


Evaluating the Potential Anticancer Properties of Salvia triloba in Human-Osteosarcoma U2OS Cell Line and Ovarian Adenocarcinoma SKOV3 Cell Line

 , , , ,
, , , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material Harvest
2.2. Crude Extract Preparation
2.3. Cell Culture
2.4. MTT Viability Assay
2.5. Selectivity Index
2.6. Combination of S. triloba Acetone Extract and Paclitaxel in SKOV3 Cells
2.7. Trypan Blue Exclusion Assay
2.8. Scratch-Wound Healing Assay
2.9. Transwell Migration Assay
2.10. Reverse Transcription Quantitative Polymerase Chain Reaction (RT-qPCR)
2.11. Transcription Factor Binding Sites Annotation
2.12. Hemolysis Assay
2.13. TLC-UV Fractionation of Acetone Crude Extract
2.14. Statistical Analysis
3. Results
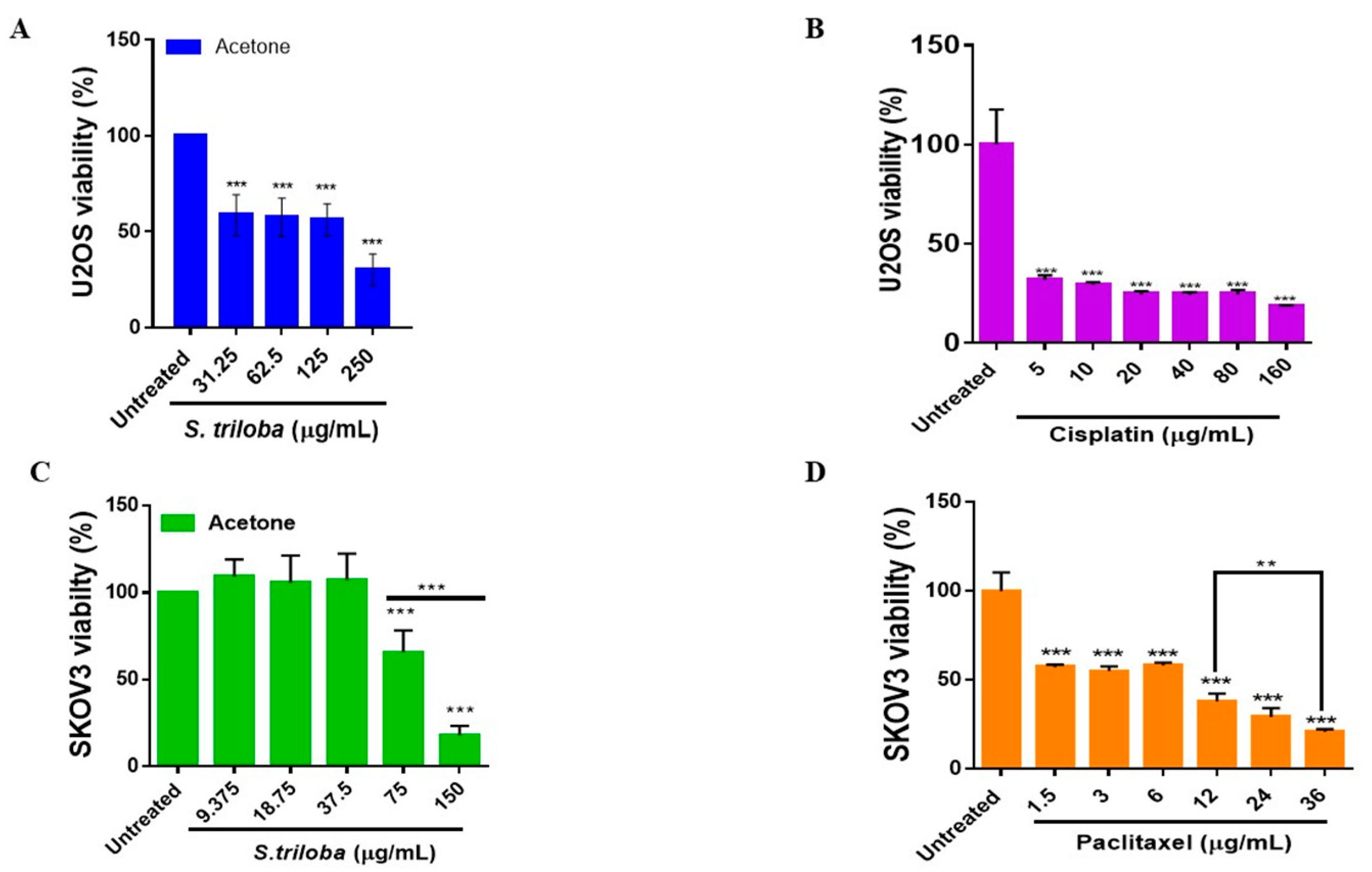
3.1. S. triloba Acetone Extract Significantly Reduces U2OS and SKOV3 Cell Viability
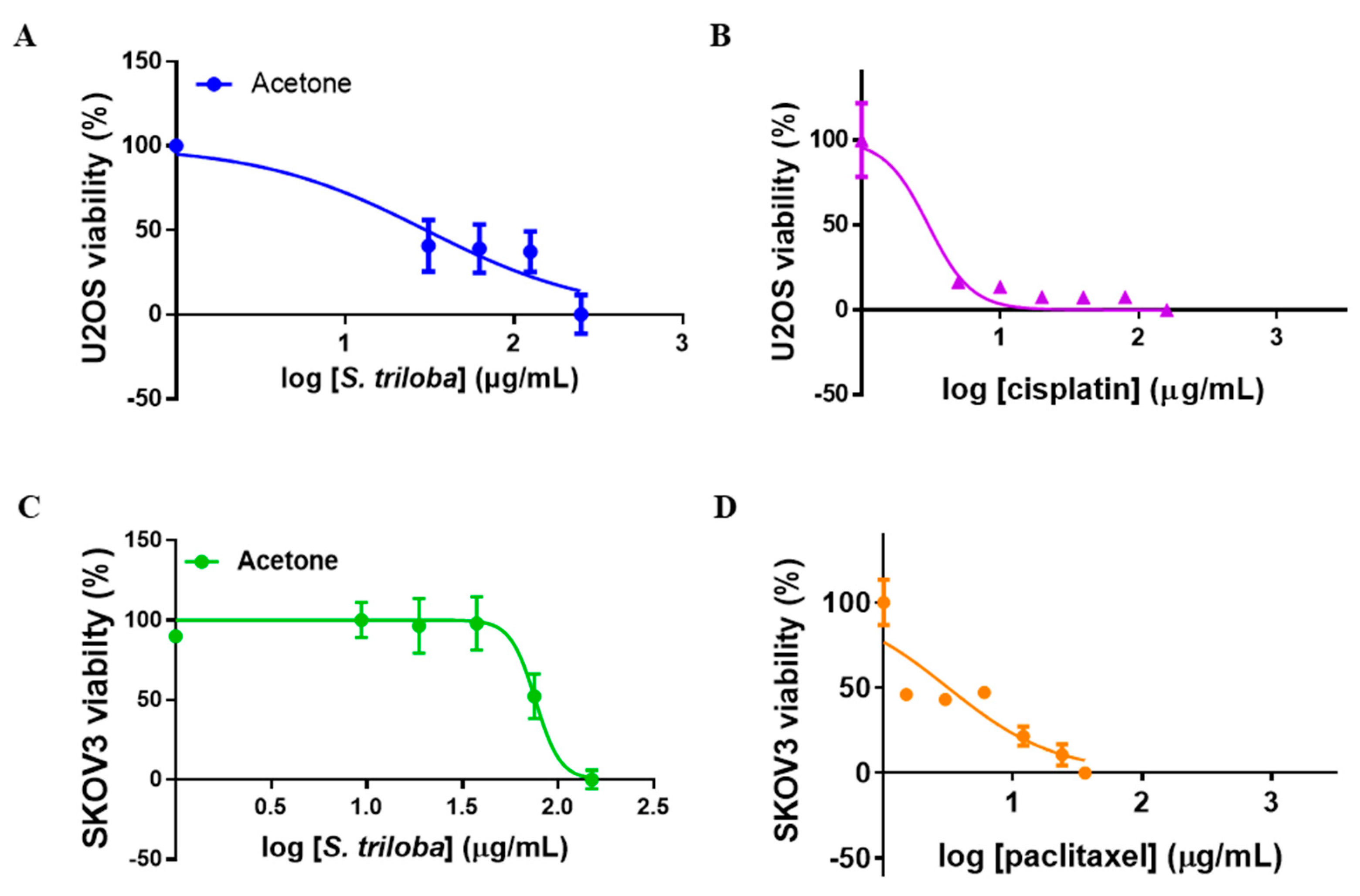
3.2. Selectivity Index of S. triloba Acetone Extract for U2OS and SKOV3 Cells
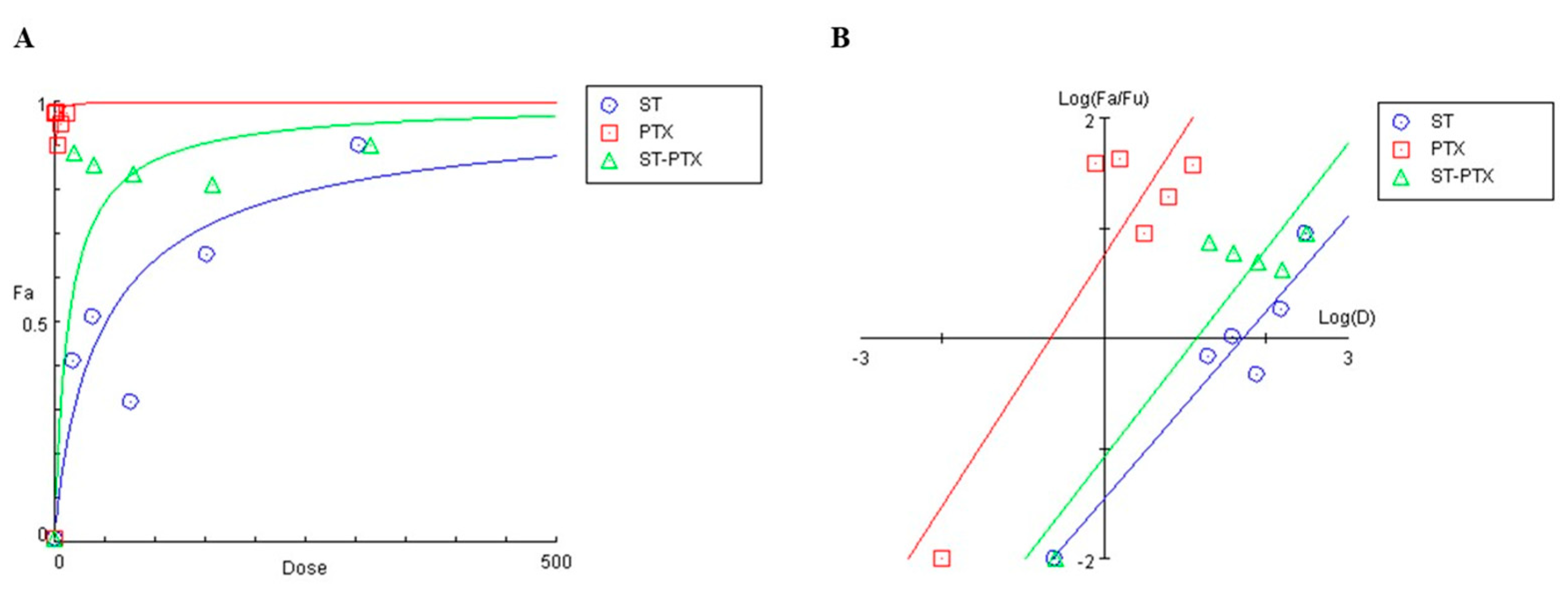
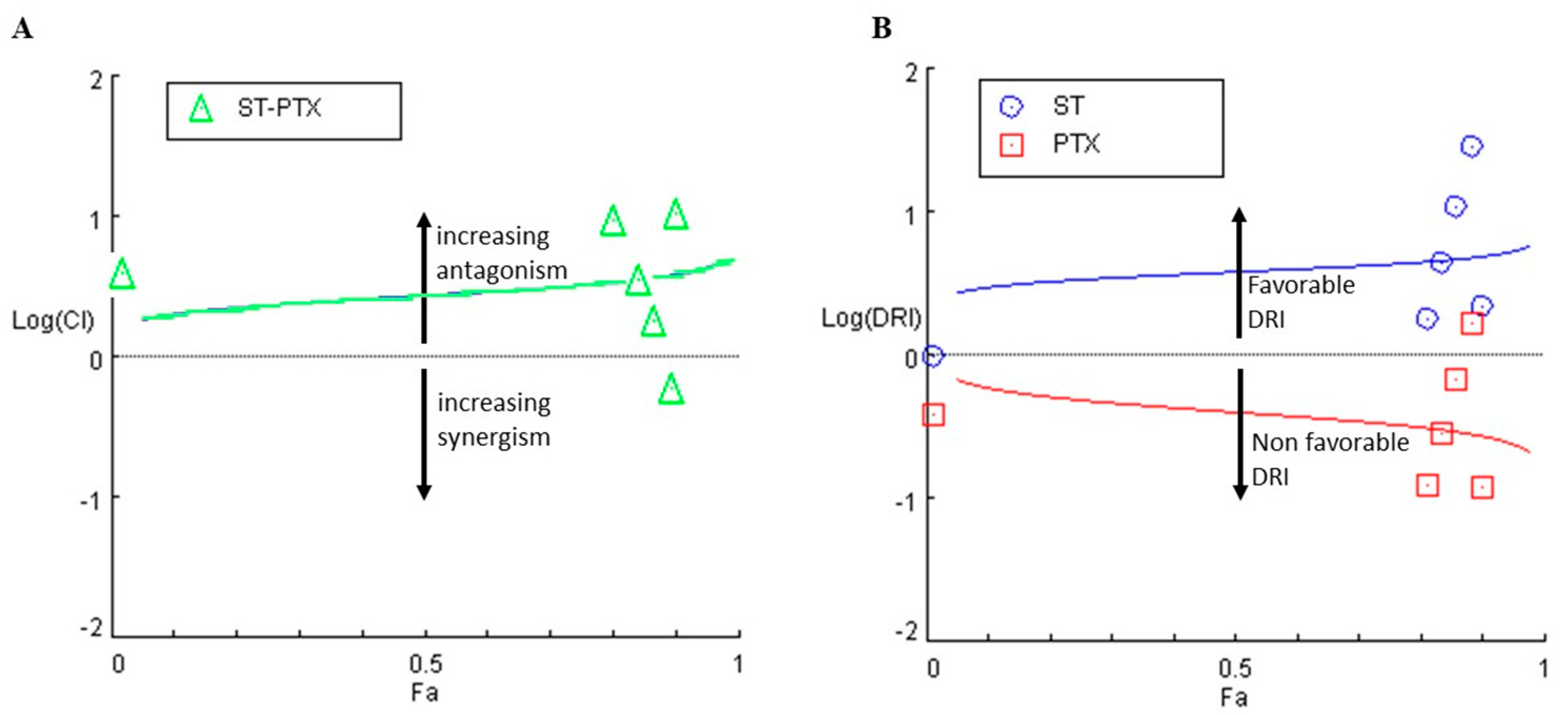
3.3. The Combination of S. triloba Acetone Extract and Paclitaxel Exhibits Strong Synergism in SKOV3 Cells
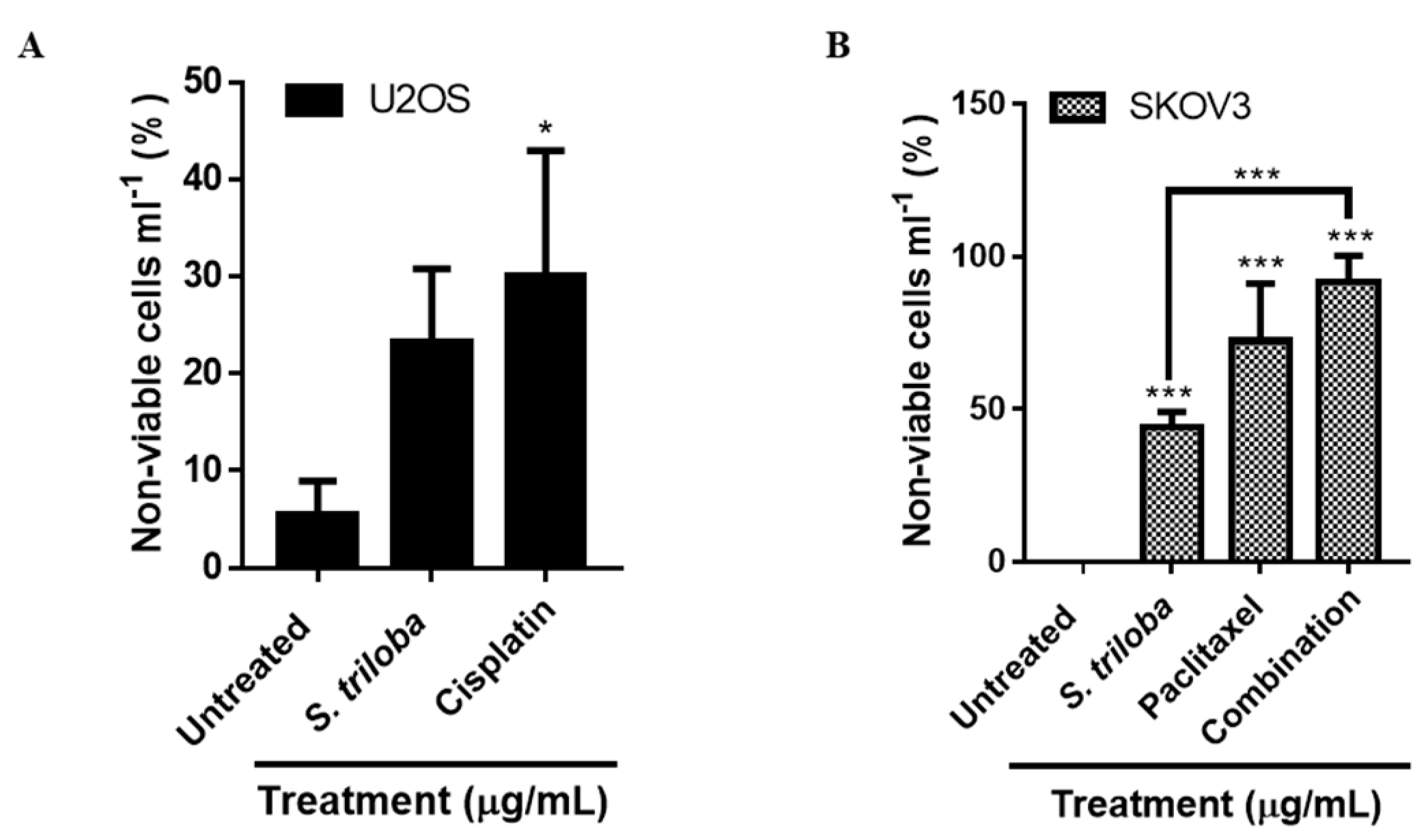
3.4. S. triloba Acetone Extract Increases the Percentage of Non-Viable Cells in Both U2OS and SKOV3 Cells
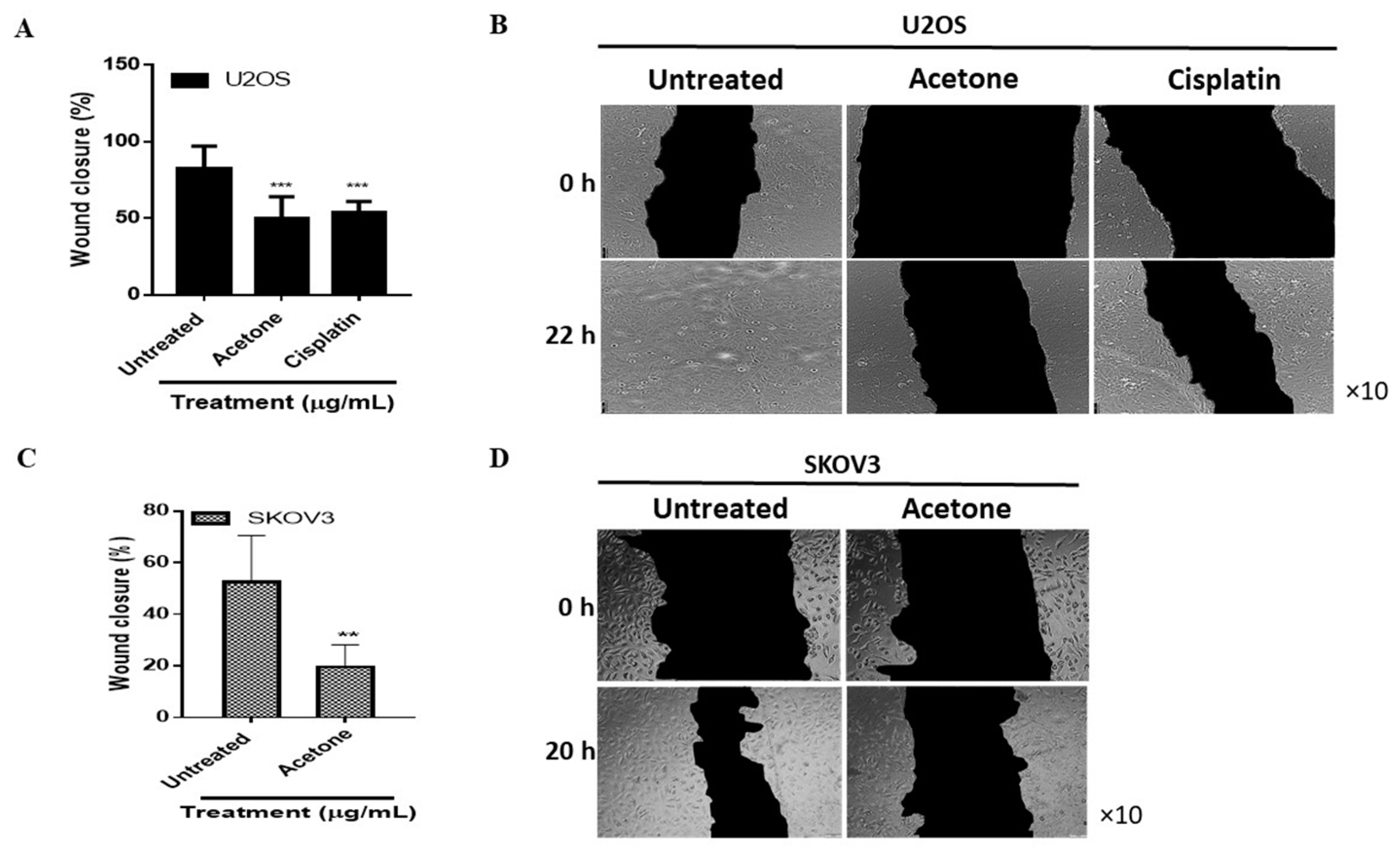
3.5. S. triloba Acetone Extract Decreases Wound Closure in Both U2OS and SKOV3 Cells
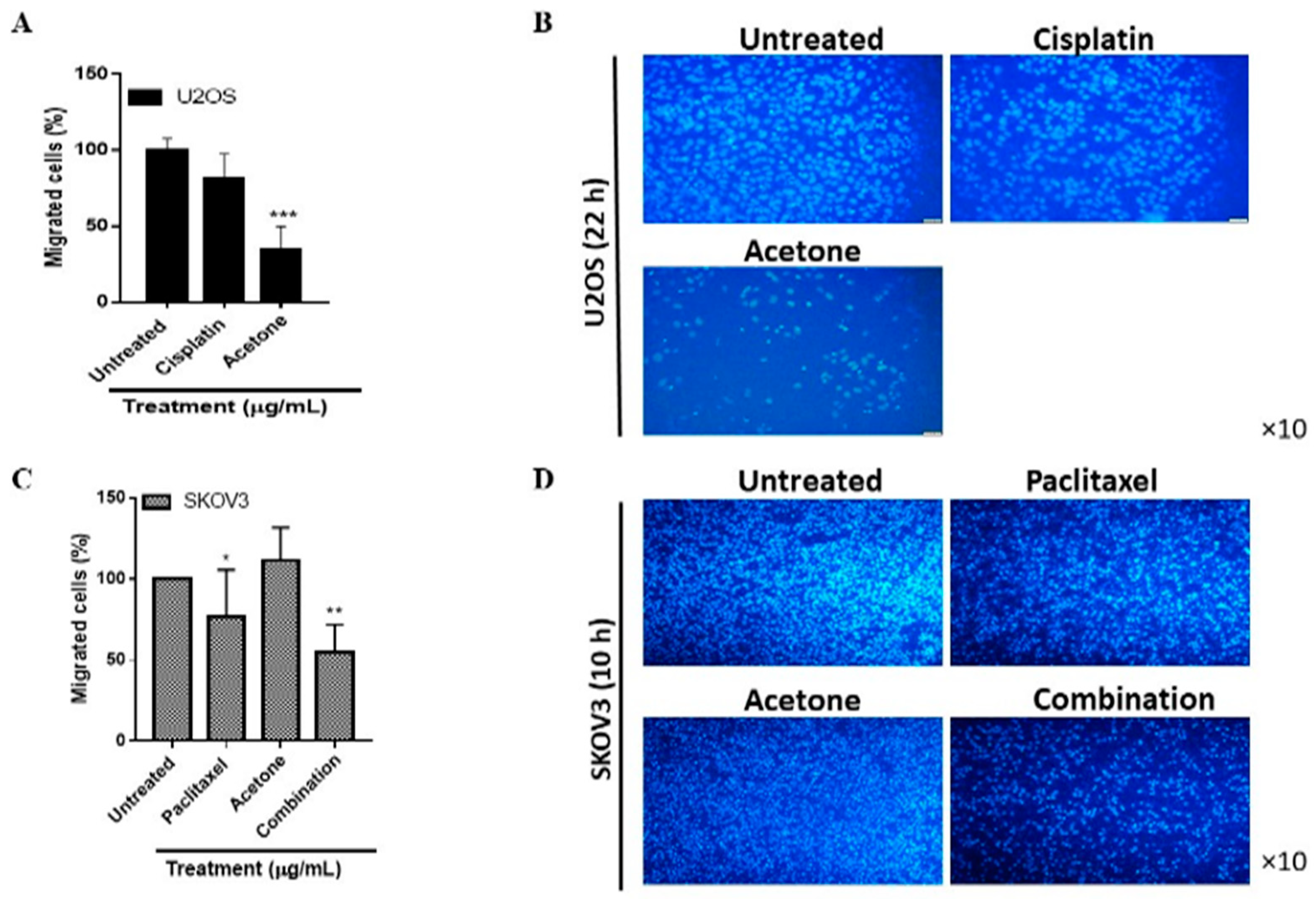
3.6. S. triloba Acetone Extract and the Combination Treatment Respectively Decrease U2OS and SKOV3 Cell Migration
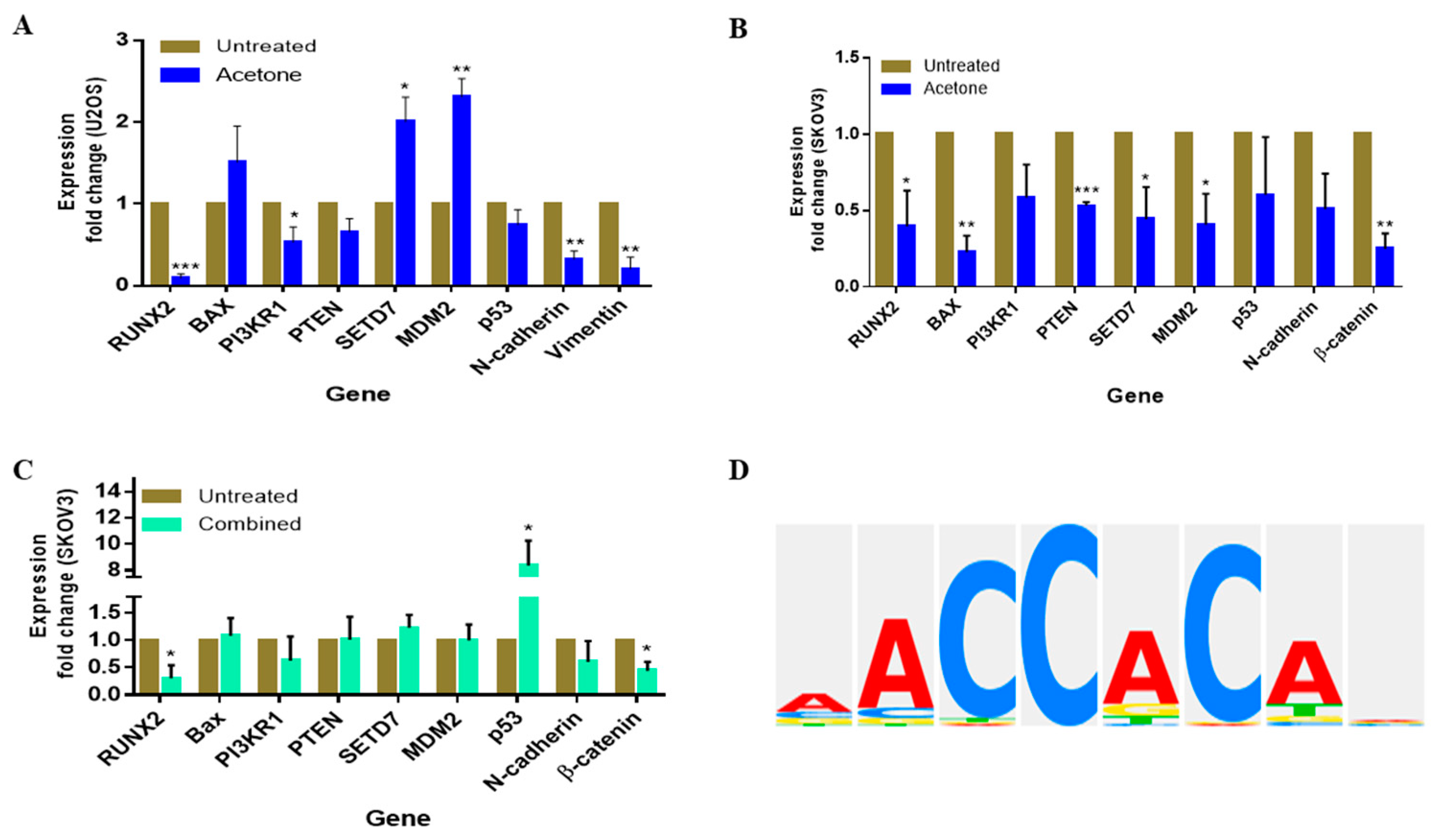
3.7. S. triloba Acetone Extract and Its Combination with Paclitaxel Affect the Steady-State mRNA Expression of Oncogenic Targets in Both U2OS and SKOV3 Cells
3.8. Identification of RUNX2 Transcription Factor Binding Sites within the MDM2 Gene
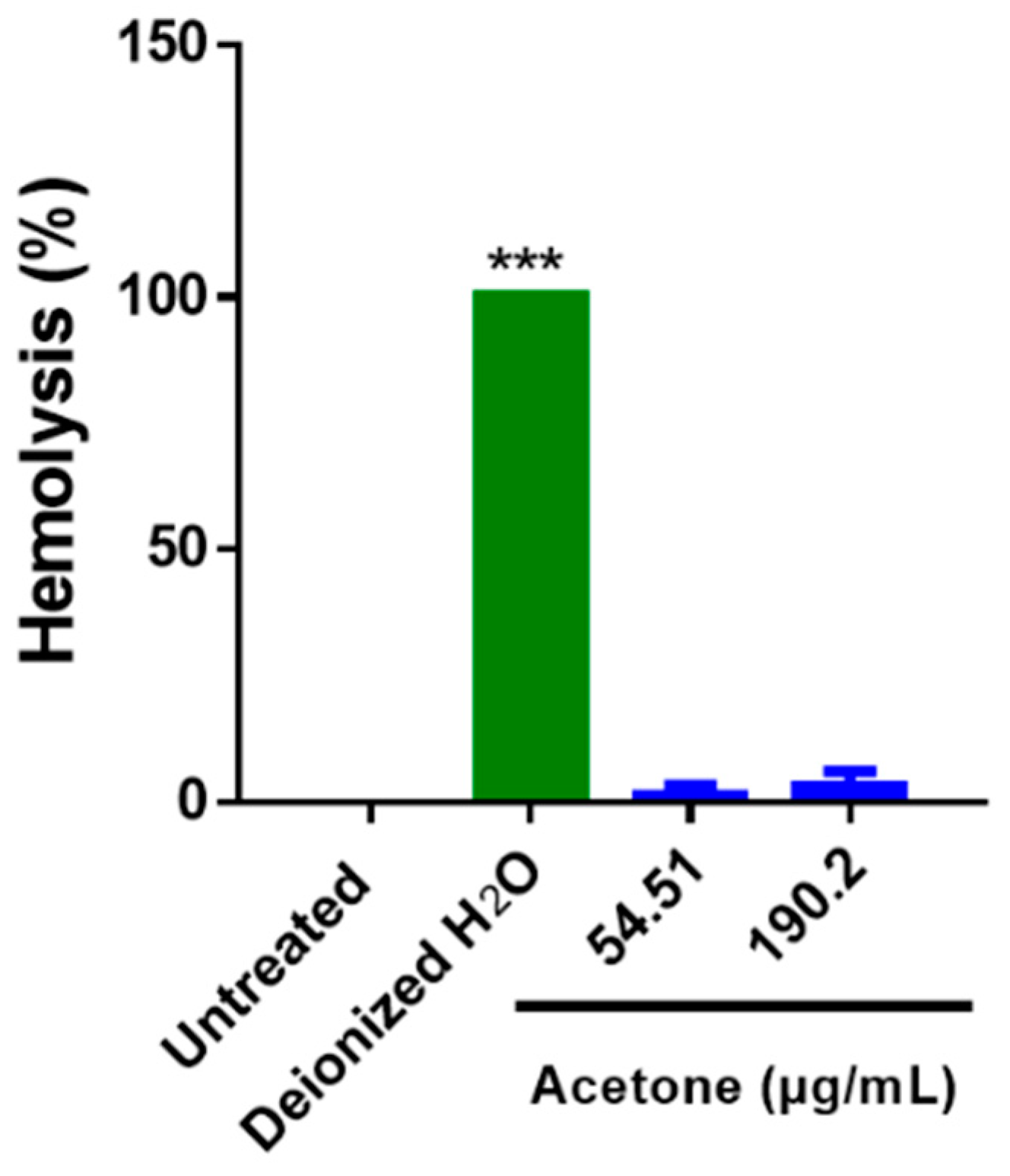
3.9. S. triloba Acetone Extract Does Not Induce Hemolysis in Human Erythrocytes
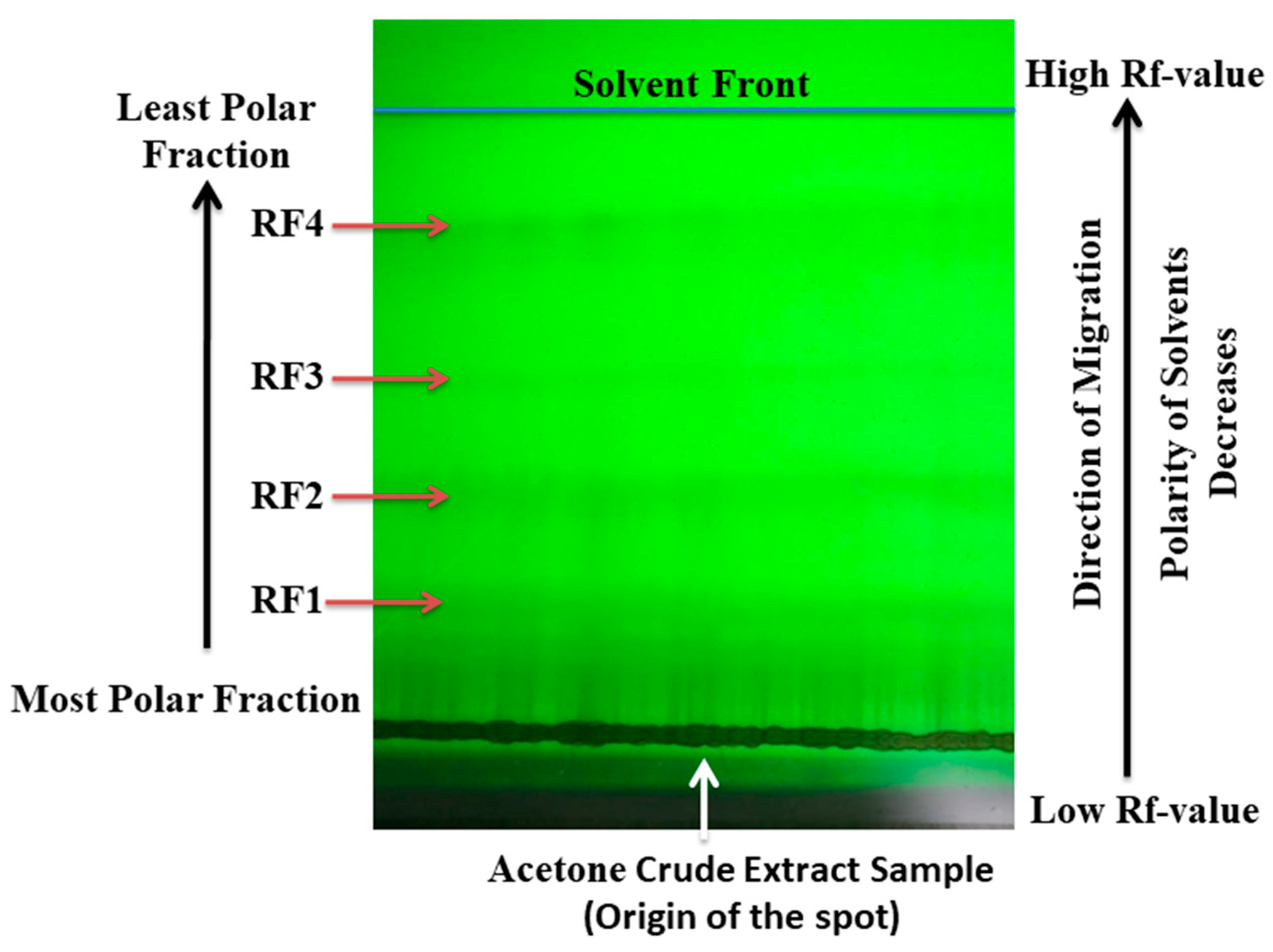
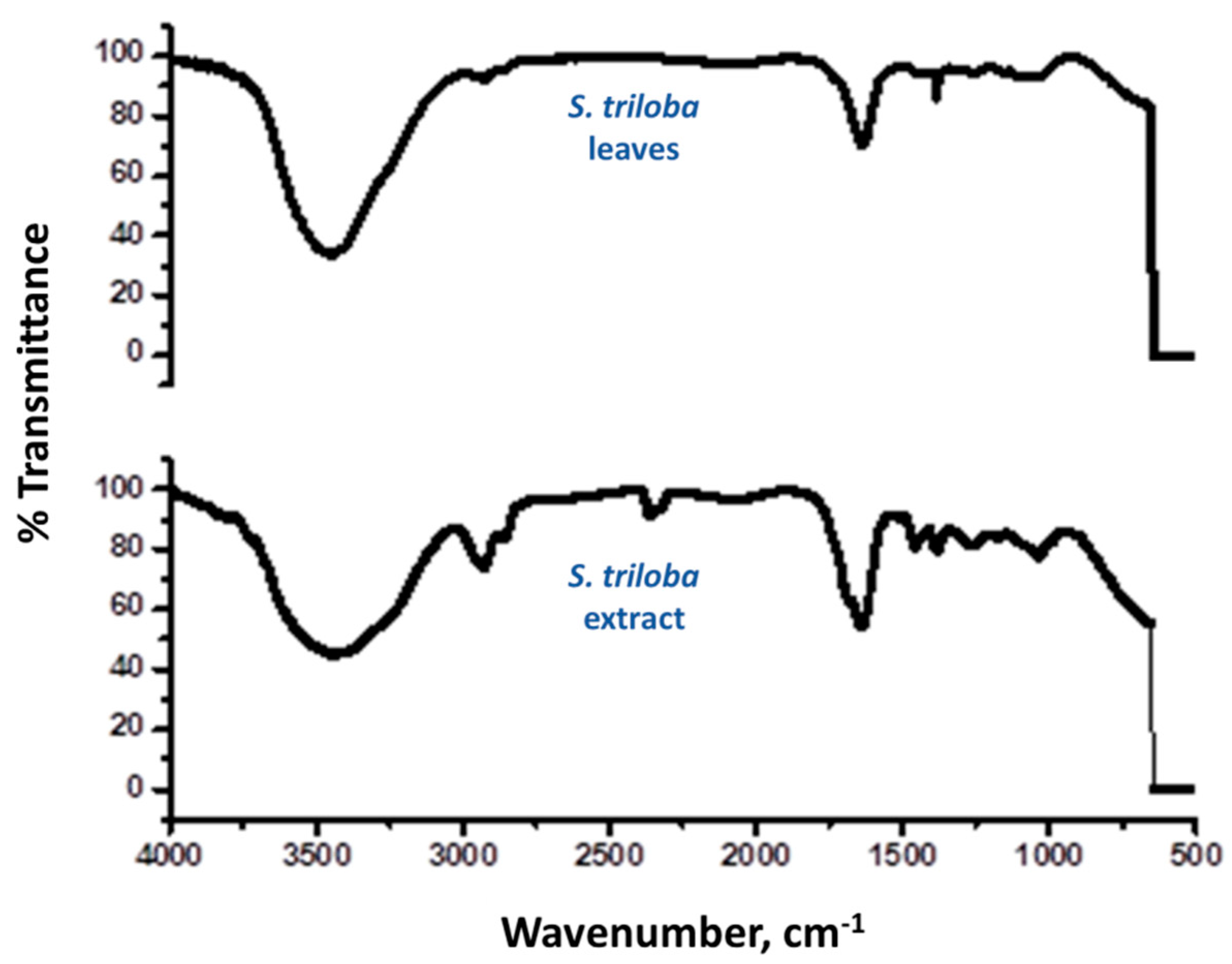
3.10. TLC-UV Fractionation and FTIR Measurements of Crude S. triloba Acetone Extract
4. Discussion
4.1. S. triloba Acetone Extract Significantly Reduces U2OS and SKOV3 Cell Viability
4.2. Selectivity Index of S. triloba for U2OS and SKOV3 Cells
4.3. The Combination of S. triloba Acetone Extract and Paclitaxel Exhibits Strong Synergism in SKOV3 Cells
4.4. S. triloba Acetone Extract and Its Combination with Paclitaxel Increase the Percentage of Non-Viable Cells in SKOV3 Cells
4.5. S. triloba and Its Combination with Paclitaxel Respectively Display Anti-Migratory Properties in U2OS and SKOV3 Cells
4.6. S. triloba and Its Combination with Paclitaxel Modulate the Steady-State mRNA Expression of Genes Involved in Cell Proliferation and Migration
4.7. S. triloba Acetone Extract Does Not Induce Hemolysis in Human Erythrocytes
4.8. TLC-UV Fractionation of Crude S. triloba Acetone Extract
4.9. FTIR of S. triloba Leaves and Its Crude Acetone Extract
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global Cancer Statistics 2020: GLOBOCAN Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries. CA Cancer J. Clin. 2021, 71, 209–249. [Google Scholar] [CrossRef] [PubMed]
- IARC, T.I.A. for R. on C. Latest Global Cancer Data: Cancer Burden Rises to 19.3 Million New Cases. Available online: https://www.iarc.who.int/wp-content/uploads/2020/12/pr292_E.pdf (accessed on 9 September 2021).
- Jaffe, N.; Bruland, O.S.; Bielack, S.S. (Eds.) Pediatric and Adolescent Osteosarcoma; Cancer treatment and research; Springer: New York, NY, USA, 2009; ISBN 978-1-4419-0283-2. [Google Scholar]
- Eleutério, S.J.P.; Senerchia, A.A.; Almeida, M.T.; Costa, C.M.D.; Lustosa, D.; Calheiros, L.M.; Barreto, J.H.S.; Brunetto, A.L.; Macedo, C.R.P.D.; Petrilli, A.S. Osteosarcoma in Patients Younger than 12 Years Old without Metastases Have Similar Prognosis as Adolescent and Young Adults: Osteosarcoma in Children and AYA. Pediatr. Blood Cancer 2015, 62, 1209–1213. [Google Scholar] [CrossRef] [PubMed]
- American Cancer Society Key Statistics for Osteosarcoma. Available online: https://www.cancer.org/cancer/osteosarcoma/about/key-statistics.html (accessed on 19 May 2020).
- Taran, S.; Taran, R.; Malipatil, N. Pediatric Osteosarcoma: An Updated Review. Indian J. Med. Paediatr. Oncol. 2017, 38, 33. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lauvrak, S.U.; Munthe, E.; Kresse, S.H.; Stratford, E.W.; Namløs, H.M.; Meza-Zepeda, L.A.; Myklebost, O. Functional Characterisation of Osteosarcoma Cell Lines and Identification of MRNAs and MiRNAs Associated with Aggressive Cancer Phenotypes. Br. J. Cancer 2013, 109, 2228–2236. [Google Scholar] [CrossRef] [PubMed]
- Momenimovahed, Z.; Tiznobaik, A.; Taheri, S.; Salehiniya, H. Ovarian Cancer in the World: Epidemiology and Risk Factors. Int. J. Womens Health 2019, 11, 287–299. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- WOCC World Ovarian Cancer Coalition. Available online: https://worldovariancancercoalition.org/about-ovarian-cancer/key-stats/ (accessed on 1 September 2020).
- Shaw, T.J.; Senterman, M.K.; Dawson, K.; Crane, C.A.; Vanderhyden, B.C. Characterization of Intraperitoneal, Orthotopic, and Metastatic Xenograft Models of Human Ovarian Cancer. Mol. Ther. 2004, 10, 1032–1042. [Google Scholar] [CrossRef]
- ATCC U-2 OS (ATCC® HTB-96TM). Available online: https://www.lgcstandards-atcc.org/Products/All/HTB-96.aspx?geo_country=eg#characteristics (accessed on 16 April 2020).
- Allan, L.A.; Fried, M. P53-Dependent Apoptosis or Growth Arrest Induced by Different Forms of Radiation in U2OS Cells: P21WAF1/CIP1 Repression in UV Induced Apoptosis. Oncogene 1999, 18, 5403–5412. [Google Scholar] [CrossRef] [Green Version]
- ATCC, A.T.C.C. SK-OV-3 [SKOV-3; SKOV3]. Available online: https://www.atcc.org/products/htb-77 (accessed on 24 August 2021).
- Jelovac, D.; Armstrong, D.K. Recent Progress in the Diagnosis and Treatment of Ovarian Cancer. CA. Cancer J. Clin. 2011, 61, 183–203. [Google Scholar] [CrossRef]
- Cragg, G.M.; Newman, D.J. Plants as a Source of Anti-Cancer Agents. J. Ethnopharmacol. 2005, 100, 72–79. [Google Scholar] [CrossRef] [Green Version]
- Ahl, H.S.-A.; Hussein, M.S.; Gendy, A.S.H.; Tkachenko, K.G. Quality of Sage (Salvia officinalis L.) Essential Oil Grown in Egypt. Int. J. Plant Sci. Ecol. 2015, 1, 119–123. [Google Scholar]
- Shahidi, F. (Ed.) Handbook of Antioxidants for Food Preservation; Woodhead Publishing series in food science, technology and nutrition; Elsevier: Cambridge, UK; Waltham, MA, USA, 2015; ISBN 978-1-78242-089-7. [Google Scholar]
- Gali-Muhtasib, H. Anticancer and Medicinal Properties of Essential Oil and Extracts of East Mediterranean Sage (Salvia triloba). In Advances in Phytomedicine; Elsevier: Amsterdam, The Netherlands; Boston, MA, USA, 2006; Volume 2, pp. 169–180. ISBN 978-0-444-51619-0. [Google Scholar]
- The British Pharmaceutical Codex, 1923: An Imperial Dispensatory for the Use of Medical Practitioners and Pharmacists. Nature 1923, 112, 859. [CrossRef]
- Council of Europe; European Pharmacopoeia Commission; European Directorate for the Quality of Medicines & Healthcare. European Pharmacopoeia, 7th ed.; Council of Europe, European Directorate for the Quality of Medicines and Healthcare: Strasbourg, France, 2010; ISBN 978-92-871-6700-2. [Google Scholar]
- Lopresti, A.L. Salvia (Sage): A Review of Its Potential Cognitive-Enhancing and Protective Effects. Drugs RD 2017, 17, 53–64. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Topçu, G.; Öztürk, M.; Kuşman, T. Terpenoids, essential oil composition, fatty acid profile, and biological activities of Anatolian Salvia fruticosa Mill. Turk. J. Chem. 2013, 37, 619–632. [Google Scholar] [CrossRef]
- Atmaca, H.; Bozkurt, E. Apoptotic and Anti-Angiogenic Effects of Salvia Triloba Extract in Prostate Cancer Cell Lines. Tumor Biol. 2016, 37, 3639–3646. [Google Scholar] [CrossRef]
- Abu-Dahab, R.; Abdallah, M.R.; Kasabri, V.; Mhaidat, N.M.; Afifi, F.U. Mechanistic Studies of Antiproliferative Effects of Salvia Triloba and Salvia Dominica (Lamiaceae) on Breast Cancer Cell Lines (MCF7 and T47D). Z. Fur Nat. C 2014, 69, 443–451. [Google Scholar] [CrossRef]
- Al-Kalaldeh, J.Z.; Abu-Dahab, R.; Afifi, F.U. Volatile Oil Composition and Antiproliferative Activity of Laurus Nobilis, Origanum Syriacum, Origanum Vulgare, and Salvia Triloba against Human Breast Adenocarcinoma Cells. Nutr. Res. 2010, 30, 271–278. [Google Scholar] [CrossRef]
- Zihlif, M.; Afifi, F.; Abu-Dahab, R.; Abdul Majid, A.M.S.; Somrain, H.; Saleh, M.M.; Nassar, Z.D.; Naffa, R. The Antiangiogenic Activities of Ethanolic Crude Extracts of Four Salvia Species. BMC Complement. Altern. Med. 2013, 13, 358. [Google Scholar] [CrossRef] [Green Version]
- Danin, A.; Fragman-Sapir, O. Flora of Israel Online. Available online: https://flora.org.il/en/plants/salfru/ (accessed on 2 September 2021).
- Kim, D.; Lee, C.Y. Extraction and Isolation of Polyphenolics. Curr. Protoc. Food Anal. Chem. 2002, 6, I1.2.1–I1.2.12. [Google Scholar] [CrossRef]
- Ponten, J.; Saksela, E. Two Established in Vitro Cell Lines from Human Mesenchymal Tumours. Int. J. Cancer 1967, 2, 434–447. [Google Scholar] [CrossRef]
- Fogh, J.; Fogh, J.M.; Orfeo, T. One Hundred and Twenty-Seven Cultured Human Tumor Cell Lines Producing Tumors in Nude Mice23. JNCI J. Natl. Cancer Inst. 1977, 59, 221–226. [Google Scholar] [CrossRef]
- ATCC 293 [HEK-293]. Available online: https://www.atcc.org/products/crl-1573 (accessed on 25 October 2022).
- Riss, T.L.; Moravec, R.A.; Niles, A.L.; Duellman, S.; Benink, H.A.; Worzella, T.J.; Minor, L. Cell Viability Assays. In Assay Guidance Manual; Sittampalam, G.S., Coussens, N.P., Brimacombe, K., Grossman, A., Arkin, M., Auld, D., Austin, C., Baell, J., Bejcek, B., Caaveiro, J.M.M., et al., Eds.; Eli Lilly & Company and the National Center for Advancing Translational Sciences: Bethesda, MD, USA, 2004. [Google Scholar]
- Graphpad Software Inc. GraphPad Prism Version 6.01 for Windows, GraphPad Software, La Jolla California USA. Available online: www.Graphpad.com (accessed on 24 July 2020).
- Indrayanto, G.; Putra, G.S.; Suhud, F. Validation of In-Vitro Bioassay Methods: Application in Herbal Drug Research. Profiles Drug Subst. Excip. Relat. Methodol. 2021, 46, 273–307. [Google Scholar]
- Sahu, S.C.; Hayes, A.W. Toxicity of Nanomaterials Found in Human Environment: A Literature Review. Toxicol. Res. Appl. 2017, 1, 2397847317726352. [Google Scholar] [CrossRef]
- Peng, H.; Zhang, X.; Wei, Y.; Liu, W.; Li, S.; Yu, G.; Fu, X.; Cao, T.; Deng, X. Cytotoxicity of Silver Nanoparticles in Human Embryonic Stem Cell-Derived Fibroblasts and an L-929 Cell Line. J. Nanomater. 2012, 2012, 160145. [Google Scholar] [CrossRef] [Green Version]
- Sooklert, K.; Chattong, S.; Manotham, K.; Boonwong, C.; Klaharn, I.; Jindatip, D.; Sereemaspun, A. Cytoprotective Effect of Glutaraldehyde Erythropoietin on HEK293 Kidney Cells after Silver Nanoparticle Exposure. Int. J. Nanomed. 2016, 11, 597. [Google Scholar]
- Chou, T.-C. Drug Combination Studies and Their Synergy Quantification Using the Chou-Talalay Method. Cancer Res. 2010, 70, 440–446. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chou, T.-C. Theoretical Basis, Experimental Design, and Computerized Simulation of Synergism and Antagonism in Drug Combination Studies. Pharmacol. Rev. 2006, 58, 621–681. [Google Scholar] [CrossRef]
- Chou, T.-C.; Martin, N. CompuSyn for Drug Combinations User’s Guide; ComboSyn, Inc.: Paramus, NJ, USA, 2005. [Google Scholar]
- Strober, W. Trypan Blue Exclusion Test of Cell Viability. In Current Protocols in Immunology; Coligan, J.E., Bierer, B.E., Margulies, D.H., Shevach, E.M., Strober, W., Eds.; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2001; ISBN 978-0-471-14273-7. [Google Scholar]
- Boleman, A.I.; Tănasie, G.; Găluşcan, A.; Cristea, M.I.; Bojin, F.M.; Panaitescu, C.; Păunescu, V. Studies Regarding the In Vitro Wound Healing Potential of Mouse Dental Pulp Stem-Like Progenitor Cells. Biotechnol. Biotechnol. Equip. 2012, 26, 2781–2785. [Google Scholar] [CrossRef] [Green Version]
- Ganger, M.T.; Dietz, G.D.; Ewing, S.J. A Common Base Method for Analysis of QPCR Data and the Application of Simple Blocking in QPCR Experiments. BMC Bioinform. 2017, 18, 534. [Google Scholar] [CrossRef]
- Matys, V. TRANSFAC(R) and Its Module TRANSCompel(R): Transcriptional Gene Regulation in Eukaryotes. Nucleic Acids Res. 2006, 34, D108–D110. [Google Scholar] [CrossRef] [Green Version]
- Malagoli, D. A Full-Length Protocol to Test Hemolytic Activity of Palytoxin on Human Erythrocytes. Invertebr. Surviv. J. 2007, 4, 92–94. [Google Scholar]
- Nichols, L. Visualizing TLC Plates. Available online: https://chem.libretexts.org/Bookshelves/Organic_Chemistry/Book%3A_Organic_Chemistry_Lab_Techniques_(Nichols)/02%3A_Chromatography/2.03%3A_Thin_Layer_Chromatography_(TLC)/2.3F%3A_Visualizing_TLC_Plates (accessed on 8 September 2021).
- Bryne, J.C. E-MTAB-1828-ChIP-Seq of Human (Un)Differentiated Mesenchymal Stem Cells and Normal Osteoblasts with Antibodies against Various Histone H3 Modifications and RUNX2 to Study the Regulatory Landscape of Osteogenic Differentiation. Available online: https://www.ebi.ac.uk/arrayexpress/experiments/E-MTAB-1828/ (accessed on 1 September 2020).
- Liu, Z.; Merkurjev, D.; Rosenfeld, M. Enhancer Activation Requires Trans-Recruitment of a Mega Transcription Factor Complex (ChIP-Seq). Available online: https://www.ncbi.nlm.nih.gov/geo/query/acc.cgi?acc=GSE60270 (accessed on 1 September 2020).
- Little, G.; Noushmehr, H.; Baniwal, S.; Berman, B.; Coetzee, G.; Frenkel, B. Genome-Wide Runx2 Occupancy in Prostate Cancer Cells Suggests a Role in Regulating Secretion. Available online: https://www.ncbi.nlm.nih.gov/geo/query/acc.cgi?acc=GSE33889 (accessed on 1 September 2020).
- Naziruddin, M.A.; Kian, L.K.; Jawaid, M.; Fouad, H.; Sanny, M.; Braganca, R.M. Sage Biomass Powders by Supercritical Fluid Extraction and Hydro-Distillation Techniques: A Comparative Study of Biological and Chemical Properties. Biomass Convers. Biorefin. 2021. [Google Scholar] [CrossRef]
- Essa, H.L.; Abdelfattah, M.S.; Marzouk, A.S.; Guirguis, H.A.; El-Sayed, M.M.H. Nano-Formulations of Copper Species Coated with Sulfated Polysaccharide Extracts and Assessment of Their Phytotoxicity on Wheat (Triticum aestivum L.) Seedlings in Seed Germination, Foliar and Soil Applications. Appl. Sci. 2020, 10, 6302. [Google Scholar] [CrossRef]
- Abou El Azm, N.; Fleita, D.; Rifaat, D.; Mpingirika, E.Z.; Amleh, A.; El-Sayed, M.M.H. Production of Bioactive Compounds from the Sulfated Polysaccharides Extracts of Ulva Lactuca: Post-Extraction Enzymatic Hydrolysis Followed by Ion-Exchange Chromatographic Fractionation. Molecules 2019, 24, 2132. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fleita, D.; El-Sayed, M.; Rifaat, D. Evaluation of the Antioxidant Activity of Enzymatically-Hydrolyzed Sulfated Polysaccharides Extracted from Red Algae; Pterocladia capillacea. LWT Food Sci. Technol. 2015, 63, 1236–1244. [Google Scholar] [CrossRef]
- Essa, H.L.; Guirguis, H.A.; El-Sayed, M.M.H.; Rifaat, D.; Abdelfattah, M.S. Ultrasonically-Extracted Marine Polysaccharides as Potential Green Antioxidant Alternatives. In Proceedings of the 1st International Electronic Conference on Applied Sciences, Basel, Switzerland, 10–30 November 2020; p. 23. [Google Scholar]
- Essa, H.L.; Abdelfattah, M.S.; Marzouk, A.S.; Shedeed, Z.; Guirguis, H.A.; El-Sayed, M.M.H. Biogenic Copper Nanoparticles from Avicennia Marina Leaves: Impact on Seed Germination, Detoxification Enzymes, Chlorophyll Content and Uptake by Wheat Seedlings. PLoS ONE 2021, 16, e0249764. [Google Scholar] [CrossRef] [PubMed]
- Abu-Dahab, R.; Afifi, F.; Kasabri, V.; Majdalawi, L.; Naffa, R. Comparison of the Antiproliferative Activity of Crude Ethanol Extracts of Nine Salvia Species Grown in Jordan against Breast Cancer Cell Line Models. Pharmacogn. Mag. 2012, 8, 319. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Georgiev, V.; Pavlov, A. (Eds.) Salvia Biotechnology, 1st ed.; Springer International Publishing: Cham, Switzerland, 2017; ISBN 978-3-319-73900-7. [Google Scholar]
- Prinz, H. Hill Coefficients, Dose–Response Curves and Allosteric Mechanisms. J. Chem. Biol. 2010, 3, 37–44. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weiss, J.N. The Hill Equation Revisited: Uses and Misuses. FASEB J. 1997, 11, 835–841. [Google Scholar] [CrossRef]
- Krzywik, J.; Mozga, W.; Aminpour, M.; Janczak, J.; Maj, E.; Wietrzyk, J.; Tuszyński, J.A.; Huczyński, A. Synthesis, Antiproliferative Activity and Molecular Docking Studies of Novel Doubly Modified Colchicine Amides and Sulfonamides as Anticancer Agents. Molecules 2020, 25, 1789. [Google Scholar] [CrossRef] [Green Version]
- Lica, J.J.; Wieczór, M.; Grabe, G.J.; Heldt, M.; Jancz, M.; Misiak, M.; Gucwa, K.; Brankiewicz, W.; Maciejewska, N.; Stupak, A.; et al. Effective Drug Concentration and Selectivity Depends on Fraction of Primitive Cells. Int. J. Mol. Sci. 2021, 22, 4931. [Google Scholar] [CrossRef]
- Yap, T.A.; Omlin, A.; de Bono, J.S. Development of Therapeutic Combinations Targeting Major Cancer Signaling Pathways. J. Clin. Oncol. 2013, 31, 1592–1605. [Google Scholar] [CrossRef]
- Albain, K.S.; Nag, S.M.; Calderillo-Ruiz, G.; Jordaan, J.P.; Llombart, A.C.; Pluzanska, A.; Rolski, J.; Melemed, A.S.; Reyes-Vidal, J.M.; Sekhon, J.S.; et al. Gemcitabine plus Paclitaxel versus Paclitaxel Monotherapy in Patients with Metastatic Breast Cancer and Prior Anthracycline Treatment. J. Clin. Oncol. 2008, 26, 3950–3957. [Google Scholar] [CrossRef] [PubMed]
- Bayat Mokhtari, R.; Homayouni, T.S.; Baluch, N.; Morgatskaya, E.; Kumar, S.; Das, B.; Yeger, H. Combination Therapy in Combating Cancer. Oncotarget 2017, 8, 38022–38043. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mohan, L. Plant-Based Drugs as an Adjuvant to Cancer Chemotherapy. In Alternative Medicine [Working Title]; IntechOpen: London, UK, 2020. [Google Scholar]
- Tayeh, Z.; Ofir, R. Asteriscus Graveolens Extract in Combination with Cisplatin/Etoposide/Doxorubicin Suppresses Lymphoma Cell Growth through Induction of Caspase-3 Dependent Apoptosis. Int. J. Mol. Sci. 2018, 19, 2219. [Google Scholar] [CrossRef]
- Liu, L.; Fan, J.; Ai, G.; Liu, J.; Luo, N.; Li, C.; Cheng, Z. Berberine in Combination with Cisplatin Induces Necroptosis and Apoptosis in Ovarian Cancer Cells. Biol. Res. 2019, 52, 37. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Montopoli, M.; Ragazzi, E.; Froldi, G.; Caparrotta, L. Cell-Cycle Inhibition and Apoptosis Induced by Curcumin and Cisplatin or Oxaliplatin in Human Ovarian Carcinoma Cells. Cell Prolif. 2009, 42, 195–206. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chan, M.M.; Soprano, K.J.; Weinstein, K.; Fong, D. Epigallocatechin-3-Gallate Delivers Hydrogen Peroxide to Induce Death of Ovarian Cancer Cells and Enhances Their Cisplatin Susceptibility. J. Cell. Physiol. 2006, 207, 389–396. [Google Scholar] [CrossRef] [PubMed]
- Cui, Q.; Bian, R.; Xu, F.; Li, Q.; Wang, W.; Bian, Q. New Molecular Entities and Structure–Activity Relationships of Drugs Designed by the Natural Product Derivatization Method from 2010 to 2018. In Studies in Natural Products Chemistry; Elsevier: Amsterdam, The Netherlands, 2021; Volume 69, pp. 371–415. ISBN 978-0-12-819487-4. [Google Scholar]
- Lin, S.-R.; Chang, C.-H.; Hsu, C.-F.; Tsai, M.-J.; Cheng, H.; Leong, M.K.; Sung, P.-J.; Chen, J.-C.; Weng, C.-F. Natural Compounds as Potential Adjuvants to Cancer Therapy: Preclinical Evidence. Br. J. Pharmacol. 2020, 177, 1409–1423. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Chen, X.; Gueydan, C.; Han, J. Plasma Membrane Changes during Programmed Cell Deaths. Cell Res. 2018, 28, 9–21. [Google Scholar] [CrossRef] [Green Version]
- Green, D.R.; Llambi, F. Cell Death Signaling. Cold Spring Harb. Perspect. Biol. 2015, 7, a006080. [Google Scholar] [CrossRef] [Green Version]
- Tahtamouni, L.; Ahram, M.; Koblinski, J.; Rolfo, C. Molecular Regulation of Cancer Cell Migration, Invasion, and Metastasis. Anal. Cell. Pathol. 2019, 2019, 1356508. [Google Scholar] [CrossRef] [Green Version]
- Anantharaju, P.G.; Gowda, P.C.; Vimalambike, M.G.; Madhunapantula, S.V. An Overview on the Role of Dietary Phenolics for the Treatment of Cancers. Nutr. J. 2016, 15, 99. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abreu, M.E.; Müller, M.; Alegre, L.; Munné-Bosch, S. Phenolic Diterpene and α-Tocopherol Contents in Leaf Extracts of 60 Salvia Species. J. Sci. Food Agric. 2008, 88, 2648–2653. [Google Scholar] [CrossRef]
- Barni, M.V.; Carlini, M.J.; Cafferata, E.G.; Puricelli, L.; Moreno, S. Carnosic Acid Inhibits the Proliferation and Migration Capacity of Human Colorectal Cancer Cells. Oncol. Rep. 2012, 27, 1041–1048. [Google Scholar] [CrossRef] [PubMed]
- Koutsoulas, A.; Čarnecká, M.; Slanina, J.; Tóth, J.; Slaninová, I. Characterization of Phenolic Compounds and Antiproliferative Effects of Salvia Pomifera and Salvia Fruticosa Extracts. Molecules 2019, 24, 2921. [Google Scholar] [CrossRef] [Green Version]
- Xiaomeng, F.; Lei, L.; Jinghong, A.; Juan, J.; Qi, Y.; Dandan, Y. Treatment with β-Elemene Combined with Paclitaxel Inhibits Growth, Migration, and Invasion and Induces Apoptosis of Ovarian Cancer Cells by Activation of STAT-NF-ΚB Pathway. Braz. J. Med. Biol. Res. 2020, 53, e8885. [Google Scholar] [CrossRef]
- Martin, J.W.; Zielenska, M.; Stein, G.S.; Wijnen, A.J.; Squire, J.A. The Role of RUNX2 in Osteosarcoma Oncogenesis. Sarcoma 2011, 2011, 282745. [Google Scholar] [CrossRef] [Green Version]
- Li, W.; Xu, S.; Lin, S.; Zhao, W. Overexpression of Runt-Related Transcription Factor-2 Is Associated with Advanced Tumor Progression and Poor Prognosis in Epithelial Ovarian Cancer. J. Biomed. Biotechnol. 2012, 2012, 456534. [Google Scholar] [CrossRef] [Green Version]
- Park, E.-S.; Lind, A.-K.; Dahm-Kähler, P.; Brännström, M.; Carletti, M.Z.; Christenson, L.K.; Curry, T.E.; Jo, M. RUNX2 Transcription Factor Regulates Gene Expression in Luteinizing Granulosa Cells of Rat Ovaries. Mol. Endocrinol. 2010, 24, 846–858. [Google Scholar] [CrossRef] [Green Version]
- Sionov, R.V.; Hayon, I.L.; Haupt, Y. The Regulation of P53 Growth Suppression. In Madame Curie Bioscience Database [Internet]; Landes Bioscience: Austin, TX, USA, 2000. [Google Scholar]
- Basu, A. The Relationship between BcI2, Bax and P53: Consequences for Cell Cycle Progression and Cell Death. Mol. Hum. Reprod. 1998, 4, 1099–1109. [Google Scholar] [CrossRef]
- Chipuk, J.E. Direct Activation of Bax by P53 Mediates Mitochondrial Membrane Permeabilization and Apoptosis. Science 2004, 303, 1010–1014. [Google Scholar] [CrossRef] [Green Version]
- Yu, J.; Zhang, L. PUMA, a Potent Killer with or without P53. Oncogene 2008, 27, S71–S83. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, M.; Zheng, J.; Nussinov, R.; Ma, B. Release of Cytochrome C from Bax Pores at the Mitochondrial Membrane. Sci. Rep. 2017, 7, 2635. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schuler, M.; Bossy-Wetzel, E.; Goldstein, J.C.; Fitzgerald, P.; Green, D.R. P53 Induces Apoptosis by Caspase Activation through Mitochondrial Cytochrome c Release. J. Biol. Chem. 2000, 275, 7337–7342. [Google Scholar] [CrossRef] [PubMed]
- Batista, I.A.A.; Helguero, L.A. Biological Processes and Signal Transduction Pathways Regulated by the Protein Methyltransferase SETD7 and Their Significance in Cancer. Signal Transduct. Target. Ther. 2018, 3, 19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Campaner, S.; Spreafico, F.; Burgold, T.; Doni, M.; Rosato, U.; Amati, B.; Testa, G. The Methyltransferase Set7/9 (Setd7) Is Dispensable for the P53-Mediated DNA Damage Response In Vivo. Mol. Cell 2011, 43, 681–688. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Yu, X.-H.; Yan, Y.-G.; Wang, C.; Wang, W.-J. PI3K/Akt Signaling in Osteosarcoma. Clin. Chim. Acta 2015, 444, 182–192. [Google Scholar] [CrossRef] [PubMed]
- Gasparri, M.L.; Bardhi, E.; Ruscito, I.; Papadia, A.; Farooqi, A.A.; Marchetti, C.; Bogani, G.; Ceccacci, I.; Mueller, M.D.; Benedetti Panici, P. PI3K/AKT/MTOR Pathway in Ovarian Cancer Treatment: Are We on the Right Track? Geburtshilfe Frauenheilkd. 2017, 77, 1095–1103. [Google Scholar] [CrossRef] [Green Version]
- Valenta, T.; Hausmann, G.; Basler, K. The Many Faces and Functions of β-Catenin: β-Catenin: A Life by, beyond, and against the Wnt Canon. EMBO J. 2012, 31, 2714–2736. [Google Scholar] [CrossRef] [Green Version]
- Lustig, B.; Behrens, J. The Wnt Signaling Pathway and Its Role in Tumor Development. J. Cancer Res. Clin. Oncol. 2003, 129, 199–221. [Google Scholar] [CrossRef]
- MacDonald, B.T.; Tamai, K.; He, X. Wnt/β-Catenin Signaling: Components, Mechanisms, and Diseases. Dev. Cell 2009, 17, 9–26. [Google Scholar] [CrossRef] [Green Version]
- Hadjihannas, M.V.; Bruckner, M.; Jerchow, B.; Birchmeier, W.; Dietmaier, W.; Behrens, J. Aberrant Wnt/Beta-Catenin Signaling Can Induce Chromosomal Instability in Colon Cancer. Proc. Natl. Acad. Sci. USA 2006, 103, 10747–10752. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mohinta, S. Wnt Pathway and Breast Cancer. Front. Biosci. 2007, 12, 4020. [Google Scholar] [CrossRef] [Green Version]
- Khalaf, A.M.; Fuentes, D.; Morshid, A.I.; Burke, M.R.; Kaseb, A.O.; Hassan, M.; Hazle, J.D.; Elsayes, K.M. Role of Wnt/β-Catenin Signaling in Hepatocellular Carcinoma, Pathogenesis, and Clinical Significance. J. Hepatocell. Carcinoma 2018, 5, 61–73. [Google Scholar] [CrossRef] [PubMed]
- Kleinerman, E.S. Current Advances in Osteosarcoma; Springer: New York, NY, USA, 2014; ISBN 978-3-319-04842-0. [Google Scholar]
- Arend, R.C.; Londoño-Joshi, A.I.; Straughn, J.M.; Buchsbaum, D.J. The Wnt/β-Catenin Pathway in Ovarian Cancer: A Review. Gynecol. Oncol. 2013, 131, 772–779. [Google Scholar] [CrossRef] [PubMed]
- Wu, R.; Zhai, Y.; Fearon, E.R.; Cho, K.R. Diverse Mechanisms of Beta-Catenin Deregulation in Ovarian Endometrioid Adenocarcinomas. Cancer Res. 2001, 61, 8247–8255. [Google Scholar] [PubMed]
- Blagodatski, A.; Klimenko, A.; Jia, L.; Katanaev, V.L. Small Molecule Wnt Pathway Modulators from Natural Sources: History, State of the Art and Perspectives. Cells 2020, 9, 589. [Google Scholar] [CrossRef] [Green Version]
- Fu, J.; Wang, S.; Lu, H.; Ma, J.; Ke, X.; Liu, T.; Luo, Y. In Vitro Inhibitory Effects of Terpenoids from Chloranthus Multistachys on Epithelial–Mesenchymal Transition via down-Regulation of Runx2 Activation in Human Breast Cancer. Phytomedicine 2015, 22, 165–172. [Google Scholar] [CrossRef]
- Ma, J.; Lu, H.; Wang, S.; Chen, B.; Liu, Z.; Ke, X.; Liu, T.; Fu, J. The Anthraquinone Derivative Emodin Inhibits Angiogenesis and Metastasis through Downregulating Runx2 Activity in Breast Cancer. Int. J. Oncol. 2015, 46, 1619–1628. [Google Scholar] [CrossRef] [Green Version]
- Riaz, M.; Ashfaq, U.A.; Qasim, M.; Yasmeen, E.; Ul Qamar, M.T.; Anwar, F. Screening of Medicinal Plant Phytochemicals as Natural Antagonists of P53–MDM2 Interaction to Reactivate P53 Functioning. Anti-Cancer Drugs 2017, 28, 1032–1038. [Google Scholar] [CrossRef]
- Sukumari-Ramesh, S.; Bentley, J.N.; Laird, M.D.; Singh, N.; Vender, J.R.; Dhandapani, K.M. Dietary Phytochemicals Induce P53- and Caspase-independent Cell Death in Human Neuroblastoma Cells. Int. J. Dev. Neurosci. 2011, 29, 701–710. [Google Scholar] [CrossRef] [Green Version]
- Ozaki, T.; Wu, D.; Sugimoto, H.; Nagase, H.; Nakagawara, A. Runt-Related Transcription Factor 2 (RUNX2) Inhibits P53-Dependent Apoptosis through the Collaboration with HDAC6 in Response to DNA Damage. Cell Death Dis. 2013, 4, e610. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ozaki, T.; Nakagawara, A.; Nagase, H. RUNX Family Participates in the Regulation of P53-Dependent DNA Damage Response. Int. J. Genom. 2013, 2013, 271347. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blyth, K.; Vaillant, F.; Hanlon, L.; Mackay, N.; Bell, M.; Jenkins, A.; Neil, J.C.; Cameron, E.R. Runx2 and MYC Collaborate in Lymphoma Development by Suppressing Apoptotic and Growth Arrest Pathways In Vivo. Cancer Res. 2006, 66, 2195–2201. [Google Scholar] [CrossRef] [PubMed]
- van der Deen, M.; Taipaleenmäki, H.; Zhang, Y.; Teplyuk, N.M.; Gupta, A.; Cinghu, S.; Shogren, K.; Maran, A.; Yaszemski, M.J.; Ling, L.; et al. MicroRNA-34c Inversely Couples the Biological Functions of the Runt-Related Transcription Factor RUNX2 and the Tumor Suppressor P53 in Osteosarcoma. J. Biol. Chem. 2013, 288, 21307–21319. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lengner, C.J.; Steinman, H.A.; Gagnon, J.; Smith, T.W.; Henderson, J.E.; Kream, B.E.; Stein, G.S.; Lian, J.B.; Jones, S.N. Osteoblast Differentiation and Skeletal Development Are Regulated by Mdm2–P53 Signaling. J. Cell Biol. 2006, 172, 909–921. [Google Scholar] [CrossRef]
- Dyer, K.D.; Rosenberg, H.F. Shared Features of Transcription: Mutational Analysis of the Eosinophil/Basophil Charcot-Leyden Crystal Protein Gene Promoter. J. Leukoc. Biol. 2000, 67, 691–698. [Google Scholar] [CrossRef]
- Dyer, K.D.; Rosenberg, H.F. Transcriptional Regulation of Galectin-10 (Eosinophil Charcot-Leyden Crystal Protein). Life Sci. 2001, 69, 201–212. [Google Scholar] [CrossRef]
- Makita, N.; Suzuki, M.; Asami, S.; Takahata, R.; Kohzaki, D.; Kobayashi, S.; Hakamazuka, T.; Hozumi, N. Two of Four Alternatively Spliced Isoforms of RUNX2 Control Osteocalcin Gene Expression in Human Osteoblast Cells. Gene 2008, 413, 8–17. [Google Scholar] [CrossRef]
- Ducy, P.; Zhang, R.; Geoffroy, V.; Ridall, A.L.; Karsenty, G. Osf2/Cbfa1: A Transcriptional Activator of Osteoblast Differentiation. Cell 1997, 89, 747–754. [Google Scholar] [CrossRef] [Green Version]
- Nakajima, S. N-Cadherin Expression and Epithelial-Mesenchymal Transition in Pancreatic Carcinoma. Clin. Cancer Res. 2004, 10, 4125–4133. [Google Scholar] [CrossRef] [Green Version]
- Mrozik, K.M.; Blaschuk, O.W.; Cheong, C.M.; Zannettino, A.C.W.; Vandyke, K. N-Cadherin in Cancer Metastasis, Its Emerging Role in Haematological Malignancies and Potential as a Therapeutic Target in Cancer. BMC Cancer 2018, 18, 939. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Davidson, B.; Tropé, C.G.; Reich, R. Epithelial–Mesenchymal Transition in Ovarian Carcinoma. Front. Oncol. 2012, 2, 33. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Derycke, L.D.M.; Bracke, M.E. N-Cadherin in the Spotlight of Cell-Cell Adhesion, Differentiation, Embryogenesis, Invasion and Signalling. Int. J. Dev. Biol. 2004, 48, 463–476. [Google Scholar] [CrossRef] [PubMed]
- Satelli, A.; Li, S. Vimentin in Cancer and Its Potential as a Molecular Target for Cancer Therapy. Cell. Mol. Life Sci. 2011, 68, 3033–3046. [Google Scholar] [CrossRef] [Green Version]
- Lomonosova, E.; Ryerse, J.; Chinnadurai, G. BAX/BAK–Independent Mitoptosis during Cell Death Induced by Proteasome Inhibition? Mol. Cancer Res. 2009, 7, 1268–1284. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mizuta, T.; Shimizu, S.; Matsuoka, Y.; Nakagawa, T.; Tsujimoto, Y. A Bax/Bak-Independent Mechanism of Cytochrome c Release. J. Biol. Chem. 2007, 282, 16623–16630. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cohen-Solal, K.A.; Boregowda, R.K.; Lasfar, A. RUNX2 and the PI3K/AKT Axis Reciprocal Activation as a Driving Force for Tumor Progression. Mol. Cancer 2015, 14, 137. [Google Scholar] [CrossRef] [Green Version]
- Trzeciakiewicz, A.; Habauzit, V.; Mercier, S.; Lebecque, P.; Davicco, M.-J.; Coxam, V.; Demigne, C.; Horcajada, M.-N. Hesperetin Stimulates Differentiation of Primary Rat Osteoblasts Involving the BMP Signalling Pathway. J. Nutr. Biochem. 2010, 21, 424–431. [Google Scholar] [CrossRef]
- Dai, J.; Li, Y.; Zhou, H.; Chen, J.; Chen, M.; Xiao, Z. Genistein Promotion of Osteogenic Differentiation through BMP2/SMAD5/RUNX2 Signaling. Int. J. Biol. Sci. 2013, 9, 1089–1098. [Google Scholar] [CrossRef]
- Li, J.; Hao, L.; Wu, J.; Zhang, J.; Su, J. Linarin Promotes Osteogenic Differentiation by Activating the BMP-2/RUNX2 Pathway via Protein Kinase A Signaling. Int. J. Mol. Med. 2016, 37, 901–910. [Google Scholar] [CrossRef] [Green Version]
- Auh, Q.-S.; Park, K.-R.; Yun, H.-M.; Lim, H.-C.; Kim, G.-H.; Lee, D.-S.; Kim, Y.-C.; Oh, H.; Kim, E.-C. Sulfuretin Promotes Osteoblastic Differentiation in Primary Cultured Osteoblasts and in Vivo Bone Healing. Oncotarget 2016, 7, 78320–78330. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mu, P.; Hu, Y.; Ma, X.; Shi, J.; Zhong, Z.; Huang, L. Total Flavonoids of Rhizoma Drynariae Combined with Calcium Attenuate Osteoporosis by Reducing Reactive Oxygen Species Generation. Exp. Ther. Med. 2021, 21, 618. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Zhu, R.; Wang, L.; Liu, C.; Ma, R.; Qi, B.; Chen, B.; Li, L.; Guo, Y.; Shi, S.; et al. Radix Salviae miltiorrhizae Improves Bone Microstructure and Strength through Wnt/β-Catenin and Osteoprotegerin/Receptor Activator for Nuclear Factor-ΚB Ligand/Cathepsin K Signaling in Ovariectomized Rats: Radix Salvia Miltiorrhizae Improves Bone Quality in OVX Rats. Phytother. Res. 2018, 32, 2487–2500. [Google Scholar] [CrossRef]
- Ahn, J.-H.; Kim, T.J.; Lee, J.H.; Choi, J.-H. Mutant P53 Stimulates Cell Invasion through an Interaction with Rad21 in Human Ovarian Cancer Cells. Sci. Rep. 2017, 7, 9076. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Son, D.-S.; Kabir, S.M.; Dong, Y.-L.; Lee, E.; Adunyah, S.E. Inhibitory Effect of Tumor Suppressor P53 on Proinflammatory Chemokine Expression in Ovarian Cancer Cells by Reducing Proteasomal Degradation of IκB. PLoS ONE 2012, 7, e51116. [Google Scholar] [CrossRef] [PubMed]
- Yaginuma, Y.; Westphal, H. Abnormal Structure and Expression of the p53 Gene in Human Ovarian Carcinoma Cell Lines. Cancer Res. 1992, 52, 4196. [Google Scholar] [PubMed]
- Thermo Fisher Scientific. Real-Time PCR Handbook; Thermo Fisher Scientific Inc.: Waltham, MA, USA, 2014. [Google Scholar]
- Tan, G.; Heqing, L.; Jiangbo, C.; Ming, J.; Yanhong, M.; Xianghe, L.; Hong, S.; Li, G. Apoptosis Induced by Low-Dose Paclitaxel Is Associated with P53 Upregulation in Nasopharyngeal Carcinoma Cells. Int. J. Cancer 2002, 97, 168–172. [Google Scholar] [CrossRef]
- Lanni, J.S.; Lowe, S.W.; Licitra, E.J.; Liu, J.O.; Jacks, T. P53-Independent Apoptosis Induced by Paclitaxel through an Indirect Mechanism. Proc. Natl. Acad. Sci. USA 1997, 94, 9679–9683. [Google Scholar] [CrossRef] [Green Version]
- Wahl, A.F.; Donaldson, K.L.; Faircnild, C.; Lee, F.Y.F.; Foster, S.A.; Demers, G.W.; Galloway, D.A. Loss of Normal P53 Function Confers Sensitization to Taxol by Increasing G2/M Arrest and Apoptosis. Nat. Med. 1996, 2, 72–79. [Google Scholar] [CrossRef]
- Lavarino, C.; Pilotti, S.; Oggionni, M.; Gatti, L.; Perego, P.; Bresciani, G.; Pierotti, M.A.; Scambia, G.; Ferrandina, G.; Fagotti, A.; et al. P53 Gene Status and Response to Platinum/Paclitaxel-Based Chemotherapy in Advanced Ovarian Carcinoma. J. Clin. Oncol. 2000, 18, 3936–3945. [Google Scholar] [CrossRef]
- Jeswani, G.; Alexander, A.; Saraf, S.; Saraf, S.; Qureshi, A. Ajazuddin Recent Approaches for Reducing Hemolytic Activity of Chemotherapeutic Agents. J. Control. Release 2015, 211, 10–21. [Google Scholar] [CrossRef] [PubMed]
- Brewer, M.S. Natural Antioxidants: Sources, Compounds, Mechanisms of Action, and Potential Applications. Compr. Rev. Food Sci. Food Saf. 2011, 10, 221–247. [Google Scholar] [CrossRef]
- Waksmundzka-Hajnos, M.; Sherma, J.; Kowalska, T. Thin Layer Chromatography in Phytochemistry; CRC Press: Boca Raton, FL, USA, 2008; ISBN 978-1-4200-4677-9. [Google Scholar]
- El-Sayed, N.H.; Khalifa, T.I.; Ibrahim, M.T.; Mabry, T.J. Constituents from Salvia triloba. Fitoterapia 2001, 72, 850–853. [Google Scholar] [CrossRef]
- Tabei, S.M.; Alizadeh, A. Phytochemical Constituents and Antimicrobial Activity of Salvia. Bangladesh J. Bot. 2018, 47, 847–854. [Google Scholar] [CrossRef]
- Çadirci, E.; Süleyman, H.; Gürbüz, P.; Uz, A.K.; Güvenalp, Z.; DemïRezer, L.Ö. Anti-Inflammatory Effects of Different Extracts from Three Salvia Species. Turk. J. Biol. 2012, 36, 59–64. [Google Scholar] [CrossRef]
- Afonso, A.F.; Pereira, O.R.; Cardoso, S.M. Salvia Species as Nutraceuticals: Focus on Antioxidant, Antidiabetic and Anti-Obesity Properties. Appl. Sci. 2021, 11, 9365. [Google Scholar] [CrossRef]
- Rustaie, A.; Hadjiakhoondi, A.; Akbarzadeh, T.; Safavi, M.; Samadi, N.; Sabourian, R.; Khanavi*, M. Phytochemical Constituents and Biological Activities of Salvia Suffruticosa. Res. J. Pharmacogn. 2018, 5, 25–32. [Google Scholar] [CrossRef]
- Rupasinghe, V.; Jiang, Y.; Zhang, L. The Anticancer Properties of Phytochemical Extracts from Salvia Plants. Bot. Targets Ther. 2016, 6, 25–44. [Google Scholar] [CrossRef] [Green Version]
- Sajewicz, M.; Staszek, D.; Wróbel, M.S.; Waksmundzka-Hajnos, M.; Kowalska, T. The HPLC/DAD Fingerprints and Chemometric Analysis of Flavonoid Extracts from the Selected Sage (Salvia) Species. Chromatogr. Res. Int. 2012, 2012, 230903. [Google Scholar] [CrossRef] [Green Version]
- Capek, P.; Hříbalová, V. Water-Soluble Polysaccharides from Salvia officinalis L. Possessing Immunomodulatory Activity. Phytochemistry 2004, 65, 1983–1992. [Google Scholar] [CrossRef]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| S. triloba Acetone Extract | ||||||
|---|---|---|---|---|---|---|
| Paclitaxel | IC50 × 4 | IC50 × 2 | IC50 | IC50 × 0.5 | IC50 × 0.25 | |
| IC50 × 4 | Combination 1 | |||||
| IC50 × 2 | Combination 2 | |||||
| IC50 | Combination 3 | |||||
| IC50 × 0.5 | Combination 4 | |||||
| IC50 × 0.25 | Combination 5 | |||||
| Parameter | Value |
|---|---|
| Matrix library | TRANSFAC MATRIX TABLE, Release 2020.2 |
| Sequence file | MTBP_PM000918140 |
| Profile | vertebrate_non_redundant_minFP.prf |
| Only high-quality matrices | Yes |
| Cut-offs | Minimize false positives |
| S. triloba | Paclitaxel | Combination ** | |||
|---|---|---|---|---|---|
| Dose (µg/mL) | Effect * | Dose (µg/mL) | Effect * | Total Dose (µg/mL) | Effect * |
| 0.25 | 0.01 | 0.01 | 0.01 | 0.2537 | 0.01 |
| 19 | 0.41077 | 0.787 | 0.97536 | 19.787 | 0.88325 |
| 38 | 0.51115 | 1.574 | 0.97752 | 39.574 | 0.85706 |
| 76 | 0.3187 | 3.148 | 0.9 | 79.148 | 0.83451 |
| 152 | 0.65196 | 6.296 | 0.95046 | 158.296 | 0.81048 |
| 304 | 0.9 | 12.592 | 0.97428 | 316.592 | 0.9 |
| Drug | Dm * (µg/mL) | m ** | r *** |
|---|---|---|---|
| S. triloba | 50.9282 | 0.85854 | 0.9587 |
| Paclitaxel | 0.22062 | 1.1417 | 0.89187 |
| S. triloba + Paclitaxel | 13.8357 | 0.95052 | 0.91799 |
| Combination Total Dose (µg/mL) | Effect * | CI Value | CI Description ** |
|---|---|---|---|
| 0.2537 | 0.01 | 3.5694 | Strong antagonism |
| 19.787 | 0.8833 | 0.6415 | Synergism |
| 39.574 | 0.8571 | 1.5787 | Antagonism |
| 79.148 | 0.8345 | 3.6856 | Strong antagonism |
| 158.296 | 0.8105 | 8.5413 | Strong antagonism |
| 316.592 | 0.9 | 8.7917 | Strong antagonism |
| Effect * | DRI S. triloba | DRI Paclitaxel |
|---|---|---|
| 0.01 | 0.9904 | 0.3907 |
| 0.8833 | 28.3034 | 1.6498 |
| 0.8571 | 10.7945 | 0.6729 |
| 0.8345 | 4.4115 | 0.2891 |
| 0.8105 | 1.8205 | 0.1251 |
| 0.9 | 2.1655 | 0.1201 |
| Factor Name | RUNX2 | |||||
|---|---|---|---|---|---|---|
| TFBS * | 1 | 2 | 3 | 4 | 5 | 6 |
| Core similarity score | 1.00 | 1.00 | 0.895 | 1.00 | 1.00 | 1.00 |
| Matrix similarity score | 0.998 | 0.918 | 0.906 | 0.894 | 0.893 | 0.913 |
| Start position ** | 68799245 | 68799934 | 68806888 | 68807452 | 68808063 | 68809252 |
| End position ** | 68799254 | 68799943 | 68806897 | 68807461 | 68808072 | 68809261 |
| Sequence | CAACCACAAG | GAACCACTTA | AAGCCACATA | CCACCACGCC | GAGGTGGTGC | TAACCACCTC |
| Reference | ArrayExpress [47] | ArrayExpress [47] | GEO [48] | GEO [49] | ArrayExpress [47] | ArrayExpress [47] |
| Distance Traveled by Fraction (cm) | Distance Traveled by Solvent Front (cm) | Retention Factor |
|---|---|---|
| 2.54 | 11.43 | 0.2222 |
| 4.445 | 11.43 | 0.3889 |
| 6.5024 | 11.43 | 0.5689 |
| 9.2202 | 11.43 | 0.8067 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Saleh, N.A.M.; El-bary, R.B.E.-d.A.; Mpingirika, E.Z.; Essa, H.L.; El-Sayed, M.M.H.; Sherbetjian, M.S.; Elfandi, H.F.; Wahed, M.A.A.; Arafeh, R.; Amleh, A. Evaluating the Potential Anticancer Properties of Salvia triloba in Human-Osteosarcoma U2OS Cell Line and Ovarian Adenocarcinoma SKOV3 Cell Line. Appl. Sci. 2022, 12, 11545. https://doi.org/10.3390/app122211545
Saleh NAM, El-bary RBE-dA, Mpingirika EZ, Essa HL, El-Sayed MMH, Sherbetjian MS, Elfandi HF, Wahed MAA, Arafeh R, Amleh A. Evaluating the Potential Anticancer Properties of Salvia triloba in Human-Osteosarcoma U2OS Cell Line and Ovarian Adenocarcinoma SKOV3 Cell Line. Applied Sciences. 2022; 12(22):11545. https://doi.org/10.3390/app122211545
Chicago/Turabian StyleSaleh, Naela Adel Mohammed, Rowan Bahaa El-din Abd El-bary, Eric Zadok Mpingirika, Hanaa L. Essa, Mayyada M. H. El-Sayed, Mirna Sarkis Sherbetjian, Hanin Fadel Elfandi, Muhammad Adel Abdel Wahed, Rami Arafeh, and Asma Amleh. 2022. "Evaluating the Potential Anticancer Properties of Salvia triloba in Human-Osteosarcoma U2OS Cell Line and Ovarian Adenocarcinoma SKOV3 Cell Line" Applied Sciences 12, no. 22: 11545. https://doi.org/10.3390/app122211545
APA StyleSaleh, N. A. M., El-bary, R. B. E. -d. A., Mpingirika, E. Z., Essa, H. L., El-Sayed, M. M. H., Sherbetjian, M. S., Elfandi, H. F., Wahed, M. A. A., Arafeh, R., & Amleh, A. (2022). Evaluating the Potential Anticancer Properties of Salvia triloba in Human-Osteosarcoma U2OS Cell Line and Ovarian Adenocarcinoma SKOV3 Cell Line. Applied Sciences, 12(22), 11545. https://doi.org/10.3390/app122211545