
Sustainable Microalgae and Cyanobacteria Biotechnology

 , , , ,
, , , ,  and
and
Abstract
:1. Introduction
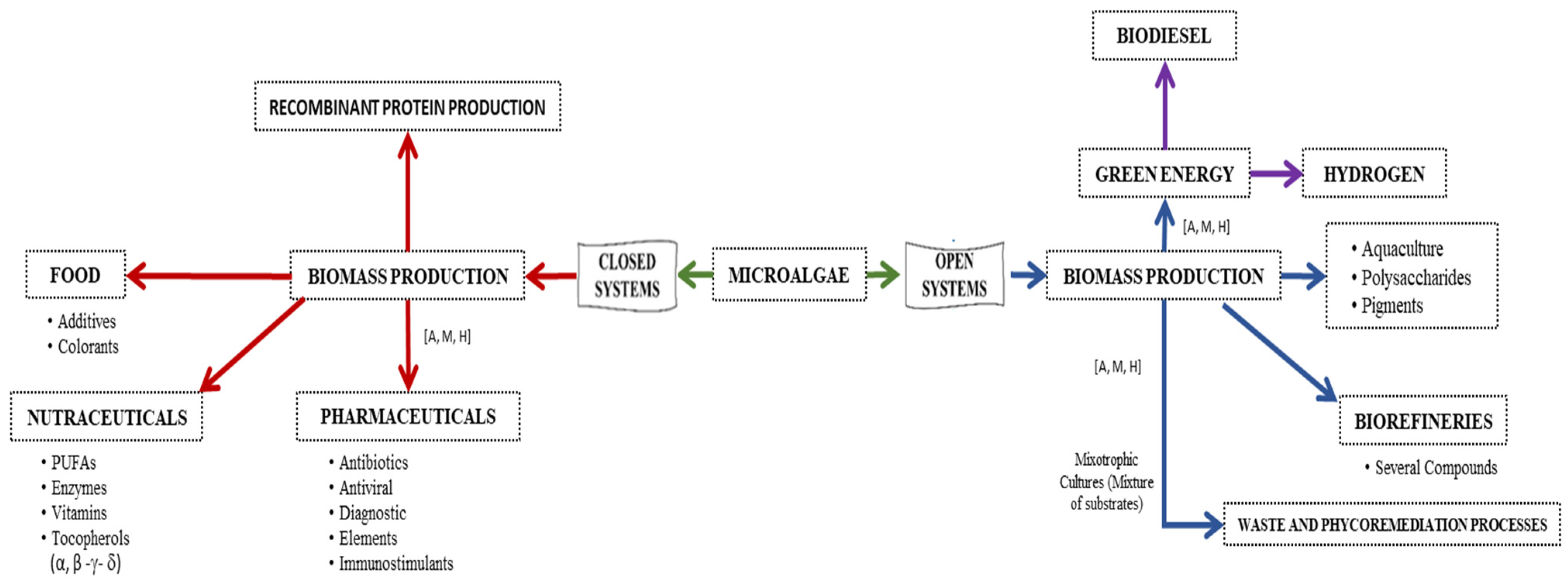
2. Applications of Biotechnology on Microalgae and Cyanobacteria Utilization
2.1. High Valuable Compounds from Microalgae
2.1.1. Pigments
Chlorophylls
Carotenoids
Phycobiliproteins
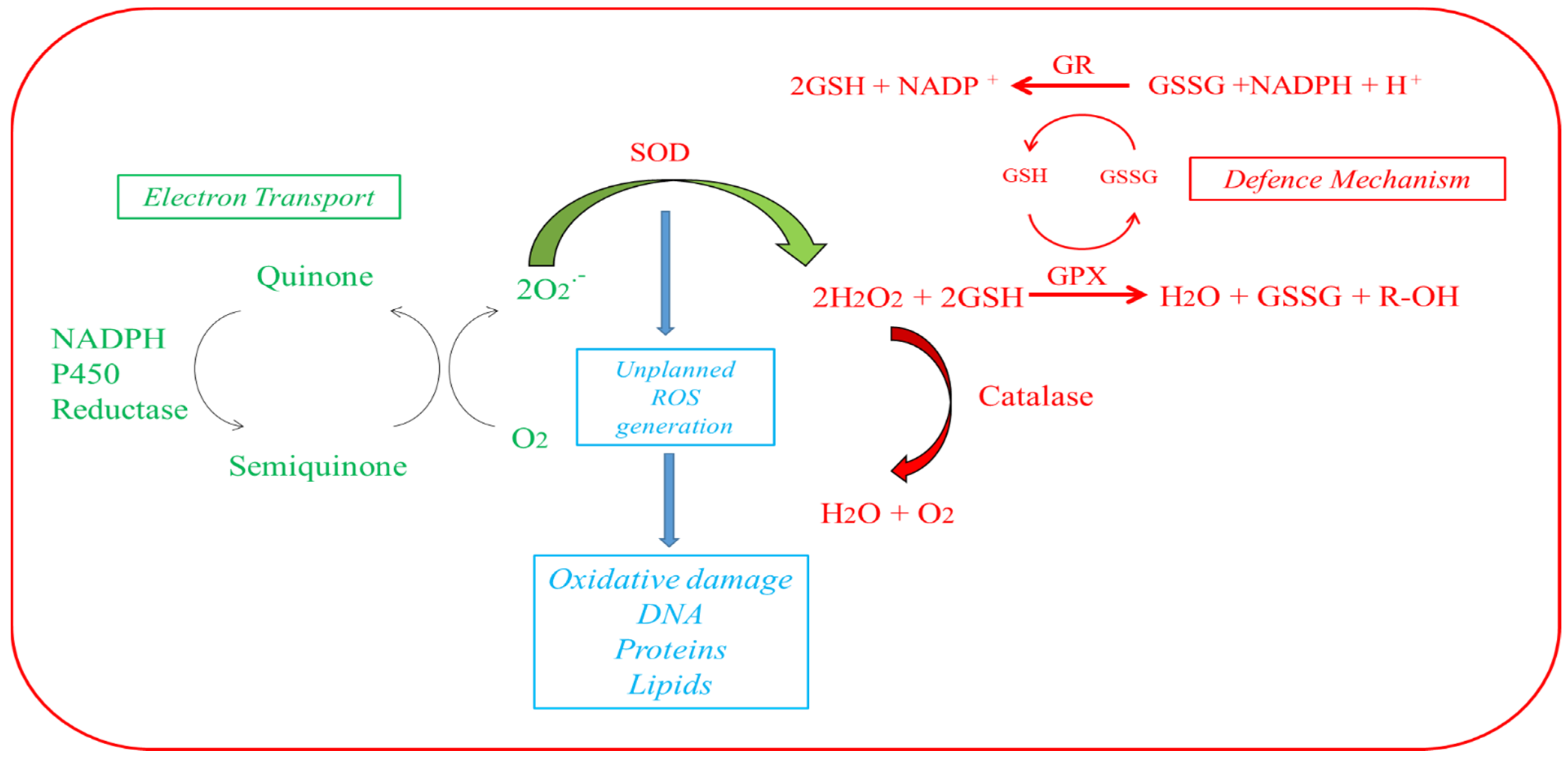
2.1.2. Superoxide Dismutase
2.1.3. Antibacterial, Antiviral, Anticancer, and Anti-Inflammatory Activities
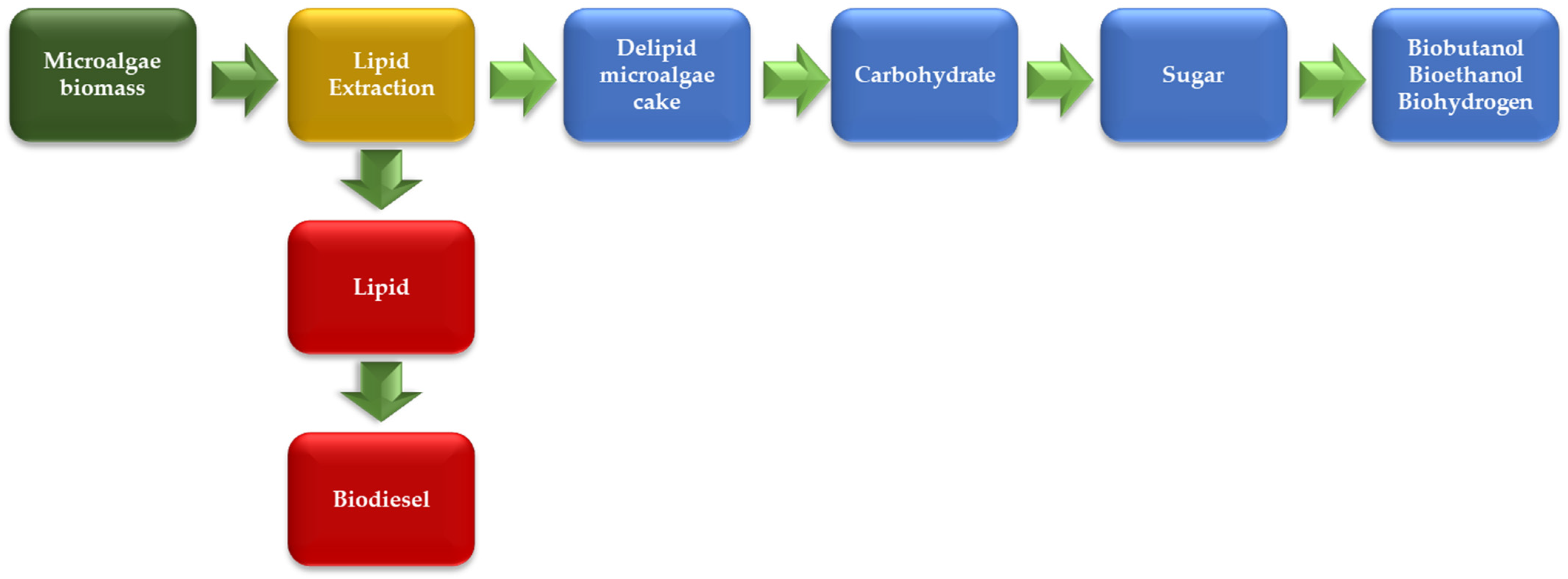
2.2. Biofuels from Microalgae
3. Implementation of an Efficient Production System: Mixotrophic Cultivation of Microalgae for Biodiesel Production
- (i)
- For the autotrophic fraction
- (ii)
- For the heterotrophic fraction
- and with the condition at final time = tf signifying that light irradiance is limiting and growth depends on organic carbon source consumption. In other words:
- the light saturation constant, , biomass produced per energy unit and illuminated area, this constant can be used as photobioreactor design criteria, because it involves the light energy absorbed by a specific microalgal specie and the amount of light irradiated in a specific photobioreactor geometry.
Kinetic of Biofuels Production by Microalgae
4. Other Important Applications of Microalgal Biotechnology
4.1. Additives for Animal Feeding
4.2. Phycoremediation
4.3. Waste Management
5. New Tools to Improve Microalgal Applications: Recombinant Protein Production in Microalgae
6. Concluding Remarks
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lutzu, G.A.; Zhang, L.; Zhang, Z.; Liu, T. Feasibility of attached cultivation for polysaccharides production by Porphyridium cruentum. Bioprocess Biosyst. Eng. 2017, 40, 73–83. [Google Scholar] [CrossRef] [PubMed]
- Montoya-Arroyo, A.; Lehnert, K.; Muñoz-González, A.; Schmid-Staiger, U.; Vetter, W.; Frank, J. Tocochromanol Profiles in Chlorella sorokiniana, Nannochloropsis limnetica and Tetraselmis suecica Confirm the Presence of 11′-α-Tocomonoenol in Cultured Microalgae Independently of Species and Origin. Foods 2022, 11, 396. [Google Scholar] [CrossRef] [PubMed]
- Alsenani, F.; Tupally, K.R.; Chua, E.T.; Eltanahy, E.; Alsufyani, H.; Parekh, H.S.; Peer, M.; Schenk, P.M. Evaluation of microalgae and cyanobacteria as potential sources of antimicrobial compounds. Saudi Pharm. J. 2020, 28, 1834–1841. [Google Scholar] [CrossRef] [PubMed]
- Nigam, S.; Sinha, S.; Srivastava, A.; Srivastava, A. Cultivation and Production Techniques of Marine Algae. In Encyclopedia of Marine Biotechnology; Kim, S.-K., Ed.; John Wiley & Sons Ltd.: London, UK, 2020; Chapter 9; pp. 327–340. [Google Scholar] [CrossRef]
- Güven, K.C.; Coban, B.; Özdemir, O. Pharmacology of Marine Macroalgae. In Encyclopedia of Marine Biotechnology; Kim, S.-K., Ed.; John Wiley & Sons Ltd.: London, UK, 2020; Chapter 20; pp. 585–615. [Google Scholar] [CrossRef]
- Dębowski, M.; Zieliński, M.; Świca, I.; Kazimierowicz, J. Algae Biomass as a Potential Source of Liquid Fuels. Phycology 2021, 1, 105–118. [Google Scholar] [CrossRef]
- Da Silva, T.L.; Gouveia, L.; Reis, A. Integrated microbial processes for biofuels and high value-added products: The way to improve the cost effectiveness of biofuel production. Appl. Microbiol. Biotechnol. 2014, 98, 1043–1053. [Google Scholar] [CrossRef]
- Kapoor, S.; Singh, M.; Srivastava, A.; Chavali, M.; Chandrasekhar, K.; Verma, P. Extraction and characterization of microalgae derived phenolics for pharmaceutical applications: A systematic review. J. Basic Microbiol. 2021, 1–20. [Google Scholar] [CrossRef]
- Madeira, M.S.; Cardoso, C.; Lopes, P.A.; Coelho, D.; Afonso, C.; Bandarra, N.M.; Prates, J.A.M. Microalgae as feed ingredientes for livestock production and meat quality: A review. Livest. Sci. 2017, 205, 11–121. [Google Scholar] [CrossRef]
- Tibbetts, S.M.; MacPherson, T.; McGinn, P.J.; Fredeen, A.H. In vitro digestion of microalgal biomass from freshwater species isolated in Alberta, Canada for monogastric and ruminant animal feed applications. Algal Res. 2016, 19, 324–332. [Google Scholar] [CrossRef] [Green Version]
- Couteau, C.; Coiffard, L. Microalgal Application in Cosmetics. In Microalgae in Health and Disease Prevention; Levine, I.A., Fleurence, J., Eds.; Academic Press: Cambridge, MA, USA, 2018; Chapter 15; pp. 317–323. [Google Scholar] [CrossRef]
- Bianchi, T.S.; Canuel, E.A. Photosynthetic Pigments: Chlorophylls, Carotenoids, and Phycobilins. In Chemical Biomarkers in Aquatic Ecosystems; Princeton Scholarship Online: Princeton, NJ, USA, 2011. [Google Scholar] [CrossRef]
- Eriksen, N.T. Research trends in the dominating microalgal pigments, β-carotene, astaxanthin, and phycocyanin used in feed, in foods, and in health applications. J. Nutr. Food Sci. 2016, 6, 507. [Google Scholar] [CrossRef]
- Singh, D.P.; Khattar, J.S.; Rajput, A.; Chaudhary, R.; Singh, R. High production of carotenoids by the green microalga Asterarcys quadricellulare PUMCC 5.1.1 under optimized culture conditions. PLoS ONE 2019, 14, e0221930. [Google Scholar] [CrossRef] [Green Version]
- Sahu, S.K.; Mantri, V.A.; Zheng, P.; Yao, N. Algae Biotechnology. In Encyclopedia of Marine Biotechnology; Kim, S.-K., Ed.; John Wiley & Sons Ltd.: London, UK, 2020; Chapter 1; pp. 1–31. [Google Scholar] [CrossRef]
- Igreja, W.; Maia, F.; Lopes, A.; Chisté, R. Biotechnological production of carotenoids using low cost-substrates is influenced by cultivation parameters: A review. Internat. J. Mol. Sci. 2021, 22, 8819. [Google Scholar] [CrossRef] [PubMed]
- Da Silva, J.C.; Lombardi, A.T. Chlorophylls in Microalgae: Occurrence, Distribution, and Biosynthesis. In Pigments from Microalgae Handbook; Jacob-Lopes, E., Queiroz, M., Zepka, L., Eds.; Springer: Cham, Switzerland, 2020; Chapter 1; pp. 1–18. [Google Scholar] [CrossRef]
- Masojídek, J.; Torzillo, G.; Koblížek, M. Photosynthesis in Microalgae. Appl. Phycol. Biotechnol. 2013, 20, 21–36. [Google Scholar]
- De Lima Petito, N.; Da Silva Dias, D.; Costa, V.G.; Falcão, D.Q.; De Lima Araujo, K.G. Increasing solubility of red bell pepper carotenoids by complexation with 2-hydroxypropyl-β-cyclodextrin. Food Chem. 2016, 208, 124–131. [Google Scholar] [CrossRef] [PubMed]
- Mishra, V.; Bacheti, R.; Husen, A. Medicinal uses of chlorophyll: A critical overview. In Chlorophyll: Structure, Function and Medicinal Uses; Le, H., Salcedo, E., Eds.; Nova Science Publishers, Inc.: New York, NY, USA, 2011; pp. 177–196. [Google Scholar]
- Ferruzzi, M.G.; Böhm, V.; Courtney, P.D.; Schwartz, S.J. Antioxidant and antimutagenic activity of dietary chlorophyll derivatives determined by radical scavenging and bacterial reverse mutagenesis assays. J. Food Sci. 2002, 67, 2589–2595. [Google Scholar] [CrossRef]
- Xu, Y.; Harvey, P.J. Carotenoid Production by Dunaliella salina under Red Light. Antioxidants 2019, 8, 123. [Google Scholar] [CrossRef] [Green Version]
- Janik, E.; Grudziński, W.; Gruszecki, W.I.; Krupa, Z. The xanthophyll cycle pigments in Secale cereale leaves under combined Cd and high light stress conditions. J. Photochem. Photobiol. B Biol. 2008, 90, 47–52. [Google Scholar] [CrossRef]
- Lokstein, H.; Renger, G.; Götze, J.P. Photosynthetic Light-Harvesting (Antenna) Complexes-Structures and Functions. Molecules 2021, 26, 3378. [Google Scholar] [CrossRef]
- Krinsky, N.I. Carotenoid Protection against Oxidation. Pure Appl. Chem. 1979, 51, 649–660. [Google Scholar] [CrossRef]
- Sommella, E.; Conte, G.; Salviati, E.; Pepe, G.; Bertamino, A.; Ostacolo, C.; Sansone, F.; Prete, F.; Aquino, R.; Campiglia, P. Fast Profiling of Natural Pigments in Different Spirulina (Arthrospira platensis) Dietary Supplements by DI-FT-ICR and Evaluation of their Antioxidant Potential by Pre-Column DPPH-UHPLC Assay. Molecules 2018, 23, 1152. [Google Scholar] [CrossRef] [Green Version]
- Moo-Young, M. Supercritical Fluid Technology in Bioseperation. In Comprehensive Biotechnology, 3rd ed.; Ramsey, E.D., Guo, W., Liu, J.Y., Wu, X.H., Eds.; Pergamon: Oxford, UK, 2019; pp. 713–724. [Google Scholar] [CrossRef]
- Stahl, W.; Sies, H. Bioactivity and protective effects of natural carotenoids. Biochim. Biophys. Acta (BBA)—Mol. Basis Dis. 2005, 1740, 101–107. [Google Scholar] [CrossRef] [Green Version]
- Shah, M.M.R.; Liang, Y.; Cheng, J.J.; Daroch, M. Astaxanthin-Producing Green Microalga Haematococcus pluvialis: From Single Cell to High Value Commercial Products. Front. Plant Sci. 2016, 7, 531. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liang, C.; Zhao, F.; Wei, W.; Wen, Z.; Qin, S. Carotenoid Biosynthesis in Cyanobacteria: Structural and Evolutionary Scenarios Based on Comparative Genomics. Int. J. Biol. Sci. 2006, 2, 197. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moreno, J.C.; Pizarro, L.; Fuentes, P.; Handford, M.; Cifuentes, V.; Stange, C. Levels of Lycopene β-Cyclase 1 Modulate Carotenoid Gene Expression and Accumulation in Daucus carota. PLoS ONE 2013, 8, e058144. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maresca, J.A.; Romberger, S.P.; Bryant, D.A. Isorenieratene Biosynthesis in Green Sulfur Bacteria Requires the Cooperative Actions of Two Carotenoid Cyclases. J. Bactereriol. 2008, 190, 6384–6391. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pagels, F.; Vasconcelos, V.; Guedes, A.C. Carotenoids from Cyanobacteria: Biotechnological Potential and Optimization Strategies. Biomolecules 2021, 11, 735. [Google Scholar] [CrossRef] [PubMed]
- Hayashi, M.; Chang, Y.M.; Wang, T.K.; Lin, S.H.; Knee, J.L. Ultrafast Spectroscopy and its Applications. Encycl. Phys. Sci. Technol. 2003, 17, 217–226. [Google Scholar] [CrossRef]
- El-Mohsnawy, E.; Abu-Khudir, R. A highly purified C-phycocyanin from thermophilic cyanobacterium Thermosynechococcus elongatus and its cytotoxic activity assessment using an in vitro cell-based approach. J. Taibah Univ. Sci. 2020, 14, 1218–1225. [Google Scholar] [CrossRef]
- Lin, J.Y.; Tan, S.I.; Yi, Y.C.; Hsiang, C.C.; Chang, C.H.; Chen, C.Y.; Chang, J.S.; Ng, I.S. High-level production and extraction of C-phycocyanin from cyanobacteria Synechococcus sp. PCC7002 for antioxidation, antibacterial and lead adsorption. Environ. Res. 2021, 206, 112283. [Google Scholar] [CrossRef]
- Miller, L.S.; Holt, S.C. Effect of carbon dioxide on pigment and membrane content in Synechococcus lividus. Arch. Microbiol. 1997, 115, 185–198. [Google Scholar] [CrossRef]
- Boussiba, S.; Richmond, A.E. C-phycocyanin as a storage protein in the blue-green alga Spirulina platensis. Arch. Microbiol. 1980, 125, 143–147. [Google Scholar] [CrossRef]
- Santiago-Santos, M.C.; Ponce-Noyola, T.; Olvera-Ramírez, R.; Ortega-López, J.; Cañizares-Villanueva, R.O. Extraction and purification of phycocyanin from Calothrix sp. Process Biochem. 2004, 39, 2047–2052. [Google Scholar] [CrossRef]
- Hao, S.; Yan, Y.; Li, S.; Zhao, L.; Zhang, C.; Liu, L.; Wang, C. The In Vitro Anti-Tumor Activity of Phycocyanin against Non-Small Cell Lung Cancer Cells. Mar. Drugs 2018, 16, 178. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singh, N.K.; Parmar, A.; Madamwar, D. Optimization of medium components for increased production of C-phycocyanin from Phormidium ceylanicum and its purification by single step process. Bioresour. Technol. 2009, 100, 1663–1669. [Google Scholar] [CrossRef] [PubMed]
- Ramos, A.; Acién, F.G.; Fernandez-Sevilla, J.M.; Gonzalez, C.V.; Bermejo, R. Development of a process for large-scale purification of C-phycocyanin from Synechocystis aquatilis using expanded bed adsorption chromatography. J. Chromatogr. B. 2011, 879, 511–519. [Google Scholar] [CrossRef]
- Mulshine, J.L.; Bingula, R.; Dupuis, C.; Chantal, P.; Berthon, J.Y.; Filaire, M.; Pigeon, L.; Filaire, E. Study of the Effects of Betaine and/or C-Phycocyanin on the Growth of Lung Cancer A549 Cells In Vitro and In Vivo. J. Oncol. 2016, 2016, 8162952. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Jovcevski, B.; Pukala, T.L. C-Phycocyanin from Spirulina Inhibits α-Synuclein and Amyloid-β Fibril Formation but Not Amorphous Aggregation. J. Nat. Prod. 2019, 82, 66–73. [Google Scholar] [CrossRef]
- Yang, W.; Wang, F.; Liu, L.N.; Sui, N. Responses of Membranes and the Photosynthetic Apparatus to Salt Stress in Cyanobacteria. Front. Plant Sci. 2020, 11, 713–723. [Google Scholar] [CrossRef]
- Okamoto, O.K.; Asano, C.S.; Aidar, E.; Colepicolo, P. Effect of cadmium on growth and superoxide dismutase activity of the marine microalgae Tetraselmis gracilis (Prasinophyceae). J. Phycol. 1996, 32, 74–79. [Google Scholar] [CrossRef]
- Saha, S.; Moane, S.; Murray, P. Effect of macro- and micro-nutrient limitation on superoxide dismutase activities and carotenoid levels in microalga Dunaliella salina CCAP 19/18. Bioresour. Technol. 2013, 147, 23–28. [Google Scholar] [CrossRef]
- Rossa, M.M.; De Oliveira, M.C.; Okamoto, O.K.; Lopes, P.F.; Colepicolo, P. Effect of visible light on superoxide dismutase (SOD) activity in the red alga Gracilariopsis tenuifrons (Gracilariales, Rhodophyta). J. Appl. Phycol. 2002, 14, 151–157. [Google Scholar] [CrossRef]
- Santiago-Morales, I.S.; López-Hernández, J.F.; Trujillo-Valle, L.; Márquez-Rocha, F.J. Tocopherols, Phycocyanin and Superoxide Dismutase from Microalgae: As Potential Food Antioxidants. Appl. Food Biotechnol. 2018, 5, 19–27. [Google Scholar] [CrossRef]
- Wood, N.J.; Sørensen, J. Catalase and superoxide dismutase activity in ammonia-oxidising bacteria. FEMS Microbiol. Ecol. 2001, 38, 53–58. [Google Scholar] [CrossRef]
- Lauritano, C.; Andersen, J.H.; Hansen, E.; Albrigtsen, M.; Escalera, L.; Esposito, F.; Hanssen, H.K.; Romano, G.; Ianora, A. Bioactivity Screening of Microalgae for Antioxidant, Anti-Inflammatory, Anticancer, Anti-Diabetes, and Antibacterial Activities. Front. Mar. Sci. 2016, 3, 68. [Google Scholar] [CrossRef] [Green Version]
- Carpine, R.; Sieber, S. Antibacterial and antiviral metabolites from cyanobacteria: Their application and their impact on human health. Curr. Res. Biotechnol. 2021, 3, 65–81. [Google Scholar] [CrossRef]
- Bhuyar, P.; Rahim, M.H.A.; Maniam, G.P.; Ramaraj, R.; Govindan, N. Exploration of bioactive compounds and antibacterial activity of marine blue-green microalgae (Oscillatoria sp.) isolated from coastal region of west Malaysia. SN Appl. Sci. 2020, 2, 1906. [Google Scholar] [CrossRef]
- Kellam, S.J.; Walker, J.M. Antibacterial activity from marine microalgae in laboratory culture. Br. Phycol. J. 2007, 24, 191–194. [Google Scholar] [CrossRef]
- Mashhadinejad, A.; Zamani, H.; Sarmad, J. Effect of growth conditions and extraction solvents on enhancement of antimicrobial activity of the microalgae Chlorella vulgaris. Pharm. Biomed. Res. 2016, 2, 65–73. [Google Scholar] [CrossRef] [Green Version]
- Maadane, A.; Merghoub, N.; El Mernissi, N.; Ainane, T.; Amzazi, S.; Wahby, I.; Bakri, Y. Antimicrobial activity of marine microalgae isolated from Moroccan coastlines. J. Microbiol. Biotechnol. Food Sci. 2017, 6, 1257–1260. [Google Scholar] [CrossRef] [Green Version]
- Plaza, M.; Santoyo, S.; Jaime, L.; García-Blairsy, R.; Herrero, M.; Señoráns, F.J.; Ibáñez, E. Screening for bioactive compounds from algae. J. Pharm. Biomed. Anal. 2010, 51, 450–455. [Google Scholar] [CrossRef]
- Ip, P.F.; Chen, F. Production of astaxanthin by the green microalga Chlorella zofingiensis in the dark. Process Biochem. 2005, 40, 733–738. [Google Scholar] [CrossRef]
- Olasehinde, T.A.; Odjadjare, E.C.; Mabinya, L.V.; Olaniran, A.O.; Okoh, A.I. Chlorella sorokiniana and Chlorella minutissima exhibit antioxidant potentials, inhibit cholinesterases and modulate disaggregation of β-amyloid fibrils. Electron. J. Biotechnol. 2019, 40, 1–9. [Google Scholar] [CrossRef]
- Parikh, A.; Madamwar, D. Partial characterization of extracellular polysaccharides from cyanobacteria. Biores. Technol. 2006, 97, 1822–1827. [Google Scholar] [CrossRef]
- Sarada, R.; Pillai, M.G.; Ravishankar, G.A. Phycocyanin from Spirulina sp.: Influence of processing of biomass on phycocyanin yield, analysis of efficacy of extraction methods and stability studies on phycocyanin. Process Biochem. 1999, 34, 795–801. [Google Scholar] [CrossRef]
- Taufiqurrahmi, N.; Religia, P.; Mulyani, G.; Suryana, D.; Tanjung, F.A.; Arifin, Y. Phycocyanin extraction in Spirulina produced using agricultural waste. IOP Conf. Ser. Mater. Sci. Eng. 2017, 206, 012097. [Google Scholar] [CrossRef] [Green Version]
- Ha, K.S.; Thompson, G.A. Diacylglycerol Metabolism in the Green Alga Dunaliella salina under Osmotic Stress Possible Role of Diacylglycerols in Phospholipase C-Mediated Signal Transduction. Plant Physiol. 1991, 97, 921–927. [Google Scholar] [CrossRef] [Green Version]
- Zienkiewicz, K.; Zienkiewicz, A.; Poliner, E.; Du, Z.Y.; Vollheyde, K.; Herrfurth, C.; Marmon, S.; Farré, E.M.; Feussner, I.; Benning, C. Nannochloropsis, a rich source of diacylglycerol acyltransferases for engineering of triacylglycerol content in different hosts. Biotechnol. Biofuels 2017, 10, 8. [Google Scholar] [CrossRef] [Green Version]
- Hosseinzadeh, G.N.; Valizadeh, M.; Dorani, E.; Hejazi, M.A. Biochemical profiling of three indigenous Dunaliella isolates with main focus on fatty acid composition towards potential biotechnological application. Biotechnol. Rep. 2020, 26, e00479. [Google Scholar] [CrossRef]
- Chisti, Y. Biodiesel from microalgae. Biotechnol. Adv. 2007, 25, 294–306. [Google Scholar] [CrossRef]
- Huang, W.; Kim, J. Nickel oxide nanoparticle-based method for simultaneous harvesting and disruption of microalgal cells. Bioresour. Technol. 2016, 218, 1290–1293. [Google Scholar] [CrossRef]
- Soru, S.; Malavasi, V.; Caboni, P.; Concas, A.; Cao, G. Behavior of the extremophile green alga Coccomyxa melkonianii SCCA 048 in terms of lipids production and morphology at different pH values. Extremophiles 2019, 23, 79–89. [Google Scholar] [CrossRef]
- De Farias Silva, C.E.; Bertucco, A. Bioethanol from microalgae and cyanobacteria: A review and technological outlook. Process Biochem. 2016, 51, 1833–1842. [Google Scholar] [CrossRef]
- Phwan, C.K.; Chew, K.W.; Sebayang, A.H.; Ong, H.C.; Ling, T.C.; Malek, M.A.; Ho, Y.C.; Show, P.L. Effects of acids pre-treatment on the microbial fermentation process for bioethanol production from microalgae. Biotechnol. Biofuels 2019, 12, 191–198. [Google Scholar] [CrossRef] [Green Version]
- El-Mekkawi, S.A.; Abdo, S.M.; Samhan, F.A.; Ali, G.H. Optimization of some fermentation conditions for bioethanol production from microalgae using response surface method. Bull. Natl. Res. Cent. 2019, 43, 164–171. [Google Scholar] [CrossRef]
- Kassim, M.A.; Tan, K.M.; Serri, N.A. Enzymatic hydrolysis of dilute alkaline pretreated Chlorella sp. biomass for biosugar production and fed-batch bioethanol fermentation. Asia-Pac. J. Sci. Technol. 2019, 24, 1–16. [Google Scholar] [CrossRef]
- Harun, R.; Jason, W.S.Y.; Cherrington, T.; Danquah, M.K. Exploring alkaline pre-treatment of microalgal biomass for bioethanol production. Appl. Enerey 2011, 88, 3464–3467. [Google Scholar] [CrossRef]
- Onay, M. Bioethanol production from Nannochloropsis gaditana in municipal wastewater. Energy Procedia 2018, 153, 253–257. [Google Scholar] [CrossRef]
- Fetyan, N.; El-Khair, A.; El-Sayed, B.; Ibrahim, F.M.; Sadik, M.W. Bioethanol production from defatted biomass of Nannochloropsis oculata microalgae grown under mixotrophic conditions. Environ. Sci. Pollut. Res. 2021, 1, 2588–2597. [Google Scholar] [CrossRef]
- Varela-Bojórquez, N.; Vélez-De La Rocha, R.; Ángel, M.; Escalante, A.; Sañudo-Barajas, J.A. Production of bioethanol from biomass of microalgae Dunaliella tertiolecta. Int. J. Environ. Agric. Res. 2016, 2, 110–116. [Google Scholar]
- Kassim, M.A.; Meng, T.K. Carbon dioxide (CO2) biofixation by microalgae and its potential for biorefinery and biofuel production. Sci. Total Environ. 2017, 584–585, 1121–1129. [Google Scholar] [CrossRef]
- Kim, E.J.; Kim, S.; Choi, H.G.; Han, S.J. Co-production of biodiesel and bioethanol using psychrophilic microalga Chlamydomonas sp. KNM0029C isolated from Arctic Sea ice. Biotechnol. Biofuels 2020, 13, 20. [Google Scholar] [CrossRef]
- Lee, O.K.; Kim, A.L.; Seong, D.H.; Lee, C.G.; Jung, Y.T.; Lee, J.W.; Lee, E.Y. Chemo-enzymatic saccharification and bioethanol fermentation of lipid-extracted residual biomass of the microalga, Dunaliella tertiolecta. Bioresour. Technol. 2013, 132, 197–201. [Google Scholar] [CrossRef] [PubMed]
- Lewis Oscar, F.; Praveenkum, R.; Thajuddin, N. Bioethanol Production Using Starch Extracted from Microalga Stigeoclonium sp., Kutz. BUM11007 Cultivated in Domestic Wastewater. Res. J. Environ. Sci. 2015, 9, 216–224. [Google Scholar] [CrossRef] [Green Version]
- Li, S.; Huang, L.; Ke, C.; Pang, Z.; Liu, L. Pathway dissection, regulation, engineering and application: Lessons learned from biobutanol production by solventogenic clostridia. Biotechnol. Biofuels 2020, 13, 39. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hong, Y.; Chen, C.; Wu, Y. Biobutanol production from sulfuric acid-pretreated red algal biomass by a newly isolated Clostridium sp. strain WK. Biotechnol. Appl. Biochem. 2020, 67, 738–743. [Google Scholar] [CrossRef] [PubMed]
- Figueroa-Torres, G.M.; Wan Mahmood, W.M.A.; Pittman, J.K.; Theodoropoulos, C. Microalgal biomass as a biorefinery platform for biobutanol and biodiesel production. Biochem. Eng. J. 2020, 153, 107396. [Google Scholar] [CrossRef]
- Kristiawan, O.; Tambunan, U.S.F. Biobutanol Production from Microalgae Nannochloropsis sp. Biomasses by Clostridium acetobutylicum Fermentation. Sci. Contrib. Oil Gas 2020, 43, 91–98. [Google Scholar] [CrossRef]
- Onay, M. Biomass and bio-butanol production from Borodinellopsis texensis CCALA 892 in synthetic wastewater: Determination of biochemical composition. J. Nat. Appl. Sci. 2020, 24, 306–316. [Google Scholar] [CrossRef]
- Zhu, L. Microalgal culture strategies for biofuel production: A review. Biofuels Bioprod. Biorefining 2015, 9, 801–814. [Google Scholar] [CrossRef]
- Hosikian, A.; Lim, S.; Halim, R.; Danguah, M.L. Chlorophyll extraction from microalgae: A review on the Process Engineering Aspects. Int. J. Chem. Eng. 2010, 391632. [Google Scholar] [CrossRef] [Green Version]
- Zili, F.; Bouzidi, N.; Ammar, J.; Zakhama, W.; Ghoul, M.; Sayadi, S.; Ouada, H.B. Mixotrophic cultivation promotes growth, lipid productivity, and PUFA production of a thermophilic Chlorophyta strain related to the genus Graesiella. J. Appl. Phycol. 2016, 29, 35–43. [Google Scholar] [CrossRef]
- Pérez-García, O.; de-Bashan, L.E.; Hernández, J.P.; Bashan, Y. Efficiency of growth and nutrient uptake from wastewater by heterotrophic, autotrophic, and mixotrophic cultivation of Chlorella vulgaris immobilized with Azospirillum brasilense. J. Phycol. 2010, 46, 800–812. [Google Scholar] [CrossRef]
- Venkata, M.S.; Rohit, M.V.; Chiranjeevi, P.; Chandra, R.; Navaneeth, B. Heterotrophic microalgae cultivation to synergize biodiesel production with waste remediation: Progress and perspectives. Bioresour. Technol. 2015, 184, 169–178. [Google Scholar] [CrossRef] [PubMed]
- Ogbonna, J.C.; McHenry, M.P. Culture Systems Incorporating Heterotrophic Metabolism for Biodiesel Oil Production by Microalgae. In Biomass and Biofuels from Microalgae; Advances in Engineering and Biology; Moheimani, N.R., McHenry, M.P., Boe, K., Bahri, P.A., Eds.; Springer International Publishing: New York, NY, USA, 2015; pp. 63–74. [Google Scholar]
- Márquez, F.; Sasaki, K.; Kakizono, T.; Nishio, N.; Nagai, S. Growth characteristics of Spirulina platensis in mixotrophic and heterotrophic conditions. J. Ferment. Bioeng. 1993, 76, 408–410. [Google Scholar] [CrossRef]
- Márquez, F.; Sasaki, K.; Nishio, N.; Nagai, S. Enhancement of biomass and pigment production during growth of Spirulina platensis in Mixotrophic Culture. J. Chem. Technol. Biotechnol. 1995, 62, 159–164. [Google Scholar] [CrossRef]
- Laraib, N.; Manzoor, M.; Javid, A.; Jabeen, F.; Bukhari, S.M.; Ali, W.; Hussain, A. Mixotrophic cultivation of Chlorella vulgaris in sugarcane molasses preceding nitrogen starvation: Biomass productivity, lipid content and fatty acid analyses. Environ. Prog. Sustain. Energy 2021, 40, e13625. [Google Scholar] [CrossRef]
- Laraib, N.; Hussain, A.; Javid, A.; Bukahari, S.M.; Ali, W.; Mazoor, M.; Jabeen, F. Mixotrophic Cultivation of Scenedesmus dimorphus for Enhancing Biomass Productivity and Lipid Yield. Iran. J Sci. Technol. Trans. Sci. 2012, 45, 397–403. [Google Scholar] [CrossRef]
- Manzoor, M.; Ahamad, Q.; Aslam, A.; Jabeen, F.; Rasul, A.; Schenk, P.M.; Qazi, J.I. Mixotrophic cultivation of Scenedesmus dimorphus in sugarcane bagasse hydrolysate. Environ. Prog. Sustain. Energy 2020, 39, e13334. [Google Scholar] [CrossRef]
- Arora, N.; Philippidis, G.P. Insights into the physiology of Chlorella vulgaris cultivated in sweet sorghum bagasse hydrolysate for sustainable algal biomass and lipid production. Sci. Rep. 2021, 11, 6779. [Google Scholar] [CrossRef]
- Tosuner, V.; Ozturk Urek, R. Simultaneous Lipid Production and Valorization of Crude Glycerol by Mixotrophic and Heterotrophic Cultivation of Arthrospira platensis. J. Sci. Ind. Res. 2021, 80, 850–857. [Google Scholar]
- Ogbonna, I.O.; Ogbonna, J.C. Effects of Carbon Source on Growth Characteristics and Lipid Accumulation by Microalga Dictyosphaerium sp. with Potential for Biodiesel Production. Energy Power Eng. 2018, 10, 29–42. [Google Scholar] [CrossRef] [Green Version]
- Hamilton, P.; Proulx, M.; Earle, C. Enumerating phytoplankton with an upright compound microscope using a modified settling chamber. Hydrobiologia 2001, 444, 171–175. [Google Scholar] [CrossRef]
- Jakobsen, H.; Carstensen, J. FlowCAM. Sizing cells and understanding the impact of size distribution on biovolume of planktonic community structure. Aquat. Microbial. Ecol. 2011, 65, 75–87. [Google Scholar] [CrossRef] [Green Version]
- Singh, J.; Saxena, R.C. An introduction to microalgae. In Handbook of Marine Microalgae; Elsevier Science: Amsterdam, The Netherlands, 2015; pp. 11–24. [Google Scholar]
- Beardall, J.; Stojkovic, S. Microalgae under global environmental change: Implications for growth and productivity, populations and trophic flow. ScienceAsia 2006, 32 (Suppl. 1), 1–10. [Google Scholar] [CrossRef]
- Molazadeh, M.; Ahmadzadeh, H.; Pourianfar, H.R.; Lyon, S.; Rampelotto, P.H. The use of microalgae for coupling wastewater treatment with CO2 biofixation. Front. Bioeng. Biotechnol. 2019, 7, 42. [Google Scholar] [CrossRef]
- Harrison, P.J.; Berges, J.A. Marine culture media. In Algal Culturing Techniques; Anderson, R., Ed.; Academic Press: New York, NY, USA, 2005; Chapter 3; pp. 21–33. [Google Scholar]
- Liu, J.; Sun, Z.; Gerken, H. Omega-3 Polyunsaturated Free Fatty Acidsfrom Algae. In Recent Advances in Microalgal Biotechnology; Liu, J., Sun, Z., Gerken, H., Eds.; OMICS Group eBook: Foster City, CA, USA, 2015. [Google Scholar]
- Liang, S.; Xueming, L.; Chen, F.; Chen, Z. Current microalgal health food R&D activities in China. Hydrobiologia 2004, 512, 45–48. [Google Scholar]
- Spolaore, P.; Joannis-Cassan, C.; Duran, E.; Isambert, A. Commercial applications of microalgae. J. Biosci. Bioeng. 2006, 101, 87–96. [Google Scholar] [CrossRef] [Green Version]
- Borowitzka, M.A. Vitamins and fine chemicals from microalgae. In Microalgal Biotechnology; Borowitzka, M.A., Borowitzka, L.J., Eds.; Cambridge University Press: Cambridge, UK, 1998; pp. 96–153. [Google Scholar]
- Guil-Guerrero, J.L.; Navarro-Juarez, R.; Lopez-Martinez, J.C.; Campra-Madrid, P.; Rebolloso-Fuentes, M.M. Functionnal properties of the biomass of three microalgal species. J. Food Eng. 2004, 65, 511–517. [Google Scholar] [CrossRef]
- Mahajan, G.; Kamat, M. g-Linolenic acid production from Spirulina platensis. Appl. Microbiol. Biotechnol. 1995, 43, 466–469. [Google Scholar] [CrossRef]
- Griffiths, M.J.; Harrison, S.T. Lipid productivity as a key characteristic for choosing algal species for biodiesel production. J. Appl. Phycol. 2009, 21, 493–507. [Google Scholar] [CrossRef]
- West, M.; Zubeck, M. Evaluation of microalgae for use as nutraceuticals and nutritional supplements. J. Nutr. Food Sci. 2012, 2, 147. [Google Scholar] [CrossRef] [Green Version]
- Borowitzka, M.A. High-value products from microalgae-their development and commercialisation. J. Appl. Phycol. 2013, 25, 743–756. [Google Scholar] [CrossRef]
- López-Hernandez, J.F.; Peña-Marin, E.S.; Martínez-García, R.; Camarillo-Coop, S.; Márquez-Rocha, F.J.; Alvarez-González, C.A. Properties of microalgae as functional ingredient for the tropical Gar (Atractosteus tropicus, Gill 1863) aquaculture. In Proceedings of the XVI International Symposium on Aquaculture Nutrition, Tabasco, México, 29–31 March 2022. [Google Scholar]
- Gaikwad, R.W.; Manjiry, D.G.; Bhagat, S.L. Carbon dioxide capture, tolerance and sequestration using microalgae a review. Int. J. Pharm. Chem. Biol. Sci. 2016, 6, 345–349. [Google Scholar]
- Wang, J.; Yang, H.; Wang, F. Mixotrophic cultivation of microalgae for biodiesel production: Status and prospects. Appl. Biochem. Biotechnol. 2014, 172, 3307–3329. [Google Scholar] [CrossRef] [PubMed]
- Miao, X.; Wu, Q. High yield bio-oil production from fast pyrolysis by metabolic controlling of Chlorella protothecoides. J. Biotechnol. 2004, 110, 85–93. [Google Scholar] [CrossRef] [PubMed]
- Verma, R.; Kumari, K.V.L.K.; Srivastava, A.; Kumar, A. Photoautotrophic, mixotrophic, and heterotrophic culture media optimization for enhanced microalgae production. J. Environ. Chem. Eng. 2020, 8, 104149. [Google Scholar] [CrossRef]
- Ji, M.K.; Kim, H.C.; Sapireddy, V.R.; Yun, H.S.; Abou-Shanab, R.A.; Choi, J.; Lee, W.; Timmes, T.C.; Jeon, B.H. Simultaneous nutrient removal and lipid production from pretreated piggery wastewater by Chlorella vulgaris YSW-04. Appl. Microbiol. Biotechnol. 2013, 97, 2701–2710. [Google Scholar] [CrossRef]
- Park, H.; Lee, C. Theoretical calculations on the feasibility of microalgal biofuels: Utilization of marine resources could help realizing the potential of microalgae. Biotechnol. J. 2016, 11, 1461–1470. [Google Scholar] [CrossRef]
- Khan, M.I.; Shin, J.H.; Kim, J.D. The promising future of microalgae: Current status, challenges, and optimization of a sustainable and renewable industry for biofuels, feed, and other products. Microb. Cell Factories 2018, 17, 36. [Google Scholar] [CrossRef]
- Venkata-Mohan, S. Reorienting Waste Remediation towards Harnessing Bioenergy: A Paradigm Shift. In Industrial Wastewater Treatment, Recycling and Reuse; Vivek, V., Ranade, V., Bhandari, M., Eds.; Butterworth-Heinemann: Oxford, UK, 2014; Chapter 6; pp. 235–281. [Google Scholar]
- Venkata-Mohan, S.; Goud, R.K. Pretreatment of biocatalyst as viable option for sustained production of biohydrogen from wastewater treatment. In Biogas Production: Pretreatment Methods in Anaerobic Digestion; Mudhoo, A., Ed.; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2012. [Google Scholar]
- Goud, R.K.; Venkata-Mohan, S. Acidic and alkaline shock pretreatment to enrich acidogenic biohydrogen producing mixed culture: Long term synergetic evaluation of microbial inventory, dehydrogenase activity and bio-electro kinetics. RSC Adv. 2012, 2, 6336–6353. [Google Scholar] [CrossRef]
- Sarkar, O.; Goud, R.K.; Subash, G.V.; Venkata-Mohan, S. Relative effect of different inorganic acids on selective enrichment of acidogenic biocatalyst for fermentative biohydrogen production from wastewater. Bioresour. Technol. 2013, 147, 321–331. [Google Scholar] [CrossRef]
- Lalit-Babu, V.; VenkataMohan, S.; Sarma, P.N. Influence of reactor configuration on fermentative hydrogen production during wastewater treatment. Int. J. Hydrogen Energy 2009, 34, 3305–3312. [Google Scholar] [CrossRef]
- Saratale, G.D.; Chen, S.; Lo, Y.; Saratale, J.L.G.; Chang, J. Outlook of biohydrogen production from lignocellulosic feedstock using dark fermentation—A review. J. Sci. Ind. Res. 2008, 67, 962–979. [Google Scholar]
- Abdel-Raouf, N.; Al-Homaidan, A.A.; Ibraheem, I.B. Microalgae and wastewater treatment. Saudi J. Biol. Sci. 2012, 19, 257–275. [Google Scholar] [CrossRef] [Green Version]
- Velvizhi, G.; Venkata-Mohan, S. In situ system buffering capacity dynamics on bioelectrogenic activity during the remediation of wastewater in microbial fuel cell. Environ. Prog. Sustain. Energy 2013, 32, 454–462. [Google Scholar] [CrossRef]
- Velvizhi, G.; Venkata-Mohan, S. Electrogenic activity and electron losses under increasing organic load of recalcitrant pharmaceutical wastewater. Int. J. Hydrogen Energy 2013, 37, 5969–5978. [Google Scholar] [CrossRef]
- Naik, S.N.; Goud, V.V.; Rout, P.K.; Dalai, A.K. Production of first- and second-generation biofuels: A comprehensive review. Renew. Sustain. Energy Rev. 2010, 14, 578–597. [Google Scholar] [CrossRef]
- Singh, A.P.; Singh, N.; Murphy, J.D. Renewable fuels from algae: An answer to debatable land-based fuel. Bioresour. Technol. 2011, 102, 10–16. [Google Scholar] [CrossRef]
- Rashid, N.; Rehman, M.S.U.; Memon, S.; Ur Rahman, Z.; Lee, K.; Han, J.I. Current status, barriers and developments in biohydrogen production by microalgae. Renew. Sustain. Energy Rev. 2013, 22, 571–579. [Google Scholar] [CrossRef]
- Kim, M.K.; Park, J.W.; Park, C.S.; Kim, S.J.; Jeune, K.H.; Chang, M.U.; Acreman, J. Enhanced production of Scenedesmus spp. (green microalgae) using a new medium containing fermented swine wastewater. Bioresour. Technol. 2007, 98, 2220–2228. [Google Scholar] [CrossRef]
- Pulz, O.; Gross, G. Valuable products from biotechnology of microalgae. Appl. Microbiol. Biotechnol. 2004, 65, 635–648. [Google Scholar] [CrossRef]
- Usher, P.K.; Ross, A.B.; Camargo-Valero, M.A.; Tomlin, A.S.; Gale, W.F. An overview of the potential environmental impacts of large-scale microalgae cultivation. Biofuels 2014, 5, 331–349. [Google Scholar] [CrossRef]
- Cheah, W.Y.; Show, P.L.; Chang, J.S.; Ling, T.C.; Juan, J.C. Biosequestration of atmospheric CO2 and flue gas containing CO2 by microalgae. Bioresour. Technol. 2015, 184, 190–201. [Google Scholar] [CrossRef] [PubMed]
- Phuc, V.P. Medical Biotechnology: Techniques and Applications. In Omics Technologies and Bioengineering; Barh, D., Azevedo, V., Eds.; Academic Press: Cambridge, MA, USA, 2018; Chapter 19; pp. 449–469. [Google Scholar] [CrossRef]
- Bartfai, T.; Lees, G.V. (Eds.) The need for medicines grows. In The Future of Drug Discovery; Academic Press: Cambridge, MA, USA, 2013; Chapter 2; pp. 31–53. [Google Scholar] [CrossRef]
- Vaishnav, P.; Demain, A.L. Industrial Biotechnology, (overview). In Encyclopedia of Microbiology, 3rd ed.; Schaechter, M., Ed.; Academic Press: Cambridge, MA, USA, 2009; pp. 335–348. [Google Scholar] [CrossRef]
- Rosales-Mendoza, S.; García-Silva, I.; González-Ortega, O.; Sandoval-Vargas, J.M.; Malla, A.; Vimolmangkang, S. The Potential of Algal Biotechnology to Produce Antiviral Compounds and Biopharmaceuticals. Molecules 2020, 25, 4049. [Google Scholar] [CrossRef] [PubMed]
- Shih, S.M.; Doran, P.M. Foreign protein production using plant cell and organ cultures: Advantages and limitations. Biotechnol. Adv. 2009, 27, 1036–1042. [Google Scholar] [CrossRef] [PubMed]
- Dhara, V.G.; Naik, H.M.; Majewska, N.I.; Betenbaugh, M.J. Recombinant antibody production in CHO and NS0 cells: Differences and similarities. BioDrugs 2018, 32, 571–584. [Google Scholar] [CrossRef]
- Kis, Z.; Shattock, R.; Shah, N.; Kontoravdi, C. Emerging technologies for low-cost, rapid vaccine manufacture. Biotechnol. J. 2019, 14, 1800376. [Google Scholar] [CrossRef] [Green Version]
- Chen, Q.; Davis, K.R. The potential of plants as a system for the development and production of human biologics. F1000Research 2016, 5, F1000. [Google Scholar] [CrossRef] [Green Version]
- Burnett, M.J.B.; Burnett, A.C. Therapeutic recombinant protein production in plants: Challenges and opportunities. Plants People Planet 2020, 2, 121–132. [Google Scholar] [CrossRef]
- Shanmugaraj, B.; Bulaon, C.J.; Phoolcharoen, W. Plant molecular farming: A viable platform for recombinant biopharmaceutical production. Plants 2020, 9, 842. [Google Scholar] [CrossRef]
- Specht, E.; Miyake-Stoner, S.; Mayfield, S. Micro-algae come of age as a platform for recombinant protein production. Biotechnol. Lett. 2010, 32, 1373–1383. [Google Scholar] [CrossRef] [Green Version]
- Specht, E.; Mayfield, S. Algae-based oral recombinant vaccines. Front. Microbiol. 2014, 5, 60. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ahmad, N.; Mehmood, M.A.; Malik, S. Recombinant protein production in microalgae: Emerging trends. Protein Pept. Lett. 2020, 27, 105–110. [Google Scholar] [CrossRef] [PubMed]
- Malla, A.; Rosales-Mendoza, S.; Phoolcharoen, W.; Vimolmangkang, S. Efficient Transient Expression of Recombinant Proteins Using DNA Viral Vectors in Freshwater Microalgal Species. Front. Plant Sci. 2021, 12, 650820. [Google Scholar] [CrossRef] [PubMed]
- Diamos, A.G.; Mason, H.S. Modifying the replication of geminiviral vectors reduces cell death and enhances expression of biopharmaceutical proteins in Nicotiana benthamiana Leaves. Front. Plant Sci. 2019, 9, 1974. [Google Scholar] [CrossRef]
- Rosenberg, J.N.; Oyler, G.A.; Wilkinson, L.; Betenbaugh, M.J. A green light for engineered algae: Redirecting metabolism to fuel a biotechnology revolution. Curr. Opin. Biotechnol. 2008, 19, 430–436. [Google Scholar] [CrossRef]
- Potvin, G.; Zhang, Z. Strategies for high-level recombinant protein expression in transgenic microalgae: A review. Biotechnol. Adv. 2010, 28, 910–918. [Google Scholar] [CrossRef]
- Yaakob, Z.; Ali, E.; Zainal, A.; Mohamad, M.; Takriff, M.S. An overview: Biomolecules from microalgae for animal feed and aquaculture. J. Biol. Res. Thessaloniki. 2014, 21, 6. [Google Scholar] [CrossRef] [Green Version]
- Rasala, B.A.; Mayfield, S.P. Photosynthetic biomanufacturing in green algae; production of recombinant proteins for industrial, nutritional, and medical uses. Photosynth. Res. 2015, 123, 227–239. [Google Scholar] [CrossRef]
- Salazar-González, J.A.; Bañuelos-Hernández, B.; Rosales-Mendoza, S. Current status of viral expression systems in plants and perspectives for oral vaccines development. Plant Mol. Biol. 2015, 87, 203–217. [Google Scholar] [CrossRef]
- Mochdia, K.; Tamaki, S. Transcription Factor-Based Genetic Engineering in Microalgae. Plants. 2021, 10, 1602. [Google Scholar] [CrossRef]
- Valledor, L.; Furuhashi, T.; Recuenco-Munoz, L.; Wienkoop, S.; Weckwerth, W. System-level network analysis of nitrogen starvation and recovery in Chlamydomonas reinhardtii reveals potential new targets for increased lipid accumulation. Biotechnol. Biofuels. 2014, 7, 171. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jia, B.; Xie, X.; Wu, M.; Lin, Z.; Yin, J.; Huang, Y.; Hu, Z. Understanding the functions of endogenous DOF transcript factor in Chlamydomonas reinhardtii. Biotechnol. Biofuels 2019, 12, 67. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.U. Lipid Metabolism in Plants. Plants 2020, 9, 871. [Google Scholar] [CrossRef] [PubMed]
- Bajhaiya, A.K.; Dean, A.P.; Zeef, L.A.H.; Webster, R.E.; Pittman, J.K. PSR1 is a Global Transcriptional Regulator of Phosphorus Deficiency Responses and Carbon Storage Metabolism in Chlamydomonas reinhardtii. Plant Physiol. 2016, 170, 1216–1234. [Google Scholar] [CrossRef] [Green Version]
- Hidayati, N.A.; Oshima, Y.; Iwai, M.; Yamano, T.; Kajikawa, M.; Sakurai, N.; Suda, K.; Sesoko, K.; Hori, K.; Obayashi, T.; et al. Lipid Remodeling Regulator 1 (LRL1) Is Differently Involved in the Phosphorus-Depletion Response from PSR1 in Chlamydomonas Reinhardtii. Plant J. 2019, 100, 610–626. [Google Scholar] [CrossRef] [Green Version]
- Sun, X.; Perera, S.; Haas, N.; Lefebvre, P.A.; Silflow, C.D. Using an RSP3 reporter gene system to investigate molecular regulation of hydrogenase expression in Chlamydomonas reinhardtii. Algal Res. 2013, 2, 341–351. [Google Scholar] [CrossRef]
- Toepel, J.; Illmer-Kephalides, M.; Jaenicke, S.; Straube, J.; May, P.; Goesmann, A.; Kruse, O. New insights into Chlamydomonas reinhardtii hydrogen production processes by combined microarray/RNA-seq transcriptomics. Plant Biotechnol. J. 2013, 11, 717–733. [Google Scholar] [CrossRef]
- Daroch, M.; Geng, S.; Wang, G. Recent advances in liquid biofuel production from algal feedstocks. Appl. Energy 2013, 102, 1371–1381. [Google Scholar] [CrossRef]
- Matos, J.; Cardoso, C.; Gomes, A.; Campos, A.; Falé, P.; Afonso, C.; Bandarra, N.M. Bioprospection of Isochrysis galbana and its potential as a nutraceutical. Food Funct. 2019, 10, 7333–7342. [Google Scholar] [CrossRef]
- Aslam, A.; Bahadar, A.; Liaqat, R.; Saleem, M.; Waqas, A.; Zwawi, M. Algae as an attractive source for cosmetics to counter environmental stress. Sci. Total Environ. 2021, 772, 144905. [Google Scholar] [CrossRef]
- Schuler, L.M.; Gangadhar, K.N.; Duarte, P.; Placines, C.; Molina-Marquez, A.M.; Leon-Banares, R.; Sousa, V.S.; Varela, J.; Barreira, L. Improvement of carotenoid extraction from a recently isolated, robust microalga, Tetraselmis sp. CTP4 (chlorophyta). Bioprocess Biosyst. Eng. 2020, 43, 785–796. [Google Scholar] [CrossRef] [PubMed]
- Borowitzka, M. Commercial-scale production of microalgae for bioproducts. In Blue Biotechnology: Production and Use of Marine Molecules; La Barre, S., Bates, S.S., Eds.; Wiley-VCH: Weinheim, Germany, 2018; pp. 33–65. [Google Scholar] [CrossRef]
- Butler, T.; Golan, Y. Astaxanthin Production from Microalgae. In Microalgae Biotechnology for Food, Health and High Value Products; Springer: Berlin/Heidelberg, Germany, 2020; pp. 175–242. [Google Scholar]
- Laje, K.; Seger, M.; Dungan, B.; Cooke, P.; Polle, J.; Holguin, F.O. Phytoene accumulation in the novel microalga Chlorococcum sp. using the pigment synthesis inhibitor fluridone. Mar. Drugs 2019, 17, 187. [Google Scholar] [CrossRef] [PubMed] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Sector | Stages | ||
|---|---|---|---|
| First Stage | Second Stage | Third Stage | |
| Bioenergy | Bioprospecting of biofuels | Optimization of the production process | Use of biocatalysts, or new routes of biofuels production |
| Health care, therapeutics | Bioprospecting of health care compounds | Protein recombinant technology | Clinical assays for biomedical purposes |
| Environment | Biodegradation and biotransformation assays | Selection of phycoremediation or wastewater treatment process | On-site applications in which an integrated process has been developed |
| Aquaculture | Selection of strains and evaluation for specific fish or crustacean aquaculture | Balanced diets design | Feed additive production and implementation in aquaculture farms |
| Cosmetics | Bioprospecting of natural products from microalgae | Skin applications | Medical care supplements for skin applications |
| Foods | Natural products | Probiotics | Food supplements |
| Characteristics | Chlorophyll | Carotenoids | Phycobilins |
|---|---|---|---|
| Common found place | Green plant and cyanobacteria | Brown algae (phaeophyta) and green algae (chlorophyta) red algae (rhodophyta) | Red algae (rhodophyta) and cryptomonads and cyanobacteria |
| Structural formulae description | Tetrapyrrole ring with a central magnesium atom | Polyene chain consisting of 9–11 double bonds and terminating in rings | Tetrapyrrole unit with open chain of four pyrrole rings (tetrapyrrole) |
| Absorption wavelength | 450–475 nm (blue/blue–green); 615–675 nm (red) | 400–550 nm (blue to green light) | 500 nm to 650 nm (green–red) |
| Water solubility | Insoluble in water | Insoluble in water | Soluble in water |
| Examples | Five types of chlorophylls a, b, c and d | Xanthophylls (molecules containing oxygen) and carotenes (oxygen free molecule). | phycoerythrin, phycocyanin, allophycocyanin |
| Functions | Colour pigment used in photosynthesis as a photoreceptor | Biological abilities, including photosynthesis, photoprotection, plant coloration, and cell signaling. | Supplement the light-capturing ability especially red, orange, yellow, and greenlight. |
| Microalgae | Metabolite Compound | References |
|---|---|---|
| Chlorella zofingiensis | Astaxanthin | [58] |
| Chlorella minutissima | Phytol | [59] |
| Spirulina | Polysaccharides | [60] |
| Spirulina platensis | Phycocyanin, oleic acid, linolenic acid | [61,62] |
| Dunaliella sp. | Diacylglycerol | [63] |
| Nannochloropsis | Diacylglycerol | [64] |
| Dunaliela salina | Palmitic acid | [65] |
| Microalgae Strain | Pretreatment Condition | Bioethanol | References |
|---|---|---|---|
| Chlorella | Dilute acid 5% sulphuric acid | 0.28 g/g biomass | [70] |
| Mixed microalgae | Dilute sulphuric acid 0.5 N H2SO4 at 120 °C for 4 h | 0.18 g/g biomass | [71] |
| Chlorella sp. | Dilute alkaline treatment at 2% NaOH for 2 h at 120 °C | 0.081 g/g biomass | [72] |
| Chlorococcum infusionum | 0.74% NaOH at 120 °C for 30 min | 0.26 g/g biomass | [73] |
| Nannochloropsis gaditana | 1 M NaOH, 120 °C for 30 min | 0.094 g/g biomass | [74] |
| Defatted Nannochloropsis oculata | 4% H2SO4 | 0.062 g/g sugar | [75] |
| Dunaliella tertiolecta | Pretreatment using 1% (v/v) sulfuric acid at 120 °C for 15 min | 0.62 g/g biomass | [76] |
| Microalgal Specie | Biomass Productivity mg/L/d | Carbon Source | Metabolic Cultivation | Lipid Content % | PFA Content mg/g | Reference |
|---|---|---|---|---|---|---|
| C. vulgaris | 137.43 | molasses | Mixotrophic | 39 | 59.7 | [94] |
| C. vulgaris | 91.57 | CO2 | Autotrophic | 19 | 36 | [94] |
| Scenedesmus dimorphus | 119.25 | Sugar cane Bagasse | Mixotrophic | 40.02 | [97] | |
| Scenedesmus dimorphus | 140.37 | Apple-pomace hydrolysate | Mixotrophic | 41 | 64 | [95,96] |
| Scenedesmus dimorphus | 96.4–96.55 | CO2 | Autotrophic | 28 | 37.65 | [95] |
| Chlorella vulgaris | Sorghum bagasse | Mixotrophic | 34.4 | [96] | ||
| Arthrospira platensis | 1330 | Sucrose | Mixotrophic | 3.68 mg/g cell | [98] | |
| Arthrospira platensis | 153 | CO2 | Autotrophic | 3.12 mg/g cell | [98] | |
| Graesiella sp. | 170 | CO2 + Glucose | Mixotrophic | 45.8 | 1.47 | [88] |
| Graesiella sp. | 120 | CO2 | Autotrophic | 19.4 | 16.66 | [88] |
| Dictyosphaerium sp. | 230 | CO2 + Glucose 10 g/L | Mixotrophic | 32 | [99] | |
| Dictyosphaerium sp. | 230 | CO2 + Glucose 20 g/L | Mixotrophic | 42 | [99] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
López-Hernández, J.-F.; Kean-Meng, T.; Asencio-Alcudia, G.-G.; Asyraf-Kassim, M.; Alvarez-González, C.-A.; Márquez-Rocha, F.-J. Sustainable Microalgae and Cyanobacteria Biotechnology. Appl. Sci. 2022, 12, 6887. https://doi.org/10.3390/app12146887
López-Hernández J-F, Kean-Meng T, Asencio-Alcudia G-G, Asyraf-Kassim M, Alvarez-González C-A, Márquez-Rocha F-J. Sustainable Microalgae and Cyanobacteria Biotechnology. Applied Sciences. 2022; 12(14):6887. https://doi.org/10.3390/app12146887
Chicago/Turabian StyleLópez-Hernández, Jenny-Fabiola, Tan Kean-Meng, Gloria-Gertrudys Asencio-Alcudia, Mohd Asyraf-Kassim, Carlos-Alfonso Alvarez-González, and Facundo-Joaquín Márquez-Rocha. 2022. "Sustainable Microalgae and Cyanobacteria Biotechnology" Applied Sciences 12, no. 14: 6887. https://doi.org/10.3390/app12146887
APA StyleLópez-Hernández, J. -F., Kean-Meng, T., Asencio-Alcudia, G. -G., Asyraf-Kassim, M., Alvarez-González, C. -A., & Márquez-Rocha, F. -J. (2022). Sustainable Microalgae and Cyanobacteria Biotechnology. Applied Sciences, 12(14), 6887. https://doi.org/10.3390/app12146887