
Ethanol Extract of Pinus koraiensis Leaves Mitigates High Fructose-Induced Hepatic Triglyceride Accumulation and Hypertriglyceridemia

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. EPK
2.2. Ethics Statement
2.3. Animal Models
2.4. Biochemical Analysis
2.5. RNA Isolation and Real-Time RT-PCR
2.6. Hepatic Histology
2.7. Western Blotting
2.8. Oil Red O Staining
2.9. Cell Culture
2.10. Statistical Analysis
3. Results
3.1. The HFrD Administered for Six Weeks Did Not Affect Body Weight
3.2. The HFrD Did Not Affect TC, HDL, and LDL Levels
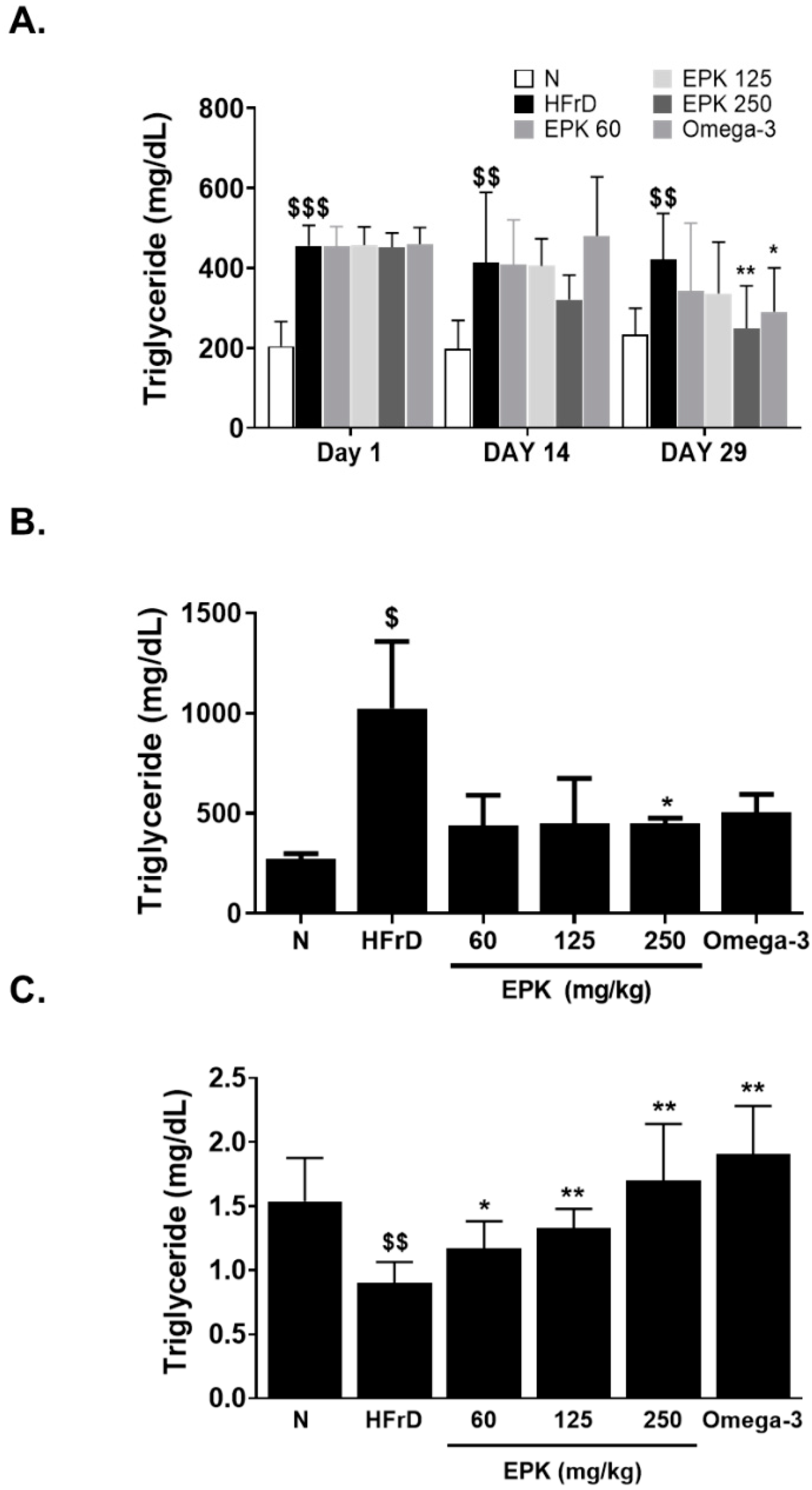
3.3. EPK Decreased Serum and Hepatic TG Levels and Increased TG Excretion through Feces in High Fructose–Induced Hypertriglyceridemia Animal Models
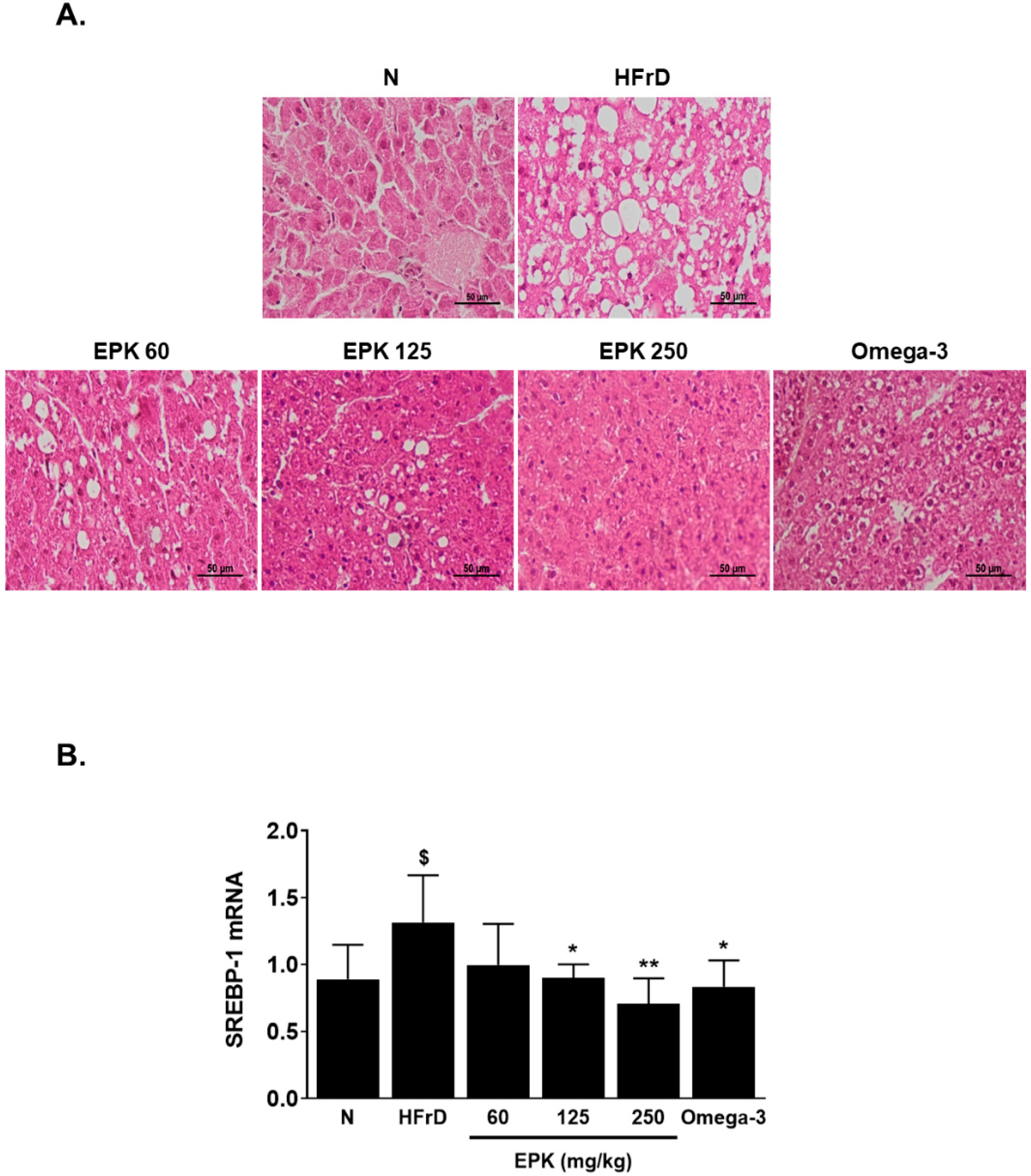
3.4. EPK Mitigated Fructose-Induced Fatty Liver
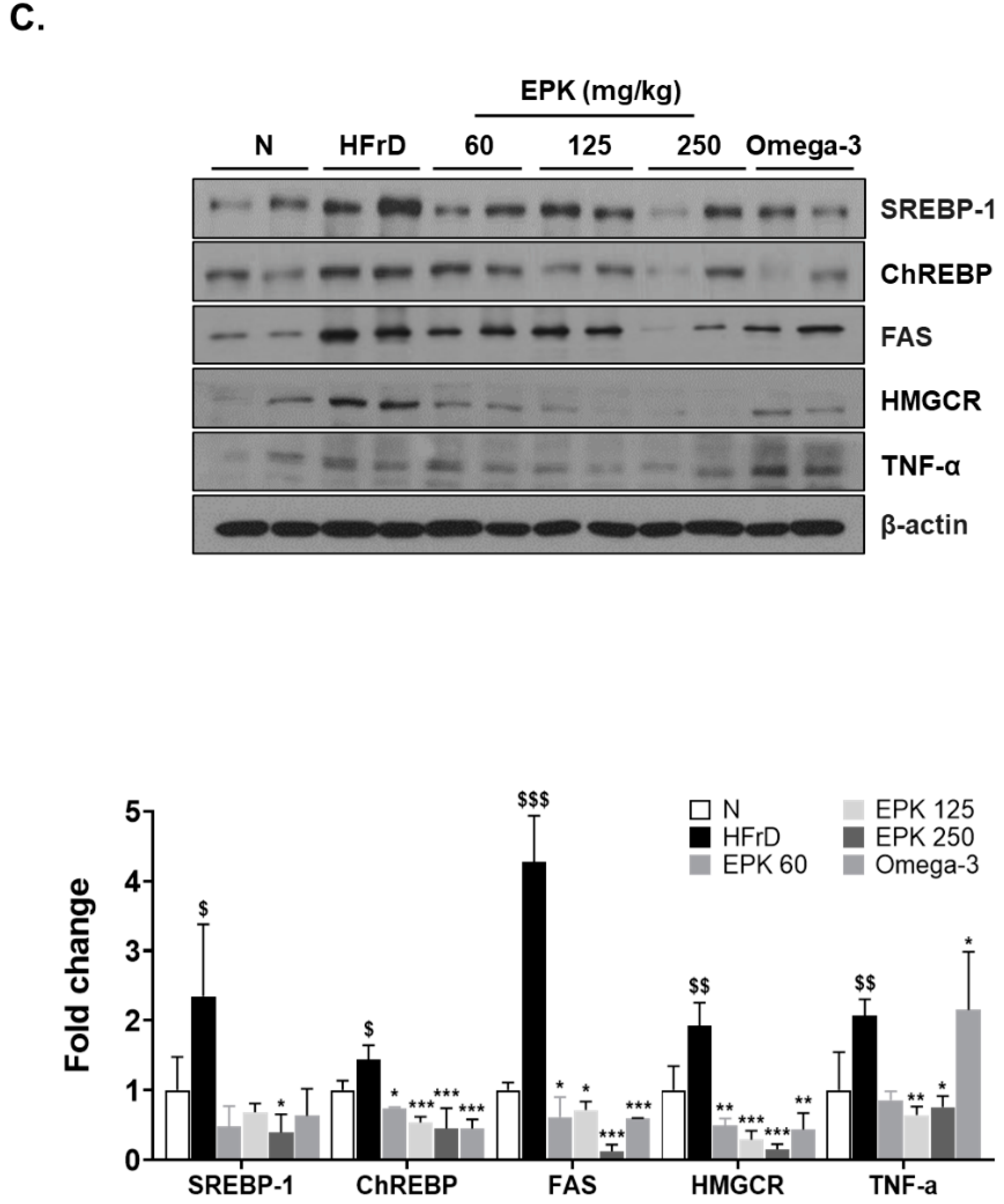
3.5. EPK Inhibited DNL-Related Proteins in HFrD-Induced Fatty Liver
3.6. EPK Inhibited TNF-α, a Pro-Inflammatory Marker, in HFrD-Induced Fatty Liver
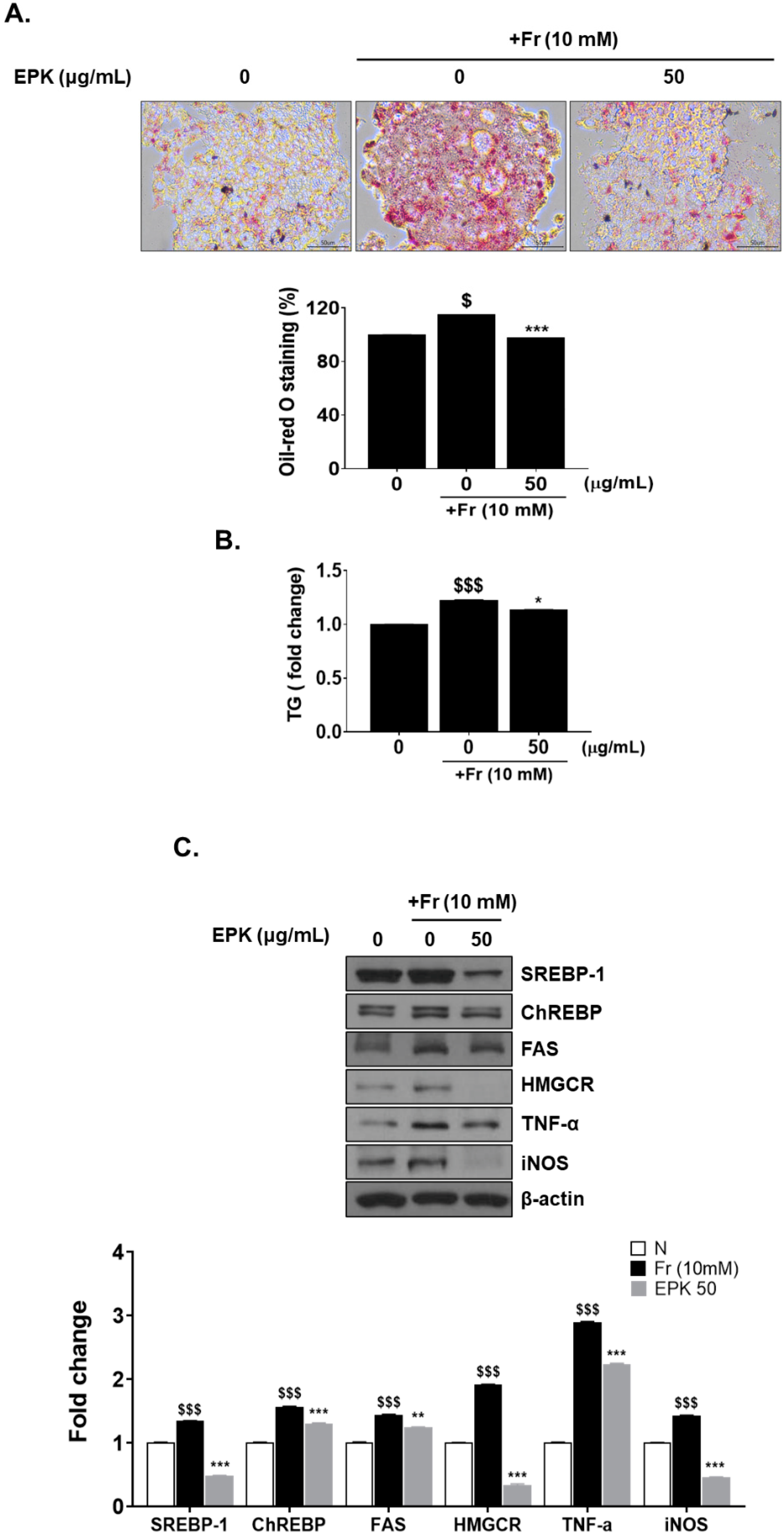
3.7. EPK Inhibited Fr-Induced Cellular Lipid Accumulation in HepG2 Cells
3.8. EPK Decreased Fr-Induced Intracellular TG Level
3.9. EPK Inhibited Fr-Induced Core Factors of DNL, such as SREBP-1 and ChREBP, and Their Target Enzymes
3.10. EPK Reduced Fr-Induced TNF-α and iNOS (Inflammatory Factors)
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Khosrow, A.; Lisa, F.; Heather, B. Fructose, insulin resistance, and metabolic dyslipidemia. Nutr. Metab. 2005, 2, 5. [Google Scholar]
- Hallfrisch, J. Metabolic effects of dietary fructose. FASEB J. 1990, 4, 2652–2660. [Google Scholar] [CrossRef] [PubMed]
- Mayes, P.A. Intermediary metabolism of fructose. Am. J. Clin. Nutr. 1993, 58, 754S–765S. [Google Scholar] [CrossRef] [PubMed]
- Tetri, L.H.; Basaranoglu, M.; Brunt, E.M.; Yerian, L.M.; Neuschwander-Tetri, B.A. Severe NAFLD with hepatic necroinflammatory changes in mice fed trans fats and a high-fructose corn syrup equivalent. Am. J. Physiol.-Gastrointest. Liver Physiol. 2008, 295, G987–G995. [Google Scholar] [CrossRef]
- Sobrecases, H.; Lê, K.-A.; Bortolotti, M.; Schneiter, P.; Ith, M.; Kreis, R.; Boesch, C.; Tappy, L. Effects of short-term overfeeding with fructose, fat and fructose plus fat on plasma and hepatic lipids in healthy men. Diabetes Metab. 2010, 36, 244–246. [Google Scholar] [CrossRef]
- Softic, S.; Cohen, D.E.; Kahn, C.R. Role of dietary fructose and hepatic de novo lipogenesis in fatty liver disease. Dig. Dis. Sci. 2016, 61, 1282–1293. [Google Scholar] [CrossRef] [Green Version]
- Jegatheesan, P.; De Bandt, J.P. Fructose and NAFLD: The multifaceted aspects of fructose metabolism. Nutrients 2017, 9, 230. [Google Scholar] [CrossRef] [Green Version]
- Zavaroni, I.; Chen, Y.-D.I.; Reaven, G.M. Studies of the mechanism of fructose-induced hypertriglyceridemia in the rat. Metabolism 1982, 31, 1077–1083. [Google Scholar] [CrossRef]
- Bhatt, D.; Tannock, L. Risk of fasting and non-fasting hypertriglyceridemia in coronary vascular disease and pancreatitis. In Endotext; MDText.com, Inc.: South Dartmouth, MA, USA, 2018. [Google Scholar]
- Siri-Tarino, P.W.; Krauss, R.M. The early years of lipoprotein research: From discovery to clinical application. J. Lipid Res. 2016, 57, 1771–1777. [Google Scholar] [CrossRef] [Green Version]
- Herman, M.A.; Samuel, V.T. The sweet path to metabolic demise: Fructose and lipid synthesis. Trends Endocrinol. Metab. 2016, 27, 719–730. [Google Scholar] [CrossRef] [Green Version]
- Jegatheesan, P.; Beutheu, S.; Ventura, G.; Nubret, E.; Sarfati, G.; Bergheim, I.; De Bandt, J.-P. Citrulline and nonessential amino acids prevent fructose-induced nonalcoholic fatty liver disease in rats. J. Nutr. 2015, 145, 2273–2279. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, X.; Li, Y.; Yang, W.; Xiao, C.; Fu, S.; Deng, Q.; Ding, H.; Wang, Z.; Liu, G.; Li, X. SREBP-1c overexpression induces triglycerides accumulation through increasing lipid synthesis and decreasing lipid oxidation and VLDL assembly in bovine hepatocytes. J. Steroid Biochem. Mol. Biol. 2014, 143, 174–182. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.-S.; Krawczyk, S.A.; Doridot, L.; Fowler, A.J.; Wang, J.X.; Trauger, S.A.; Noh, H.-L.; Kang, H.J.; Meissen, J.K.; Blatnik, M. ChREBP regulates fructose-induced glucose production independently of insulin signaling. J. Clin. Investig. 2019, 126, 4372–4386. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Iizuka, K.; Takao, K.; Kato, T.; Horikawa, Y.; Takeda, J. ChREBP reciprocally regulates liver and plasma triacylglycerol levels in different manners. Nutrients 2018, 10, 1699. [Google Scholar] [CrossRef] [Green Version]
- Cho, S.-M.; Lee, E.-O.; Kim, S.-H.; Lee, H.-J. Essential oil of Pinus koraiensis inhibits cell proliferation and migration via inhibition of p21-activated kinase 1 pathway in HCT116 colorectal cancer cells. BMC Complementary Altern. Med. 2014, 14, 275. [Google Scholar] [CrossRef] [Green Version]
- Lee, M.-S.; Cho, S.-M.; Lee, M.-h.; Lee, E.-O.; Kim, S.-H.; Lee, H.-J. Ethanol extract of Pinus koraiensis leaves containing lambertianic acid exerts anti-obesity and hypolipidemic effects by activating adenosine monophosphate-activated protein kinase (AMPK). BMC Complementary Altern. Med. 2016, 16, 51. [Google Scholar] [CrossRef] [Green Version]
- Joo, H.-E.; Lee, H.-J.; Sohn, E.J.; Lee, M.-H.; Ko, H.-S.; Jeong, S.-J.; Lee, H.-J.; Kim, S.-H. Anti-diabetic potential of the essential oil of Pinus koraiensis leaves toward streptozotocin-treated mice and HIT-T15 pancreatic β cells. Biosci. Biotechnol. Biochem. 2013, 77, 1997–2001. [Google Scholar] [CrossRef] [Green Version]
- Folch, J.; Lees, M.; Sloane Stanley, G.H. A simple method for the isolation and purification of total lipids from animal tissues. J. Biol. Chem. 1957, 226, 497–509. [Google Scholar] [CrossRef]
- Lee, S.-O.; Xu, Y.; Han, H.; Jeong, S.-T.; Lee, Y.-K.; Paik, J.K.; Cha, J.-S.; Lee, H.-J. Fermented Rhus verniciflua Stokes extract alleviates nonalcoholic fatty liver through the AMPK/SREBP1/PCSK9 pathway in HFD-induced nonalcoholic fatty liver animal model. Appl. Sci. 2020, 10, 6833. [Google Scholar] [CrossRef]
- Xu, Y.; Cha, J.-S.; Lee, S.-O.; Ryu, S.-I.; Lee, Y.-K.; Han, H.; Kim, J.-E.; Lee, M.-H.; Lee, E.-O.; Lee, H.-J. HMT Exerts an Anticancer Effect by Targeting PAK-1. Appl. Sci. 2021, 11, 6034. [Google Scholar] [CrossRef]
- Lee, M.-S.; Kim, J.-S.; Cho, S.-M.; Lee, S.O.; Kim, S.-H.; Lee, H.-J. Fermented Rhus verniciflua Stokes extract exerts an Antihepatic Lipogenic effect in oleic-acid-induced HepG2 cells via Upregulation of AMP-activated protein kinase. J. Agric. Food Chem. 2015, 63, 7270–7276. [Google Scholar] [CrossRef] [PubMed]
- Ballestri, S.; Nascimbeni, F.; Romagnoli, D.; Baldelli, E.; Lonardo, A. The role of nuclear receptors in the pathophysiology, natural course, and drug treatment of NAFLD in humans. Adv. Ther. 2016, 33, 291–319. [Google Scholar] [CrossRef] [PubMed]
- Ter Horst, K.W.; Serlie, M.J. Fructose consumption, lipogenesis, and non-alcoholic fatty liver disease. Nutrients 2017, 9, 981. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, J.; Fox, C.S.; Jacques, P.F.; Speliotes, E.K.; Hoffmann, U.; Smith, C.E.; Saltzman, E.; McKeown, N.M. Sugar-sweetened beverage, diet soda, and fatty liver disease in the Framingham Heart Study cohorts. J. Hepatol. 2015, 63, 462–469. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Choi, Y.; Abdelmegeed, M.A.; Song, B.-J. Diet high in fructose promotes liver steatosis and hepatocyte apoptosis in C57BL/6J female mice: Role of disturbed lipid homeostasis and increased oxidative stress. Food Chem. Toxicol. 2017, 103, 111–121. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Handayani, D.; Febrianingsih, E.; Kurniawati, A.D.; Kusumastuty, I.; Nurmalitasari, S.; Widyanto, R.M.; Oktaviani, D.N.; Innayah, A.M.; Sulistyowati, E. High-fructose diet initially promotes increased aortic wall thickness, liver steatosis, and cardiac histopathology deterioration, but does not increase body fat index. J. Public Health Res. 2021, 10, 2181. [Google Scholar] [CrossRef] [PubMed]
- Ren, L.-P.; Song, G.-Y.; Hu, Z.-J.; Zhang, M.; Peng, L.; Chen, S.-C.; Wei, L.; Li, F.; Sun, W. The chemical chaperon 4-phenylbutyric acid ameliorates hepatic steatosis through inhibition of de novo lipogenesis in high-fructose-fed rats. Int. J. Mol. Med. 2013, 32, 1029–1036. [Google Scholar] [CrossRef] [Green Version]
- Nakamura, K.; Fukunishi, S.; Yokohama, K.; Ohama, H.; Tsuchimoto, Y.; Asai, A.; Tsuda, Y.; Higuchi, K. A long-lasting dipeptidyl peptidase-4 inhibitor, teneligliptin, as a preventive drug for the development of hepatic steatosis in high-fructose diet-fed ob/ob mice. Int. J. Mol. Med. 2017, 39, 969–983. [Google Scholar] [CrossRef] [Green Version]
- Yu, X.; Ren, L.-P.; Wang, C.; Zhu, Y.-J.; Xing, H.-Y.; Zhao, J.; Song, G.-Y. Role of X-box binding protein-1 in fructose-induced de novo lipogenesis in HepG2 cells. Chin. Med. J. 2018, 131, 2310–2319. [Google Scholar] [CrossRef]
- Vasiljević, A.; Bursać, B.; Djordjevic, A.; Milutinović, D.V.; Nikolić, M.; Matić, G.; Veličković, N. Hepatic inflammation induced by high-fructose diet is associated with altered 11βHSD1 expression in the liver of Wistar rats. Eur. J. Nutr. 2014, 53, 1393–1402. [Google Scholar] [CrossRef]
- Gao, H.; Guan, T.; Li, C.; Zuo, G.; Yamahara, J.; Wang, J.; Li, Y. Treatment with ginger ameliorates fructose-induced Fatty liver and hypertriglyceridemia in rats: Modulation of the hepatic carbohydrate response element-binding protein-mediated pathway. Evid.-Based Complement. Altern. Med. 2012, 2012, 570948. [Google Scholar] [CrossRef] [PubMed]
- Dentin, R.; Benhamed, F.; Hainault, I.; Fauveau, V.; Foufelle, F.; Dyck, J.R.; Girard, J.; Postic, C. Liver-specific inhibition of ChREBP improves hepatic steatosis and insulin resistance in ob/ob mice. Diabetes 2006, 55, 2159–2170. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Iizuka, K.; Miller, B.; Uyeda, K. Deficiency of carbohydrate-activated transcription factor ChREBP prevents obesity and improves plasma glucose control in leptin-deficient (ob/ob) mice. Am. J. Physiol.-Endocrinol. Metab. 2006, 291, E358–E364. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tappy, L.; Lê, K.-A. Metabolic effects of fructose and the worldwide increase in obesity. Physiol. Rev. 2010, 90, 23–46. [Google Scholar] [CrossRef] [Green Version]
- Postic, C.; Girard, J. Contribution of de novo fatty acid synthesis to hepatic steatosis and insulin resistance: Lessons from genetically engineered mice. J. Clin. Investig. 2008, 118, 829–838. [Google Scholar] [CrossRef] [Green Version]
- Postic, C.; Dentin, R.; Denechaud, P.-D.; Girard, J. ChREBP, a transcriptional regulator of glucose and lipid metabolism. Annu. Rev. Nutr. 2007, 27, 179–192. [Google Scholar] [CrossRef]
- Kanuri, G.; Spruss, A.; Wagnerberger, S.; Bischoff, S.C.; Bergheim, I. Role of tumor necrosis factor α (TNFα) in the onset of fructose-induced nonalcoholic fatty liver disease in mice. J. Nutr. Biochem. 2011, 22, 527–534. [Google Scholar] [CrossRef]
- Cigliano, L.; Spagnuolo, M.S.; Crescenzo, R.; Cancelliere, R.; Iannotta, L.; Mazzoli, A.; Liverini, G.; Iossa, S. Short-term fructose feeding induces inflammation and oxidative stress in the hippocampus of young and adult rats. Mol. Neurobiol. 2018, 55, 2869–2883. [Google Scholar] [CrossRef]
- Van der Borght, K.; Köhnke, R.; Göransson, N.; Deierborg, T.; Brundin, P.; Erlanson-Albertsson, C.; Lindqvist, A. Reduced neurogenesis in the rat hippocampus following high fructose consumption. Regul. Pept. 2011, 167, 26–30. [Google Scholar] [CrossRef]
- Povero, D.; Feldstein, A.E. Novel molecular mechanisms in the development of non-alcoholic steatohepatitis. Diabetes Metab. J. 2016, 40, 1–11. [Google Scholar] [CrossRef] [Green Version]
- d’Alessio, P.A.; Ostan, R.; Bisson, J.-F.; Schulzke, J.D.; Ursini, M.V.; Béné, M.C. Oral administration of d-limonene controls inflammation in rat colitis and displays anti-inflammatory properties as diet supplementation in humans. Life Sci. 2013, 92, 1151–1156. [Google Scholar] [CrossRef] [PubMed]
- Vallianou, I.; Peroulis, N.; Pantazis, P.; Hadzopoulou-Cladaras, M. Camphene, a plant-derived monoterpene, reduces plasma cholesterol and triglycerides in hyperlipidemic rats independently of HMG-CoA reductase activity. PLoS ONE 2011, 6, e20516. [Google Scholar] [CrossRef] [PubMed] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | Routes of Administration | Dosage (mg/kg) | No. of Mice |
|---|---|---|---|
| Normal | Oral administration | 0 | 8 |
| HFrD (High fructose diet) | 0 | 8 | |
| EPK 60 | 60 | 8 | |
| EPK 125 | 125 | 8 | |
| EPK 250 | 250 | 8 | |
| Omega-3 | 500 | 8 |
| Group | Time after Administration (Days) | ||
|---|---|---|---|
| Body Weight (g) | % of BW Change | ||
| Day1 | Day22 | ||
| N | 427.08 ± 31.83 | 538.16 ± 47.62 | +25.92 ± 3.65 |
| HFrD | 418.27 ± 9.82 | 530.20 ± 28.36 | +26.76 ± 6.07 |
| EPK 60 | 405.36 ± 17.08 | 503.16 ± 16.79 | +24.74 ± 1.90 |
| EPK 125 | 390.32 ± 16.53 | 492.51 ± 29.85 | +26.21 ± 6.51 |
| EPK 250 | 408.18 ± 14.16 | 518.51 ± 26.61 | +27.01 ± 4.27 |
| Omega-3 | 417.59 ± 9.94 | 510.38 ± 44.97 | +22.34 ± 11.77 |
| Group | Concentration (mg/dL)/Time after Administration (Days) | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| Total Cholesterol | HDL-C | LDL-C | |||||||
| Day1 | Day29 | % of Total Cholesterol Change | Day1 | Day29 | % of HDL-C Change | Day1 | Day29 | % of LDL-C Change | |
| N | 102 ± 14.42 | 100.29 ± 15.84 | −1.13 ± 13.95 | 33.46 ± 3.86 | 29.21 ± 3.97 | −12.24 ± 10.65 | 9.82 ± 1.58 | 9.36 ± 2.96 | −5.80 ± 17.73 |
| HFrD | 82.57 ± 13.21 | 90.86 ± 10.14 | +11.97 ± 18.07 | 29.18 ± 3.64 | 27.9 ± 2.42 | −2.00 ± 13.87 | 5.84 ± 0.80 | 6.98 ± 1.38 | +18.92 ± 10.47 |
| EPK 60 | 89.57 ± 12.81 | 85.14 ± 15.02 | −4.09 ± 16.18 | 32.21 ± 5.89 | 27.2 ± 4.87 | −14.52 ± 13.87 | 5.78 ± 1.86 | 5.7 ± 1.45 | +0.44 ± 16.66 |
| EPK 125 | 85.86 ± 16.43 | 92.71 ± 20.69 | +8.58 ± 16.03 | 30.65 ± 5.06 | 32.87 ± 6.11 | +6.81 ± 16.61 | 5.62 ± 0.49 | 6.06 ± 0.79 | +8.10 ± 13.70 |
| EPK 250 | 86.57 ± 2.88 | 94.43 ± 21.53 | +9.42 ± 26.77 | 33.58 ± 1.82 | 30.99 ± 3.94 | −7.65 ± 15.92 | 5.42 ± 0.98 | 5.9 ± 0.65 | +10.24 ± 12.89 |
| Omega-3 | 90.71 ± 9.23 | 71.29 ± 3.95 | −20.49 ± 11.54 | 33.56 ± 2.15 | 25.65 ± 1.83 | −24.41 ± 7.58 | 5.58 ± 0.67 | 5.8 ± 0.58 | +4.31 ± 6.57 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, M.-H.; Park, S.; Xu, Y.; Kim, J.-E.; Han, H.; Lee, J.-H.; Paik, J.K.; Lee, H.-J. Ethanol Extract of Pinus koraiensis Leaves Mitigates High Fructose-Induced Hepatic Triglyceride Accumulation and Hypertriglyceridemia. Appl. Sci. 2022, 12, 6745. https://doi.org/10.3390/app12136745
Lee M-H, Park S, Xu Y, Kim J-E, Han H, Lee J-H, Paik JK, Lee H-J. Ethanol Extract of Pinus koraiensis Leaves Mitigates High Fructose-Induced Hepatic Triglyceride Accumulation and Hypertriglyceridemia. Applied Sciences. 2022; 12(13):6745. https://doi.org/10.3390/app12136745
Chicago/Turabian StyleLee, Min-Ho, Sunyeong Park, Yinzhu Xu, Jung-Eun Kim, Hengmin Han, Jae-Hyeon Lee, Jean Kyung Paik, and Hyo-Jeong Lee. 2022. "Ethanol Extract of Pinus koraiensis Leaves Mitigates High Fructose-Induced Hepatic Triglyceride Accumulation and Hypertriglyceridemia" Applied Sciences 12, no. 13: 6745. https://doi.org/10.3390/app12136745
APA StyleLee, M. -H., Park, S., Xu, Y., Kim, J. -E., Han, H., Lee, J. -H., Paik, J. K., & Lee, H. -J. (2022). Ethanol Extract of Pinus koraiensis Leaves Mitigates High Fructose-Induced Hepatic Triglyceride Accumulation and Hypertriglyceridemia. Applied Sciences, 12(13), 6745. https://doi.org/10.3390/app12136745