, 1758) to the Grey List Metal, Zinc under Laboratory Condition. Asian J. Biol. Life Sci. 2021, 10, 132–140. [Google Scholar] [CrossRef]
- Sharma, R.; Jindal, R.; Faggio, C. Impact of cypermethrin in nephrocytes of freshwater fish Catla catla. Environ. Toxicol. Pharmacol. 2021, 88, 103739. [Google Scholar] [CrossRef]
- Stara, A.; Pagano, M.; Capillo, G.; Fabrello, J.; Sandova, M.; Albano, M.; Zuskova, E.; Velisek, J.; Matozzo, V.; Faggio, C. Acute effects of neonicotinoid insecticides on Mytilus galloprovincialis: A case study with the active compound thiacloprid and the commercial formulation calypso 480 SC. Ecotoxicol. Environ. Saf. 2020, 203, 110980. [Google Scholar] [CrossRef]
- Gharaei, A.; Karimi, M.; Mirdar, J.; Miri, M.; Faggio, C. Population growth of Brachionus calyciflorus affected by deltamethrin and imidacloprid insecticides. Iran. J. Fish. Sci. 2018, 19, 588–601. [Google Scholar]
- Karanth, S. Diazinon. In Encyclopedia of Toxicology; Elsevier BV: Amsterdam, The Netherlands, 2014; pp. 55–56. [Google Scholar]
- Rogers, H.; Schmidt, T.S.; Dabney, B.L.; Hladik, M.L.; Mahler, B.J.; Van Metre, P.C. Bifenthrin Causes Trophic Cascade and Altered Insect Emergence in Mesocosms: Implications for Small Streams. Environ. Sci. Technol. 2016, 50, 11974–11983. [Google Scholar] [CrossRef] [PubMed]
- Dinham, B. Prolonged exposure to some agricultural pesticides may increase the risk of lung cancer in agricultural workers. Evid. Based Healthc. Public Health 2005, 9, 203–205. [Google Scholar] [CrossRef]
- Čolović, M.; Krstić, D.; Petrović, S.; Leskovac, A.; Joksić, G.; Savić, J.; Franko, M.; Trebše, P.; Vasić, V. Toxic effects of diazinon and its photodegradation products. Toxicol. Lett. 2010, 193, 9–18. [Google Scholar] [CrossRef]
- Poet, T.; Kousba, A.; Dennison, S.; Timchalk, C. Physiologically Based Pharmacokinetic/Pharmacodynamic Model for the Organophosphorus Pesticide Diazinon. NeuroToxicology 2004, 25, 1013–1030. [Google Scholar] [CrossRef] [PubMed]
- Shah, M.D.; Iqbal, M. Diazinon-induced oxidative stress and renal dysfunction in rats. Food Chem. Toxicol. 2010, 48, 3345–3353. [Google Scholar] [CrossRef] [PubMed]
- Al-Otaibi, A.M.; Al-Balawi, H.F.A.; Ahmad, Z.; Suliman, E.M. Toxicity bioassay and sub-lethal effects of diazinon on blood profile and histology of liver, gills and kidney of catfish, Clarias gariepinus. Braz. J. Biol. 2019, 79, 326–336. [Google Scholar] [CrossRef] [Green Version]
- Faria, M.; Prats, E.; Ramírez, J.R.R.; Bellot, M.; Bedrossiantz, J.; Pagano, M.; Valls, A.; Gomez-Canela, C.; Porta, J.M.; Mestres, J.; et al. Androgenic activation, impairment of the monoaminergic system and altered behavior in zebrafish larvae exposed to environmental concentrations of fenitrothion. Sci. Total Environ. 2021, 775, 145671. [Google Scholar] [CrossRef] [PubMed]
- Farkhondeh, T.; Aschner, M.; Sadeghi, M.; Mehrpour, O.; Naseri, K.; Amirabadizadeh, A.; Roshanravan, B.; Aramjoo, H.; Samarghandian, S. The effect of diazinon on blood glucose homeostasis: A systematic and meta-analysis study. Environ. Sci. Pollut. Res. 2021, 28, 4007–4018. [Google Scholar] [CrossRef]
- Vajargah, M.F.; Namin, J.I.; Mohsenpour, R.; Yalsuyi, A.M.; Prokić, M.D.; Faggio, C. Histological effects of sublethal concentrations of insecticide Lindane on intestinal tissue of grass carp (Ctenopharyngodon idella). Veter. Res. Commun. 2021, 1–8. [Google Scholar] [CrossRef]
- Stara, A.; Pagano, M.; Albano, M.; Savoca, S.; Di Bella, G.; Albergamo, A.; Koutkova, Z.; Sandova, M.; Velisek, J.; Fabrello, J.; et al. Effects of long-term exposure of Mytilus galloprovincialis to thiacloprid: A multibiomarker approach. Environ. Pollut. 2021, 289, 117892. [Google Scholar] [CrossRef]
- Boussabbeh, M.; Ben Salem, I.; Hamdi, M.; Ben Fradj, S.; Abid-Essefi, S.; Bacha, H. Diazinon, an organophosphate pesticide, induces oxidative stress and genotoxicity in cells deriving from large intestine. Environ. Sci. Pollut. Res. 2015, 23, 2882–2889. [Google Scholar] [CrossRef]
- Radovanović, T.B.; Gavrilović, B.R.; Petrović, T.G.; Despotović, S.G.; Gavrić, J.P.; Kijanović, A.; Mirč, M.; Kolarov, N.T.; Faggio, C.; Prokić, M.D. Impact of desiccation pre-exposure on deltamethrin-induced oxidative stress in Bombina variegata juveniles. Comp. Biochem. Physiol. Part. C Toxicol. Pharmacol. 2021, 250, 109191. [Google Scholar] [CrossRef]
- Kim, S.; Jung, J.; Lee, I.; Jung, D.; Youn, H.; Choi, K. Thyroid disruption by triphenyl phosphate, an organophosphate flame retardant, in zebrafish (Danio rerio) embryos/larvae, and in GH3 and FRTL-5 cell lines. Aquat. Toxicol. 2015, 160, 188–196. [Google Scholar] [CrossRef]
- Flynn, K.; Johnson, R.; Lothenbach, D.; Swintek, J.; Whiteman, F.; Etterson, M. The effects of combinations of limited ration and diazinon exposure on acetylcholinesterase activity, growth and reproduction in Oryzias latipes, the Japanese medaka. J. Appl. Toxicol. 2020, 40, 535–547. [Google Scholar] [CrossRef]
- Sharma, S.; Dar, O.I.; Andotra, M.; Sharma, S.; Kaur, A.; Faggio, C. Environmentally relevant concentrations of Triclosan induce cyto-genotoxicity and biochemical alterations in the hatchlings of Labeo rohita. Appl. Sci. 2021, 11, 10478. [Google Scholar] [CrossRef]
- Sharma, R.; Jindal, R.; Faggio, C. Cassia fistula ameliorates chronic toxicity of cypermethrin in Catla catla. Comp. Biochem. Physiol. Part. C Toxicol. Pharmacol. 2021, 248, 109113. [Google Scholar] [CrossRef] [PubMed]
- Sinha, R.; Jindal, R.; Faggio, C. Protective Effect of Emblica officinalis in Cyprinus carpio against Hepatotoxicity Induced by Malachite Green: Ultrastructural and Molecular Analysis. Appl. Sci. 2021, 11, 3507. [Google Scholar] [CrossRef]
- Sinha, R.; Jindal, R.; Faggio, C. Nephroprotective effect of Emblica officinalis fruit extract against malachite green toxicity in piscine model: Ultrastructure and oxidative stress study. Microsc. Res. Tech. 2021, 84, 1911–1919. [Google Scholar] [CrossRef]
- Strungaru, S.-A.; Pohontiu, C.M.; Nicoara, M.; Teodosiu, C.; Baltag, E.S.; Jijie, R.; Plavan, G.; Pacioglu, O.; Faggio, C. Response of aquatic macroinvertebrates communities to multiple anthropogenic stressors in a lowland tributary river. Environ. Toxicol. Pharmacol. 2021, 87, 103687. [Google Scholar] [CrossRef]
- Crane, M.; Giddings, J.M. “Ecologically Acceptable Concentrations” When Assessing the Environmental Risks of Pesticides Under European Directive 91/414/EEC. Hum. Ecol. Risk Assess. Int. J. 2004, 10, 733–747. [Google Scholar] [CrossRef]
- Dhara, K.; Saha, S.; Pal, P.; Chukwuka, A.V.; Panigrahi, A.K.; Saha, N.C.; Faggio, C. Biochemical, physiological (haematological, oxygen-consumption rate) and behavioural effects of mercury exposures on the freshwater snail, Bellamya bengalensis. Comp. Biochem. Physiol. Part C Toxicol. Pharmacol. 2021, 251, 109195. [Google Scholar] [CrossRef]
- APHA; AWWA. Standard Methods for the Examination of Water and Wastewater, 19th ed.; APHA: Washington, DC, USA, 1995. [Google Scholar]
- Palacio, J.A.; Henao, B.; Vélez, J.H.; González, J.; Parra, C.M. Acute toxicity and bioaccumulation of pesticide Diazinon in red tilapia (Oreochromis niloticus x Mossambicus albina). Environ. Toxicol. 2002, 17, 334–340. [Google Scholar] [CrossRef]
- Saha, S. Study on the effects of some commonly used new generation pesticides on fish and fish food organisms and their role on agricultural runoff in the change of water quality and piscine diversity of Kankana Baor and Beri Baor, North 24 Parganas, West Bengal. In Department of Zoology; The University of Burdwan: Burdwan, India, 2021. [Google Scholar]
- Gomez, K.A.; Gomez, A.A. Statistical Procedures for Agricultural Research, 2nd ed.; John Wiley & Sons: New York, NY, USA, 1984. [Google Scholar]
- Bain, B.J.; Bates, I.; Laffan, M.A. Dacie and Lewis Practical Haematology E-Book; Elsevier Health Sciences: Amsterdam, The Netherlands, 2016. [Google Scholar]
- Hale, V. Dacie and Lewis Practical Haematology. Aust. J. Med Sci. 2008, 29, 19. [Google Scholar]
- Bhunia, F.; Saha, N.C.; Kaviraj, A. Toxicity of Thiocyanate to Fish, Plankton, Worm, and Aquatic Ecosystem. Bull. Environ. Contam. Toxicol. 2000, 64, 197–204. [Google Scholar] [CrossRef] [PubMed]
- Kaviraj, A.; Bhunia, F.; Saha, N.C. Toxicity of Methanol to Fish, Crustacean, Oligochaete Worm, and Aquatic Ecosystem. Int. J. Toxicol. 2004, 23, 55–63. [Google Scholar] [CrossRef] [PubMed]
- Lal, B.; Singh, A.K. Immunological and physiological validation of enzyme-linked immunosorbent assay for growth hormone of the Asian catfish, Clarias batrachus. Fish. Physiol. Biochem. 2005, 31, 289–293. [Google Scholar] [CrossRef] [PubMed]
- Lowry, O.H.; Rosebrough, N.J.; Farr, A.L.; Randall, R.J. Protein measurement with the Folin phenol reagent. J. Biol. Chem. 1951, 193, 265–275. [Google Scholar] [CrossRef]
- Beliaeff, B.; Burgeot, T. Integrated biomarker response: A useful tool for ecological risk assessment. Environ. Toxicol. Chem. 2002, 21, 1316–1322. [Google Scholar] [CrossRef]
- Samanta, P.; Im, H.; Na, J.; Jung, J. Ecological risk assessment of a contaminated stream using multi-level integrated biomarker response in Carassius auratus. Environ. Pollut. 2018, 233, 429–438. [Google Scholar] [CrossRef]
- Hagger, J.A.; Jones, M.B.; Lowe, D.; Leonard, D.P.; Owen, R.; Galloway, T.S. Application of biomarkers for improving risk assessments of chemicals under the Water Framework Directive: A case study. Mar. Pollut. Bull. 2008, 56, 1111–1118. [Google Scholar] [CrossRef]
- Li, X.; Wang, M.; Chen, W.; Jiang, R. Evaluation of combined toxicity of Siduron and cadmium on earthworm (Eisenia fetida) using Biomarker Response Index. Sci. Total. Environ. 2019, 646, 893–901. [Google Scholar] [CrossRef]
- Nwani, C.D.; Ivoke, N.; Ugwu, D.O.; Atama, C.; Onyishi, G.C.; Echi, P.C.; Ogbonna, S.A. Investigation on acute toxicity and behavioral changes in a freshwater African catfish, Clarias gariepinus (Burchell, 1822), exposed to organophosphorous pesticide, Termifos®. Pak. J. Zool. 2013, 45, 959–965. [Google Scholar]
- Rauf, A.; Arain, N. Acute toxicity of diazinon and its effects on hematological parameters in the Indian carp, Cirrhinus mrigala (Hamilton). Turk. J. Veter. Anim. Sci. 2013, 37, 535–540. [Google Scholar] [CrossRef]
- Morgan, J.E.; Hall, N.D.; Collins, A.J.; Bacon, P.A. The nonspecific inhibitory effect of synovial tissue extracts on leucocyte migration in vitro. Ann. Rheum. Dis. 1980, 39, 323–328. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chatterjee, S.; Basak, P.; Chaklader, M.; Das, P.; Pereira, J.A.; Chaudhuri, S.; Law, S. Pesticide induced alterations in marrow physiology and depletion of stem and stromal progenitor population: An experimental model to study the toxic effects of pesticide. Environ. Toxicol. 2014, 29, 84–97. [Google Scholar] [CrossRef] [PubMed]
- Shamoushaki, M.M.N.; Soltani, M.; Kamali, A.; Imanpoor, M.R.; Sharifpour, I.; Khara, H. Effects of organophosphate, diazinon on some haematological and biochemical changes in Rutilus frisii kutum (Kamensky, 1901) male brood stocks. Iran. J. Fish. Sci. 2012, 11, 105–117. [Google Scholar]
- Svoboda, M.; Lusková, V.; Drastichová, J.; Žlábek, V. The Effect of Diazinon on Haematological Indices of Common Carp (Cyprinus carpio L.). Acta Veter. Brno 2001, 70, 457–465. [Google Scholar] [CrossRef] [Green Version]
- Štěpánová, S.; Plhalová, L.; Doleželová, P.; Prokeš, M.; Maršálek, P.; Škorič, M.; Svobodová, Z. The Effects of Subchronic Exposure to Terbuthylazine on Early Developmental Stages of Common Carp. Sci. World J. 2012, 2012, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Babatunde, T.A.; Chukwuka, A.V. Evaluation of Response Patterns in Somatic and Otolith Features of Laboratory- Reared and Wild Clarias gariepinus Exposed to Industrial Effluent. Res. J. Appl. Sci. Eng. Technol. 2013, 5, 626–634. [Google Scholar] [CrossRef]
- Adeogun, A.O.; Ogidan, I.M.; Ibor, O.R.; Chukwuka, A.V.; Adedara, I.A.; Farombi, E.O. Long-term exposure to industrial effluent induces oxidative stress and affects growth in Clarias gariepinus. Res. J. Environ. Earth Sci. 2012, 4, 738–746. [Google Scholar]
- van der Oost, R.; Beyer, J.; Vermeulen, N.P. Fish bioaccumulation and biomarkers in environmental risk assessment: A review. Environ. Toxicol. Pharmacol. 2003, 13, 57–149. [Google Scholar] [CrossRef]
- Scott, G.R.; Sloman, K.A. The effects of environmental pollutants on complex fish behaviour: Integrating behavioural and physiological indicators of toxicity. Aquat. Toxicol. 2004, 68, 369–392. [Google Scholar] [CrossRef]
- Saxena, A. Biochemical changes in liver and muscles of Channa gachua during reproductive cycle. Aquaculture 2000, 1, 51–56. [Google Scholar]
- Devi, M.; Gopal, V.; Ratna, G. Liver-somatic index of Channa striatus as a biomonitoring tool of heavy metal and pesticide toxicity. J. Ecotoxicol. Environ. Monit. 1991, 1, 135–141. [Google Scholar]
- Barse, A.; Chakrabarti, T.; Ghosh, T.; Pal, A.; Jadhao, S. One-tenth dose of LC50 of 4-tert-butylphenol causes endocrine disruption and metabolic changes in Cyprinus carpio. Pestic. Biochem. Physiol. 2006, 86, 172–179. [Google Scholar] [CrossRef]
- Abdel-Hameid, N.-A.H. Physiological and histopathological alterations induced by phenol exposure in Oreochromis aureus juveniles. Turk. J. Fish. Aquat. Sci. 2007, 7, 131–138. [Google Scholar]
- Sadekarpawar, S.; Parikh, P. Gonadosomatic and hepatosomatic indices of freshwater fish Oreochromis mossambicus in response to a plant nutrient. World J. Zool. 2013, 8, 110–118. [Google Scholar]
- Frontera, J.L.; Vatnick, I.; Chaulet, A.; Rodríguez, E.M. Effects of Glyphosate and Polyoxyethylenamine on Growth and Energetic Reserves in the Freshwater Crayfish Cherax quadricarinatus (Decapoda, Parastacidae). Arch. Environ. Contam. Toxicol. 2011, 61, 590–598. [Google Scholar] [CrossRef] [PubMed]
- Majumder, R.; Kaviraj, A. Acute and sublethal effects of organophosphate insecticide chlorpyrifos on freshwater fish Oreochromis niloticus. Drug Chem. Toxicol. 2018, 42, 487–495. [Google Scholar] [CrossRef] [PubMed]
- McCarthy, I.D.; Fuiman, L.A. Growth and protein metabolism in red drum (Sciaenops ocellatus) larvae exposed to environmental levels of atrazine and malathion. Aquat. Toxicol. 2008, 88, 220–229. [Google Scholar] [CrossRef]
- Foss, A.; Siikavuopio, S.I.; Sæther, B.-S.; Evensen, T.H. Effect of chronic ammonia exposure on growth in juvenile Atlantic cod. Aquaculture 2004, 237, 179–189. [Google Scholar] [CrossRef]
- Muhammad, U.; Yasid, N.; Daud, H.; Shukor, M. Glyphosate Herbicide Induces Changes in the Growth Pattern and Somatic Indices of Crossbred Red Tilapia (O. niloticus × O. mossambicus). Animals 2021, 11, 1209. [Google Scholar] [CrossRef]
- Lal, B.; Sarang, M.K.; Kumar, P. Malathion exposure induces the endocrine disruption and growth retardation in the catfish, Clarias batrachus (Linn.). Gen. Comp. Endocrinol. 2013, 181, 139–145. [Google Scholar] [CrossRef]
- Senthilkumaran, B. Pesticide- and sex steroid analogue-induced endocrine disruption differentially targets hypothalamo–hypophyseal–gonadal system during gametogenesis in teleosts—A review. Gen. Comp. Endocrinol. 2015, 219, 136–142. [Google Scholar] [CrossRef]
- Zhu, L.; Li, W.; Zha, J.; Wang, M.; Yuan, L.; Wang, Z. Butachlor causes disruption of HPG and HPT axes in adult female rare minnow (Gobiocypris rarus). Chem. Interact. 2014, 221, 119–126. [Google Scholar] [CrossRef]
- Liu, L.; Zhu, B.; Gong, Y.-X.; Liu, G.-L.; Wang, G.-X. Neurotoxic effect of triazophos on goldfish (Carassius auratus) and tissue specific antioxidant responses. Ecotoxicol. Environ. Saf. 2015, 116, 68–75. [Google Scholar] [CrossRef] [PubMed]
- Yu, L.-Q.; Zhao, G.-F.; Feng, M.; Wen, W.; Li, K.; Zhang, P.-W.; Peng, X.; Huo, W.-J.; Zhou, H.-D. Chronic exposure to pentachlorophenol alters thyroid hormones and thyroid hormone pathway mRNAs in zebrafish. Environ. Toxicol. Chem. 2014, 33, 170–176. [Google Scholar] [CrossRef]
- Liang, X.; Yu, L.; Gui, W.; Zhu, G. Exposure to difenoconazole causes changes of thyroid hormone and gene expression levels in zebrafish larvae. Environ. Toxicol. Pharmacol. 2015, 40, 983–987. [Google Scholar] [CrossRef] [PubMed]
- Power, D.; Llewellyn, L.; Faustino, M.; Nowell, M.; Bjornsson, B.T.; Einarsdottir, I.; Canario, A.; Sweeney, G. Thyroid hormones in growth and development of fish. Comp. Biochem. Physiol. Part. C Toxicol. Pharmacol. 2001, 130, 447–459. [Google Scholar] [CrossRef]
- Olakolu, F.C.; Chukwuka, A.V. Trace metal concentrations and antioxidant activity in ovarian tissue of blue crab Callinectes amnicola from Lagos lagoon and implications for reproductive success. Zool. Ecol. 2014, 24, 278–284. [Google Scholar] [CrossRef]
- Maruthanayagam, C.; Sharmila, G. Effect of monocrotophos on biochemical markers DNA and RNA in Cyprinus carpio during exposure and recovery period. Fish. Manag. 2004, 21, 163. [Google Scholar]
- Soyingbe, A. Effects of Sublethal Concentrations of Diazinon on Total Protein in Tilapia Fish (Oreochromis Niloticus). IOSR J. Environ. Sci. Toxicol. Food Technol. 2012, 1, 22–25. [Google Scholar] [CrossRef]
- Matin, M.A.; Sattar, S.; Husain, K. The role of adrenals in diazinon-induced changes in carbohydrate metabolism in rats. Arch. Ind. Hyg. Toxicol. 1990, 41, 347–355. [Google Scholar]
- Abdelkhalek, N.; Eissa, I.A.; Ahmed, E.; Kilany, O.; El-Adl, M.; Dawood, M.; Hassan, A.M.; Abdel-Daim, M.M. Protective role of dietary Spirulina platensis against diazinon-induced Oxidative damage in Nile tilapia; Oreochromis niloticus. Environ. Toxicol. Pharmacol. 2017, 54, 99–104. [Google Scholar] [CrossRef]
- Burgos-Aceves, M.A.; Lionetti, L.; Faggio, C. Multidisciplinary haematology as prognostic device in environmental and xenobiotic stress-induced response in fish. Sci. Total. Environ. 2019, 670, 1170–1183. [Google Scholar] [CrossRef]
- Aliko, V.; Mehmeti, E.; Qirjo, M.; Faggio, C. ‘Drink and sleep like a fish’: Goldfish as a behavior model to study pharmaceutical effects in freshwater ecosystems. J. Biol. Res. Boll. Della Soc. Ital. Biol. Sper. 2019, 92. [Google Scholar] [CrossRef] [Green Version]
- Aliko, V.; Qirjo, M.; Sula, E.; Morina, V.; Faggio, C. Antioxidant defense system, immune response and erythron profile modulation in gold fish, Carassius auratus, after acute manganese treatment. Fish. Shellfish. Immunol. 2018, 76, 101–109. [Google Scholar] [CrossRef] [PubMed]
- Sula, E.; Aliko, V.; Pagano, M.; Faggio, C. Digital light microscopy as a tool in toxicological evaluation of fish erythrocyte morphological abnormalities. Microsc. Res. Tech. 2019, 83, 362–369. [Google Scholar] [CrossRef]
- Silva-Herdade, A.S.; Andolina, G.; Faggio, C.; Calado, Â.; Saldanha, C. Erythrocyte deformability—A partner of the inflammatory response. Microvasc. Res. 2016, 107, 34–38. [Google Scholar] [CrossRef]
- Ololade, I.; Oginni, O. Toxic stress and hematological effects of nickel on African catfish, Clarias gariepinus, fingerlings. J. Environ. Chem. Ecotoxicol. 2010, 2, 014–019. [Google Scholar] [CrossRef]
- Adeogun, A.; Salami, O.A.; Chukwuka, A.V.; Alaka, O.O. Haematological and Serum Biochemical Profile of the Blue Crab, Callinectes amnicola from two Tropical Lagoon Ecosystems. Afr. J. Biomed. Res. 2015, 18, 233–247. [Google Scholar]
- Alam, M.M.; Jahan, S.N.; Hussain, M.A.; De, M.; Goutham-Bharathi, M.P.; Magalhães, A.B.; Simon, K.D. Length-length relationship, length-weight relationship and condition factor of freshwater fish species of Bangladesh. Aquac. Aquar. Conserv. Legis. 2013, 6, 498–509. [Google Scholar]
- Barton, B.A. Stress in Fishes: A Diversity of Responses with Particular Reference to Changes in Circulating Corticosteroids. Integr. Comp. Biol. 2002, 42, 517–525. [Google Scholar] [CrossRef]
- De Schamphelaere, K.A.; Koene, J.M.; Heijerick, D.G.; Janssen, C.R. Reduction of growth and haemolymph Ca levels in the freshwater snail Lymnaea stagnalis chronically exposed to cobalt. Ecotoxicol. Environ. Saf. 2008, 71, 65–70. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shiao, J.-C.; Hwang, P.-P. Thyroid hormones are necessary for the metamorphosis of tarpon Megalops cyprinoides leptocephali. J. Exp. Mar. Biol. Ecol. 2006, 331, 121–132. [Google Scholar] [CrossRef]
- Yamano, K. The Role of Thyroid Hormone in Fish Development with Reference to Aquaculture. Jpn. Agric. Res. Q. 2005, 39, 161–168. [Google Scholar] [CrossRef] [Green Version]
- Schmid, A.C.; Lutz, I.; Kloas, W.; Reinecke, M. Thyroid hormone stimulates hepatic IGF-I mRNA expression in a bony fish, the tilapia Oreochromis mossambicus, in vitro and in vivo. Gen. Comp. Endocrinol. 2003, 130, 129–134. [Google Scholar] [CrossRef]
- Heydarnejad, M.S.; Khosravian-Hemamai, M.; Nematollahi, A. Effects of cadmium at sub-lethal concentration on growth and biochemical parameters in rainbow trout (Oncorhynchus mykiss). Ir. Veter-J. 2013, 66, 11. [Google Scholar] [CrossRef] [Green Version]
- Zutshi, B.; Prasad, S.G.R.; Nagaraja, R. Alteration in hematology of Labeo rohita under stress of pollution from Lakes of Bangalore, Karnataka, India. Environ. Monit. Assess. 2010, 168, 11–19. [Google Scholar] [CrossRef]
- David, M.; Kumar, R.S.; Mushigeri, S.B.; Kuri, R.C. Blood glucose and glycogen levels as indicators of stress in the freshwater fish, Labeo rohita under fenvalerate intoxication. J. Ecotoxicol. Environ. Monit. 2005, 15, 1–5. [Google Scholar]
- Ibor, O.R.; Adeogun, A.O.; Regoli, F.; Arukwe, A. Xenobiotic biotransformation, oxidative stress and obesogenic molecular biomarker responses in Tilapia guineensis from Eleyele Lake, Nigeria. Ecotoxicol. Environ. Saf. 2019, 169, 255–265. [Google Scholar] [CrossRef]
- Sudarshan, S.; Kulkarni, R.S. Determination of Condition Factor (K) Somatic Condition Factor (Ks) Hepatic and Gonado Somatic Indices in The Fresh Water Fish Notopterus Notopterus. Int. J. Sci. Res. 2012, 2, 524–526. [Google Scholar] [CrossRef]
- Bebianno, M.; Cabra, H. Application of an integrated biomarker response index (IBR) to assess temporal variation of environmental quality in two Portuguese aquatic systems. Ecol. Indic. 2012, 19, 215–225. [Google Scholar]
- Pal, S.; Samanta, P.; Kole, D.; Mukherjee, A.K.; Ghosh, A.R. Acute Toxicity and Oxidative Stress Responses in Tadpole of Skittering Frog, Euphlyctis cyanophlyctis (Schneider, 1799) to Sodium Fluoride Exposure. Bull. Environ. Contam. Toxicol. 2018, 100, 202–207. [Google Scholar] [CrossRef] [PubMed]
- Dey, S.; Ballav, P.; Samanta, P.; Mandal, A.; Patra, A.; Das, S.; Mondal, A.K.; Ghosh, A.R. Time-Dependent Naphthalene Toxicity in Anabas testudineus (Bloch): A Multiple Endpoint Biomarker Approach. ACS Omega 2021, 6, 317–326. [Google Scholar] [CrossRef]
- Nayak, S.; Dash, S.N.; Pati, S.S.; Priyadarshini, P.; Patnaik, L. Lipid peroxidation and antioxidant levels in Anabas testudineus (Bloch) under naphthalene (PAH) stress. Aquac. Res. 2021, 52, 5739–5749. [Google Scholar] [CrossRef]
- Parolini, M.; Pedriali, A.; Binelli, A. Application of a Biomarker Response Index for Ranking the Toxicity of Five Pharmaceutical and Personal Care Products (PPCPs) to the Bivalve Dreissena polymorpha. Arch. Environ. Contam. Toxicol. 2013, 64, 439–447. [Google Scholar] [CrossRef] [PubMed]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameters | 15 Days | 30 Days | 45 Days | ||||||
|---|---|---|---|---|---|---|---|---|---|
| Dose (mg/L) | |||||||||
| 0.00 | 0.64 | 1.28 | 0.00 | 0.64 | 1.28 | 0.00 | 0.64 | 1.28 | |
| Temperature (°C) | 28.2 ± 0.12 | 28.2 ± 0.15 | 28.1 ± 0.19 | 28.1 ± 0.21 | 28.3 ± 0.14 | 28.3 ± 0.21 | 28.3 ± 0.22 | 28.2 ± 0.28 | 28.7 ± 0.27 |
| pH | 7.5 ± 0.16 | 8.1 ± 0.14 | 8.3 ± 0.21 | 7.5 ± 0.19 | 8.4 ± 0.18 | 8.4 ± 0.18 | 7.7 ± 0.21 | 8.4 ± 0.16 | 8.4 ± 0.12 |
| Dissolved Oxygen (mg/L) | 5.5 ± 0.18 | 5.9 ± 0.22 | 6.1 ± 0.41 | 5.9 ± 0.36 | 5.9 ± 0.24 | 5.8 ± 0.22 | 5.9 ± 0.23 | 5.9 ± 0.43 | 5.9 ± 0.52 |
| Nitrate (mg/L) | 2.4 ± 0.12 | 2.5 ± 0.15 | 2.4 ± 0.10 | 2.4 ± 0.10 | 2.3 ± 0.06 | 2.5 ± 0.09 | 2.6 ± 0.10 | 2.6 ± 0.12 | 2.7 ± 0.13 |
| Alkalinity (mg/L) | 146 ± 2.20 | 208 ± 3.71 | 206 ± 0.43 | 145 ± 2.76 | 207 ± 3.43 | 207 ± 4.21 | 148 ± 2.89 | 207 ± 3.54 | 206 ± 3.24 |
| Free CO2 (mg/L) | 13.2 ± 0.21 | 0.4 ± 0.12 | 0.9 ± 0.19 | 13.5 ± 0.32 | 0.4 ± 0.12 | 0.8 ± 0.19 | 13.9 ± 0.21 | 0.4 ± 0.23 | 0.7 ± 0.19 |
| Hardness (mg/L) | 114 ± 1.23 | 134 ± 1.76 | 140 ± 0.32 | 115 ± 2.37 | 135 ± 2.23 | 140 ± 1.75 | 115 ± 1.24 | 135 ± 1.65 | 138 ± 2.43 |
| Ammonia (mg/L) | 0.25 ± 0.02 | 0.29 ± 0.03 | 0.24 ± 0.04 | 0.29 ± 0.02 | 0.31 ± 0.04 | 0.33 ± 0.03 | 0.31 ± 0.02 | 0.32 ± 0.03 | 0.33 ± 0.01 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Saha, S.; Chukwuka, A.V.; Mukherjee, D.; Patnaik, L.; Nayak, S.; Dhara, K.; Saha, N.C.; Faggio, C. Chronic Effects of Diazinon® Exposures Using Integrated Biomarker Responses in Freshwater Walking Catfish, Clarias batrachus. Appl. Sci. 2021, 11, 10902. https://doi.org/10.3390/app112210902
Saha S, Chukwuka AV, Mukherjee D, Patnaik L, Nayak S, Dhara K, Saha NC, Faggio C. Chronic Effects of Diazinon® Exposures Using Integrated Biomarker Responses in Freshwater Walking Catfish, Clarias batrachus. Applied Sciences. 2021; 11(22):10902. https://doi.org/10.3390/app112210902
Chicago/Turabian StyleSaha, Shubhajit, Azubuike V. Chukwuka, Dip Mukherjee, Lipika Patnaik, Susri Nayak, Kishore Dhara, Nimai Chandra Saha, and Caterina Faggio. 2021. "Chronic Effects of Diazinon® Exposures Using Integrated Biomarker Responses in Freshwater Walking Catfish, Clarias batrachus" Applied Sciences 11, no. 22: 10902. https://doi.org/10.3390/app112210902
APA StyleSaha, S., Chukwuka, A. V., Mukherjee, D., Patnaik, L., Nayak, S., Dhara, K., Saha, N. C., & Faggio, C. (2021). Chronic Effects of Diazinon® Exposures Using Integrated Biomarker Responses in Freshwater Walking Catfish, Clarias batrachus. Applied Sciences, 11(22), 10902. https://doi.org/10.3390/app112210902