
Diauxie Studies in Biogas Production from Gelatin and Adaptation of the Modified Gompertz Model: Two-Phase Gompertz Model



Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Biogas Production Trials
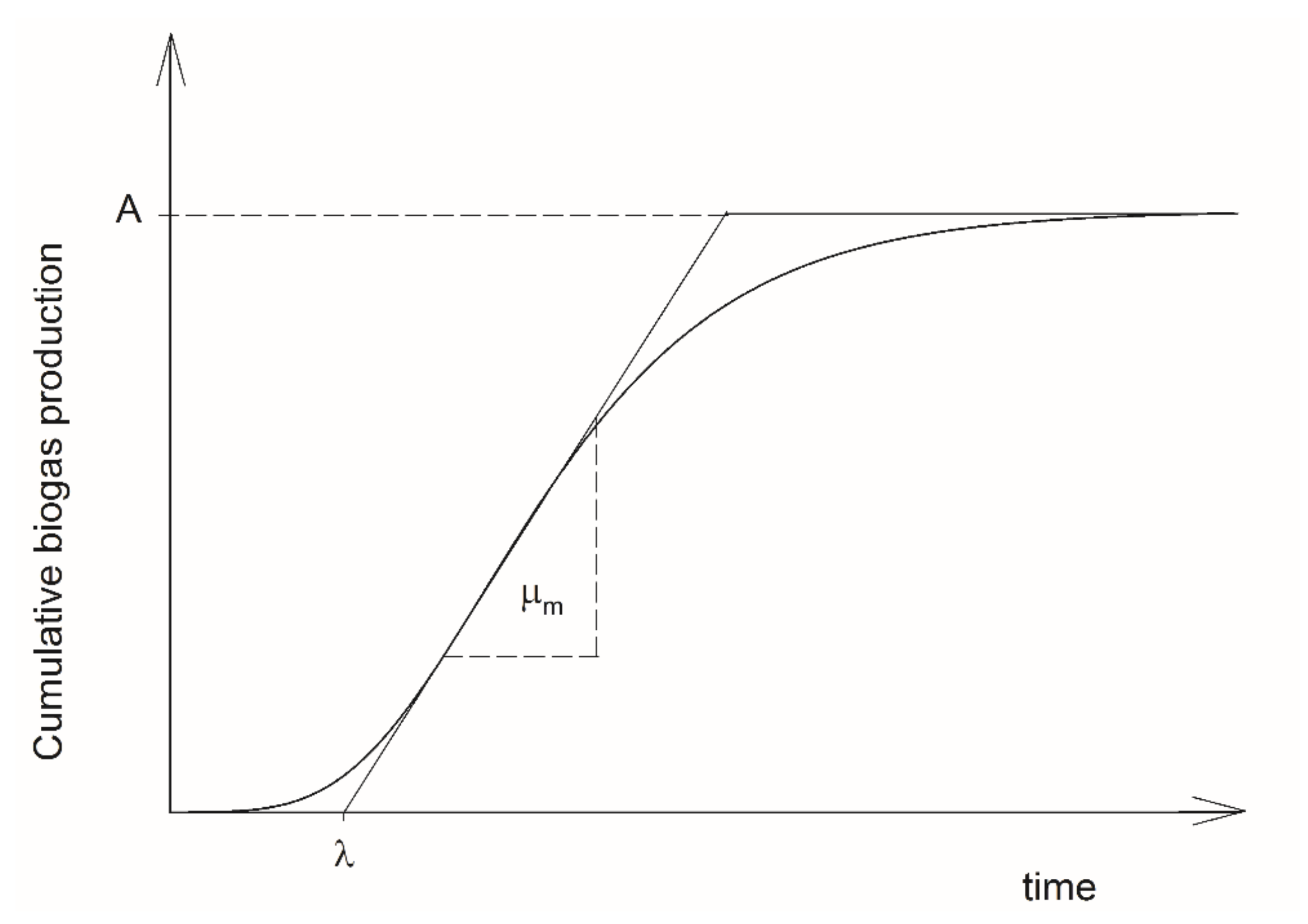
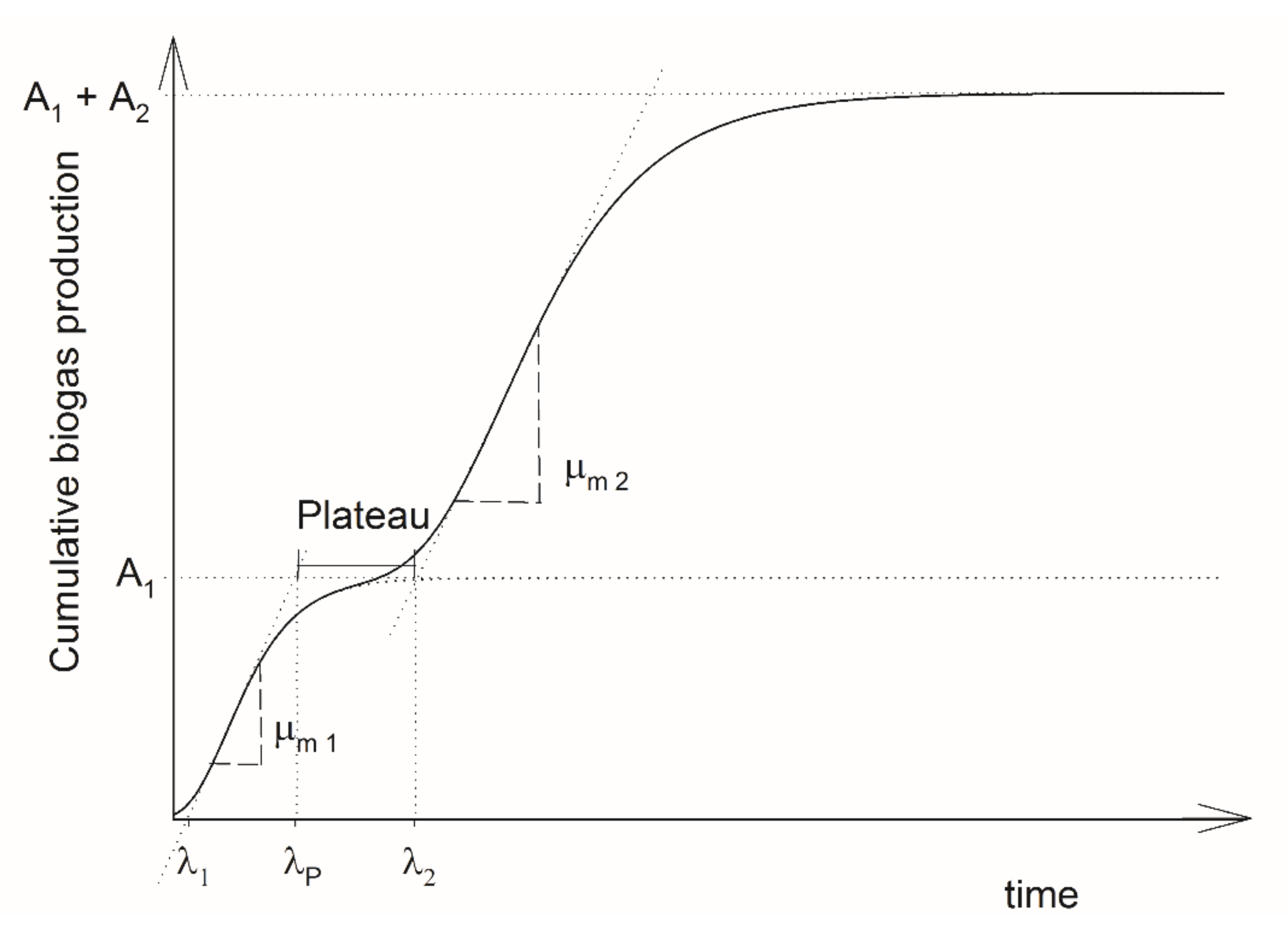
2.3. Two-Phase Gompertz Model
2.4. Statistical Methods
2.5. Analyses of the Final Biomass
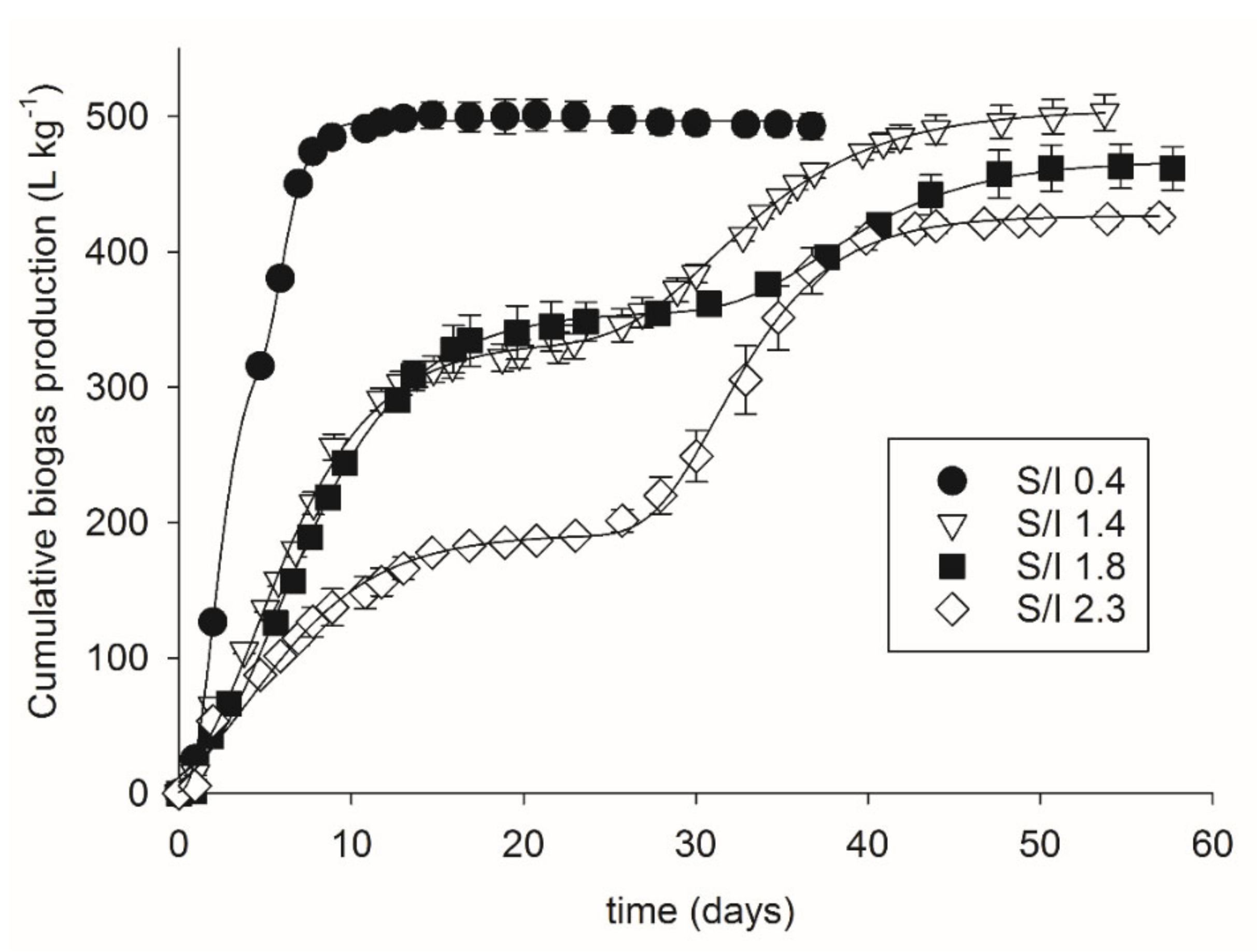
3. Results
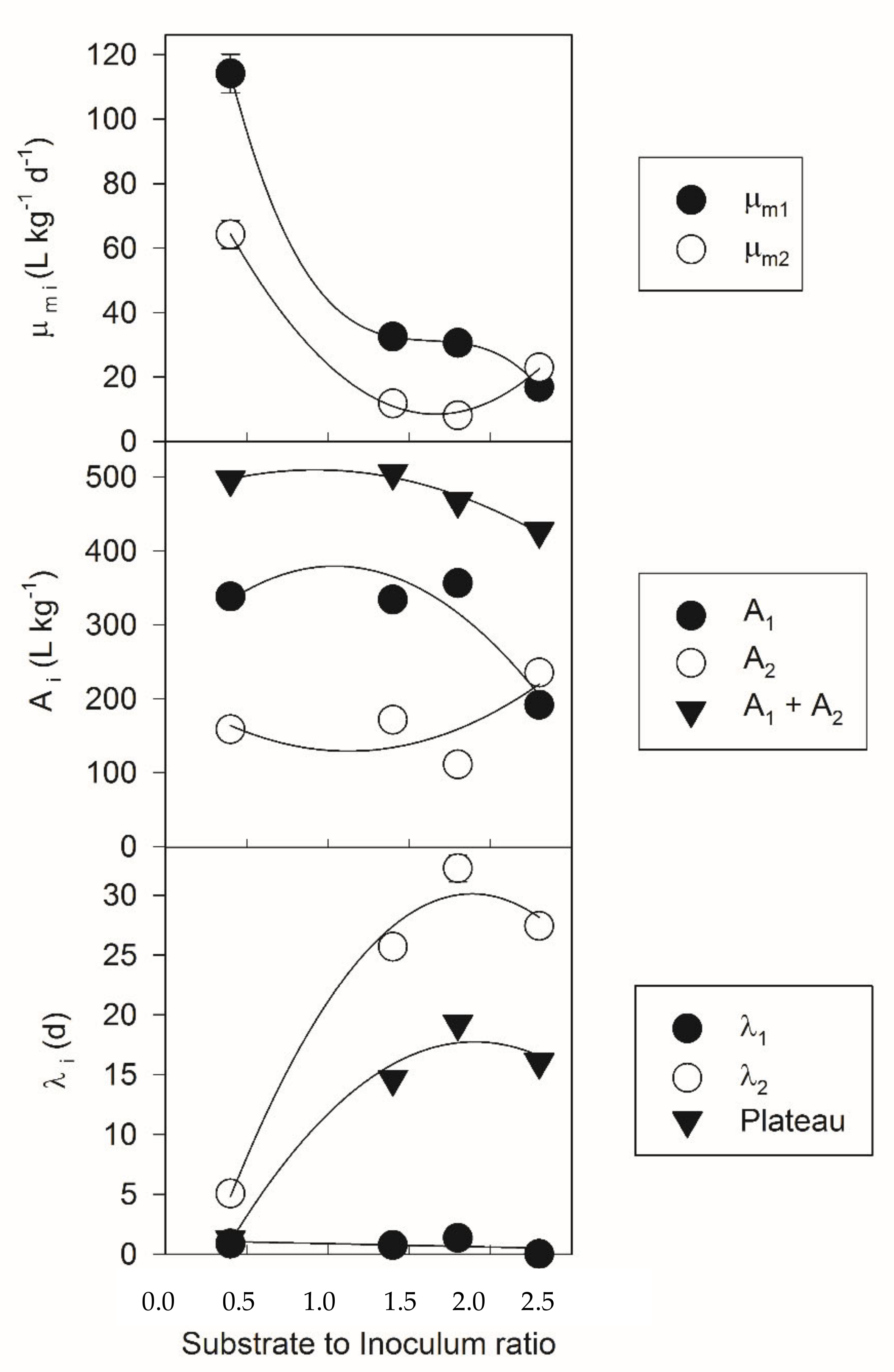
3.1. Two-Phase Gompertz Model
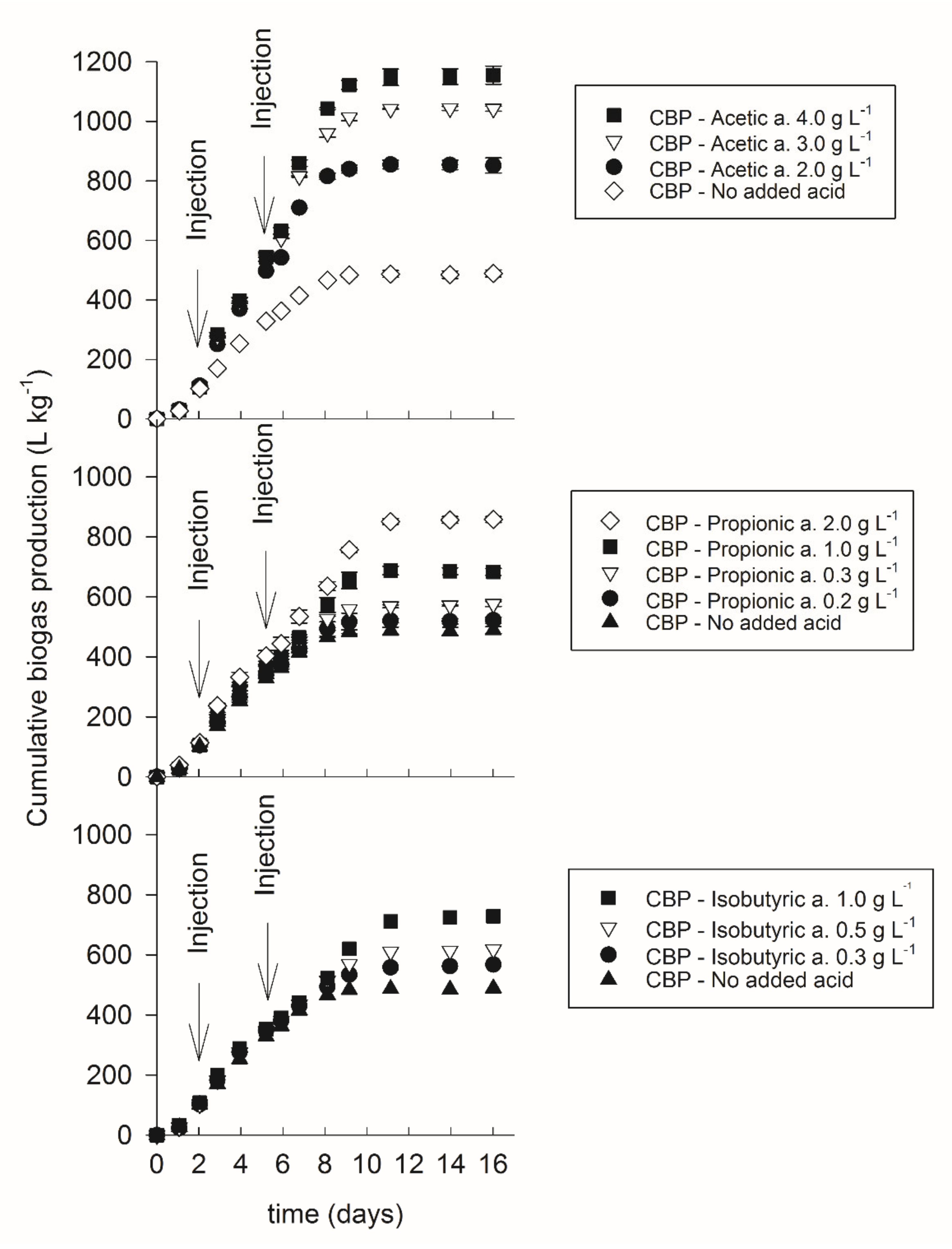
3.2. Inhibition by Volatile Fatty Acids
4. Discussion
5. Conclusions
Author Contributions
Funding
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Tjørve, K.M.C.; Tjørve, E. The Use of Gompertz Models in Growth Analyses, and New Gompertz-Model Approach: An Addition to the Unified-Richards Family. PLoS ONE 2017, 12, e0178691. [Google Scholar] [CrossRef] [PubMed]
- Lay, J.-J.; Li, Y.-Y.; Noike, T. Effect of Moisture Content and Chemical Nature on Methane Fermentation Characteristics of Municipal Solid Wastes. Doboku Gakkai Ronbunshu 1996, 1996, 101–108. [Google Scholar] [CrossRef] [Green Version]
- Zwietering, M.H.; Jongenburger, I.; Rombouts, F.M.; van’t Riet, K. Modeling of the Bacterial Growth Curve. Appl. Environ. Microbiol. 1990, 56, 1875–1881. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luz, F.C.; Cordiner, S.; Manni, A.; Mulone, V.; Rocco, V.; Braglia, R.; Canini, A. Ampelodesmos Mauritanicus Pyrolysis Biochar in Anaerobic Digestion Process: Evaluation of the Biogas Yield. Energy 2018, 161, 663–669. [Google Scholar] [CrossRef]
- Wang, D.; Liu, X.; Zeng, G.; Zhao, J.; Liu, Y.; Wang, Q.; Chen, F.; Li, X.; Yang, Q. Understanding the Impact of Cationic Polyacrylamide on Anaerobic Digestion of Waste Activated Sludge. Water Res. 2018, 130, 281–290. [Google Scholar] [CrossRef] [Green Version]
- Gomes, C.S.; Repke, J.-U.; Meyer, M. Investigation of Different Pre-Treatments of Chromium Leather Shavings to Improve Biogas Production. J. Leather Sci. Eng. 2020, 2, 14. [Google Scholar] [CrossRef]
- Barua, V.B.; Kalamdhad, A.S. Anaerobic Biodegradability Test of Water Hyacinth after Microbial Pretreatment to Optimise the Ideal F/M Ratio. Fuel 2018, 217, 91–97. [Google Scholar] [CrossRef]
- Lo, H.M.; Kurniawan, T.A.; Sillanpää, M.E.T.; Pai, T.Y.; Chiang, C.F.; Chao, K.P.; Liu, M.H.; Chuang, S.H.; Banks, C.J.; Wang, S.C.; et al. Modeling Biogas Production from Organic Fraction of MSW Co-Digested with MSWI Ashes in Anaerobic Bioreactors. Bioresour. Technol. 2010, 101, 6329–6335. [Google Scholar] [CrossRef]
- Shahbaz, M.; Ammar, M.; Zou, D.; Korai, R.M.; Li, X. An Insight into the Anaerobic Co-Digestion of Municipal Solid Waste and Food Waste: Influence of Co-Substrate Mixture Ratio and Substrate to Inoculum Ratio on Biogas Production. Appl. Biochem. Biotechnol. 2019, 187, 1356–1370. [Google Scholar] [CrossRef]
- Lü, F.; Hao, L.; Zhu, M.; Shao, L.; He, P. Initiating Methanogenesis of Vegetable Waste at Low Inoculum-to-Substrate Ratio: Importance of Spatial Separation. Bioresour. Technol. 2012, 105, 169–173. [Google Scholar] [CrossRef]
- Wang, X.; Zhang, L.; Xi, B.; Sun, W.; Xia, X.; Zhu, C.; He, X.; Li, M.; Yang, T.; Wang, P.; et al. Biogas Production Improvement and C/N Control by Natural Clinoptilolite Addition into Anaerobic Co-Digestion of Phragmites Australis, Feces and Kitchen Waste. Bioresour. Technol. 2015, 180, 192–199. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Zhang, R.; He, Y.; Liu, X.; Chen, C.; Liu, G. Thermophilic Solid-State Anaerobic Digestion of Alkaline-Pretreated Corn Stover. Energy Fuels 2014, 28, 3759–3765. [Google Scholar] [CrossRef]
- Ziemiński, K.; Kowalska-Wentel, M. Effect of Enzymatic Pretreatment on Anaerobic Co-Digestion of Sugar Beet Pulp Silage and Vinasse. Bioresour. Technol. 2015, 180, 274–280. [Google Scholar] [CrossRef]
- Ismail, Z.Z.; Talib, A.R. Recycled Medical Cotton Industry Waste as a Source of Biogas Recovery. J. Clean. Prod. 2016, 112, 4413–4418. [Google Scholar] [CrossRef]
- Ma, H.; Li, Z.; Yin, F.; Kao, W.; Yin, Y.; Bai, X. Study on Anaerobic Treatment of Hazardous Steel-Mill Waste Rolling Oil (SmWRO) for Multi-Benefit Disposal Route. Bioresour. Technol. 2014, 151, 106–112. [Google Scholar] [CrossRef] [PubMed]
- Moraes, B.S.; Triolo, J.M.; Lecona, V.P.; Zaiat, M.; Sommer, S.G. Biogas Production within the Bioethanol Production Chain: Use of Co-Substrates for Anaerobic Digestion of Sugar Beet Vinasse. Bioresour. Technol. 2015, 190, 227–234. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Zhang, R.; Liu, X.; Chen, C.; Xiao, X.; Feng, L.; He, Y.; Liu, G. Evaluating Methane Production from Anaerobic Mono- and Co-Digestion of Kitchen Waste, Corn Stover, and Chicken Manure. Energy Fuels 2013, 27, 2085–2091. [Google Scholar] [CrossRef]
- Takeuchi, Y.; Andriamanohiarisoamanana, F.J.; Yasui, S.; Iwasaki, M.; Nishida, T.; Ihara, I.; Umetsu, K. Feasibility Study of a Centralized Biogas Plant Performance in a Dairy Farming Area. J. Mater. Cycles Waste Manag. 2018, 20, 314–322. [Google Scholar] [CrossRef]
- Gomes, C.S.; Repke, J.-U.; Meyer, M. Diauxie during Biogas Production from Collagen-Based Substrates. Renew. Energy 2019, 141, 20–27. [Google Scholar] [CrossRef]
- Lee, C.; Kim, J.; Shin, S.G.; O’Flaherty, V.; Hwang, S. Quantitative and Qualitative Transitions of Methanogen Community Structure during the Batch Anaerobic Digestion of Cheese-Processing Wastewater. Appl. Microbiol. Biotechnol. 2010, 87, 1963–1973. [Google Scholar] [CrossRef]
- Marin, J.; Kennedy, K.J.; Eskicioglu, C. Effect of Microwave Irradiation on Anaerobic Degradability of Model Kitchen Waste. Anaerob. Dig. AD Solid Waste 2010, 30, 1772–1779. [Google Scholar] [CrossRef] [PubMed]
- Misi, S.N.; Forster, C.F. Batch Co-Digestion of Multi-Component Agro-Wastes. Bioresour. Technol. 2001, 80, 19–28. [Google Scholar] [CrossRef]
- Walter, A.; Silberberger, S.; Juárez, M.F.-D.; Insam, H.; Franke-Whittle, I.H. Biomethane Potential of Industrial Paper Wastes and Investigation of the Methanogenic Communities Involved. Biotechnol. Biofuels 2016, 9, 21. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ashekuzzaman, S.M.; Poulsen, T.G. Optimizing Feed Composition for Improved Methane Yield during Anaerobic Digestion of Cow Manure Based Waste Mixtures. Bioresour. Technol. 2011, 102, 2213–2218. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.J.; Kim, S.H. Minimization of Diauxic Growth Lag-Phase for High-Efficiency Biogas Production. J. Environ. Manag. 2017, 187, 456–463. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Xu, Q.; Wang, D.; Wu, Y.; Yang, Q.; Liu, Y.; Wang, Q.; Li, X.; Li, H.; Zeng, G.; et al. Unveiling the Mechanisms of How Cationic Polyacrylamide Affects Short-Chain Fatty Acids Accumulation during Long-Term Anaerobic Fermentation of Waste Activated Sludge. Water Res. 2019, 155, 142–151. [Google Scholar] [CrossRef] [PubMed]
- El-Mashad, H.M. Kinetics of Methane Production from the Codigestion of Switchgrass and Spirulina Platensis Algae. Bioresour. Technol. 2013, 132, 305–312. [Google Scholar] [CrossRef] [PubMed]
- Kafle, G.K.; Kim, S.H. Effects of Chemical Compositions and Ensiling on the Biogas Productivity and Degradation Rates of Agricultural and Food Processing By-Products. Bioresour. Technol. 2013, 142, 553–561. [Google Scholar] [CrossRef]
- DIN EN ISO 4684. Leather–Chemical Tests–Determination of Volatile Matter; German Institute for Standardization: Berlin, Germany, 2005. [Google Scholar]
- DIN EN ISO 4047. Leather-Determination of Sulphated Total Ash and sulphated Water-Insoluble Ash; German Institute for Standardization: Berlin, Germany, 1998. [Google Scholar]
- VDI 4630. Fermentation of Organic Materials, Characterization of the Substrate, Sampling, Collection of Material Data, Fermentation Tests; The Association of German Engineers: Düsseldorf, Germany, 2006. [Google Scholar]
- Stegemann, H. Microdetermination of hydroxyproline with chloramine-T and p-dimethylaminobenzaldehyde. Hoppe. Seylers Z. Physiol. Chem. 1958, 311, 41–45. [Google Scholar] [CrossRef]
- Reich, G. From Collagen to Leather-the Theoretical Background; BASF: Ludwigshafen, Germany, 2007. [Google Scholar]
- FNR. Guide to Biogas from Production to Use; Fachagentur Nachwachsende Rohstoffe e. V. (FNR): Gülzow-Prüzen, Germany, 2012. [Google Scholar]
- Drosg, B.; Braun, R.; Bochmann, G. Analysis and characterisation of biogas feedstocks. In The Biogas Handbook: Science, Production and Applications; Wellinger, A., Murphy, J.D., Baxter, D., Eds.; Woodhead Publishing Limited: Cambridge, UK, 2013; pp. 52–84. ISBN 978-0-85709-498-8. [Google Scholar]
- Kaiser, F.; Metzner, T.; Effenberger, M.; Gronauer, A. Sicherung Der Prozessstabilität in Landwirtschaftlichen Biogasanlagen; LfL-Information; Bayerische Landesanstalt für Landwirtschaft: Freising-Weihenstephan, Germany, 2008. [Google Scholar]
- Deublein, D.; Steinhauser, A. Biogas from Waste and Renewable Resources: An Introduction; John Wiley & Sons, Ltd.: Weinheim, Germany, 2008; ISBN 978-3-527-62170-5. [Google Scholar]
- Lalitha, K.; Swaminathan, K.R.; Padma Bai, R. Kinetics of Biomethanation of Solid Tannery Waste and the Concept of Interactive Metabolic Control. Appl. Biochem. Biotechnol. 1994, 47, 73–87. [Google Scholar] [CrossRef]
- Córdoba, V.; Fernández, M.; Santalla, E. The Effect of Substrate/Inoculum Ratio on the Kinetics of Methane Production in Swine Wastewater Anaerobic Digestion. Environ. Sci. Pollut. Res. 2018, 25, 21308–21317. [Google Scholar] [CrossRef] [PubMed]
- Chang, J.I.; Tsai, J.J.; Wu, K.H. Mathematical Model for Carbon Dioxide Evolution from the Thermophilic Composting of Synthetic Food Wastes Made of Dog Food. Waste Manag. 2005, 25, 1037–1045. [Google Scholar] [CrossRef] [PubMed]
- Vázquez, J.A.; Lorenzo, J.M.; Fuciños, P.; Franco, D. Evaluation of Non-Linear Equations to Model Different Animal Growths with Mono and Bisigmoid Profiles. J. Theor. Biol. 2012, 314, 95–105. [Google Scholar] [CrossRef] [PubMed]
- Fernandes, T.J.; Pereira, A.A.; Muniz, J.A. Double Sigmoidal Models Describing the Growth of Coffee Berries. Ciênc. Rural 2017, 47. [Google Scholar] [CrossRef] [Green Version]
- Mischan, M.M.; Passos, J.R.d.S.; Pinho, S.Z.d.; Carvalho, L.R.d. Inflection and Stability Points of Diphasic Logistic Analysis of Growth. Sci. Agric. 2015, 72, 215–220. [Google Scholar] [CrossRef] [Green Version]
- Pellera, F.-M.; Gidarakos, E. Effect of Substrate to Inoculum Ratio and Inoculum Type on the Biochemical Methane Potential of Solid Agroindustrial Waste. J. Environ. Chem. Eng. 2016, 4, 3217–3229. [Google Scholar] [CrossRef]
- Gomes, C.S.; Repke, J.-U.; Meyer, M. Different Pre-Treatments of Chrome Tanned Leather Waste and Their Use in the Biogas Production. In Proceedings of the XXXIV IULTCS Congress, Chennai, India, 7 February 2017; Universal Publishing: Chennai, India, 2017; p. 62. [Google Scholar]
- The UniProt Consortium. Available online: https://www.uniprot.org (accessed on 26 November 2020).
- Nisman, B. The Stickland Reaction. Bacteriol. Rev. 1954, 18, 16–42. [Google Scholar] [CrossRef]
- Breure, A.M.; Mooijman, K.A.; van Andel, J.G. Protein Degradation in Anaerobic Digestion: Influence of Volatile Fatty Acids and Carbohydrates on Hydrolysis and Acidogenic Fermentation of Gelatin. Appl. Microbiol. Biotechnol. 1986, 24, 426–431. [Google Scholar] [CrossRef]
- Franke-Whittle, I.H.; Walter, A.; Ebner, C.; Insam, H. Investigation into the Effect of High Concentrations of Volatile Fatty Acids in Anaerobic Digestion on Methanogenic Communities. Waste Manag. 2014, 34, 2080–2089. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Water Content (%) 1 | Organic Matter (%) 1 | |
|---|---|---|
| Substrate—Gelatin | 90.5 ± 0.0 | 99.0 ± 0.0 |
| Inoculum—Mesophilic anaerobic sludge | 96.6 ± 0.0 | 51.0 ± 2.5 |
| S/I | A1/A2 | λ1/Plateau | µm1/µm2 |
|---|---|---|---|
| 0.4 | 10.7 | 0.6 | 2.0 |
| 1.4 | 2.0 | 0.1 | 2.8 |
| 1.8 | 3.2 | 0.1 | 3.8 |
| 2.3 | 0.8 | 0.0 | 0.7 |
| S/I | Organic Matter (%) 1 | Hydroxyproline (%) 2 | Degradation Degree (%) | pH |
|---|---|---|---|---|
| 0.4 | 46.0 ± 1.1 | 0.04 ± 0.01 | 98.8 | 8.3 |
| 1.4 | 48.8 ± 0.2 | 0.05 ± 0.01 | 99.6 | 8.4 |
| 1.8 | 45.5 ± 0.2 | n.d. | n.d. | 8.5 |
| 2.3 | 49.1 ± 0.4 | 0.06 ± 0.00 | 99.6 | 8.4 |
| Shapiro–Wilk Test | Pearson Correlation | Linear Regression Analysis | ||||
|---|---|---|---|---|---|---|
| W | p | r | p | Equation | R2 | |
| Injection of acetic acid | 0.938 | 0.64 | 0.996 | 0.004 | y = 170.42 x + 501.21 | 0.993 |
| Injection of propionic acid | 0.905 | 0.439 | 0.995 | 0 | y = 181.48 x + 498.31 | 0.991 |
| Injection of isobutyric acid | 0.991 | 0.96 | 0.999 | 0.001 | y = 238.79 x + 493.39 | 0.998 |
| Injection of Acetic Acid | ||||
|---|---|---|---|---|
| Acid Conc. (g L−1) | Organic Matter (%) 1 | Hydroxyproline(%) 1 | Degradation Degree (%) | Biogas Formation Potential (L kg−1) |
| 4 | 50.7 ± 0.7 | 0.05 ± 0.01 | 98.2 | 1156 ± 30 |
| 3 | 50.1 ± 0.2 | 0.05 ± 0.00 | 98.2 | 1042 ± 8 |
| 2 | 50.4 ± 0.2 | 0.06 ± 0.00 | 97.8 | 852 ± 25 |
| Injection of Propionic Acid | ||||
| 2 | 50.6 ± 0.3 | 0.05 ± 0.00 | 98.2 | 858 ± 10 |
| 1 | 50.4 ± 0.2 | 0.05 ± 0.01 | 98.2 | 683 ± 12 |
| 0.3 | 49.9 ± 0.3 | 0.06 ± 0.01 | 98.3 | 576 ± 8 |
| 0.2 | 49.3 ± 0.3 | 0.02 ± 0.00 | 99.6 | 522 ± 10 |
| Injection of Isobutyric Acid | ||||
| 1 | 49.5 ± 0.0 | 0.04 ± 0.00 | 98.7 | 729 ± 49 |
| 0.5 | 49.6 ± 0.5 | 0.02 ± 0.01 | 99.2 | 618 ± 20 |
| 0.3 | 49.9 ± 0.2 | 0.03 ± 0.01 | 99.1 | 568 ± 34 |
| No Added Acid | ||||
| 0 | 49.8 ± 0.3 | 0.01 ± 0.00 | 99.6 | 489 ± 12 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gomes, C.S.; Strangfeld, M.; Meyer, M. Diauxie Studies in Biogas Production from Gelatin and Adaptation of the Modified Gompertz Model: Two-Phase Gompertz Model. Appl. Sci. 2021, 11, 1067. https://doi.org/10.3390/app11031067
Gomes CS, Strangfeld M, Meyer M. Diauxie Studies in Biogas Production from Gelatin and Adaptation of the Modified Gompertz Model: Two-Phase Gompertz Model. Applied Sciences. 2021; 11(3):1067. https://doi.org/10.3390/app11031067
Chicago/Turabian StyleGomes, Carolina Scaraffuni, Martin Strangfeld, and Michael Meyer. 2021. "Diauxie Studies in Biogas Production from Gelatin and Adaptation of the Modified Gompertz Model: Two-Phase Gompertz Model" Applied Sciences 11, no. 3: 1067. https://doi.org/10.3390/app11031067
APA StyleGomes, C. S., Strangfeld, M., & Meyer, M. (2021). Diauxie Studies in Biogas Production from Gelatin and Adaptation of the Modified Gompertz Model: Two-Phase Gompertz Model. Applied Sciences, 11(3), 1067. https://doi.org/10.3390/app11031067