

In Silico Modeling and Quantification of Synergistic Effects of Multi-Combination Compounds: Case Study of the Attenuation of Joint Pain Using a Combination of Phytonutrients

Abstract
:1. Introduction
Research Aim
2. Materials and Methods
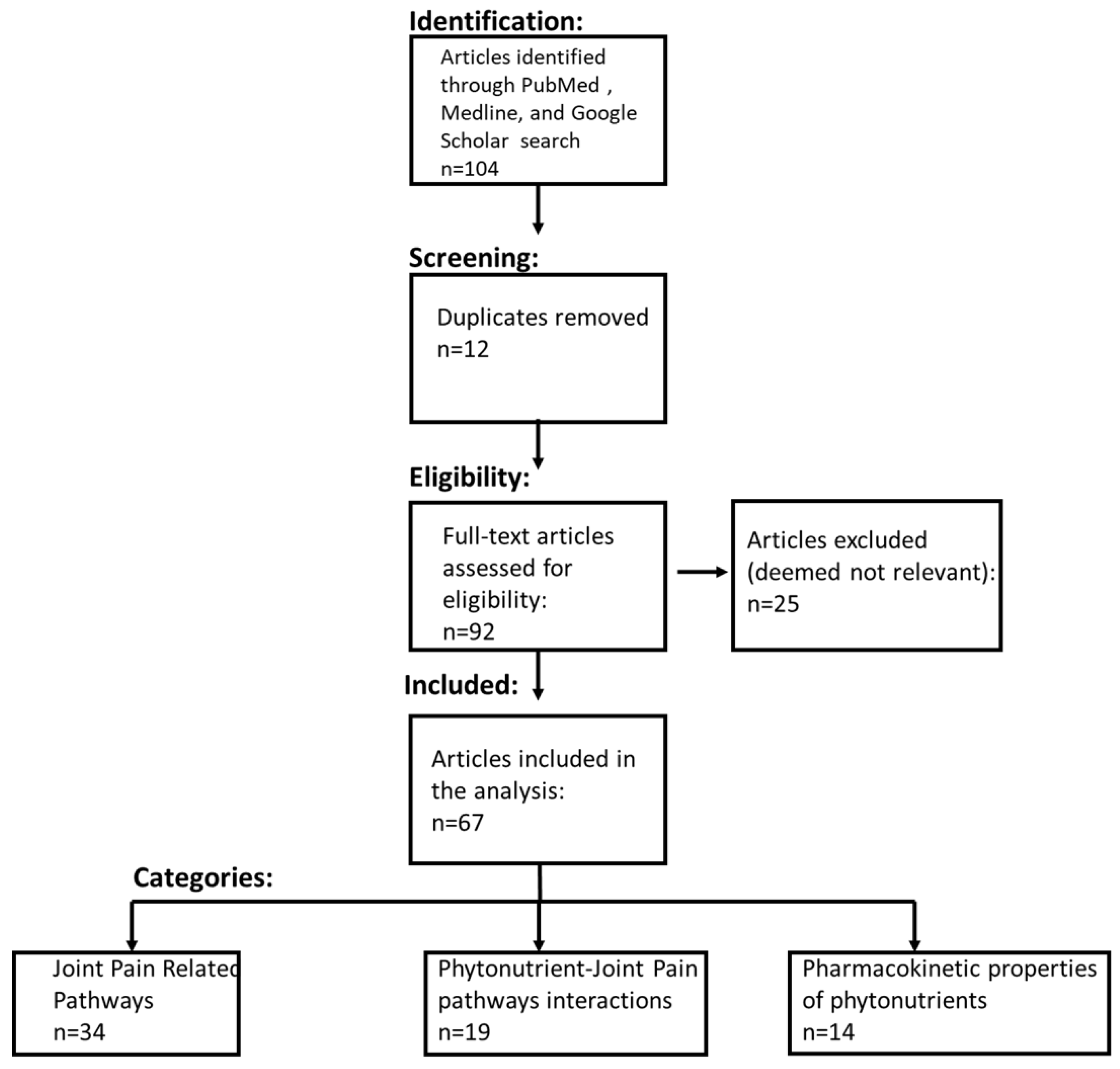
2.1. Systematic Literature Review Process and Inclusion Criteria
2.2. CytoSolve In Silico Modeling Protocol
2.2.1. Control Conditions
2.2.2. Computer Simulations to Study the Effect of Apigenin and Hesperidin on Integrated Model of Joint Pain
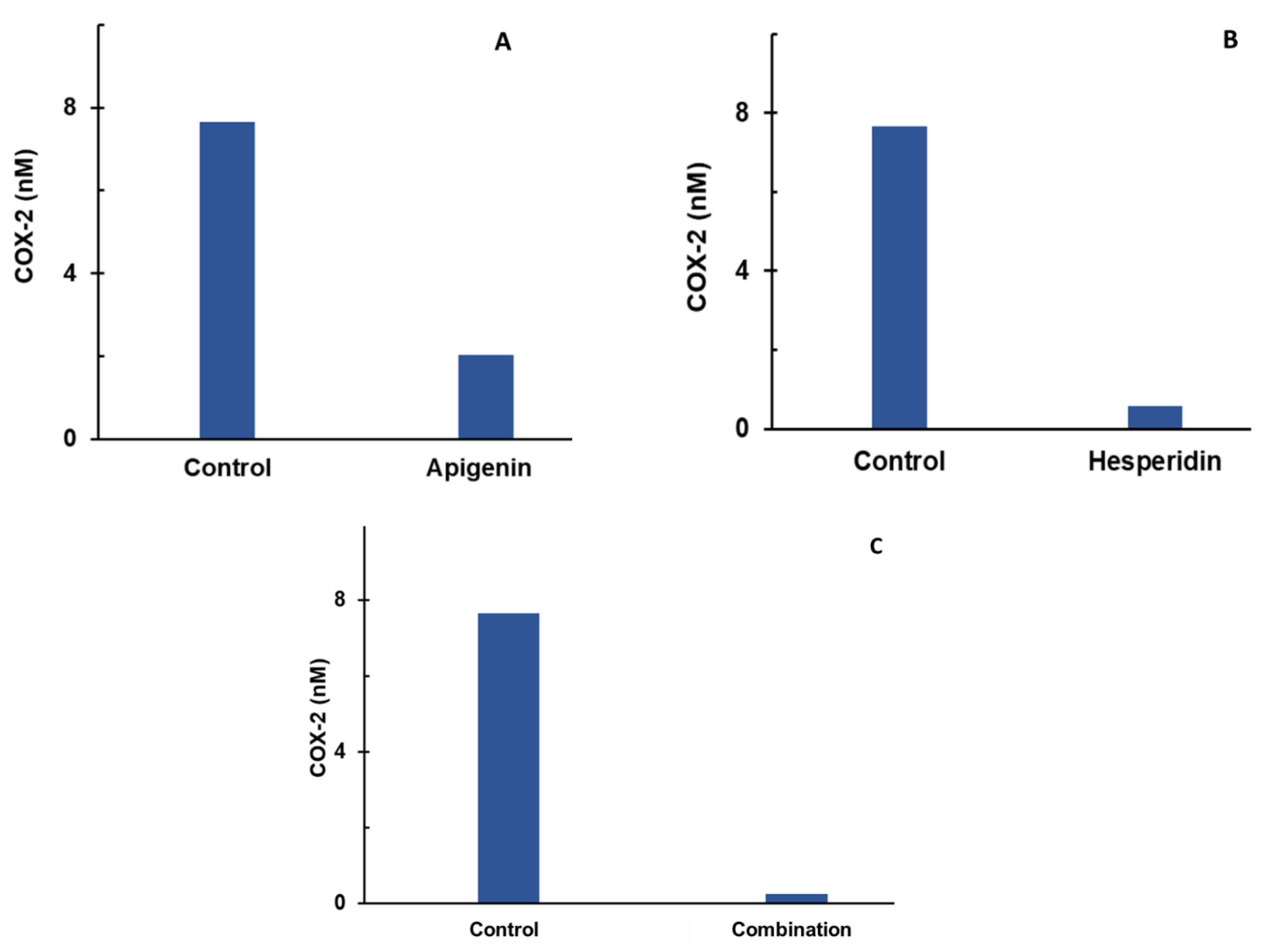
- Individual and combination effects of apigenin and hesperidin on COX-2 production;
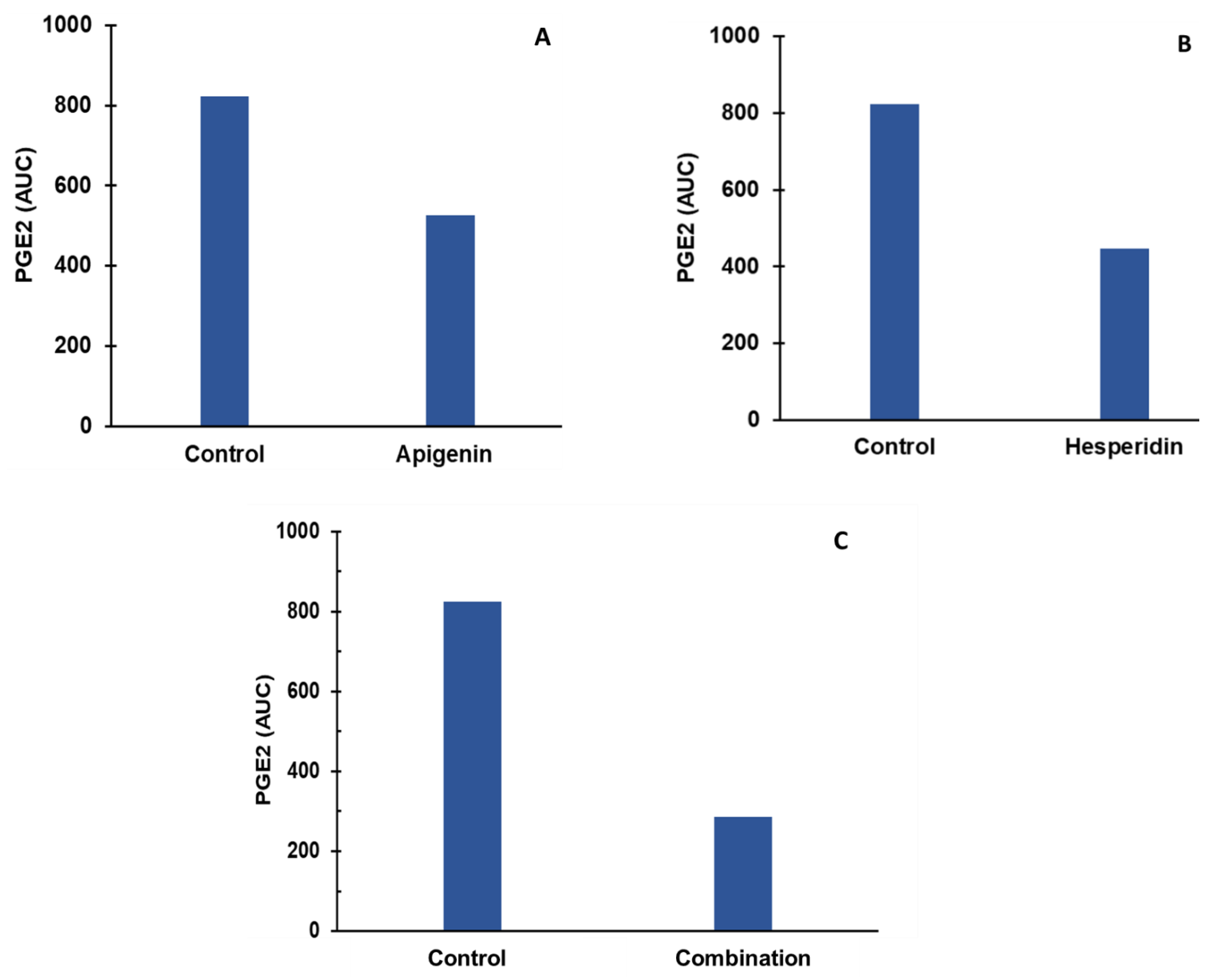
- Individual and combination effects of apigenin and hesperidin on PGE-2 production;
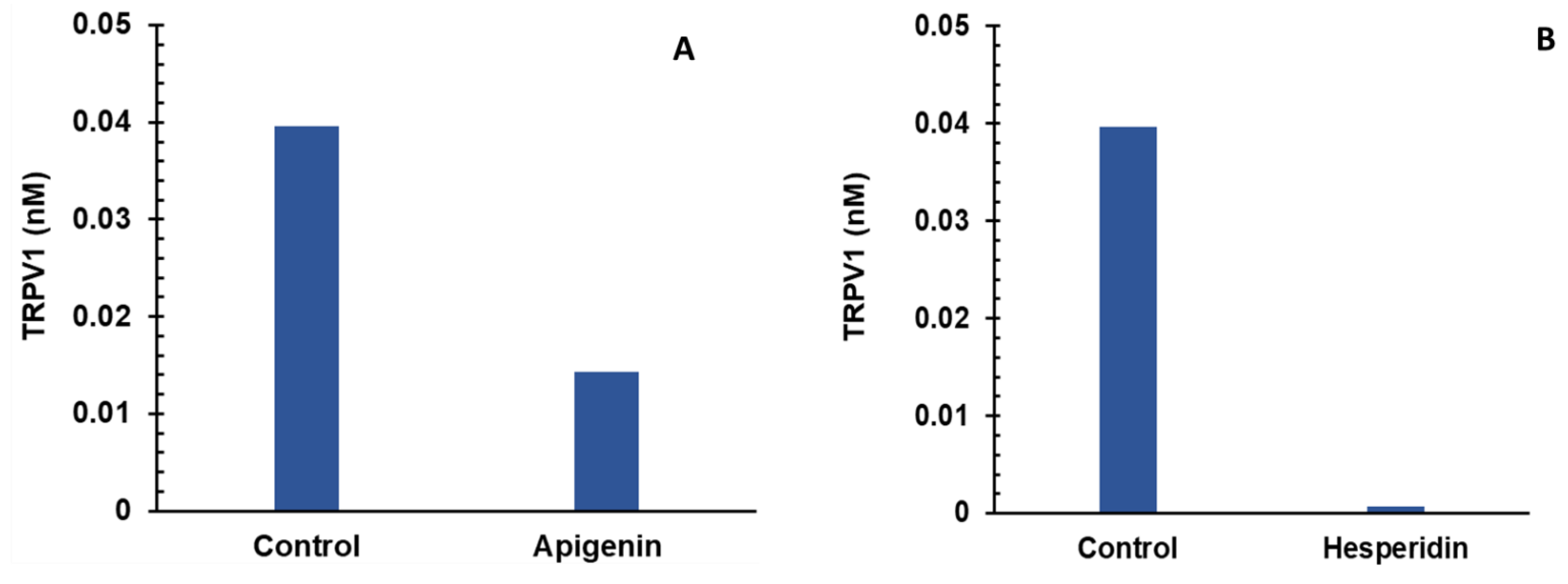
- Individual and combination effects of apigenin and hesperidin on TRPV1 production;
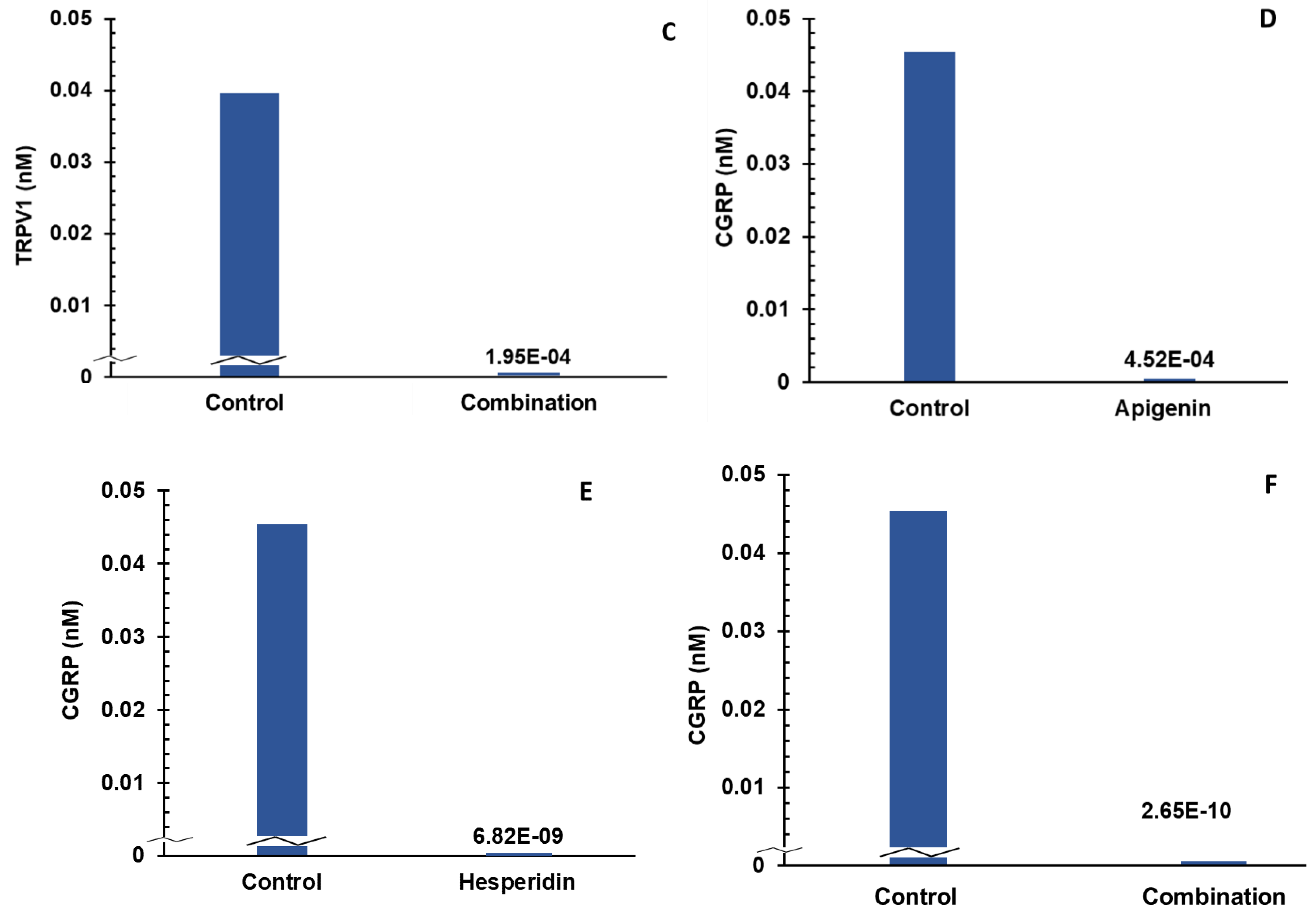
- Individual and combination effects of apigenin and hesperidin on CGRP production;
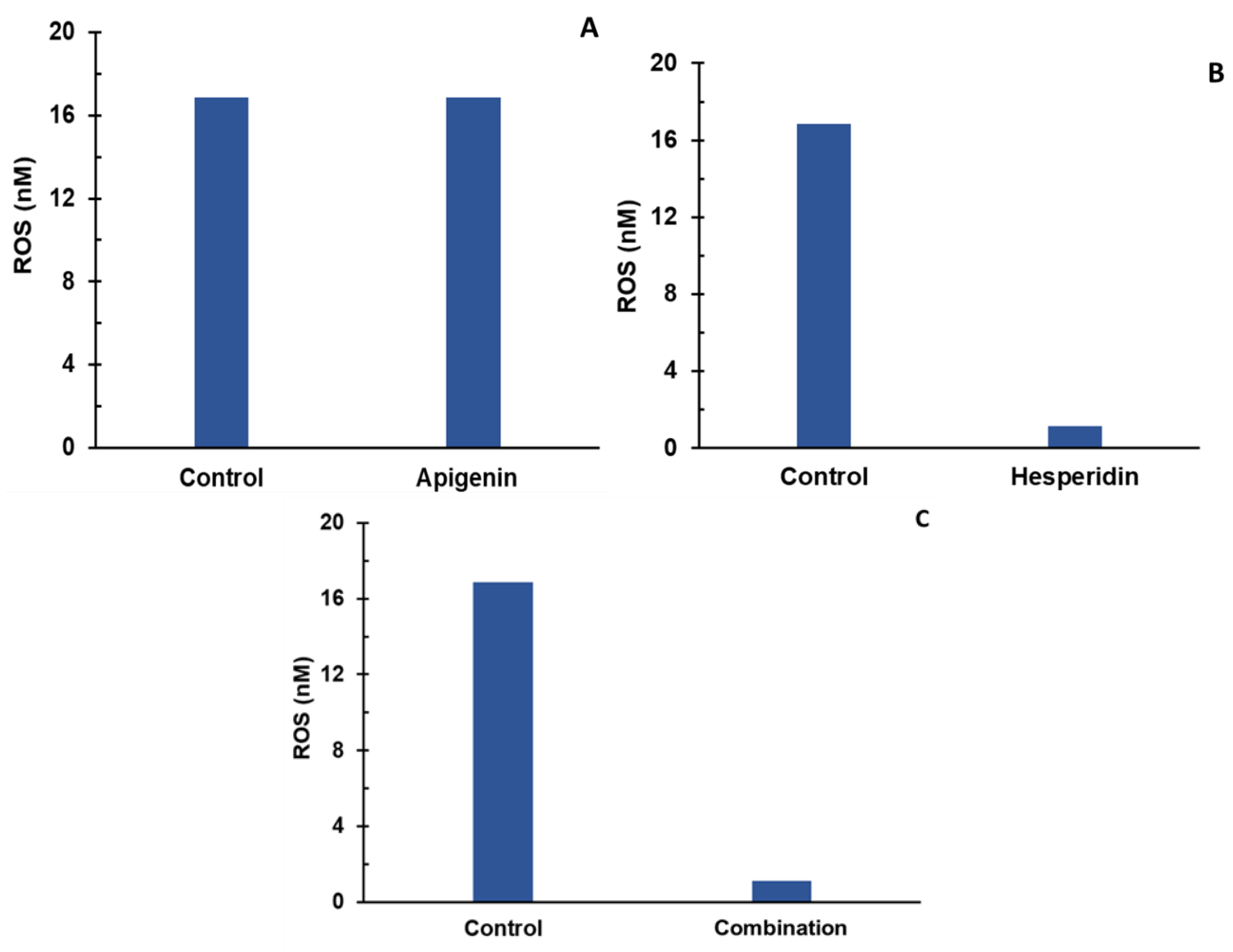
- Individual and combination effects of apigenin and hesperidin on ROS production.
2.2.3. Computer Simulations to Determine Synergistic Effects of Apigenin and Hesperidin on an Integrated Model of Joint Pain
- Sequential addition of apigenin first and then hesperidin to estimate COX-2 production;
- Sequential addition of hesperidin first and then apigenin to estimate COX-2 production;
- Sequential addition of apigenin first and then hesperidin to estimate PGE2 production;
- Sequential addition of hesperidin first and then apigenin to PGE2 production;
- Sequential addition of apigenin first and then hesperidin to estimate TRPV1 production;
- Sequential addition of hesperidin first and then apigenin to estimate TRPV1 production;
- Sequential addition of apigenin first and then hesperidin to estimate CGRP production;
- Sequential addition of hesperidin first and then apigenin to estimate CGRP production;
- Sequential addition of apigenin first and then hesperidin to estimate ROS production;
- Sequential addition of hesperidin first and then apigenin to estimate ROS production.
3. Results
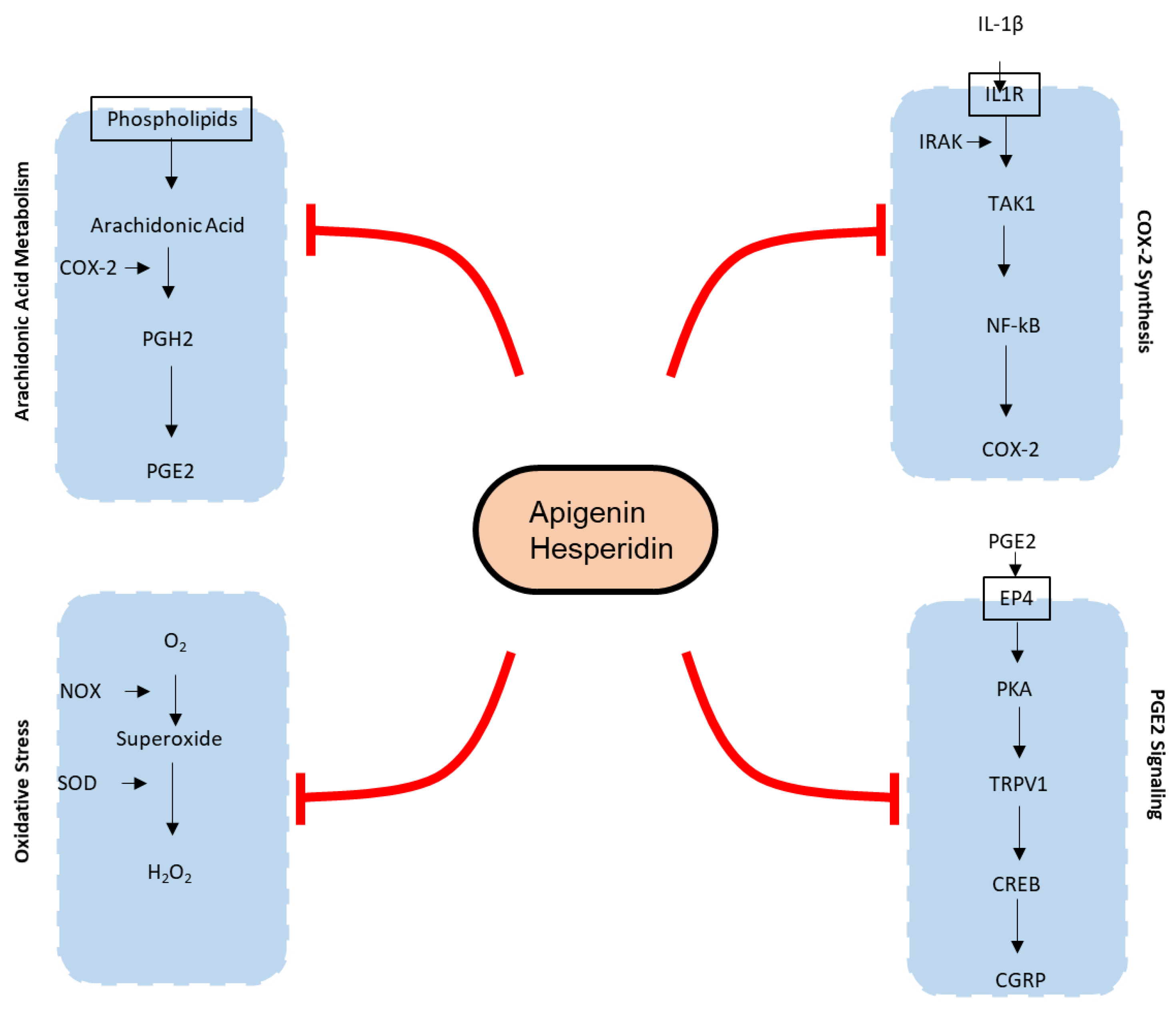
3.1. Effect of Apigenin and Hesperidin on the COX-2 Synthesis Pathway
3.2. Effect of Apigenin and Hesperidin on Arachidonic Acid Metabolism Pathways
3.3. Effect of Apigenin and Hesperidin on PGE2 Signaling Pathways
3.4. Effect of Apigenin and Hesperidin on Oxidative Stress Signaling Pathway
4. Discussion
5. Conclusions and Future Work
5.1. Conclusions
5.2. Future Work
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Kidd, B.L.; Langford, R.M.; Wodehouse, T. Arthritis and pain. Current approaches in the treatment of arthritic pain. Arthritis Res. Ther. 2007, 9, 214. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hurley, M.; Sheldon, H.; Connolly, M.; Carter, A.; Hallett, R. Providing easier access to community-based healthcare for people with joint pain: Experiences of delivering ESCAPE-pain in community venues by exercise professionals. Musculoskelet. Care 2022, 20, 408–415. [Google Scholar] [CrossRef] [PubMed]
- Yoon, E.; Doherty, J.B. Arthritis pain. J. Gerontol. Soc. Work 2008, 50 (Suppl. S1), 79–103. [Google Scholar] [CrossRef]
- Lee, A.S.; Ellman, M.B.; Yan, D.; Kroin, J.S.; Cole, B.J.; van Wijnen, A.J.; Im, H.J. A Current Review of Molecular Mechanisms Regarding Osteoarthritis and Pain. Gene 2013, 527, 440. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, X.; Kim, J.S.; Van Wijnen, A.J.; Im, H.J. Osteoarthritic Tissues Modulate Functional Properties of Sensory Neurons Associated with Symptomatic OA Pain. Mol. Biol. Rep. 2011, 38, 5335. [Google Scholar] [CrossRef]
- Felson, D.T. The sources of pain in knee osteoarthritis. Curr. Opin. Rheumatol. 2005, 17, 624–628. [Google Scholar] [CrossRef] [PubMed]
- Dray, A.; Read, S.J. Arthritis and pain. Future targets to control osteoarthritis pain. Arthritis Res. Ther. 2007, 9, 212. [Google Scholar] [CrossRef] [Green Version]
- Cho, H.; Walker, A.; Williams, J.; Hasty, K.A.; Shuang, Y. Study of Osteoarthritis Treatment with Anti-Inflammatory Drugs: Cyclooxygenase-2 Inhibitor and Steroids. Biomed Res. Int. 2015, 2015, 595273. [Google Scholar] [CrossRef] [Green Version]
- Jang, Y.; Kim, M.; Hwang, S.W. Molecular mechanisms underlying the actions of arachidonic acid-derived prostaglandins on peripheral nociception. J. Neuroinflamm. 2020, 17, 30. [Google Scholar] [CrossRef]
- Jenkins, D.W.; Feniuk, W.; Humphrey, P.P.A. Characterization of the prostanoid receptor types involved in mediating calcitonin gene-related peptide release from cultured rat trigeminal neurones. Br. J. Pharmacol. 2001, 134, 1296–1302. [Google Scholar] [CrossRef]
- Moriyama, T.; Higashi, T.; Togashi, K.; Iida, T.; Segi, E.; Sugimoto, Y.; Tominaga, T.; Narumiya, S.; Tominaga, M. Sensitization of TRPV1 by EP1 and IP reveals peripheral nociceptive mechanism of prostaglandins. Mol. Pain 2005, 1, 3. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Al-Lazikani, B.; Banerji, U.; Workman, P. Combinatorial drug therapy for cancer in the post-genomic era. Nat. Biotechnol. 2012, 30, 679–692. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.K.; Park, S.U. Recent insights into the biological functions of apigenin. EXCLI J. 2020, 19, 984. [Google Scholar]
- Hong, S.; Dia, V.P.; Baek, S.J.; Zhong, Q. Nanoencapsulation of apigenin with whey protein isolate: Physicochemical properties, in vitro activity against colorectal cancer cells, and bioavailability. LWT 2022, 154, 112751. [Google Scholar] [CrossRef] [PubMed]
- Abid, R.; Ghazanfar, S.; Farid, A.; Sulaman, S.M.; Idrees, M.; Amen, R.A.; Muzammal, M.; Shahzad, M.K.; Mohamed, M.O.; Khaled, A.A.; et al. Pharmacological Properties of 4′, 5, 7-Trihydroxyflavone (Apigenin) and Its Impact on Cell Signaling Pathways. Molecules 2022, 27, 4304. [Google Scholar] [CrossRef]
- Pyrzynska, K. Hesperidin: A Review on Extraction Methods, Stability and Biological Activities. Nutrients 2022, 14, 2387. [Google Scholar] [CrossRef]
- Ayyadurai, V.A.S.; Deonikar, P. Bioactive compounds in green tea may improve transplant tolerance: A computational systems biology analysis. Clin. Nutr. ESPEN 2021, 46, 439–452. [Google Scholar] [CrossRef]
- Ayyadurai, V.A.S.; Deonikar, P.; Bannuru, R.R. Attenuation of low-grade chronic inflammation by phytonutrients: A computational systems biology analysis. Clin. Nutr. ESPEN 2022, 49, 425–435. [Google Scholar] [CrossRef]
- Moher, D.; Liberati, A.; Tetzlaff, J.; Altman, D.G.; Altman, D.; Antes, G.; Atkins, D.; Barbour, V.; Barrowman, N.; Berlin, J.A.; et al. Preferred reporting items for systematic reviews and meta-analyses: The PRISMA statement. PLoS Med. 2009, 6, e1000097. [Google Scholar] [CrossRef] [Green Version]
- Imamura, M.; Targino, R.A.; Hsing, W.T.; Imamura, S.; Azevedo, R.S.; Villas Boas, L.S.; Tozetto-Mendoza, T.R.; Alfieri, F.M.; Filippo, T.R.; Battistella, L.R. Concentration of cytokines in patients with osteoarthritis of the knee and fibromyalgia. Clin. Interv. Aging 2014, 9, 939. [Google Scholar]
- King, S.S.; Abughazaleh, A.A.; Webel, S.K.; Jones, K.L. Circulating fatty acid profiles in response to three levels of dietary omega-3 fatty acid supplementation in horses. J. Anim. Sci. 2008, 86, 1114–1123. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, K.; Ma, W.; Liang, H.; Ouyang, Q.; Tang, C.; Lai, L. Dynamic Simulations on the Arachidonic Acid Metabolic Network. PLoS Comput. Biol. 2007, 3, 523–530. [Google Scholar] [CrossRef] [PubMed]
- Jansen, F.; Yang, X.; Franklin, B.S.; Hoelscher, M.; Schmitz, T.; Bedorf, J.; Nickenig, G.; Werner, N. High glucose condition increases NADPH oxidase activity in endothelial microparticles that promote vascular inflammation. Cardiovasc. Res. 2013, 98, 94–106. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yuan, S.; Chen, H. Mathematical rules for synergistic, additive, and antagonistic effects of multi-drug combinations and their application in research and development of combinatorial drugs and special medical food combinations. Food Sci. Hum. Wellness 2019, 8, 136–141. [Google Scholar] [CrossRef]
- Walsh, D.A.; McWilliams, D.F. Pain in rheumatoid arthritis. Curr. Pain Headache Rep. 2012, 16, 509–517. [Google Scholar] [CrossRef]
- O’Neill, T.W.; Felson, D.T. Mechanisms of Osteoarthritis (OA) Pain. Curr. Osteoporos. Rep. 2018, 16, 611. [Google Scholar] [CrossRef] [Green Version]
- Zhaoyang, R.; Martire, L.M. Daily Sedentary Behavior Predicts Pain and Affect in Knee Arthritis. Ann. Behav. Med. 2019, 53, 642–651. [Google Scholar] [CrossRef]
- McAlindon, T.E.; Bannuru, R.R.; Sullivan, M.C.; Arden, N.K.; Berenbaum, F.; Bierma-Zeinstra, S.M.; Hawker, G.A.; Henrotin, Y.; Hunter, D.J.; Kawaguchi, H.; et al. OARSI guidelines for the non-surgical management of knee osteoarthritis. Osteoarthr. Cartil. 2014, 22, 363–388. [Google Scholar] [CrossRef] [Green Version]
- Bindu, S.; Mazumder, S.; Bandyopadhyay, U. Non-steroidal anti-inflammatory drugs (NSAIDs) and organ damage: A current perspective. Biochem. Pharmacol. 2020, 180, 114147. [Google Scholar] [CrossRef]
- Bañuls-Mirete, M.; Ogdie, A.; Guma, M. Micronutrients: Essential Treatment for Inflammatory Arthritis? Curr. Rheumatol. Rep. 2020, 22, 87. [Google Scholar] [CrossRef]
- Badimon, L.; Vilahur, G.; Padro, T. Systems biology approaches to understand the effects of nutrition and promote health. Br. J. Clin. Pharmacol. 2017, 83, 38–45. [Google Scholar] [CrossRef] [Green Version]
- Johnson, M.; McElhenney, W.H.; Egnin, M. Influence of Green Leafy Vegetables in Diets with an Elevated ω-6:ω-3 Fatty Acid Ratio on Rat Blood Pressure, Plasma Lipids, Antioxidant Status and Markers of Inflammation. Nutrients 2019, 11, 301. [Google Scholar] [CrossRef] [PubMed]
- Rondanelli, M.; Faliva, M.A.; Miccono, A.; Naso, M.; Nichetti, M.; Riva, A.; Guerriero, F.; De Gregori, M.; Peroni, G.; Perna, S. Food pyramid for subjects with chronic pain: Foods and dietary constituents as anti-inflammatory and antioxidant agents. Nutr. Res. Rev. 2018, 31, 131–151. [Google Scholar] [CrossRef] [PubMed]
- Ayyadurai, V.A.S.; Dewey, C.F. CytoSolve: A Scalable Computational Method for Dynamic Integration of Multiple Molecular Pathway Models. Cell. Mol. Bioeng. 2011, 4, 28–45. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nordsletten, D.A.; Yankama, B.; Umeton, R.; Ayyadurai, V.V.S.; Dewey, C.F. Multiscale Mathematical Modeling to Support Drug Development. IEEE Trans. Biomed. Eng. 2011, 58, 3508–3512. [Google Scholar] [CrossRef]
- Sweeney, M.D.; Ayyadurai, S.; Zlokovic, B.V. Pericytes of the Neurovascular Unit: Key Functions and Signaling Pathways. Nat. Neurosci. 2016, 19, 771–783. [Google Scholar] [CrossRef]
- Koo, A.; Nordsletten, D.; Umeton, R.; Yankama, B.; Ayyadurai, S.; García-Cardeña, G.; Dewey, C.F. In Silico Modeling of Shear-Stress-Induced Nitric Oxide Production in Endothelial Cells through Systems Biology. Biophys. J. 2013, 104, 2295–2306. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- CytoSolve, Inc. Food and Drug Administration Center for Drug Evaluation and Research Request for Determination of Exempt Status of Investigational New Drug Application (IND) for Cyto-001 as Treatment for Patients with Pancreatic Cancer (PIND: 118833); CytoSolve, Inc.: Beltsville, MD, USA, 2013. [Google Scholar]
- Ayyadurai, V.A.S.; Deonikar, P.; McLure, K.G.; Sakamoto, K.M. Molecular Systems Architecture of Interactome in the Acute Myeloid Leukemia Microenvironment. Cancers 2022, 14, 756. [Google Scholar] [CrossRef] [PubMed]
- Ayyadurai, V.A.S.; Deonikar, P. Do GMOs Accumulate Formaldehyde and Disrupt Molecular Systems Equilibria? Systems Biology May Provide Answers. Agric. Sci. 2015, 6, 630–662. [Google Scholar] [CrossRef] [Green Version]
- Kothandaram, S.; Deonikar, P.; Mohan, M.; Venugopal, V.; Ayyadurai, V.A.S. In Silico Modeling of C1 Metabolism. Am. J. Plant Sci. 2015, 6, 1444–1465. [Google Scholar] [CrossRef] [Green Version]
- Mohan, M.; Kothandaram, S.; Venugopal, V.; Deonikar, P.; Ayyadurai, V.A.S. Integrative Modeling of Oxidative Stress and C1 Metabolism Reveals Upregulation of Formaldehyde and Downregulation of Glutathione. Am. J. Plant Sci. 2015, 6, 1527–1542. [Google Scholar] [CrossRef] [Green Version]
- Ayyadurai, V.A.S.; Hansen, M.; Fagan, J.; Deonikar, P. In-Silico Analysis & In-Vivo Results Concur on Glutathione Depletion in Glyphosate Resistant GMO Soy, Advancing a Systems Biology Framework for Safety Assessment of GMOs. Am. J. Plant Sci. 2016, 7, 1571–1589. [Google Scholar] [CrossRef]
- Cornish-Bowden, A. One Hundred Years of Michaelis–Menten Kinetics. Perspect. Sci. 2015, 4, 3–9. [Google Scholar] [CrossRef]
- Michaelis, L.; Menten, M.L.; Goody, R.S.; Johnson, K.A. Die Kinetik Der Invertinwirkung/ The Kinetics of Invertase Action. Biochemistry 1913, 49, 352. [Google Scholar]
- Hucka, M.; Finney, A.; Sauro, H.M.; Bolouri, H.; Doyle, J.C.; Kitano, H.; Arkin, A.P.; Bornstein, B.J.; Bray, D.; Cornish-Bowden, A.; et al. The Systems Biology Markup Language (SBML): A Medium for Representation and Exchange of Biochemical Network Models. Bioinformatics 2003, 19, 524–531. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ayyadurai, V.A.S. Services-Based Systems Architecture for Modeling the Whole Cell: A Distributed Collaborative Engineering Systems Approach. Commun. Med. Care Compunetics 2010, 1, 115–168. [Google Scholar]
- Shmulevich, I.; Aitchison, J.D. DETERMINISTIC AND STOCHASTIC MODELS OF GENETIC REGULATORY NETWORKS. Methods Enzymol. 2009, 467, 335. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oden, J.T.; John, T.; Reddy, J.N.; Junuthula, N. An Introduction to the Mathematical Theory of Finite Elements, 1976th ed.; Dover Publications: New York, NY, USA, 2011; ISBN 9780486142210. [Google Scholar]
- Cumming, G.; Fidler, F.; Vaux, D.L. Error Bars in Experimental Biology. J. Cell Biol. 2007, 177, 7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thanh, V.H.; Zunino, R.; Priami, C. Efficient Finite-Difference Method for Computing Sensitivities of Biochemical Reactions. Proc. R. Soc. A Math. Phys. Eng. Sci. 2018, 474. [Google Scholar] [CrossRef] [Green Version]
- Kablar, N.A.; Kvrgi, V. Mathematical Model of IL − 1 — N F ΚB Biological Module. J. Theor. Biol. 2012, 1, 31–37. [Google Scholar]
- Hoffmann, A.; Levchenko, A.; Scott, M.L.; Baltimore, D. The IκB-NF-ΚB Signaling Module: Temporal Control and Selective Gene Activation. Science 2002. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boerboom, D.; Brown, K.A.; Vaillancourt, D.; Poitras, P.; Goff, A.K.; Watanabe, K.; Doré, M.; Sirois, J. Expression of Key Prostaglandin Synthases in Equine Endometrium During Late Diestrus and Early Pregnancy. Biol. Reprod. 2004, 70, 391–399. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Learn, C.A.; Mizel, S.B.; McCall, C.E. MRNA and Protein Stability Regulate the Differential Expression of Pro- and Anti-Inflammatory Genes in Endotoxin-Tolerant THP-1 Cells. J. Biol. Chem. 2000, 275, 12185–12193. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ghorbani, A.; Nazari, M.; Jeddi-Tehrani, M.; Zand, H. The Citrus Flavonoid Hesperidin Induces P53 and Inhibits NF-ΚB Activation in Order to Trigger Apoptosis in NALM-6 Cells: Involvement of PPARγ-Dependent Mechanism. Eur. J. Nutr. 2012, 51, 39–46. [Google Scholar] [CrossRef] [PubMed]
- Shukla, S.; Kanwal, R.; Shankar, E.; Datt, M.; Chance, M.R.; Fu, P.; MacLennan, G.T.; Gupta, S. Apigenin Blocks IKKα Activation and Suppresses Prostate Cancer Progression. Oncotarget 2015, 6, 31216. [Google Scholar] [CrossRef] [PubMed]
- Alcaraz, M.J.; Ferrándiz, M.L. Modification of Arachidonic Metabolism by Flavonoids. J. Ethnopharmacol. 1987, 21, 209–229. [Google Scholar] [CrossRef]
- Franke, R.; Schilcher, H. Chamomile Indrsurtia Profiles, 1st ed.; CRC Press: Boca Raton, FA, USA; p. 304. ISBN 9780429215674.
- McLaughlin, S.; Wang, J.; Gambhir, A.; Murray, D. PIP(2) and Proteins: Interactions, Organization, and Information Flow. Annu. Rev. Biophys. Biomol. Struct. 2002, 31, 151–175. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Willoughby, D.; Cooper, D.M.F. Organization and Ca2+ Regulation of Adenylyl Cyclases in CAMP Microdomains. Physiol. Rev. 2007, 87, 965–1010. [Google Scholar] [CrossRef] [Green Version]
- Kawaguchi, S.Y.; Hirano, T. Gating of Long-Term Depression by Ca2+/Calmodulin-Dependent Protein Kinase II through Enhanced CGMP Signalling in Cerebellar Purkinje Cells. J. Physiol. 2013, 591, 1707–1730. [Google Scholar] [CrossRef]
- Wei, X.; Zhang, X.; Zuscik, M.J.; Drissi, M.H.; Schwarz, E.M.; O’Keefe, R.J. Fibroblasts Express RANKL and Support Osteoclastogenesis in a COX-2-Dependent Manner after Stimulation with Titanium Particles. J. Bone Miner. Res. 2005, 20, 1136–1148. [Google Scholar] [CrossRef]
- Calebiro, D.; Nikolaev, V.O.; Gagliani, M.C.; De Filippis, T.; Dees, C.; Tacchetti, C.; Persani, L.; Lohse, M.J. Persistent CAMP-Signals Triggered by Internalized G-Protein-Coupled Receptors. PLoS Biol. 2009, 7. [Google Scholar] [CrossRef] [Green Version]
- Markevich, N.I.; Hoek, J.B.; Kholodenko, B.N. Signaling Switches and Bistability Arising from Multisite Phosphorylation in Protein Kinase Cascades. J. Cell Biol. 2004, 164, 353–359. [Google Scholar] [CrossRef] [PubMed]
- Salonikidis, P.S.; Zeug, A.; Kobe, F.; Ponimaskin, E.; Richter, D.W. Quantitative Measurement of CAMP Concentration Using an Exchange Protein Directly Activated by a CAMP-Based FRET-Sensor. Biophys. J. 2008, 95, 5412–5423. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Biggin, M.D. Animal Transcription Networks as Highly Connected, Quantitative Continua. Dev. Cell 2011, 21, 611–626. [Google Scholar] [CrossRef] [Green Version]
- Dolan, A.T.; Diamond, S.L. Systems Modeling of Ca(2+) Homeostasis and Mobilization in Platelets Mediated by IP3 and Store-Operated Ca(2+) Entry. Biophys. J. 2014, 106, 2049–2060. [Google Scholar] [CrossRef]
- Greget, R.; Pernot, F.; Bouteiller, J.M.C.; Ghaderi, V.; Allam, S.; Keller, A.F.; Ambert, N.; Legendre, A.; Sarmis, M.; Haeberle, O.; et al. Simulation of Postsynaptic Glutamate Receptors Reveals Critical Features of Glutamatergic Transmission. PLoS ONE 2011, 6, e28380. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chang, C.W.; Poteet, E.; Schetz, J.A.; Gümüş, Z.H.; Weinstein, H. Towards a Quantitative Representation of the Cell Signaling Mechanisms of Hallucinogens: Measurement and Mathematical Modeling of 5-HT1A and 5-HT2A Receptor-Mediated ERK1/2 Activation. Neuropharmacology 2009, 56 (Suppl. S1), 213–225. [Google Scholar] [CrossRef] [Green Version]
- Wu, G.; Lü, J.M.; Van Der Donk, W.A.; Kulmacz, R.J.; Tsai, A.L. Cyclooxygenase Reaction Mechanism of Prostaglandin H Synthase from Deuterium Kinetic Isotope Effects. J. Inorg. Biochem. 2011, 105, 382–390. [Google Scholar] [CrossRef] [Green Version]
- Buschow, R. The Heterogeneity of TRPV1 and Its Activation in Nociceptive Neurons. Ph.D. Thesis, Freien Universität Berlin, Berlin, Germany, 2014. [Google Scholar]
- Zuo, P.; Picher, M.; Okada, S.F.; Lazarowski, E.R.; Button, B.; Boucher, R.C.; Elston, T.C. Mathematical Model of Nucleotide Regulation on Airway Epithelia: IMPLICATIONS FOR AIRWAY HOMEOSTASIS*. J. Biol. Chem. 2008, 283, 26805. [Google Scholar] [CrossRef] [Green Version]
- Marshall, F.H.; Patel, K.; Lundstrom, K.; Camacho, J.; Foord, S.M.; Lee, M.G. Characterization of [3H]-Prostaglandin E2 Binding to Prostaglandin EP4 Receptors Expressed with Semliki Forest Virus. Br. J. Pharmacol. 1997, 121, 1673–1678. [Google Scholar] [CrossRef] [Green Version]
- Ponsioen, B.; Zhao, J.; Riedl, J.; Zwartkruis, F.; van der Krogt, G.; Zaccolo, M.; Moolenaar, W.H.; Bos, J.L.; Jalink, K. Detecting CAMP-Induced Epac Activation by Fluorescence Resonance Energy Transfer: Epac as a Novel CAMP Indicator. EMBO Rep. 2004, 5, 1176–1180. [Google Scholar] [CrossRef] [Green Version]
- Ogawa, N.; Kurokawa, T.; Fujiwara, K.; Polat, O.K.; Badr, H.; Takahashi, N.; Mori, Y. Functional and Structural Divergence in Human TRPV1 Channel Subunits by Oxidative Cysteine Modification. J. Biol. Chem. 2016, 291, 4197–4210. [Google Scholar] [CrossRef] [Green Version]
- Long, X.; Fan, M.; Bigsby, R.M.; Nephew, K.P. Apigenin Inhibits Antiestrogen-Resistant Breast Cancer Cell Growth through Estrogen Receptor-Alpha-Dependent and Estrogen Receptor-Alpha-Independent Mechanisms. Mol. Cancer Ther. 2008, 7, 2096–2108. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, J.K. Suppression of Protein Kinase C and Nuclear Oncogene Expression as Possible Action Mechanisms of Cancer Chemoprevention by Curcumin. Arch. Pharm. Res. 2004, 27, 683–692. [Google Scholar] [CrossRef]
- Jin, Y.R.; Han, X.H.; Zhang, Y.H.; Lee, J.J.; Lim, Y.; Chung, J.H.; Yun, Y.P. Antiplatelet Activity of Hesperetin, a Bioflavonoid, Is Mainly Mediated by Inhibition of PLC-Gamma2 Phosphorylation and Cyclooxygenase-1 Activity. Atherosclerosis 2007, 194, 144–152. [Google Scholar] [CrossRef] [PubMed]
- Boonpawa, R.; Spenkelink, A.; Punt, A.; Rietjens, I.M.C.M. Physiologically Based Kinetic Modeling of Hesperidin Metabolism and Its Use to Predict in Vivo Effective Doses in Humans. Mol. Nutr. Food Res. 2017, 61. [Google Scholar] [CrossRef] [Green Version]
- Martínez, I.; Saracho, R.; Moina, I.; Montenegro, J.; Llach, F. Is There a Lesser Hyperparathyroidism in Diabetic Patients with Chronic Renal Failure? Nephrol. Dial. Transplant. 1998, 13, 9–11. [Google Scholar] [CrossRef] [Green Version]
- Rehmann, H. Epac-Inhibitors: Facts and Artefacts. Sci. Rep. 2013, 3. [Google Scholar] [CrossRef] [Green Version]
- Enslen, H.; Tokumitsu, H.; Soderling, T.R. Phosphorylation of CREB by CaM-Kinase IV Activated by CaM-Kinase IV Kinase. Biochem. Biophys. Res. Commun. 1995, 207, 1038–1043. [Google Scholar] [CrossRef] [PubMed]
- Tomaiuolo, M.; Kottke, M.; Matheny, R.W.; Reifman, J.; Mitrophanov, A.Y. Computational Identification and Analysis of Signaling Subnetworks with Distinct Functional Roles in the Regulation of TNF Production. Mol. Biosyst. 2016, 12, 826–838. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Neeb, L.; Hellen, P.; Boehnke, C.; Hoffmann, J.; Schuh-Hofer, S.; Dirnagl, U.; Reuter, U. IL-1β Stimulates COX-2 Dependent PGE2 Synthesis and CGRP Release in Rat Trigeminal Ganglia Cells. PLoS ONE 2011, 6. [Google Scholar] [CrossRef] [PubMed]
- Ma, W. Chronic Prostaglandin E2 Treatment Induces the Synthesis of the Pain-Related Peptide Substance P and Calcitonin Gene-Related Peptide in Cultured Sensory Ganglion Explants. J. Neurochem. 2010, 115, 363–372. [Google Scholar] [CrossRef] [PubMed]
- Shimozawa, N.; Okajima, K.; Harada, N. Estrogen and Isoflavone Attenuate Stress-Induced Gastric Mucosal Injury by Inhibiting Decreases in Gastric Tissue Levels of CGRP in Ovariectomized Rats. Am. J. Physiol. Gastrointest. Liver Physiol. 2007, 292. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dargemont, C.; Kühn, L.C. Export of MRNA from Microinjected Nuclei of Xenopus Laevis Oocytes. J. Cell Biol. 1992, 118, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Deisseroth, K.; Heist, E.K.; Tsien, R.W. Translocation of Calmodulin to the Nucleus Supports CREB Phosphorylation in Hippocampal Neurons. Nature 1998, 392, 198–202. [Google Scholar] [CrossRef] [PubMed]
- Falcke, M.; Huerta, R.; Rabinovich, M.I.; Abarbanel, H.D.I.; Elson, R.C.; Selverston, A.I. Modeling Observed Chaotic Oscillations in Bursting Neurons: The Role of Calcium Dynamics and IP3. Biol. Cybern. 2000, 82, 517–527. [Google Scholar] [CrossRef]
- Dupont, G.; Houart, G.; De Koninck, P. Sensitivity of CaM Kinase II to the Frequency of Ca2+ Oscillations: A Simple Model. Cell Calcium 2003, 34, 485–497. [Google Scholar] [CrossRef]
- Saucerman, J.J.; Bers, D.M. Calmodulin Mediates Differential Sensitivity of CaMKII and Calcineurin to Local Ca2+ in Cardiac Myocytes. Biophys. J. 2008, 95, 4597–4612. [Google Scholar] [CrossRef] [Green Version]
- Shi, X.M.; Zheng, Y.F.; Liu, Z.R.; Yang, W.Z. A Model of Calcium Signaling and Degranulation Dynamics Induced by Laser Irradiation in Mast Cells. Chinese Sci. Bull. 2008, 53, 2315–2325. [Google Scholar] [CrossRef] [Green Version]
- Babbs, C.F.; Steiner, M.G. Simulation of Free Radical Reactions in Biology and Medicine: A New Two-Compartment Kinetic Model of Intracellular Lipid Peroxidation. Free Radic. Biol. Med. 1990, 8, 471–485. [Google Scholar] [CrossRef] [Green Version]
- Antunes, F.; Salvador, A.; Marinho, H.S.; Alves, R.; Pinto, R.E. Lipid Peroxidation in Mitochondrial Inner Membranes I. An Integrative Kinetic Model. Free Radic. Biol. Med. 1996, 21, 917–943. [Google Scholar] [CrossRef]
- Shi, Y.; Mellier, G.; Huang, S.; White, J.; Pervaiz, S.; Tucker-Kellogg, L. Computational Modelling of LY303511 and TRAIL-Induced Apoptosis Suggests Dynamic Regulation of CFLIP. Bioinformatics 2013, 29, 347–354. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kavdia, M. Mathematical and Computational Models of Oxidative and Nitrosative Stress. Crit. Rev. Biomed. Eng. 2011, 39, 461–472. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aydemir, T.; Kuru, K.; Aydem, T. Purification and Partial Characterization of Catalase from Chicken Erythrocytes and the Effect of Various Inhibitors on Enzyme Activity. Turkish J. Chem. 2003, 27, 85–98. [Google Scholar]
- Henle, E.S.; Luo, Y.; Linn, S. Fe2+, Fe3+, and Oxygen React with DNA-Derived Radicals Formed during Iron-Mediated Fenton Reactions†. Biochemistry 1996, 35, 12212–12219. [Google Scholar] [CrossRef] [PubMed]
- Xue, C.; Chou, C.S.; Kao, C.Y.; Sen, C.K.; Friedman, A. Propagation of Cutaneous Thermal Injury: A Mathematical Model. Wound Repair Regen. 2012, 20, 114–122. [Google Scholar] [CrossRef]
- Buettner, G.R.; Ng, C.F.; Wang, M.; Rodgers, V.G.J.; Schafer, F.Q. A New Paradigm: Manganese Superoxide Dismutase Influences the Production of H2O2 in Cells and Thereby Their Biological State. Free Radic. Biol. Med. 2006, 41, 1338–1350. [Google Scholar] [CrossRef] [Green Version]
- Edwards, A.; Cao, C.; Pallone, T.L. Cellular Mechanisms Underlying Nitric Oxide-Induced Vasodilation of Descending Vasa Recta. Am. J. Physiol. Physiol. 2011, 300, F441–F456. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ozhogina, O.A.; Kasaikina, O.T. Β-Carotene As an Interceptor of Free Radicals. Free Radic. Biol. Med. 1995, 19, 575–581. [Google Scholar] [CrossRef]
- Haila, K. Effects of Carotenoids and Carotenoid- Tocopherol Interaction on Lipid Oxidation In Vitro; Semantic Scholar: Seattle, WA, USA, 1999; Volume 62, ISBN 9514586670. [Google Scholar]
- MacFarlane, N.G.; Miller, D.J. Depression of Peak Force without Altering Calcium Sensitivity by the Superoxide Anion in Chemically Skinned Cardiac Muscle of Rat. Circ. Res. 1992, 70, 1217–1224. [Google Scholar] [CrossRef] [Green Version]
- Koshkin, V.; Lotan, O.; Pick, E. Electron Transfer in the Superoxide-Generating NADPH Oxidase Complex Reconstituted in Vitro. Biochim. Biophys. Acta - Bioenerg. 1997, 1319, 139–146. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| In Silico Experiment | COX-2 (nM) |
|---|---|
| Apigenin First, Hesperidin Second | 1.93 |
| Hesperidin First, Apigenin Second | 1.54 |
| Apigenin and Hesperidin Simultaneously | 0.28 |
| In Silico Experiment | PGE2 (AUC) |
|---|---|
| Apigenin First, Hesperidin Second | 289 |
| Hesperidin First, Apigenin Second | 217 |
| Apigenin and Hesperidin Simultaneously | 286 |
| In Silico Experiment | TRPV1 (nM) |
|---|---|
| Apigenin First, Hesperidin Second | 0.0027 |
| Hesperidin First, Apigenin Second | 0.0325 |
| Apigenin and Hesperidin Simultaneously | 0.00019 |
| In Silico Experiment | CGRP (nM) |
|---|---|
| Apigenin First, Hesperidin Second | 1.17 × 10−6 |
| Hesperidin First, Apigenin Second | 0.066 |
| Apigenin and Hesperidin Simultaneously | 2.65 × 10−10 |
| In Silico Experiment | ROS (nM) |
|---|---|
| Apigenin First, Hesperidin Second | 1.34 |
| Hesperidin First, Apigenin Second | 12.69 |
| Apigenin and Hesperidin Simultaneously | 1.117 |
| Joint Pain Mechanisms of Action | Apigenin | Hesperidin |
|---|---|---|
| COX-2 Production | Inhibits IKK activation | Inhibits NF-kB |
| Arachidonic Acid Metabolism | Inhibits COX-2 | Inhibits COX-2 |
| PGE2 Signaling | Inhibits PKC activation | Inhibits PLC activation |
| Oxidative Stress Pathway | ROS Production | ROS Production |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ayyadurai, V.A.S.; Deonikar, P. In Silico Modeling and Quantification of Synergistic Effects of Multi-Combination Compounds: Case Study of the Attenuation of Joint Pain Using a Combination of Phytonutrients. Appl. Sci. 2022, 12, 10013. https://doi.org/10.3390/app121910013
Ayyadurai VAS, Deonikar P. In Silico Modeling and Quantification of Synergistic Effects of Multi-Combination Compounds: Case Study of the Attenuation of Joint Pain Using a Combination of Phytonutrients. Applied Sciences. 2022; 12(19):10013. https://doi.org/10.3390/app121910013
Chicago/Turabian StyleAyyadurai, V. A. Shiva, and Prabhakar Deonikar. 2022. "In Silico Modeling and Quantification of Synergistic Effects of Multi-Combination Compounds: Case Study of the Attenuation of Joint Pain Using a Combination of Phytonutrients" Applied Sciences 12, no. 19: 10013. https://doi.org/10.3390/app121910013
APA StyleAyyadurai, V. A. S., & Deonikar, P. (2022). In Silico Modeling and Quantification of Synergistic Effects of Multi-Combination Compounds: Case Study of the Attenuation of Joint Pain Using a Combination of Phytonutrients. Applied Sciences, 12(19), 10013. https://doi.org/10.3390/app121910013