
Halophyte Plants and Their Residues as Feedstock for Biogas Production—Chances and Challenges



Abstract
:1. Introduction
2. Natural Habitats of Halophytes, Their Role and Adaptive Mechanisms
2.1. Natural Habitats of Halophytes
2.2. Salt Adaptation Mechanisms
3. The Potential Use of Halophytes
3.1. Valuable Ingredients and Products of Halophytes
3.2. What Makes Halophytes Interesting as a Biomass Resource for Biogas Production?
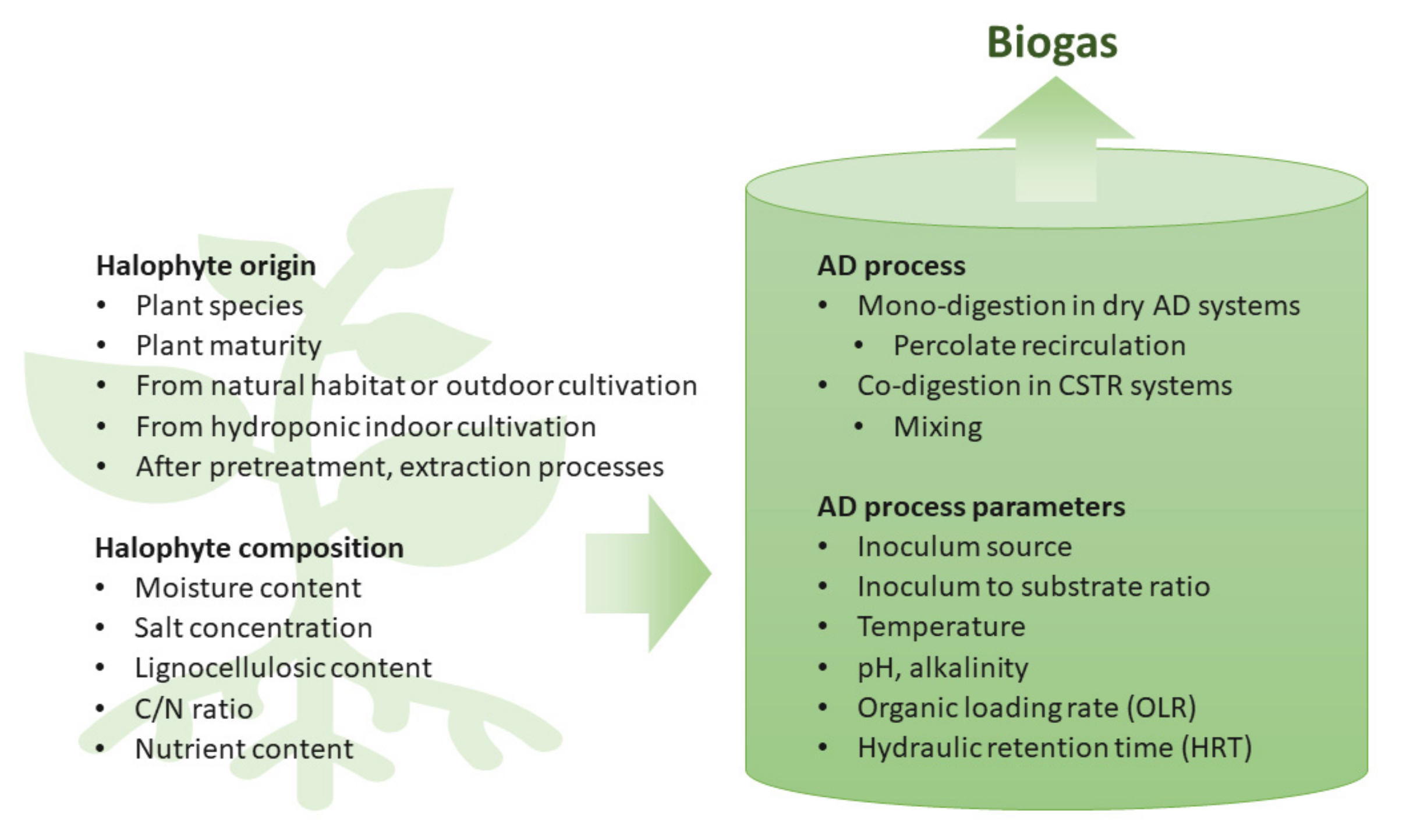
4. Cultivation of Halophytes for Biogas Production
5. Halophyte Composition
5.1. Organic Matter Content
5.2. Ash Content
5.3. Lignocellulose Content
5.4. Carbohydrate Content
5.5. Lignin Content
5.6. Extractives
6. Conversion of Halophytes into Biogas
6.1. The AD Process of Halophytes
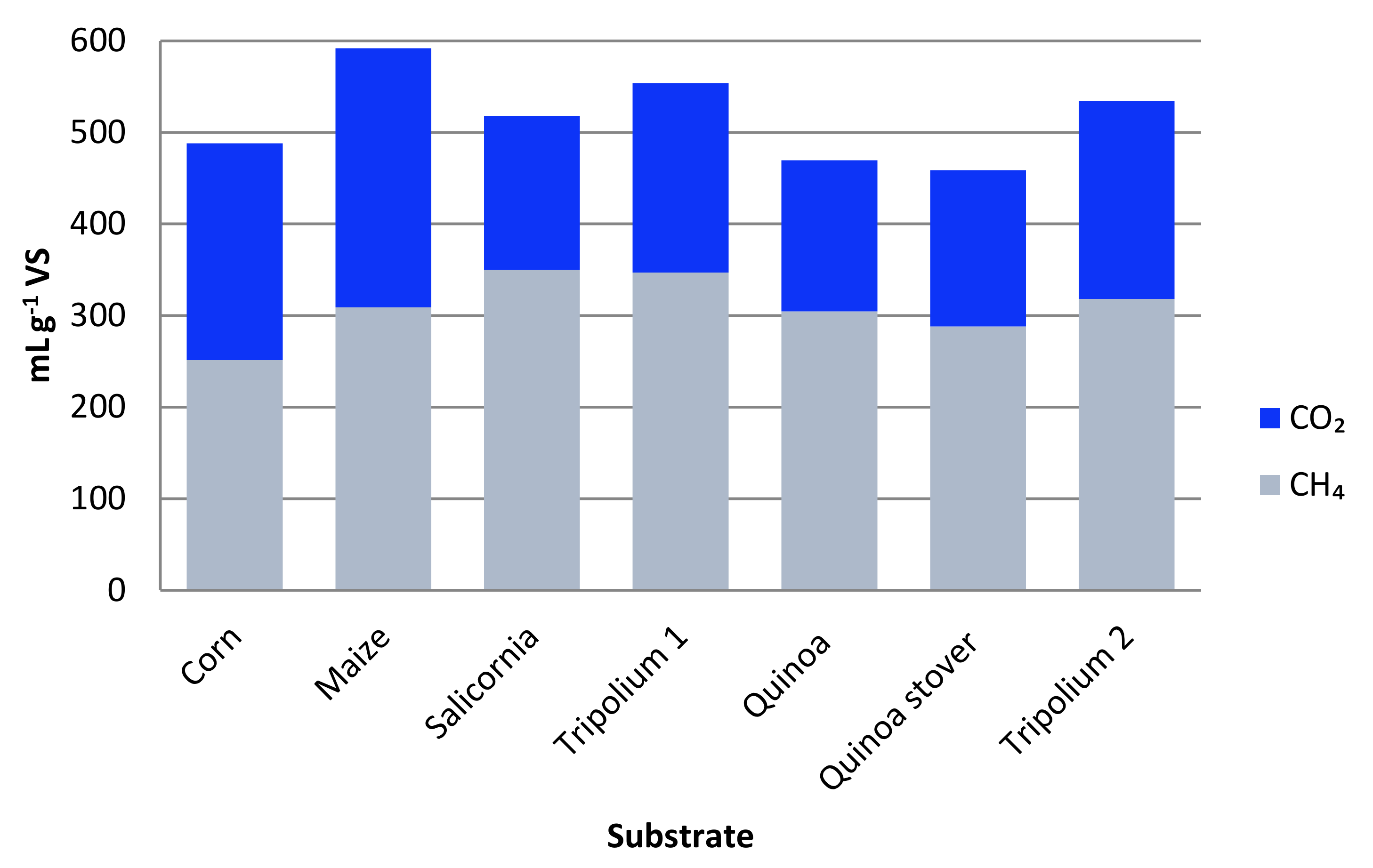
6.2. The Biogas Potential of Halophytes
6.3. Inhibiting Effects of Halophytes on the AD Process
6.3.1. Optimization of AD Microorganism Composition
6.3.2. Co-Digestion of Halophytes
7. Full-Scale Application of Biogas Production from Halophytes
- CSTR (continuous stirred tank reactor) systems for the treatment of slurries, such as manure or sewage sludge, with an optional admixing of other solid organic substrates;
- Dry AD systems for mono digestion of solid biomass or organic waste, such as OFMSW (organic fraction of municipal solid waste), with a total solids (TS) content of more than 15%;
- High-rate reactor systems of the UASB (up-flow anaerobic sludge bed) or EGSB (expanded granular sludge bed) type for the anaerobic treatment of wastewater.
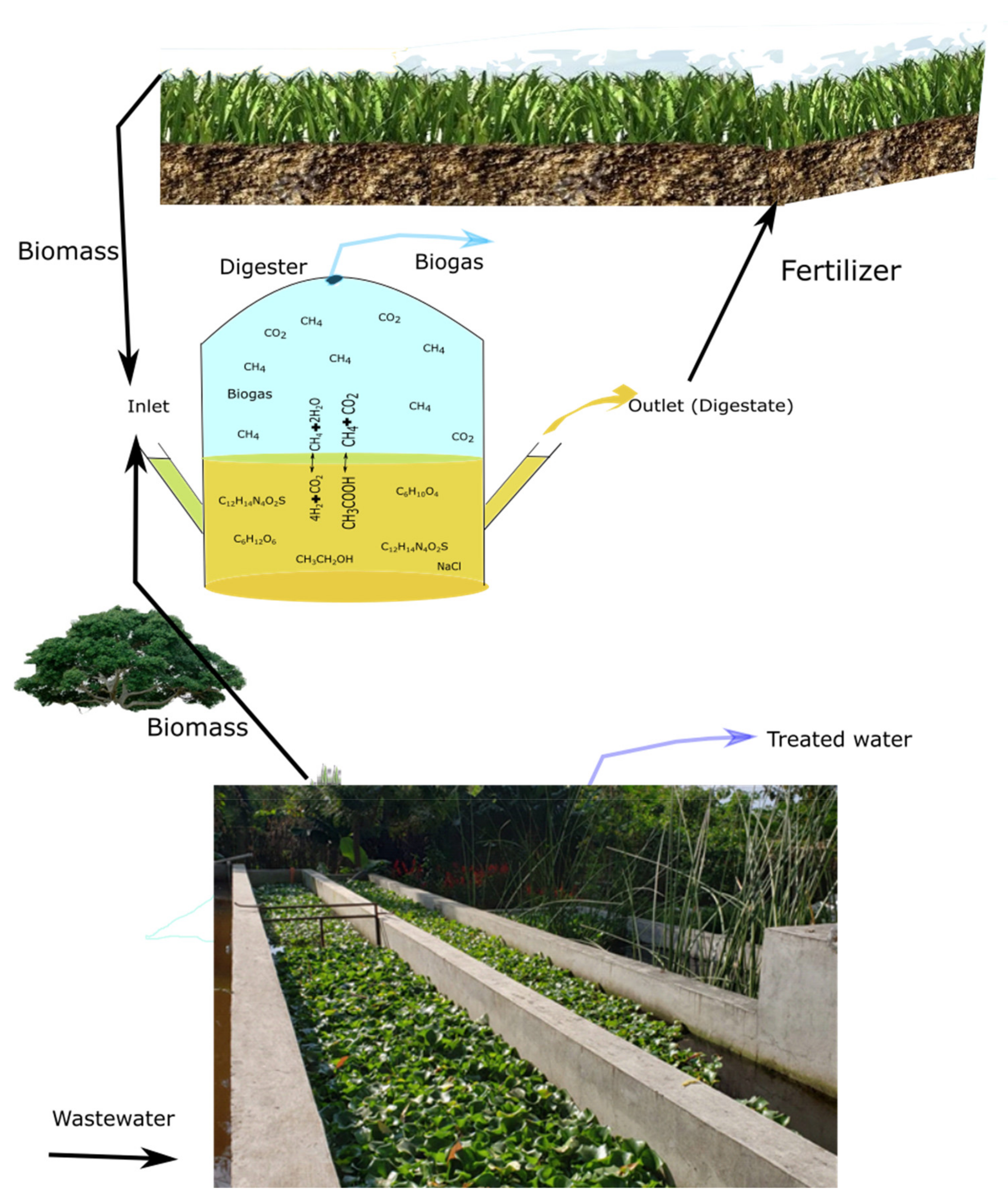
8. The Biogas Process as Part of a Biorefinery for Conversion of Halophyte Biomass
9. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Abideen, Z.; Ansari, R.; Khan, M.A. Halophytes: Potential Source of Ligno-Cellulosic Biomass for Ethanol Production. Biomass Bioenerg. 2011, 35, 1818–1822. [Google Scholar] [CrossRef]
- Smith, C. One-Fifth of Global Farm Soil Degraded by Salt—Our World. Available online: https://ourworld.unu.edu/en/one-fifth-of-global-farm-soil-degraded-by-salt (accessed on 26 February 2021).
- Sharma, R.; Wungrampha, S.; Singh, V.; Pareek, A.; Sharma, M.K. Halophytes As Bioenergy Crops. Front. Plant Sci. 2016, 7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Flowers, T.J.; Colmer, T.D. Salinity Tolerance in Halophytes. New Phytol. 2008, 179, 945–963. [Google Scholar] [CrossRef] [PubMed]
- IEA Bioenergy 28th Update. Biomass Bioenerg. 2007, 31, I–XI. [CrossRef]
- Kapler, A. Habitats of halophytes. In Halophytes and Climate Change: Adaptive Mechanisms and Potential Uses; Hasanuzzaman, M., Shabala, S., Fujita, M., Eds.; CABI: Wallingford, UK, 2019; pp. 19–37. ISBN 9781786394330. [Google Scholar]
- Colmer, T.D.; Flowers, T.J. Flooding Tolerance in Halophytes. New Phytol. 2008, 179, 964–974. [Google Scholar] [CrossRef] [PubMed]
- Glenn, E.P.; Brown, J.J.; Blumwald, E. Salt Tolerance and Crop Potential of Halophytes. Crit. Rev. Plant Sci. 1999, 18, 227–255. [Google Scholar] [CrossRef]
- Buhmann, A.; Papenbrock, J. Biofiltering of Aquaculture Effluents by Halophytic Plants: Basic Principles, Current Uses and Future Perspectives. Environ. Exp. Bot. 2013, 92, 122–133. [Google Scholar] [CrossRef]
- Sun, W.Q.; Li, X.; Ong, B. Preferential Accumulation of D-Pinitol in Acrostichum aureum Gametophytes in Response to Salt Stres. Physiol. Plant. 1999, 105, 51–57. [Google Scholar] [CrossRef]
- Rezaie, E.E.; Haghighikhah, M.; Ghorbani, S.; Kafi, M. Effect of Seed Priming on Seed Germination Properties of Two Medicinal Species in the Presence of Salinity. J. Med. Plant Res. 2013, 7, 1234–1238. [Google Scholar]
- Invasive Species Compendium. Available online: https://www.cabi.org/isc (accessed on 13 March 2021).
- Wilson, C. Growth Stage Modulates Salinity Tolerance of New Zealand Spinach (Tetragonia tetragonioides, Pall.) and Red Orach (Atriplex hortensis L.). Ann. Bot. 2000, 85, 501–509. [Google Scholar] [CrossRef] [Green Version]
- Hasanuzzaman, M.; Nahar, K.; Alam, M.M.; Bhowmik, P.C.; Hossain, M.A.; Rahman, M.M.; Prasad, M.N.V.; Ozturk, M.; Fujita, M. Potential Use of Halophytes to Remediate Saline Soils. BioMed Res. Int. 2014, 2014, 1–12. [Google Scholar] [CrossRef]
- O’Leary, J.W.; Glenn, E.P.; Watson, M.C. Agricultural Production of Halophytes Irrigated with Seawater. Plant Soil 1985, 89, 311–321. [Google Scholar] [CrossRef]
- Zhu, Z.; Chen, J.; Zheng, H.-L. Physiological and Proteomic Characterization of Salt Tolerance in a Mangrove Plant, Bruguiera gymnorrhiza (L.) Lam. Tree Physiol. 2012, 32, 1378–1388. [Google Scholar] [CrossRef] [Green Version]
- Varone, L.; Catoni, R.; Bonito, A.; Gini, E.; Gratani, L. Photochemical Performance of Carpobrotus edulis in Response to Various Substrate Salt Concentrations. S. Afr. J. Bot. 2017, 111, 258–266. [Google Scholar] [CrossRef]
- Adolf, V.I.; Jacobsen, S.-E.; Shabala, S. Salt Tolerance Mechanisms in Quinoa (Chenopodium quinoa Willd.). Environ. Exp. Bot. 2013, 92, 43–54. [Google Scholar] [CrossRef]
- Nawaz, I.; Iqbal, M.; Bliek, M.; Schat, H. Salt and Heavy Metal Tolerance and Expression Levels of Candidate Tolerance Genes among Four Extremophile Cochlearia Species with Contrasting Habitat Preferences. Sci. Total Environ. 2017, 584–585, 731–741. [Google Scholar] [CrossRef]
- De Vos, A.C. Sustainable Exploitation of Saline Resources: Ecology, Ecophysiology and Cultivation of Potential Halophyte Crops; Wohrmann Print Service: Zutphen, The Netherlands, 2011; ISBN 9789085707462. [Google Scholar]
- De Vos, A.C.; Broekman, R.; Groot, M.P.; Rozema, J. Ecophysiological Response of Crambe maritima to Airborne and Soil-Borne Salinity. Ann. Bot. 2010, 105, 925–937. [Google Scholar] [CrossRef] [Green Version]
- Ben-Hamed, K.; Debez, A.; Chibani, F.; Abdelly, C. Salt Response of Crithmum maritimum, an Oleaginous Halophyte. Trop. Ecol. 2004, 45, 151–159. [Google Scholar]
- Ben Amor, N.; Ben Hamed, K.; Debez, A.; Grignon, C.; Abdelly, C. Physiological and Antioxidant Responses of the Perennial Halophyte Crithmum maritimum to Salinity. Plant Sci. 2005, 168, 889–899. [Google Scholar] [CrossRef]
- Rozema, J. An Eco-Physiological Investigation into the Salt Tolerance of Glaux maritima L. Acta Bot. Neerl. 1975, 24, 407–416. [Google Scholar] [CrossRef]
- Tardío, J.; Pardo-De-Santayana, M.; Morales, R. Ethnobotanical Review of Wild Edible Plants in Spain. Bot. J. Linn. Soc. 2006, 152, 27–71. [Google Scholar] [CrossRef]
- Zurayk, R.A.; Baalbaki, R. Inula Crithmoides: A Candidate Plant for Saline Agriculture. Arid Soil Res. Rehabil. 1996, 10, 213–223. [Google Scholar] [CrossRef]
- Zhang, M.; Zhang, H.; Zheng, J.-X.; Mo, H.; Xia, K.-F.; Jian, S.-G. Functional Identification of Salt-Stress-Related Genes Using the FOX Hunting System from Ipomoea pes-caprae. J. Med. Plant Res. 2018, 19, 3446. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Al Hassan, M.; Chaura, J.; López-Gresa, M.P.; Borsai, O.; Daniso, E.; Donat-Torres, M.P.; Mayoral, O.; Vicente, O.; Boscaiu, M. Native-Invasive Plants vs. Halophytes in Mediterranean Salt Marshes: Stress Tolerance Mechanisms in Two Related Species. Front. Plant Sci. 2016, 7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Agarie, S.; Shimoda, T.; Shimizu, Y.; Baumann, K.; Sunagawa, H.; Kondo, A.; Ueno, O.; Nakahara, T.; Nose, A.; Cushman, J.C. Salt Tolerance, Salt Accumulation, and Ionic Homeostasis in an Epidermal Bladder-Cell-Less Mutant of the Common Ice Plant Mesembryanthemum crystallinum. J. Exp. Bot. 2007, 58, 1957–1967. [Google Scholar] [CrossRef] [PubMed]
- Herppich, W.B.; Huyskens-Keil, S.; Schreiner, M. Effects of Saline Irrigation on Growth, Physiology and Quality of Mesembryanthemum crystallinum L., a Rare Vege-Table Crop. J. Appl.Bot. Food Qual. 2008, 82, 47–54. [Google Scholar]
- Theerawitaya, C.; Samphumphaung, T.; Cha-um, S.; Yamada, N.; Takabe, T. Responses of Nipa Palm (Nypa fruticans) Seedlings, a Mangrove Species, to Salt Stress in Pot Culture. Flora Morphol. Distrib. Funct. Ecol. Plants 2014, 209, 597–603. [Google Scholar] [CrossRef]
- Koyro, H.-W. Effect of Salinity on Growth, Photosynthesis, Water Relations and Solute Composition of the Potential Cash Crop Halophyte Plantago coronopus (L.). Environ. Exp. Bot. 2006, 56, 136–146. [Google Scholar] [CrossRef]
- Simopoulos, A.P. Omega-3 Fatty Acids and Antioxidants in Edible Wild Plants. Biol. Res. 2004, 37. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yazici, I.; Türkan, I.; Sekmen, A.H.; Demiral, T. Salinity Tolerance of Purslane (Portulaca oleracea L.) Is Achieved by Enhanced Antioxidative System, Lower Level of Lipid Peroxidation and Proline Accumulation. Environ. Exp. Bot. 2007, 61, 49–57. [Google Scholar] [CrossRef]
- Kodikara, K.A.S.; Jayatissa, L.P.; Huxham, M.; Dahdouh-Guebas, F.; Koedam, N. The Effects of Salinity on Growth and Survival of Mangrove Seedlings Changes with Age. Acta Bot. Bras. 2017, 32, 37–46. [Google Scholar] [CrossRef] [Green Version]
- Ventura, Y.; Wuddineh, W.A.; Myrzabayeva, M.; Alikulov, Z.; Khozin-Goldberg, I.; Shpigel, M.; Samocha, T.M.; Sagi, M. Effect of Seawater Concentration on the Productivity and Nutritional Value of Annual Salicornia and Perennial Sarcocornia Halophytes as Leafy Vegetable Crops. Sci. Hortic. 2011, 128, 189–196. [Google Scholar] [CrossRef]
- Reimann, C.; Breckle, S.-W. Salt Tolerance and Ion Relations of Salsola kali L.: Differences between ssp. tragus (L.) Nyman and ssp. ruthenica (Iljin) Soó. New Phytol. 1995, 130, 37–45. [Google Scholar] [CrossRef]
- Messeddi, D.; Sleimi, N.; Abdelly, C. Salt tolerance in Sesuvium portulacastrum. In Plant Nutrition: Food Security and Sustainability of Agro-Ecosystems through Basic and Applied Research; Horst, W.J., Schenk, M.K., Bürkert, A., Claassen, N., Flessa, H., Frommer, W.B., Goldbach, H., Olfs, H.-W., Römheld, V., Sattelmacher, B., et al., Eds.; Developments in Plant and Soil Sciences; Springer: Dordrecht, The Netherlands, 2001; pp. 406–407. ISBN 9780306476242. [Google Scholar]
- Słupski, J.; Achrem-Achremowicz, J.; Lisiewska, Z.; Korus, A. Effect of Processing on the Amino Acid Content of New Zealand Spinach (Tetragonia tetragonioides Pall. Kuntze): Amino Acids in New Zealand Spinach. Int. J. Food Sci. Technol. 2010, 45, 1682–1688. [Google Scholar] [CrossRef]
- Boestfleisch, C.; Papenbrock, J. Changes in Secondary Metabolites in the Halophytic Putative Crop Species Crithmum maritimum L., Triglochin maritima L. and Halimione portulacoides (L.) Aellen as Reaction to Mild Salinity. PLoS ONE 2017, 12, e0176303. [Google Scholar] [CrossRef] [Green Version]
- Koyro, H.-W.; Khan, M.; Lieth, H. Halophytic Crops: A Resource for the Future to Reduce the Water Crisis? Emir. J. Food Agric. 2011, 23, 1–16. [Google Scholar] [CrossRef] [Green Version]
- Allen, J.A.; Krauss, K.W.; Hauff, R.D. Factors Limiting the Intertidal Distribution of the Mangrove Species Xylocarpus granatum. Oecologia 2003, 135, 110–121. [Google Scholar] [CrossRef] [PubMed]
- Shabala, S.; Mackay, A. Ion Transport in Halophytes. In Advances in Botanical Research; Elsevier: New York, NY, USA, 2011; Volume 57, pp. 151–199. ISBN 9780123876928. [Google Scholar]
- Chaudhary, D.R. Ion Accumulation Pattern of Halophytes. In Halophytes and Climate Change: Adaptive Mechanisms and Potential Uses; Hasanuzzaman, M., Shabala, S., Fujita, M., Eds.; CABI: Wallingford, UK, 2019; pp. 137–151. ISBN 9781786394330. [Google Scholar]
- Koyro, H.-W.; Geissler, N.; Hussin, S.; Debez, A.; Huchzermeyer, B. Strategies of Halophytes to Survive in a Salty Environment. In Abiotic Stress and Plant Responses; Khan, N.A., Singh, S., Eds.; IK International: New Delhi, India, 2008; pp. 83–104. [Google Scholar]
- Flowers, T.J. Physiology of Halophytes. Plant Soil 1985, 89, 41–56. [Google Scholar] [CrossRef]
- Flowers, T.J.; Galal, H.K.; Bromham, L. Evolution of Halophytes: Multiple Origins of Salt Tolerance in Land Plants. Funct. Plant Biol. 2010, 37, 604. [Google Scholar] [CrossRef]
- Niu, X.; Narasimhan, M.L.; Salzman, R.A.; Bressan, R.A.; Hasegawa, P.M. NaCl Regulation of Plasma Membrane H+-ATPase Gene Expression in a Glycophyte and a Halophyte. Plant Physiol. 1993, 103, 713–718. [Google Scholar] [CrossRef]
- Parida, A.K.; Das, A.B. Salt Tolerance and Salinity Effects on Plants: A Review. Ecotoxicol. Environ. Saf. 2005, 60, 324–349. [Google Scholar] [CrossRef] [PubMed]
- Boestfleisch, C.; Wagenseil, N.B.; Buhmann, A.K.; Seal, C.E.; Wade, E.M.; Muscolo, A.; Papenbrock, J. Manipulating the Antioxidant Capacity of Halophytes to Increase Their Cultural and Economic Value through Saline Cultivation. AoB Plants 2014, 6. [Google Scholar] [CrossRef]
- Akinshina, N.; Oderich, K.T.; Zizov, A.A.; Aito, L.S.; Smail, S.I. Halophyte Biomass—a Promising Source of Renewable Energy. J. Arid Land Stud. 2014, 24, 231–235. [Google Scholar]
- Galvani, A. The Challenge of the Food Sufficiency through Salt Tolerant Crops. Rev. Environ. Sci. Bio/Technol. 2007, 6, 3–16. [Google Scholar] [CrossRef]
- Ksouri, R.; Megdiche, W.; Debez, A.; Falleh, H.; Grignon, C.; Abdelly, C. Salinity Effects on Polyphenol Content and Antioxidant Activities in Leaves of the Halophyte Cakile maritima. Plant Physiol. Biochem. 2007, 45, 244–249. [Google Scholar] [CrossRef] [PubMed]
- Bernstein, N.; Shoresh, M.; Xu, Y.; Huang, B. Involvement of the Plant Antioxidative Response in the Differential Growth Sensitivity to Salinity of Leaves vs Roots during Cell Development. Free Radic. Biol. Med. 2010, 49, 1161–1171. [Google Scholar] [CrossRef]
- Atkinson, N.J.; Urwin, P.E. The Interaction of Plant Biotic and Abiotic Stresses: From Genes to the Field. J. Exp. Bot. 2012, 63, 3523–3543. [Google Scholar] [CrossRef] [Green Version]
- Zarrouk, M.; El Almi, H.; Youssef, N.B.; Sleimi, N.; Smaoui, A.; Miled, D.B.; Abdelly, C. Lipid Composition of Seeds of Local Halophytes: Cakile maritima, Zygophyllum album and Crithmum maritimum. In Proceedings of the Cash Crop Halophytes: Recent Studies; Lieth, H., Mochtchenko, M., Eds.; Springer: Dordrecht, The Netherlands, 2003; Volume 38, pp. 121–124. [Google Scholar]
- Weber, D.J.; Gul, B.; Khan, M.; Williams, T.; Wayman, P.; Warner, S. Composition of Vegetable Oil from Seeds of Native Halophytic Shrubs. In Proceedings of Shrubland Ecosystem Genetics and Biodiversity; Rocky Mountain Research Center: Logan, UT, USA, 2001. [Google Scholar]
- Glenn, E.P.; O’Leary, J.W.; Watson, M.C.; Thompson, T.L.; Kuehl, R.O. Salicornia bigelovii Torr.: An Oilseed Halophyte for Seawater Irrigation. Science 1991, 251, 1065–1067. [Google Scholar] [CrossRef] [PubMed]
- Yajun, B.; Xiaojing, L.; Weiqiang, L. Primary analysis of four salt tolerant plants growing in Hai-He Plain, China. In Cash Crop Halophytes: Recent Studies; Lieth, H., Mochtchenko, M., Eds.; Tasks for Vegetation Science; Springer: Dordrecht, The Netherlands, 2003; Volume 38, pp. 135–138. ISBN 9789048162567. [Google Scholar]
- Debez, A.; Huchzermeyer, B.; Abdelly, C.; Koyro, H.-W. Current Challenges and Future Opportunities for a Sustainable Utilization of Halophytes. In Sabkha Ecosystems; Öztürk, M., Böer, B., Barth, H.-J., Clüsener-Godt, M., Khan, M.A., Breckle, S.-W., Eds.; Tasks for Vegetation Science; Springer: Dordrecht, The Netherlands, 2010; Volume 46, pp. 59–77. ISBN 9789048196722. [Google Scholar]
- Boughalleb, F.; Denden, M. Physiological and Biochemical Changes of Two Halophytes, Nitraria retusa (Forssk.) and Atriplex halimus (L.) Under Increasing Salinity. Agric. J. 2011, 6, 327–339. [Google Scholar] [CrossRef]
- Balasundram, N.; Sundram, K.; Samman, S. Phenolic Compounds in Plants and Agri-Industrial by-Products: Antioxidant Activity, Occurrence, and Potential Uses. Food Chem. 2006, 99, 191–203. [Google Scholar] [CrossRef]
- Meot-Duros, L.; Le Floch, G.; Magné, C. Radical Scavenging, Antioxidant and Antimicrobial Activities of Halophytic Species. J. Ethnopharmacol. 2008, 116, 258–262. [Google Scholar] [CrossRef] [Green Version]
- Bouftira, I.; Hajari, S.A.; Abdelly, C.; Sfar, S. Antioxidative And Free Radical Of Limonium axillare From Qatarian Coasts. Open Access Biomed. Publ. Post Publ. Peer Rev. 2010, 1, WMC00570. [Google Scholar] [CrossRef]
- Medini, F. Effects of Physiological Stage and Solvent on Polyphenol Composition, Antioxidant and Antimicrobial Activities of Limonium Densiflorum. J. Med. Plants Res. 2011, 5. [Google Scholar] [CrossRef]
- Jallali, I.; Megdiche, W.; M’Hamdi, B.; Oueslati, S.; Smaoui, A.; Abdelly, C.; Ksouri, R. Changes in Phenolic Composition and Antioxidant Activities of the Edible Halophyte Crithmum maritimum L. with Physiological Stage and Extraction Method. Acta Physiol. Plant. 2012, 34, 1451–1459. [Google Scholar] [CrossRef]
- HLPE Biofuels and food security. A Report by the High Level Panel of Experts on Food Security and Nutrition of the Committee on World Food Security; HLPE Biofuels and Food Security: Rome, Italy, 2013. [Google Scholar]
- Van Foreest, F. Perspectives for Biogas in Europe; Oxford Institute for Energy Studies: Oxford, UK, 2012. [Google Scholar]
- Kamel, M.; Hammad, S.; Khalaphallah, R.; Elazeem, M.A. Halophytes and Salt Tolerant Wild Plants as a Feedstock for Biogas Production. J. BioSci. Biotechnol. 2019, 8, 151–159. [Google Scholar]
- Bruinsma, J. The Resource Outlook to 2050: By How Much Do Land, Water Use and Crop Yields Need to Increase by 2050? Food and Agriculture Organization of the United Nations, Economic and Social Development Department: Rome, Italy, 2009. [Google Scholar]
- Akinshina, N. Anaerobic Degradation of Halophyte Biomass for Biogas Production. J. Arid Land Stud. J. Arid Land 2012, 22, 227–230. [Google Scholar]
- Turcios, A.E.; Papenbrock, J. Sustainable Treatment of Aquaculture Effluents—What Can We Learn from the Past for the Future? Sustainability 2014, 6, 836–856. [Google Scholar] [CrossRef] [Green Version]
- Turcios, A.E.; Weichgrebe, D.; Papenbrock, J. Effect of Salt and Sodium Concentration on the Anaerobic Methanisation of the Halophyte Tripolium pannonicum. Biomass Bioenerg. 2016, 87, 69–77. [Google Scholar] [CrossRef]
- Turcios, A.E.; Weichgrebe, D.; Papenbrock, J. Potential Use of the Facultative Halophyte Chenopodium quinoa Willd. as Substrate for Biogas Production Cultivated with Different Concentrations of Sodium Chloride under Hydroponic Conditions. Bioresour. Technol. 2016, 203, 272–279. [Google Scholar] [CrossRef] [PubMed]
- Akinshina, N.; Azizov, A.; Karasyova, T.; Klose, E. On the Issue of Halophytes as Energy Plants in Saline Environment. Biomass Bioenerg. 2016, 91, 306–311. [Google Scholar] [CrossRef]
- Cybulska, I.; Chaturvedi, T.; Alassali, A.; Brudecki, G.P.; Brown, J.J.; Sgouridis, S.; Thomsen, M.H. Characterization of the Chemical Composition of the Halophyte Salicornia bigelovii under Cultivation. Energy Fuels 2014, 28, 3873–3883. [Google Scholar] [CrossRef]
- Cybulska, I.; Chaturvedi, T.; Brudecki, G.P.; Kádár, Z.; Meyer, A.S.; Baldwin, R.M.; Thomsen, M.H. Chemical Characterization and Hydrothermal Pretreatment of Salicornia bigelovii Straw for Enhanced Enzymatic Hydrolysis and Bioethanol Potential. Bioresour. Technol. 2014, 153, 165–172. [Google Scholar] [CrossRef] [PubMed]
- Brown, J.J.; Cybulska, I.; Chaturvedi, T.; Thomsen, M.H. Halophytes for the Production of Liquid Biofuels. In Sabkha Ecosystems: Volume IV: Cash Crop Halophyte and Biodiversity Conservation; Springer: Dordrecht, The Netherlands, 2014; pp. 67–72. [Google Scholar] [CrossRef]
- Yang, S.; Li, J.; Zheng, Z.; Meng, Z. Characterization of Spartina alterniflora as Feedstock for Anaerobic Digestion. Biomass Bioenerg. 2009, 33, 597–602. [Google Scholar] [CrossRef]
- Deublein, D.; Angelika, S. (Eds.) Biogas from Waste and Renewable Resources—An Introduction; 3. Nachdr.; Wiley-VCH-Verl: Weinheim, Germany, 2010; ISBN 9783527318414. [Google Scholar]
- Al Seadi, T.; Rutz, D.; Prassl, H.; Köttner, M.; Finsterwalder, T.; Volk, S.; Janssen, R. Biogas Handbook; University of Southern Denmark Esbjerg: Esbjerg, Denmark, 2008; ISBN 9788799296200. [Google Scholar]
- Chen, G.; Zheng, Z.; Yang, S.; Fang, C.; Zou, X.; Zhang, J. Improving Conversion of Spartina alterniflora into Biogas by Co-Digestion with Cow Feces. Fuel Process. Technol. 2010, 91, 1416–1421. [Google Scholar] [CrossRef]
- Li, R.L.; Qiu, G.Y. Spartina alterniflora: Potential Bio-Energy Source for Biofuel Production in China. AMM 2011, 142, 247–251. [Google Scholar] [CrossRef]
- Scarlat, N.; Dallemand, J.-F.; Fahl, F. Biogas: Developments and Perspectives in Europe. Renew. Energy 2018, 129, 457–472. [Google Scholar] [CrossRef]
- Amon, T.; Amon, B.; Kryvoruchko, V.; Zollitsch, W.; Mayer, K.; Gruber, L. Biogas Production from Maize and Dairy Cattle Manure—Influence of Biomass Composition on the Methane Yield. Agric. Ecosyst. Environ. 2007, 118, 173–182. [Google Scholar] [CrossRef]
- LfL Agrarökonomie. Institut für Betriebswirtschaft und Agrarstruktur: Biogasausbeuten-Datenbank. Available online: https://www.lfl.bayern.de/iba/energie/049711/index.php (accessed on 27 February 2021).
- Turcios, A.E.; Weichgrebe, D.; Papenbrock, J. Biogas and Methane Production Using Biomass of the Halophyte Salicornia Spp. In Proceedings of the 14th World Congress on Anaerobic Digestion, Viña del Mar, Chile, 15 November 2015. [Google Scholar]
- Turcios, A.E.; Weichgrebe, D.; Papenbrock, J. Uptake and Biodegradation of the Antimicrobial Sulfadimidine by the Species Tripolium pannonicum Acting as Biofilter and Its Further Biodegradation by Anaerobic Digestion and Concomitant Biogas Production. Bioresour. Technol. 2016, 219, 687–693. [Google Scholar] [CrossRef]
- Chen, Y.; Cheng, J.J.; Creamer, K.S. Inhibition of Anaerobic Digestion Process: A Review. Bioresour. Technol. 2008, 99, 4044–4064. [Google Scholar] [CrossRef]
- Kugelman, I.J.; McCarty, P.L. Cation Toxicity and Stimulation in Anaerobic Waste Treatment. J. Water Pollut. Control Fed. 1965, 37, 97–116. [Google Scholar]
- McCarty, P.L. Anaerobic Waste Treatment Fundamentals—Part Four—Process Design. Public Works 1964, 95, 95–99. [Google Scholar]
- McCarty, P.L.; McKinney, R.E. Volatile Acid Toxicity in Anaerobic Digestion. J. Water Pollut. Control Fed. 1961, 33, 223–232. [Google Scholar]
- Kugelman, I.; CHIN, K.K. Toxicity, Synergism, and Antagonism in Anaerobic Waste Treatment Processes. In Anaerobic Biological Treatment Processes; Advances in Chemistry; American Chemical Society: Washington, DC, USA, 1971; Volume 105, pp. 55–90. ISBN 9780841201316. [Google Scholar]
- Zhang, Y.; Li, L.; Kong, X.; Zhen, F.; Wang, Z.; Sun, Y.; Dong, P.; Lv, P. Inhibition Effect of Sodium Concentrations on the Anaerobic Digestion Performance of Sargassum Species. Energy Fuels 2017, 31, 7101–7109. [Google Scholar] [CrossRef]
- Yerkes, D.W.; Boonyakitsombut, S.; Speece, R.E. Antagonism of Sodium Toxicity by the Compatible Solute Betaine in Anaerobic Methanogenic Systems. Water Sci. Technol. 1997, 36, 15–24. [Google Scholar] [CrossRef]
- Ward, A.J.; Lewis, D.M.; Green, F.B. Anaerobic Digestion of Algae Biomass: A Review. Algal Res. 2014, 5, 204–214. [Google Scholar] [CrossRef]
- Wang, S.; Hou, X.; Su, H. Exploration of the Relationship between Biogas Production and Microbial Community under High Salinity Conditions. Sci. Rep. 2017, 7, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oh, G.; Zhang, L.; Jahng, D. Osmoprotectants Enhance Methane Production from the Anaerobic Digestion of Food Wastes Containing a High Content of Salt. J. Chem. Technol. Biotechnol. 2008, 83, 1204–1210. [Google Scholar] [CrossRef]
- Kapdan, I.K.; Erten, B. Anaerobic Treatment of Saline Wastewater by Halanaerobium lacusrosei. Process Biochem. 2007, 42, 449–453. [Google Scholar] [CrossRef]
- Kimata-Kino, N.; Ikeda, S.; Kurosawa, N.; Toda, T. Saline Adaptation of Granules in Mesophilic UASB Reactors. Int. Biodeterior. Biodegrad. 2011, 65, 65–72. [Google Scholar] [CrossRef]
- Zhang, Y.; Alam, M.A.; Kong, X.; Wang, Z.; Li, L.; Sun, Y.; Yuan, Z. Effect of Salinity on the Microbial Community and Performance on Anaerobic Digestion of Marine Macroalgae: Effect of Salinity on Anaerobic Digestion of Marine Macroalgae. J. Chem. Technol. Biotechnol. 2017, 92, 2392–2399. [Google Scholar] [CrossRef]
- Mottet, A.; Habouzit, F.; Steyer, J.P. Anaerobic Digestion of Marine Microalgae in Different Salinity Levels. Bioresour. Technol. 2014, 158, 300–306. [Google Scholar] [CrossRef] [PubMed]
- Ward, A.; Ball, A.; Lewis, D. Halophytic Microalgae as a Feedstock for Anaerobic Digestion. Algal Res. 2015, 7, 16–23. [Google Scholar] [CrossRef]
- Omil, F.; Méndez, R.; Lema, J.M. Anaerobic Treatment of Saline Wastewaters under High Sulphide and Ammonia Content. Bioresour. Technol. 1995, 54, 269–278. [Google Scholar] [CrossRef]
- Feijoo, G.; Soto, M.; Méndez, R.; Lema, J.M. Sodium Inhibition in the Anaerobic Digestion Process: Antagonism and Adaptation Phenomena. Enzym. Microb. Technol. 1995, 17, 180–188. [Google Scholar] [CrossRef]
- Lefebvre, O.; Quentin, S.; Torrijos, M.; Godon, J.J.; Delgenès, J.P.; Moletta, R. Impact of Increasing NaCl Concentrations on the Performance and Community Composition of Two Anaerobic Reactors. Appl. Microbiol. Biotechnol. 2007, 75, 61–69. [Google Scholar] [CrossRef]
- Buxy, S.; Diltz, R.; Pullammanappallil, P. Biogasification of Marine Algae Nannochloropsis oculata. In Ceramic Transactions Series; Wicks, G., Simon, J., Zidan, R., Brigmon, R., Fischman, G., Arepalli, S., Norris, A., McCluer, M., Eds.; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2013; pp. 59–67. ISBN 9781118585160. [Google Scholar]
- Hartmann, H.; Angelidaki, I.; Ahring, B.K. Co-Digestion of the Organic Fraction of Municipal Waste With Other Waste Types. In Biomethanization of the Organic Fraction of Municipal Solid Wastes; Mata-Alvarez, J., Ed.; IWA Publishing: London, UK, 2002; pp. 181–200. [Google Scholar]
- Alhraishawi, A.A.; Alani, W.K. The Co-Fermentation of Organic Substrates: A Review Performance of Biogas Production under Different Salt Content. J. Phys.: Conf. Ser. 2018, 1032. [Google Scholar] [CrossRef] [Green Version]
- Schnürer, A.; Jarvis, Å. Microbiological Handbook for Biogas Plants; Avfall Sverige: Malmö, Sweden, 2010; ISSN 1103-4092. [Google Scholar]
- Mshandete, A.; Kivaisi, A.; Rubindamayugi, M.; Mattiasson, B. Anaerobic Batch Co-Digestion of Sisal Pulp and Fish Wastes. Bioresour. Technol. 2004, 95, 19–24. [Google Scholar] [CrossRef] [PubMed]
- Debez, A.; Belghith, I.; Friesen, J.; Montzka, C.; Elleuche, S. Facing the Challenge of Sustainable Bioenergy Production: Could Halophytes Be Part of the Solution? J. Biol. Eng. 2017, 11, 27. [Google Scholar] [CrossRef] [PubMed]
- Yang, S.; Li, J.; Zheng, Z.; Meng, Z. Lignocellulosic Structural Changes of Spartina alterniflora after Anaerobic Mono- and Co-Digestion. Int. Biodeterior. Biodegrad. 2009, 63, 569–575. [Google Scholar] [CrossRef]
- Zhang, J.; Li, W.; Lee, J.; Loh, K.-C.; Dai, Y.; Tong, Y.W. Enhancement of Biogas Production in Anaerobic Co-Digestion of Food Waste and Waste Activated Sludge by Biological Co-Pretreatment. Energy 2017, 137, 479–486. [Google Scholar] [CrossRef]
- Nasir, Z.; Ahring, B.K.; Uellendahl, H. Enhancing the Hydrolysis Process in a Dry Anaerobic Digestion Process for the Organic Fraction of Municipal Solid Waste. Bioresour. Technol. Rep. 2020, 11. [Google Scholar] [CrossRef]
- Rocamora, I.; Wagland, S.T.; Villa, R.; Simpson, E.W.; Fernández, O.; Bajón-Fernández, Y. Dry Anaerobic Digestion of Organic Waste: A Review of Operational Parameters and Their Impact on Process Performance. Bioresour. Technol. 2020, 299. [Google Scholar] [CrossRef] [PubMed]
- Sahu, P. A Comprehensive Review of Saline Effluent Disposal and Treatment: Conventional Practices, Emerging Technologies, and Future Potential. J. Water Reuse Desal. 2021, 11, 33–65. [Google Scholar] [CrossRef]
- Uellendahl, H.; Ahring, B.K. Anaerobic Digestion as Final Step of a Cellulosic Ethanol Biorefinery: Biogas Production from Fermentation Effluent in a UASB Reactor-Pilot-Scale Results. Biotechnol. Bioeng. 2010, 107, 59–64. [Google Scholar] [CrossRef] [PubMed]
- Santamaría-Fernández, M.; Molinuevo-Salces, B.; Lübeck, M.; Uellendahl, H. Biogas Potential of Green Biomass after Protein Extraction in an Organic Biorefinery Concept for Feed, Fuel and Fertilizer Production. Renew. Energy 2018, 129, 769–775. [Google Scholar] [CrossRef]
- Hagman, L. How Do Biogas Solutions Influence the Sustainability of Bio-Based Industrial Systems? Linköping Studies in Science and Technology; Licentiate Thesis; Linköping University Electronic Press: Linköping, Sweden, 2018; Volume 1822, ISBN 9789176851982. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Family | Origin | Uses | Reported Salt Tolerance to NaCl (mM) | Reference |
|---|---|---|---|---|---|
| Acrostichum aureum | Polypodiaceae | Pantropical | Ornamental, medicinal | 170 | Kapler [6]; Sun et al. [10] |
| Althaea officinalis | Malvaceae | Mediterranean, invasive in other regions | Medicinal, phytoremediator, ornamental | 200 | Kapler [6]; Rezaie et al. [11] |
| Ammophila arenaria | Poaceae | Europe and western Asia, uncertain in many places, invasive | Fodder, biomass, groundcover | ~250 | Kapler [6]; CABI [12] |
| Atriplex hortensis | Amaranthaceae | Cosmopolitan, uncertain in many places | Food, fodder, spice, biofuel | >250 | Wilson et al. [13]; Hasanuzzaman et al. [14]; Kapler [6] |
| Atriplex lentiformis | Amaranthaceae | Southwestern USA and northern Mexico | Food | 500 | O’Leary et al. [15]; Hasanuzzaman et al. [14] |
| Batis maritima | Bataceae | America | Eaten raw, cooked, or pickled | 500 | O’Leary et al. [15], Hasanuzzaman et al. [14] |
| Bruguiera gymnorrhiza | Rhizophoraceae | Australasia, Oceania, East Africa, Madagascar | Fuel, charcoal, construction, medicinal, food, fodder | 500 | Kapler [6]; Zhu et al. [16] |
| Carpobrotus spp. | Aizoaceae | South Africa, south Australian coast and coastal Chile | Groundcover, medicinal | >300 | Varone et al. [17] |
| Chenopodium quinoa | Amaranthaceae | Andean highlands | Cooking, baking, animal feed, green fodder and pellet, modified food products such as cereals, pasta and cookies, industrial use of starch, protein and saponin | 400 | Adolf et al. [18] |
| Cochlearia danica | Brassicaceae | Atlantic Europe, now invasive along motorways in Eurasia | Food, spice | 200 | Kapler [6]; Nawaz et al. [19] |
| Cochlearia officinalis | Brassicaceae | Europe | Fresh salads | 100 | de Vos [20], Hasanuzzaman et al. [14] |
| Crambe maritima | Brassicaceae | Europe from the north Atlantic through the Mediterranean to Asia and around the Black Sea | Fresh salads | >100 | de Vos et al. [21], Hasanuzzaman et al. [14] |
| Crithmum maritimum | Apiaceae | Atlantic Europe, Mediterranean regions | Ornamental, food, edible oil, fodder, biofuel Fresh and pickled as spice and for salads | 150 | Kapler [6]; Ben-Hamed et al. [22], Hamed et al. [22]; Ben Amor et al. [23]; Hasanuzzaman et al. [14] |
| Derris heterophylla | Fabaceae | Pantropical | Medicinal, natural insecticide and fish poison for fishing and biomanipulation | 220 | Kapler [6] |
| Diplotaxis tenuifolia | Brassicaceae | Europe, western Asia and northern Africa | Mixed salads, medicinal | ~150 | de Vos [20]; Hasanuzzaman et al. [14] |
| Eugeisonia insignis | Arecaceae | Sarawac’ endemic | Food, fodder | Kapler [6] | |
| Glaux maritima | Primulaceae | Native to Old and New World | Medicinal | 300 | Kapler [6]; Rozema [24] |
| Inula crithmoides | Asteraceae | Europe, Africa and Asia | Salads, pickled in vinegar | >400 | Tardío et al. [25]; Zurayk and Baalbaki [26]; Hasanuzzaman et al. [14] |
| Ipomoea pes-caprae | Convolvulaceae | Pantropical | Medicinal, fodder, green manure | 300 | Kapler [6]; Zhang et al. [27] |
| Limbarda crithmoides | Asteraceae | Mediterranean, Black Sea, Atlantic Europe | Food, fodder | 450 | Kapler [6]; Al-Hassan et al. [28] |
| Mesembryanthemum crystallinum | Aizoaceae | North Africa, south-western Africa, western Asia and southern Europe, invasive in the New World and Australia | Ornamental, food, fodder | 400 | Kapler [6]; Hasanuzzaman et al. [14]; Agarie et al. [29]; Herppich et al. [30] |
| Nypa fruticans | Arecaceae | Australasia, Oceania | Construction and fiber material, source of edible salt and sugar | >200 | Kapler [6]; Theerawitaya et al. [31] |
| Oncosperma tigillarium | Arecaceae | Australasia | Prized wood, food, construction (traditional spears) | >200 | Kapler [6] |
| Pandanus tectorius, P. odoratissimus | Pandanaceae | Australasia, Oceania | Food, fodder, fiber, medicinal, perfume | >200 | Kapler [6] |
| Pennisetum clandestinum (syn. Sporobolus virginicus) | Poaceae | Pantropical | Fodder, biomass, groundcover | >200 | Kapler [6]; Muscolo et al. [32] |
| Phoenix paludosa | Arecaceae | Australasia, Oceania | Food, fodder | >200 | Kapler [6] |
| Plantago coronopus | Plantaginaceae | Europe, northern Africa, central and west Asia | Salad, greens | 250 | Koyro [32]; Hasanuzzaman et al. [14] |
| Portulaca oleracea | Portulacaceae | Native in the Old and Australasia, uncertain in the New World | Food, fodder | <140 mM | Simopoulos [33]; Yazici et al. [34]; Hasanuzzaman et al. [14]; Kapler [6] |
| Rhizophora mucronata | Rhizophoraceae | Australasia, Oceania, East Africa, Madagascar | Construction, especially for aquaculture, food, fodder, fuel, medicine | >500 | Kapler [6]; Kodikara et al. [35] |
| Salicornia europaea agg. and Sarcocornia spp. | Amaranthaceae | S. europaea native to the Old World, many species cosmopolitan | Probiotic, fodder, biofuel, phytoremediation, ornamental | 500 | O’Leary et al. [15]; Ventura et al. [36]; Hasanuzzaman et al. [14]; Kapler [6] |
| Salsola kali | Amaranthaceae | Europe along the shores of Baltic Sea, North Sea and the Atlantic Ocean | Production of sodium carbonate, forage | >200 | Reimann & Breckle [37] |
| Sesuvium portulacastrum | Aizoaceae | Pantropical | Food, fodder | 400 | Kapler [6]; Messeddi et al. [38] |
| Sonneratia alba | Sonneratiaceae | Australasia, Oceania, East Africa | Food, fodder, fuel, roots used as buoys | >500 | Kapler [6] |
| Spinifex spp. | Poaceae | Australasia, Oceania | Groundcover | Kapler [6] | |
| Tetragonia tetragonioides | Aizoaceae | Argentina, Australia, Chile, Japan and New Zealand. | Frozen like spinach | 174 | Wilson et al. [13]; Słupski et al. [39]; Hasanuzzaman et al. [14] |
| Tetragonolobus & Lotus | Fabaceae | Native in the Old World, invasive in the New World | Fodder, phytoremediation | Kapler [6] | |
| Triglochin maritima | Juncaginaceae | Cosmopolitan, uncertain in many places | Food, fodder, biofuel | >250 | Kapler [6]; Boestfleisch & Papenbrock [40] |
| Tripolium pannonicum (syn. Aster tripolium) | Asteraceae | Eurasia and North Africa, mostly Pannon biogeographic region and Atlantic shores | Ornamental, food, fodder | 300 | Koyro et al. [41]; Hasanuzzaman et al. [14]; Kapler [6] |
| Xylocarpus granatum | Meliaceae | Pantropical | Medicinal, seed oils used to make candle, soap, amadou | ~400 | Allen et al. [42]; Kapler [6] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Turcios, A.E.; Cayenne, A.; Uellendahl, H.; Papenbrock, J. Halophyte Plants and Their Residues as Feedstock for Biogas Production—Chances and Challenges. Appl. Sci. 2021, 11, 2746. https://doi.org/10.3390/app11062746
Turcios AE, Cayenne A, Uellendahl H, Papenbrock J. Halophyte Plants and Their Residues as Feedstock for Biogas Production—Chances and Challenges. Applied Sciences. 2021; 11(6):2746. https://doi.org/10.3390/app11062746
Chicago/Turabian StyleTurcios, Ariel E., Aadila Cayenne, Hinrich Uellendahl, and Jutta Papenbrock. 2021. "Halophyte Plants and Their Residues as Feedstock for Biogas Production—Chances and Challenges" Applied Sciences 11, no. 6: 2746. https://doi.org/10.3390/app11062746
APA StyleTurcios, A. E., Cayenne, A., Uellendahl, H., & Papenbrock, J. (2021). Halophyte Plants and Their Residues as Feedstock for Biogas Production—Chances and Challenges. Applied Sciences, 11(6), 2746. https://doi.org/10.3390/app11062746