
Microbial Growth Dynamics in Minced Meat Enriched with Plant Powders
 , , , , , , ,
, , , , , , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Meat and Plant Material
2.2. Sample Preparation
2.3. pH Determination
2.4. Water Activity (aw) Determination
2.5. Microbial Enumeration
2.6. Challenge Test
Bacterial Species and Strains
2.7. Statistical Analysis
3. Results
3.1. Microbial Counts in Plant Powders
3.2. Total Viable Counts in Raw Meat Samples
3.3. Pseudomonas Counts in Raw Meat Samples
3.4. Yeast and Mould Counts in Raw Meat Samples
3.5. Differences in Microbial Counts in Raw Meat Samples
3.6. Microbial Counts in Cooked Meat Samples
3.7. pH and Water Activity
3.8. Challenge Testing
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Bohrer, B.M. Nutrient density and nutritional value of meat products and non-meat foods high in protein. Trends Food Sci. Technol. 2017, 65, 103–112. [Google Scholar] [CrossRef]
- European Commission. Commission Regulation (EC) No 2073/2005 of 15 November 2005 on microbiological criteria for foodstuffs. Off. J. Eur. Union 2005, 338, 1–26. Available online: https://eur-lex.europa.eu/legal-content/EN/TXT/PDF/?uri=CELEX:02005R2073-20200308&from=EN (accessed on 6 August 2022).
- Gill, A. The importance of bacterial culture to food microbiology in the age of genomics. Front. Microbiol. 2017, 8, 777. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Björkroth, J. Microbiological ecology of marinated meat products. Meat Sci. 2005, 70, 477–480. [Google Scholar] [CrossRef]
- Raudsepp, P.; Koskar, J.; Anton, D.; Meremäe, K.; Kapp, K.; Laurson, P.; Bleive, U.; Kaldmäe, H.; Roasto, M.; Püssa, T. Antibacterial and antioxidative properties of different parts of garden rhubarb, black currant, chokeberry and blue honeysuckle. J. Sci. Food Agric. 2018, 99, 2311–2320. [Google Scholar] [CrossRef]
- Gonelimali, F.D.; Lin, J.; Miao, W.; Xuan, J.; Charles, F.; Chen, M.; Hatab, S.R. Antimicrobial properties and mechanism of action of some plant extracts against food pathogens and spoilage microorganisms. Front. Microbiol. 2018, 9, 1639. [Google Scholar] [CrossRef]
- Pisoschi, A.M.; Pop, A.; Georgescu, C.; Turcus, V.; Olah, N.K.; Mathe, E. An overview of natural antimicrobials role in food. Eur. J. Med. Chem. 2018, 143, 922–935. [Google Scholar] [CrossRef]
- Anton, D.; Koskar, J.; Raudsepp, P.; Meremäe, K.; Kaart, T.; Püssa, T.; Roasto, M. Antimicrobial and antioxidative effects of plant powders in raw and cooked minced pork. Foods 2019, 8, 661. [Google Scholar] [CrossRef] [Green Version]
- Kalogianni, A.I.; Lazou, T.; Bossis, I.; Gelasakis, A.I. Natural phenolic compounds for the control of oxidation, bacterial spoilage, and foodborne pathogens in meat. Foods 2020, 9, 794. [Google Scholar] [CrossRef]
- Reis, F.S.; Stojković, D.; Barros, L.; Glamočlija, J.; Ćirić, A.; Soković, M.; Martins, A.; Vasconcelos, M.H.; Morales, P.; Ferreira, I.C.F.R. Can Suillus granulatus (L.) Roussel be classified as a functional food? Food Funct. 2014, 5, 2861–2869. [Google Scholar] [CrossRef] [Green Version]
- Singh, B.; Falahee, M.B.; Adams, M.R. Synergistic inhibition of Listeria monocytogenes by nisin and garlic extract. Food Microbiol. 2001, 18, 133–139. [Google Scholar] [CrossRef]
- Liaqat, A.; Zahoor, T.; Randhawa, M.A.; Shahid, M. Characterization and antimicrobial potential of bioactive components of sonicated extract from garlic (Allium sativum) against foodborne pathogens. J. Food Process. Preserv. 2019, 43, e13936. [Google Scholar] [CrossRef]
- Ankri, S.; Mirelman, D. Antimicrobial properties of allicin from garlic. Microbes Infect. 1999, 2, 125–129. [Google Scholar] [CrossRef]
- Raudsepp, P.; Kaldmäe, H.; Kikas, A.; Libek, A.-V.; Püssa, T. Nutritional quality of berries and bioactive compounds in the leaves of black currant (Ribes nigrum L.) cultivars evaluated in Estonia. J. Berry Res. 2010, 1, 53–59. [Google Scholar] [CrossRef] [Green Version]
- Gironés-Vilaplana, A.; Valentão, P.; Andrade, P.B.; Ferreres, F.; Moreno, D.A.; García-Viguera, C. Phytochemical profile of a blend of black chokeberry and lemon juice with cholinesterase inhibitory effect and antioxidant potential. Food Chem. 2012, 134, 2090–2096. [Google Scholar] [CrossRef] [PubMed]
- Mlcek, J.; Rop, O.; Jurikova, T.; Sochor, J.; Fisera, M.; Balla, S.; Baron, M.; Hrabe, J. Bioactive compounds in sweet rowanberry fruits of interspecific Rowan crosses. Cent. Eur. J. Biol. 2014, 9, 1078–1086. [Google Scholar] [CrossRef]
- Bobinaitė, R.; Grootaert, C.; Van Camp, J.; Šarkinas, A.; Liaudanskas, M.; Žvikas, V.; Viškelis, P.; Venskutonis, P.R. Chemical composition, antioxidant, antimicrobial and antiproliferative activities of the extracts isolated from the pomace of rowanberry (Sorbus aucuparia L.). Food Res. Int. 2020, 136, 109310. [Google Scholar] [CrossRef]
- European Union Reference Laboratory for Listeria monocytogenes. EURL Lm Technical Guidance Document on Challenge Tests and Durability Studies for Assessing Shelf-Life of Ready-to-Eat Foods Related to Listeria monocytogenes, Version 4 of 1 July 2021. ANSES Laboratory for Food Safety. 2021. Available online: https://food.ec.europa.eu/system/files/2021-07/biosafety_fh_mc_tech-guide-doc_listeria-in-rte-foods_en_0.pdf (accessed on 4 August 2022).
- Hurtado, M.I.; Gaviria, C.A.; Cardona, O.A. Estimating shelf life on beef and pork meat with different fat content. Vitae 2014, 21, 201–210. [Google Scholar]
- Ortiz, M. Antimicrobial activity of onion and ginger against two food borne pathogens Escherichia coli and Staphylococcus aureus. MOJ Food Process. Technol. 2015, 1, 104–112. [Google Scholar] [CrossRef] [Green Version]
- Kabrah, A.M.; Faidah, H.S.; Ashshi, A.M.; Turkistani, S.A. Antibacterial effect of onion. Sch. J. Appl. Med. Sci. 2016, 4, 4128–4133. [Google Scholar]
- Babaoğlu, A.S.; Unal, K.; Dilek, N.M.; Poçan, H.B.; Karakaya, M. Antioxidant and antimicrobial effects of blackberry, black chokeberry, blueberry, and red currant pomace extracts on beef patties subject to refrigerated storage. Meat Sci. 2022, 187, 108765. [Google Scholar] [CrossRef] [PubMed]
- Xi, Y.; Sullivan, G.A.; Jackson, A.L.; Zhou, G.H.; Sebranek, J.G. Use of natural antimicrobials to improve the control of Listeria monocytogenes in a cured cooked meat model system. Meat Sci. 2011, 88, 503–511. [Google Scholar] [CrossRef]
- Koskar, J.; Kramarenko, T.; Meremäe, K.; Kuningas, M.; Sõgel, J.; Mäesaar, M.; Anton, D.; Lillenberg, M.; Roasto, M. Prevalence and numbers of Listeria monocytogenes in various ready-to-eat foods over a 5-year period in Estonia. J. Food Prot. 2019, 4, 597–604. [Google Scholar] [CrossRef] [PubMed]
- Mäesaar, M.; Mamede, R.; Elias, T.; Roasto, M. Retrospective use of whole-genome sequencing expands the multicountry outbreak cluster of Listeria monocytogenes ST1247. Int. J. Genom. 2021, 2021, 6636138. [Google Scholar] [CrossRef] [PubMed]
- Kosikowska, U.; Smolarz, H.D.; Malm, A. Antimicrobial activity and total content of polyphenols of Rheum L. species growing in Poland. Cent. Eur. J. Biol. 2010, 5, 814–820. [Google Scholar] [CrossRef]
- Raudsepp, P.; Anton, D.; Roasto, M.; Meremäe, K.; Pedastsaar, P.; Mäesaar, M.; Raal, A.; Laikoja, K.; Püssa, T. The antioxidative and antimicrobial properties of the blue honeysuckle (Lonicera caerulea L.), Siberian rhubarb (Rheum rhaponticum L.) and some other plants, compared to ascorbic acid and sodium nitrite. Food Control 2013, 31, 129–135. [Google Scholar] [CrossRef]
- Paunović, S.M.; Mašković, P.; Nikolić, M.; Miletić, R. Bioactive compounds and antimicrobial activity of black currant (Ribes nigrum L.) berries and leaves extract obtained by different soil management system. Sci. Hortic. 2017, 222, 69–75. [Google Scholar] [CrossRef]
- Wu, V.C.-H.; Qiu, X.; Bushway, A.; Harper, L. Antibacterial effects of American cranberry (Vaccinium macrocarpon) concentrate on foodborne pathogens. LWT Food Sci. Technol. 2008, 41, 1834–1841. [Google Scholar] [CrossRef]
- Zhang, L.-L.; Zhang, L.-F.; Xu, J.-G. Chemical composition, antibacterial activity and action mechanism of different extracts from hawthorn (Crataegus pinnatifida Bge.). Sci. Rep. 2020, 10, 8876. [Google Scholar] [CrossRef]
- Ravichandran, M.; Hettiarachchy, N.S.; Ganesh, V.; Ricke, S.; Singh, S. Enhancement of antimicrobial activities of naturally occurring phenolic compounds by nanoscale delivery against Listeria monocytogenes, Escherichia coli O157:H7 and Salmonella Typhimurium in broth and chicken meat system. J. Food Saf. 2011, 31, 462–471. [Google Scholar] [CrossRef]
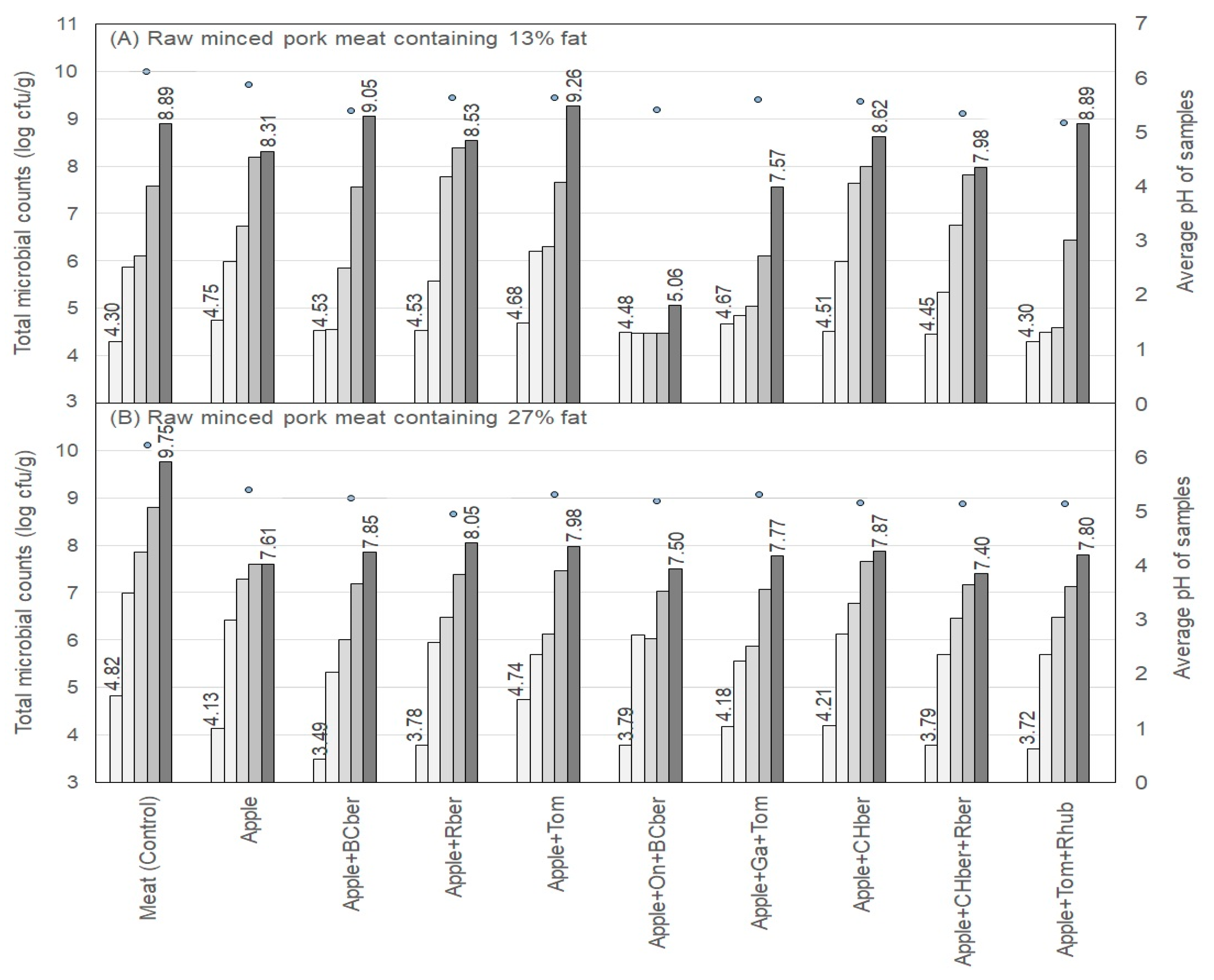
 ) of raw samples containing 13% (A) and 27% (B) fat after 1, 5, 7, 9, and 12 days of storage (columns from left to right). The values on the first and the last day are presented numerically. pH (
) of raw samples containing 13% (A) and 27% (B) fat after 1, 5, 7, 9, and 12 days of storage (columns from left to right). The values on the first and the last day are presented numerically. pH ( ) values are shown as an average throughout the storage period. Abbreviations: BCber, blackcurrant berries; Rber, rowanberries; Tom, tomato; On, onion; Ga, garlic; CHber, chokeberries; Rhub, rhubarb petioles.
) of raw samples containing 13% (A) and 27% (B) fat after 1, 5, 7, 9, and 12 days of storage (columns from left to right). The values on the first and the last day are presented numerically. pH () values are shown as an average throughout the storage period. Abbreviations: BCber, blackcurrant berries; Rber, rowanberries; Tom, tomato; On, onion; Ga, garlic; CHber, chokeberries; Rhub, rhubarb petioles.
) values are shown as an average throughout the storage period. Abbreviations: BCber, blackcurrant berries; Rber, rowanberries; Tom, tomato; On, onion; Ga, garlic; CHber, chokeberries; Rhub, rhubarb petioles.
) of raw samples containing 13% (A) and 27% (B) fat after 1, 5, 7, 9, and 12 days of storage (columns from left to right). The values on the first and the last day are presented numerically. pH () values are shown as an average throughout the storage period. Abbreviations: BCber, blackcurrant berries; Rber, rowanberries; Tom, tomato; On, onion; Ga, garlic; CHber, chokeberries; Rhub, rhubarb petioles.
 ) of raw samples containing 13% (A) and 27% (B) fat after storage times of 1, 5, 7, 9, and 12 days (columns from left to right). The values on the first and the last day are presented numerically. pH (
) of raw samples containing 13% (A) and 27% (B) fat after storage times of 1, 5, 7, 9, and 12 days (columns from left to right). The values on the first and the last day are presented numerically. pH ( ) values are averages throughout the storage period. Abbreviations: BCber, blackcurrant berries; Rber, rowanberries; Tom, tomato; On, onion; Ga, garlic; CHber, chokeberries; Rhub, rhubarb petioles.
) of raw samples containing 13% (A) and 27% (B) fat after storage times of 1, 5, 7, 9, and 12 days (columns from left to right). The values on the first and the last day are presented numerically. pH () values are averages throughout the storage period. Abbreviations: BCber, blackcurrant berries; Rber, rowanberries; Tom, tomato; On, onion; Ga, garlic; CHber, chokeberries; Rhub, rhubarb petioles.
) values are averages throughout the storage period. Abbreviations: BCber, blackcurrant berries; Rber, rowanberries; Tom, tomato; On, onion; Ga, garlic; CHber, chokeberries; Rhub, rhubarb petioles.
) of raw samples containing 13% (A) and 27% (B) fat after storage times of 1, 5, 7, 9, and 12 days (columns from left to right). The values on the first and the last day are presented numerically. pH () values are averages throughout the storage period. Abbreviations: BCber, blackcurrant berries; Rber, rowanberries; Tom, tomato; On, onion; Ga, garlic; CHber, chokeberries; Rhub, rhubarb petioles.
 ) of raw samples containing 13% (A) and 27% (B) fat 1, 5, 7, 9, and 12 days of storage (columns from left to right). The values on the first and the last day are presented numerically. pH (
) of raw samples containing 13% (A) and 27% (B) fat 1, 5, 7, 9, and 12 days of storage (columns from left to right). The values on the first and the last day are presented numerically. pH ( ) values are shown as an average throughout the storage period. Abbreviations: BCber, blackcurrant berries; Rber, rowanberries; Tom, tomato; On, onion; Ga, garlic; CHber, chokeberries; Rhub, rhubarb petioles.
) of raw samples containing 13% (A) and 27% (B) fat 1, 5, 7, 9, and 12 days of storage (columns from left to right). The values on the first and the last day are presented numerically. pH () values are shown as an average throughout the storage period. Abbreviations: BCber, blackcurrant berries; Rber, rowanberries; Tom, tomato; On, onion; Ga, garlic; CHber, chokeberries; Rhub, rhubarb petioles.
) values are shown as an average throughout the storage period. Abbreviations: BCber, blackcurrant berries; Rber, rowanberries; Tom, tomato; On, onion; Ga, garlic; CHber, chokeberries; Rhub, rhubarb petioles.
) of raw samples containing 13% (A) and 27% (B) fat 1, 5, 7, 9, and 12 days of storage (columns from left to right). The values on the first and the last day are presented numerically. pH () values are shown as an average throughout the storage period. Abbreviations: BCber, blackcurrant berries; Rber, rowanberries; Tom, tomato; On, onion; Ga, garlic; CHber, chokeberries; Rhub, rhubarb petioles.

{kind=link}
{kind=link}
{kind=link}
| Abbreviation | Composition |
|---|---|
| Meat (Control) | Minced meat with 13% and 27% fat content |
| Apple | Minced meat with 3% apple |
| Apple+BCber | Minced meat with 3% apple+2% blackcurrant berries |
| Apple+Rber | Minced meat with 3% apple+2% rowanberries |
| Apple+Tom | Minced meat with 3% apple+2% tomato |
| Apple+On+BCber | Minced meat with 3% apple+1% onion+2% blackcurrant berries |
| Apple+Ga+Tom | Minced meat with 3% apple+1% garlic+2% tomato |
| Apple+CHber | Minced meat with 3% apple+2% chokeberries |
| Apple+CHber+Rber | Minced meat with 3% apple+2% chokeberries+2% rowanberries |
| Apple+Tom+Rhub | Minced meat with 3% apple+2% tomato+1% rhubarb petioles |
| Garlic | Minced meat with 2% garlic * |
| Onion | Minced meat with 2% onion * |
| Rhub | Minced meat with 2% rhubarb petioles * |
| Plant Powder | Total Viable Count (cfu/g) | Pseudomonas Count (cfu/g) | Yeast and Mould Count (cfu/g) |
|---|---|---|---|
| Apple | 150 | <100 | 30 |
| Blackcurrant berry | 680 | <100 | 230 |
| Rowanberry | 100 | <100 | 140 |
| Tomato | 200 | <100 | 40 |
| Onion | 2100 | <100 | 350 |
| Garlic | 1200 | <100 | 1900 |
| Chokeberry | 4500 | <100 | 20 |
| Rhubarb petioles | 400 | <100 | 20 |
| Group | Minced Meat 13% | Minced Meat 27% | ||||
|---|---|---|---|---|---|---|
| Plant Powder | Total Microbial | Pseudomonas spp. | Yeasts and Moulds | Total Microbial * | Pseudomnas spp. | Yeasts and Moulds |
| Apple | 0.901 | 0.811 | 0.827 | ND | 0.030 ** | 0.142 |
| Apple+BCber | 0.892 | 1.000 | 0.421 | ND | 0.001 ** | 0.160 |
| Apple+Rber | 0.901 | 0.621 | 0.130 | ND | 0.001 ** | 0.160 |
| Apple+Tom | 0.586 | 0.630 | 0.190 | ND | 0.002 ** | 0.014 ** |
| Apple+On+BCber | 0.008 ** | 0.004 ** | 0.329 | ND | 0.002 ** | 0.142 |
| Apple+Ga+Tom | 0.315 | 0.053 | 0.421 | ND | 0.002 ** | 0.007 ** |
| Apple+CHber | 0.782 | 0.621 | 0.114 | ND | 0.002 ** | 0.067 |
| Apple+CHber+Rber | 0.964 | 1.000 | 0.087 | ND | 0.001 ** | 0.067 |
| Apple+Tom+Rhub | 0.915 | 0.004 ** | 0.022 ** | ND | 0.001 ** | 0.380 |
| Samples | Storage Day | pH * | Water Activity * aw | Total Count (cfu/g) | δ (log cfu/g) ** |
|---|---|---|---|---|---|
| Meat *** | 0 | 6.35 ± 0.048 | 0.986 ± 0.003 | <1.0 × 102 | 7.30 |
| 6 | 6.30 ± 0.021 | 0.987 ± 0.003 | 4.3 × 102 | ||
| 12 | 6.35 ± 0.001 | 0.992 ± 0.005 | 2.5 × 102 | ||
| Apple+On+BCber | 0 | 5.75 ± 0.071 | 0.987 ± 0.002 | <1.0 × 102 | 3.63 |
| 6 | 5.70 ± 0.016 | 0.985 ± 0.001 | <1.0 × 102 | ||
| 12 | 5.72 ± 0.005 | 0.984 ± 0.004 | 1.0 × 102 | ||
| Apple+Ga+Tom | 0 | 5.93 ± 0.066 | 0.986 ± 0.001 | <1.0 × 102 | 5.35 |
| 6 | 5.87 ± 0.021 | 0.990 ± 0.001 | 1.0 × 102 | ||
| 12 | 5.95 ± 0.010 | 0.979 ± 0.001 | 2.0 × 102 | ||
| Apple+Tom+Rhub | 0 | 5.51 ± 0.011 | 0.988 ± 0.002 | <1.0 × 102 | 3.74 |
| 6 | 5.42 ± 0.013 | 0.987 ± 0.001 | <1.0 × 102 | ||
| 12 | 5.48 ± 0.012 | 0.977 ± 0.001 | 1.5 × 102 | ||
| Garlic | 0 | 6.68 ± 0.015 | 0.986 ± 0.002 | 1.5 × 101 | 7.85 |
| 6 | 6.73 ± 0.001 | 0.990 ± 0.003 | 5.5 × 101 | ||
| 12 | 6.72 ± 0.003 | 0.987 ± 0.002 | 2.0 × 101 | ||
| Onion | 0 | 6.48 ± 0.011 | 0.964 ± 0.005 | 2.0 × 101 | 7.92 |
| 6 | 6.58 ± 0.008 | 0.984 ± 0.001 | 3.6 × 101 | ||
| 12 | 6.60 ± 0.017 | 0.979 ± 0.001 | 3.0 × 101 | ||
| Rhub | 0 | 5.24 ± 0.001 | 0.980 ± 0.001 | 1.0 × 101 | 2.74 |
| 6 | 5.31 ± 0.013 | 0.987 ± 0.001 | 3.5 × 101 | ||
| 12 | 5.41 ± 0.059 | 0.979 ± 0.001 | 3.0 × 101 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Koskar, J.; Meremäe, K.; Püssa, T.; Anton, D.; Elias, T.; Rätsep, R.; Mäesaar, M.; Kapp, K.; Roasto, M. Microbial Growth Dynamics in Minced Meat Enriched with Plant Powders. Appl. Sci. 2022, 12, 11292. https://doi.org/10.3390/app122111292
Koskar J, Meremäe K, Püssa T, Anton D, Elias T, Rätsep R, Mäesaar M, Kapp K, Roasto M. Microbial Growth Dynamics in Minced Meat Enriched with Plant Powders. Applied Sciences. 2022; 12(21):11292. https://doi.org/10.3390/app122111292
Chicago/Turabian StyleKoskar, Julia, Kadrin Meremäe, Tõnu Püssa, Dea Anton, Terje Elias, Reelika Rätsep, Mihkel Mäesaar, Karmen Kapp, and Mati Roasto. 2022. "Microbial Growth Dynamics in Minced Meat Enriched with Plant Powders" Applied Sciences 12, no. 21: 11292. https://doi.org/10.3390/app122111292
APA StyleKoskar, J., Meremäe, K., Püssa, T., Anton, D., Elias, T., Rätsep, R., Mäesaar, M., Kapp, K., & Roasto, M. (2022). Microbial Growth Dynamics in Minced Meat Enriched with Plant Powders. Applied Sciences, 12(21), 11292. https://doi.org/10.3390/app122111292