
Phosphorus and Nitrogen Limitation as a Part of the Strategy to Stimulate Microbial Lipid Biosynthesis




Abstract
:1. Introduction
2. Materials and Methods
2.1. Yeast strain and Culture Conditions
2.2. General Analytical Techniques
2.3. Gas Chromatography
2.4. Statistical Analyses
3. Results
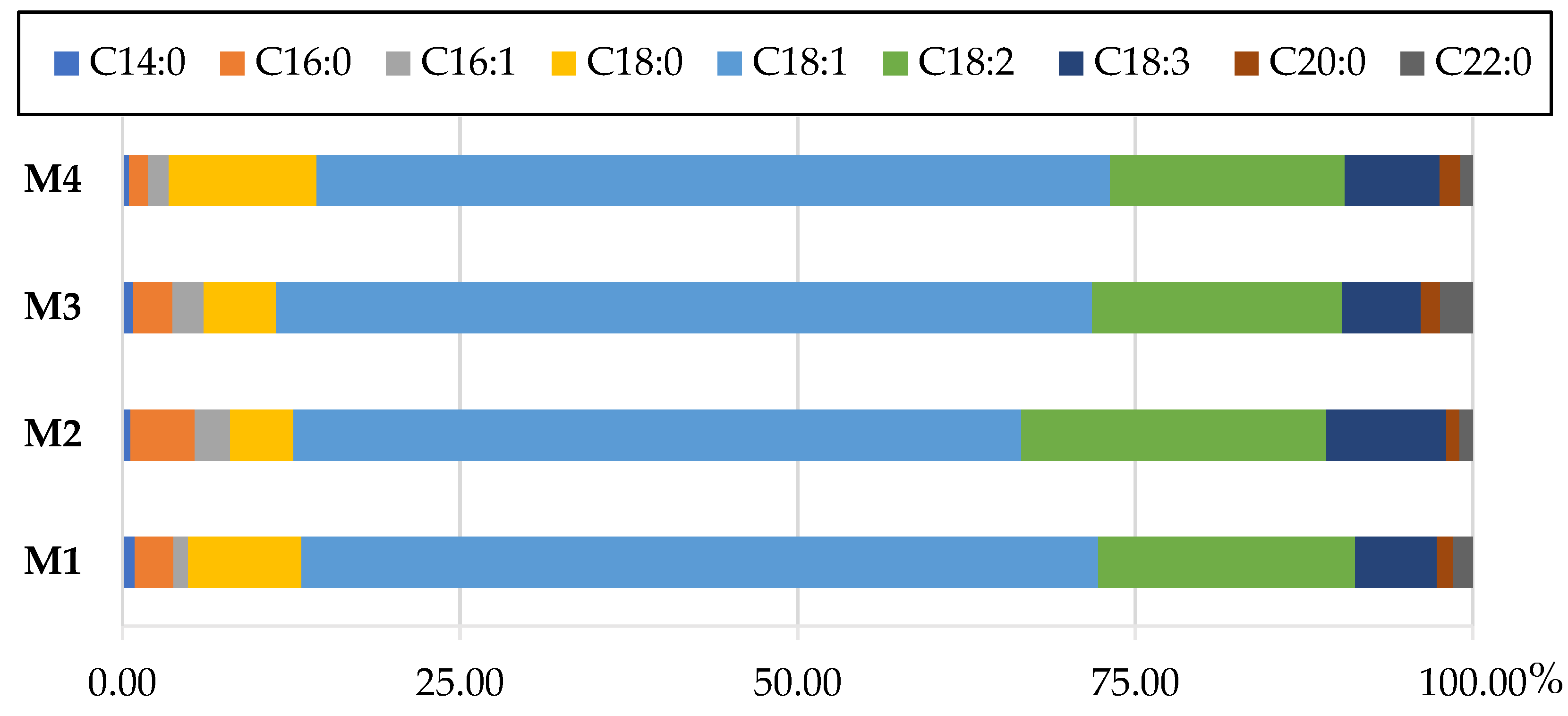
3.1. Flask Scale Experiments
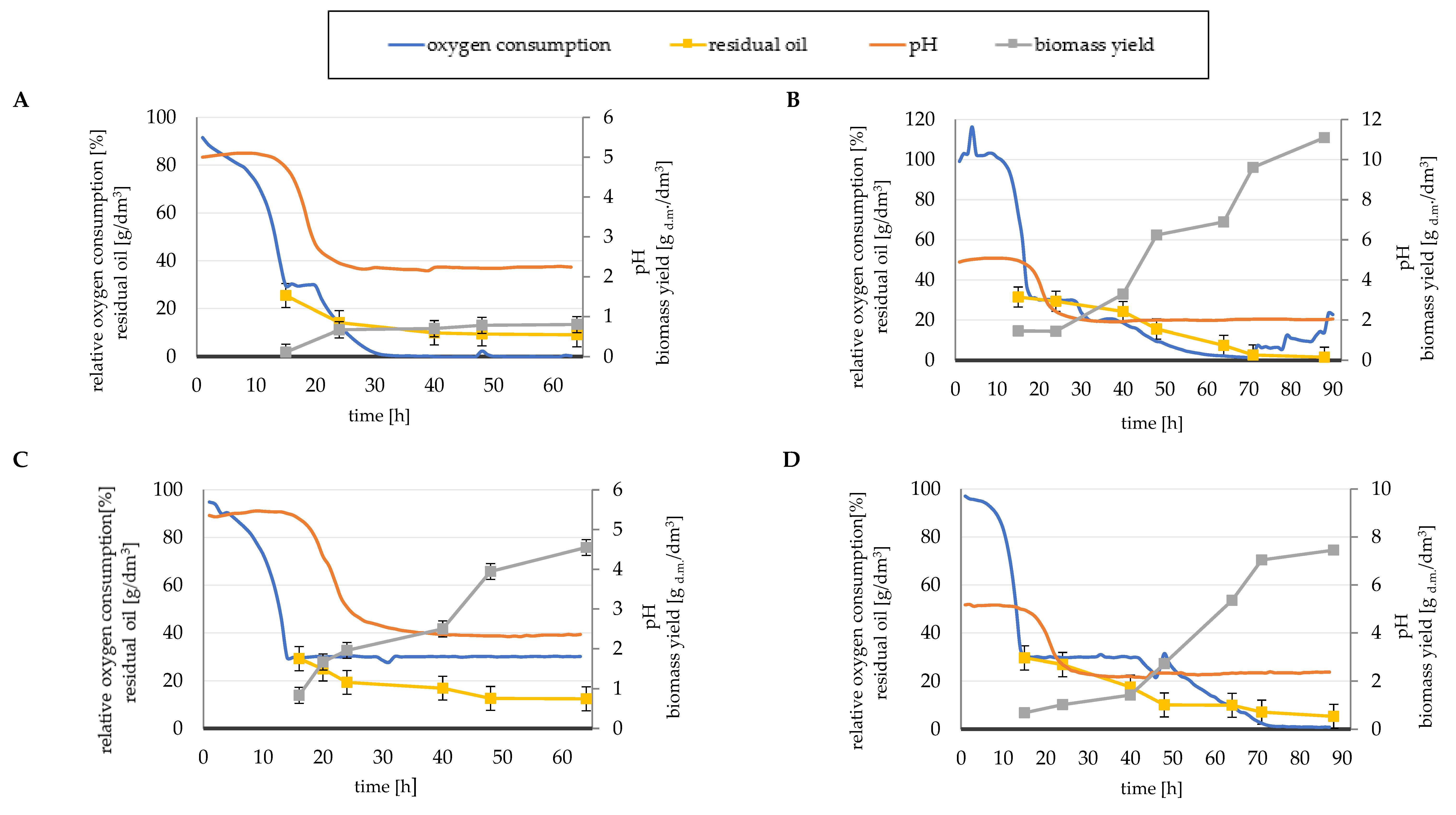
3.2. Bioreactor Scale Experiments
4. Discussion
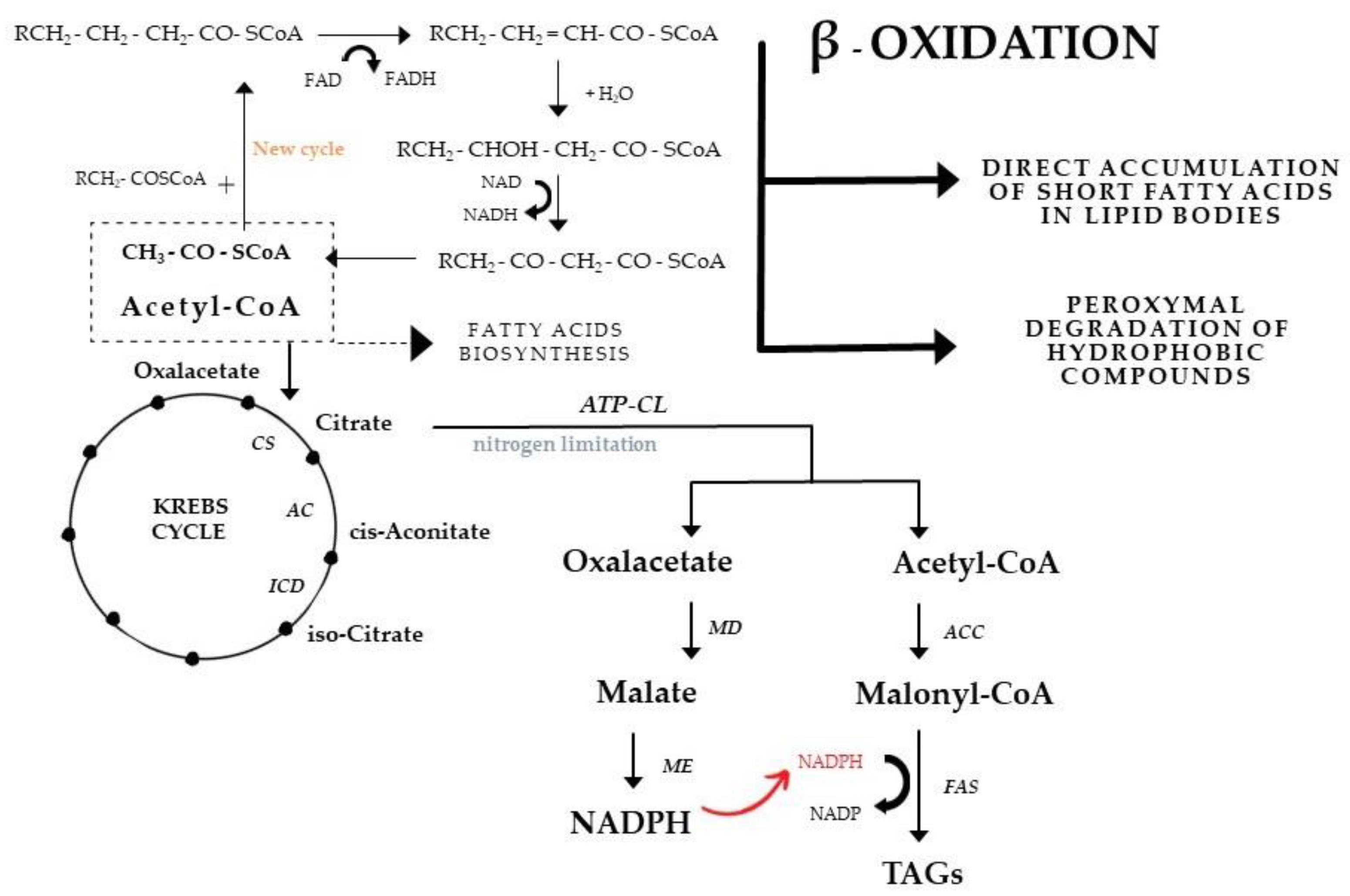
4.1. Lipid Production in Phosphorus-Limited Media
4.2. Lipid Production in Nitrogen-Limited Media
4.3. Cellular Oxygen Demand in Microbial Lipid Production
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Beopoulos, A.; Cescut, J.; Haddouche, R.; Uribelarrea, J.L.; Molina-Jouve, C.; Nicaud, J.M. Yarrowia lipolytica as a model for bio-oil production. Prog. Lipid Res. 2009, 48, 375–387. [Google Scholar] [CrossRef]
- Fabiszewska, A.; Misiukiewicz-Stępień, P.; Paplińska-Goryca, M.; Zieniuk, B.; Białecka-Florjańczyk, E. An insight into storage lipid synthesis by Yarrowia lipolytica yeast relating to lipid and sugar substrates metabolism. Biomolecules 2019, 9, 685–697. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Groenewald, M.; Boekhout, T.; Neuvéglise, C.; Gaillardin, C.; van Dijck, P.W.; Wyss, M. Yarrowia lipolytica: Safety assessment of an oleaginous yeast with a great industrial potential. Crit. Rev. Microbiol. 2014, 40, 187–206. [Google Scholar] [CrossRef] [PubMed]
- Martínez, E.J.; Raghavan, V.; González-Andrés, F.; Gómez, X. New biofuel alternatives: Integrating waste management and single cell oil production. Int. J. Mol. Sci. 2015, 16, 9385–9405. [Google Scholar] [CrossRef]
- Pawar, P.P.; Odaneth, A.A.; Vadgama, R.N.; Lali, A.M. Simultaneous lipid biosynthesis and recovery for oleaginous yeast Yarrowia lipolytica. Biotechnol. Biofuels 2019, 12, 237. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Papanikolaou, S.; Chevalot, I.; Galiotou-Panayotou, M.; Komaitis, M.; Marc, I.; Aggelis, G. Industrial derivative of tallow: A promising renewable substrate for microbial lipid, single-cell protein and lipase production by Yarrowia lipolytica. Electron. J. Biotechnol. 2007, 10, 425–435. [Google Scholar] [CrossRef] [Green Version]
- Cheirsilp, B.; Louhasakul, Y. Industrial wastes as a promising renewable source for production of microbial lipid and direct transesterification of the lipid into biodiesel. Bioresour. Technol. 2013, 142, 329–337. [Google Scholar] [CrossRef]
- Bharathiraja, B.; Sridharan, S.; Sowmya, V.; Yuvaraj, D.; Praveenkumar, R. Microbial oil—A plausible alternate resource for food and fuel application. Bioresour. Technol. 2017, 233, 423–432. [Google Scholar] [CrossRef] [PubMed]
- Subramaniam, R.; Dufreche, S.; Zappi, M.; Bajpai, R. Microbial lipids from renewable resources: Production and characterization. J. Ind. Microbiol. Biotechnol. 2010, 37, 1271–1287. [Google Scholar] [CrossRef]
- Ratledge, C.; Wynn, J.P. The biochemistry and molecular biology of lipid accumulation in oleaginous microorganisms. Adv. Appl. Microbiol. 2002, 51, 1–52. [Google Scholar] [PubMed]
- Lopes, M.; Gomes, A.S.; Silva, C.M.; Belo, I. Microbial lipids and added value metabolites production by Yarrowia lipolytica from pork lard. J. Biotechnol. 2018, 265, 76–85. [Google Scholar] [CrossRef] [Green Version]
- Ji, X.J.; Ledesma-Amaro, R. Microbial lipid biotechnology to produce polyunsaturated fatty acids. Trends Biotechnol. 2020, 38, 832–834. [Google Scholar] [CrossRef]
- Patel, A.; Pruthi, V.; Pruthi, P.A. Synchronized nutrient stress conditions trigger the diversion of CDP-DG pathway of phospholipids synthesis towards de novo TAG synthesis in oleaginous yeast escalating biodiesel production. Energy 2017, 139, 962–974. [Google Scholar] [CrossRef] [Green Version]
- Fabiszewska, A.U.; Kotyrba, D.; Nowak, D. Assortment of carbon sources in medium for Yarrowia lipolytica lipase production: A statistical approach. Ann. Microbiol. 2015, 65, 1495–1503. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Papanikolaou, S.; Muniglia, L.; Chevalot, I.; Aggelis, G.; Marc, I. Accumulation of a cocoa-butter-like lipid by Yarrowia lipolytica cultivated on agro-industrial residues. Curr. Microbiol. 2003, 46, 124–130. [Google Scholar] [CrossRef] [PubMed]
- Gajdos, P.; Nicaud, J.M.; Rossignol, T.; Čertík, M. Single cell oil production on molasses by Yarrowia lipolytica strains overexpressing DGA2 in multicopy. Appl. Microbiol. Biotechnol. 2015, 99, 8065–8074. [Google Scholar] [CrossRef] [PubMed]
- Qin, L.; Liu, L.; Zeng, A.P.; Wei, D. From low-cost substrates to single cell oils synthesized by oleaginous yeasts. Bioresour. Technol. 2017, 245, 1507–1519. [Google Scholar] [CrossRef]
- Lopes, M.; Miranda, S.M.; Belo, I. Microbial valorization of waste cooking oils for valuable compounds production—A review. Crit. Rev. Environ. Sci. Technol. 2020, 50, 2583–2616. [Google Scholar] [CrossRef]
- Liu, X.; Lv, J.; Xu, J.; Zhang, T.; Deng, Y.; He, J. Citric acid production in Yarrowia lipolytica SWJ-1b yeast when grown on waste cooking oil. Appl. Biochem. Biotechnol. 2015, 175, 2347–2356. [Google Scholar] [CrossRef]
- Xiaoyan, L.; Yu, X.; Lv, J.; Xu, J.; Xia, J.; Wu, Z.; Zhang, T.; Deng, Y. A cost-effective process for the coproduction of erythritol and lipase with Yarrowia lipolytica M53 from waste cooking oil. Food Bioprod. Process. 2017, 103, 86–94. [Google Scholar] [CrossRef]
- Nunes, P.M.B.; Martins, A.B.; Brigida, A.I.S.; Rocha Leao, M.H.M.; Amaral, P. Intracellular lipase production by Yarrowia lipolytica using different carbon sources. Chem. Eng. Trans. 2014, 38, 421–426. [Google Scholar]
- Lopes, M.; Miranda, S.M.; Alves, J.M.; Pereira, A.S.; Belo, I. Waste cooking oils as feedstock for lipase and lipid-rich biomass production. Eur. J. Lipid Sci. Technol. 2019, 121, 1800188–1800196. [Google Scholar] [CrossRef] [Green Version]
- El Bialy, H.; Gomaa, O.M.; Azab, K.S. Conversion of oil waste to valuable fatty acids using oleaginous yeast. World J. Microbiol. Biotechnol. 2011, 27, 2791–2798. [Google Scholar] [CrossRef]
- Katre, G.; Joshi, C.; Khot, M.; Zinjarde, S.; RaviKumar, A. Evaluation of single cell oil (SCO) from a tropical marine yeast Yarrowia lipolytica NCIM 3589 as a potential feedstock for biodiesel. AMB Express 2012, 2, 36–49. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tzirita, M.; Papanikolaou, S.; Chatzifragkou, A.; Quilty, B. Waste fat biodegradation and biomodification by Yarrowia lipolytica and a bacterial consortium composed of Bacillus spp. and Pseudomonas putida. Eng. Life Sci. 2018, 18, 932–942. [Google Scholar] [CrossRef] [Green Version]
- Wierzchowska, K.; Zieniuk, B.; Fabiszewska, A. Use of Non-Conventional Yeast Yarrowia lipolytica in Treatment or Upgradation of Hydrophobic Industry Wastes. Waste Biomass Valorization 2021, 1–23. Available online: https://link.springer.com/article/10.1007/s12649-021-01516-9 (accessed on 9 December 2021).
- Ratledge, C. Regulation of lipid accumulation in oleaginous micro-organisms. Biochem. Soc. Trans. 2002, 30, 1047–1050. [Google Scholar] [CrossRef] [Green Version]
- Ratledge, C.; Cohen, Z. Microbial and algal oils: Do they have a future for biodiesel or as commodity oils? Lipid Technol. 2008, 20, 155–160. [Google Scholar] [CrossRef]
- Taskin, M.; Saghafian, A.; Aydogan, M.N.; Arslan, N.P. Microbial lipid production by cold-adapted oleaginous yeast Yarrowia lipolytica B9 in non-sterile whey medium. Biofuels Bioprod. Biorefin. 2015, 9, 595–605. [Google Scholar] [CrossRef]
- Gill, C.O.; Hall, M.J.; Ratledge, C. Lipid accumulation in an oleaginous yeast (Candida 107) growing on glucose in single-stage continuous culture. Appl. Environ. Microbiol. 1977, 33, 231–239. [Google Scholar] [CrossRef] [Green Version]
- Wu, S.; Hu, C.; Jin, G.; Zhao, X.; Zhao, Z.K. Phosphate-limitation mediated lipid production by Rhodosporidium toruloides. Bioresour. Technol. 2010, 101, 6124–6129. [Google Scholar] [CrossRef]
- Bellou, S.; Triantaphyllidou, I.E.; Mizerakis, P.; Aggelis, G. High lipid accumulation in Yarrowia lipolytica cultivated under double limitation of nitrogen and magnesium. J. Biotechnol. 2016, 234, 116–126. [Google Scholar] [CrossRef]
- Kolouchová, I.; Maťátková, O.; Sigler, K.; Masák, J.; Řezanka, T. Lipid accumulation by oleaginous and non-oleaginous yeast strains in nitrogen and phosphate limitation. Folia Microbiol. 2016, 61, 431–438. [Google Scholar] [CrossRef]
- Huang, X.; Luo, H.; Mu, T.; Shen, Y.; Yuan, M.; Liu, J. Enhancement of lipid accumulation by oleaginous yeast through phosphorus limitation under high content of ammonia. Bioresour. Technol. 2018, 262, 9–14. [Google Scholar] [CrossRef]
- Hoarau, J.; Petit, T.; Grondin, I.; Marty, A.; Caro, Y. Phosphate as a limiting factor for the improvement of single cell oil production from Yarrowia lipolytica MUCL 30108 grown on pre-treated distillery spent wash. J. Water Process Eng. 2020, 37, 101392. [Google Scholar] [CrossRef]
- Timoumi, A.; Guillouet, S.E.; Molina-Jouve, C.; Fillaudeau, L.; Gorret, N. Impacts of environmental conditions on product formation and morphology of Yarrowia lipolytica. Appl. Microbiol. Biotechnol. 2018, 102, 3831–3848. [Google Scholar] [CrossRef] [PubMed]
- AOAC. International Official Methods of Analysis, 17th ed.; AOAC International: Gaithersburg, MD, USA, 2000; Volume 2. [Google Scholar]
- Folch, J.; Lees, M.; Stanley, G.S. A simple method for the isolation and purification of total lipides from animal tissues. J. Biol. Chem. 1957, 226, 497–509. [Google Scholar] [CrossRef]
- Michalik, B.; Biel, W.; Lubowicki, R.; Jacyno, E. Chemical composition and biological value of proteins of the yeast Yarrowia lipolytica growing on industrial glycerol. Can. J. Anim. Sci. 2014, 94, 99–104. [Google Scholar] [CrossRef] [Green Version]
- Papanikolaou, S.; Aggelis, G. Lipids of oleaginous yeasts. Part I: Biochemistry of single cell oil production. Eur. J. Lipid Sci. Technol. 2011, 113, 1031–1051. [Google Scholar] [CrossRef]
- Beopoulos, A.; Mrozova, Z.; Thevenieau, F.; Le Dall, M.T.; Hapala, I.; Papanikolaou, S.; Chardot, T.; Nicaud, J.M. Control of lipid accumulation in the yeast Yarrowia lipolytica. Appl. Environ. Microbiol. 2008, 74, 7779–7789. [Google Scholar] [CrossRef] [Green Version]
- Tsigie, Y.A.; Wang, C.Y.; Truong, C.T.; Ju, Y.H. Lipid production from Yarrowia lipolytica Po1g grown in sugarcane bagasse hydrolysate. Bioresour. Technol. 2011, 102, 9216–9222. [Google Scholar] [CrossRef] [PubMed]
- Santek, M.I.; Miskulin, E.; Petrovic, M.; Beluhan, S.; Santek, B. Effect of carbon and nitrogen source concentrations on the growth and lipid accumulation of yeast Trichosporon oleaginosus in continuous and batch culture. J. Soc. Chem. Ind. 2017, 92, 1620–1629. [Google Scholar]
- Dobrowolski, A.; Mituła, P.; Rymowicz, W.; Mirończuk, A.M. Efficient conversion of crude glycerol from various industrial wastes into single cell oil by yeast Yarrowia lipolytica. Bioresour. Technol. 2016, 207, 237–243. [Google Scholar] [CrossRef] [PubMed]
- Magdouli, S.; Brar, S.K.; Blais, J.F. Morphology and rheological behaviour of Yarrowia lipolytica: Impact of dissolved oxygen level on cell growth and lipid composition. Process Biochem. 2018, 65, 1–10. [Google Scholar] [CrossRef]
- Alonso, F.O.M.; Oliveira, E.B.L.; Dellamora-Ortiz, G.M.; Pereira-Meirelles, F.V. Improvement of lipase production at different stirring speeds and oxygen levels. Braz. J. Chem. Eng. 2005, 22, 9–18. [Google Scholar] [CrossRef] [Green Version]
- Bellou, S.; Makri, A.; Triantaphyllidou, I.E.; Papanikolaou, S.; Aggelis, G. Morphological and metabolic shifts of Yarrowia lipolytica induced by alteration of the dissolved oxygen concentration in the growth environment. Microbiology 2014, 160, 807–817. [Google Scholar] [CrossRef] [Green Version]
- Beopoulos, A.; Nicaud, J.M. Yeast: A new oil producer? Oléagineux Corps Gras Lipides 2012, 19, 22–28. [Google Scholar] [CrossRef] [Green Version]
- Dulermo, T.; Nicaud, J.M. Involvement of the G3P shuttle and oxidation pathway in the control of TAG synthesis and lipid accumulation in Yarrowia lipolytica. Metab. Eng. 2011, 13, 482–491. [Google Scholar] [CrossRef]
- Blazeck, J.; Hill, A.; Liu, L.; Knight, R.; Miller, J.; Pan, A.; Otoupal, P.; Alper, H.S. Harnessing Yarrowia lipolytica lipogenesis to create a platform for lipid and biofuel production. Nat. Commun. 2014, 5, 3131. [Google Scholar] [CrossRef] [Green Version]
- Zhang, H.; Zhang, L.; Chen, H.; Chen, Y.Q.; Chen, W.; Song, Y.; Ratledge, C. Enhanced lipid accumulation in the yeast Yarrowia lipolytica by over-expression of ATP: Citrate lyase from Mus musculus. J. Biotechnol. 2014, 192, 78–84. [Google Scholar] [CrossRef]
- Wasylenko, T.M.; Ahn, W.S.; Stephanopoulos, G. The oxidative pentose phosphate pathway is the primary source of NADPH for lipid overproduction from glucose in Yarrowia lipolytica. Metab. Eng. 2015, 30, 27–39. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| KH2PO4(g/dm3) | Duration of the Cultivations (Days) | (NH4)2SO4(g/dm3) | |||
| 3 | 4 | 5 | 6 | ||
|---|---|---|---|---|---|
| 3 | 4.0 | 6.0 | 8.0 | 10.0 | |
| 7 | 6.0 | 8.0 | 10 | 4.0 | |
| 9 | 8.0 | 10.0 | 4.0 | 6.0 | |
| 11 | 10.0 | 4.0 | 6.0 | 8.0 | |
| (g/dm3) | M1 | M2 | M3 | M4 |
|---|---|---|---|---|
| KH2PO4 | 3.0 | 3.0 | 3.5 | 7.0 |
| Na2HPO4 | 1.1 | 1.1 | 2.5 | 2.5 |
| (NH4)2SO4 | 3.0 | 8.0 | 10.0 | 4.0 |
| Parameter | Unit | M1 | M2 | M3 | M4 |
|---|---|---|---|---|---|
| Molar ratio in the medium [C/N/P] | mol | 114.9/1.6/1 | 114.9/4.3/1 | 81.6/3.8/1 | 53.1/1/1.07 |
| Initial concentration of carbon source [S] | g/dm3 | 50 | 50 | 50 | 50 |
| Time [t] | h | 63 | 88 | 63 | 88 |
| Biomass yield [X] | gd.m./dm3 | 0.78 | 11.10 | 4.56 | 7.45 |
| Maximum concentration of lipids produced [Lmax] | g/dm3 | 0.37 | 2.32 | 1.49 | 2.24 |
| Conversion yield of biomass per carbon substrate [YX/S] | gd.m./g | 0.0156 | 0.2220 | 0.0912 | 0.1490 |
| Conversion yield of storage lipids per biomass formed [YL/X] | g/gd.m. | 0.4744 | 0.2090 | 0.3267 | 0.3007 |
| Conversion yield of storage lipids per carbon substrate [YL/S] | g/g | 0.0074 | 0.0464 | 0.0298 | 0.0448 |
| Volumetric rate of storage lipids production [qLv] | g/dm3/h | 0.0059 | 0.0256 | 0.0236 | 0.0254 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wierzchowska, K.; Zieniuk, B.; Nowak, D.; Fabiszewska, A. Phosphorus and Nitrogen Limitation as a Part of the Strategy to Stimulate Microbial Lipid Biosynthesis. Appl. Sci. 2021, 11, 11819. https://doi.org/10.3390/app112411819
Wierzchowska K, Zieniuk B, Nowak D, Fabiszewska A. Phosphorus and Nitrogen Limitation as a Part of the Strategy to Stimulate Microbial Lipid Biosynthesis. Applied Sciences. 2021; 11(24):11819. https://doi.org/10.3390/app112411819
Chicago/Turabian StyleWierzchowska, Katarzyna, Bartłomiej Zieniuk, Dorota Nowak, and Agata Fabiszewska. 2021. "Phosphorus and Nitrogen Limitation as a Part of the Strategy to Stimulate Microbial Lipid Biosynthesis" Applied Sciences 11, no. 24: 11819. https://doi.org/10.3390/app112411819
APA StyleWierzchowska, K., Zieniuk, B., Nowak, D., & Fabiszewska, A. (2021). Phosphorus and Nitrogen Limitation as a Part of the Strategy to Stimulate Microbial Lipid Biosynthesis. Applied Sciences, 11(24), 11819. https://doi.org/10.3390/app112411819