
Obtaining Fat-Soluble Pigments—Carotenoids from the Biomass of Chlorella Microalgae

 , ,
, , 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cultivation
2.2. Concentration of the Biomass
2.3. Activation
2.4. Extraction
2.5. Fractionation of the Pigments
2.6. Analysis of the Obtained Carotenoids
2.7. Statistical Analysis
3. Results and Discussion
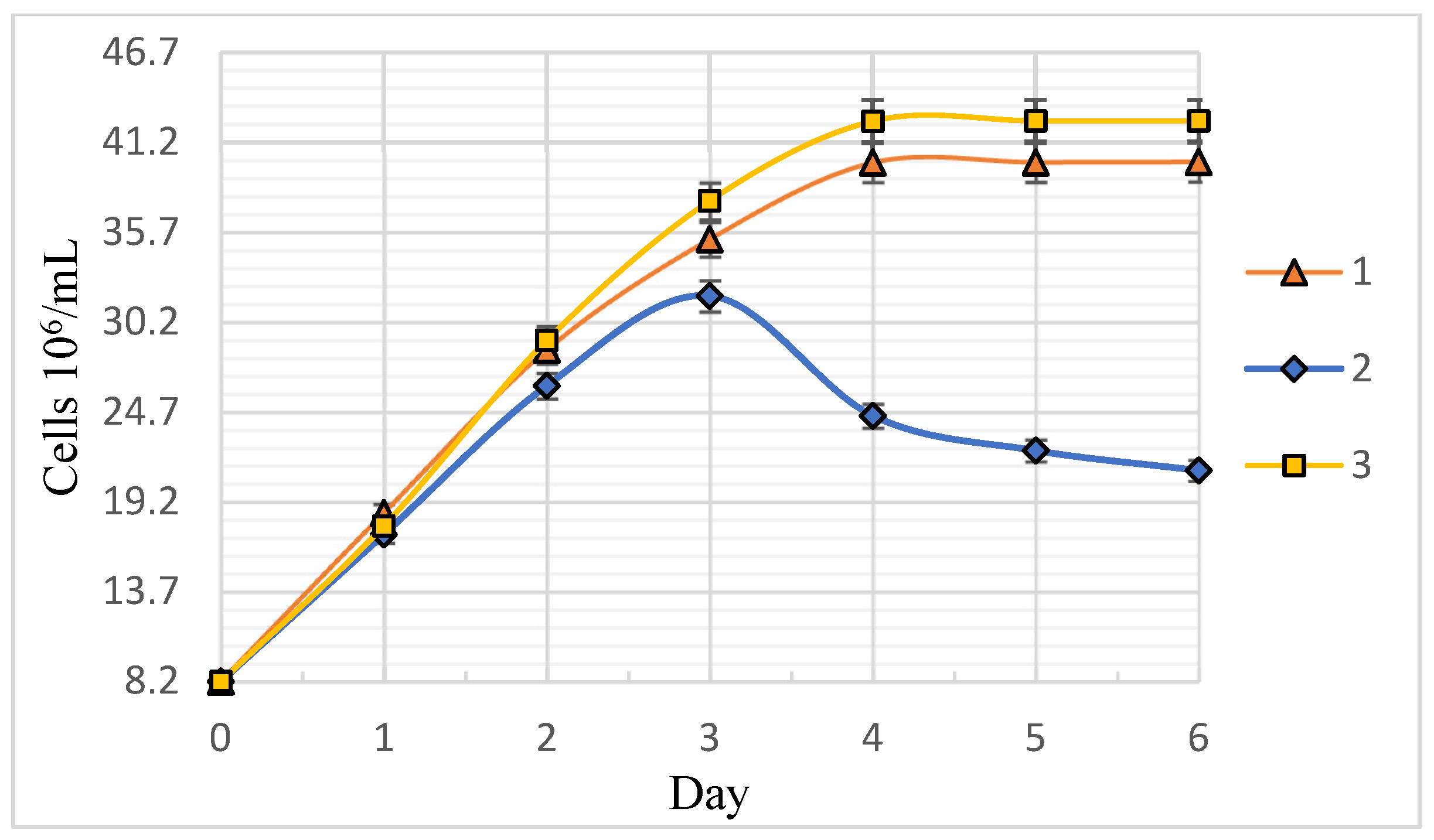
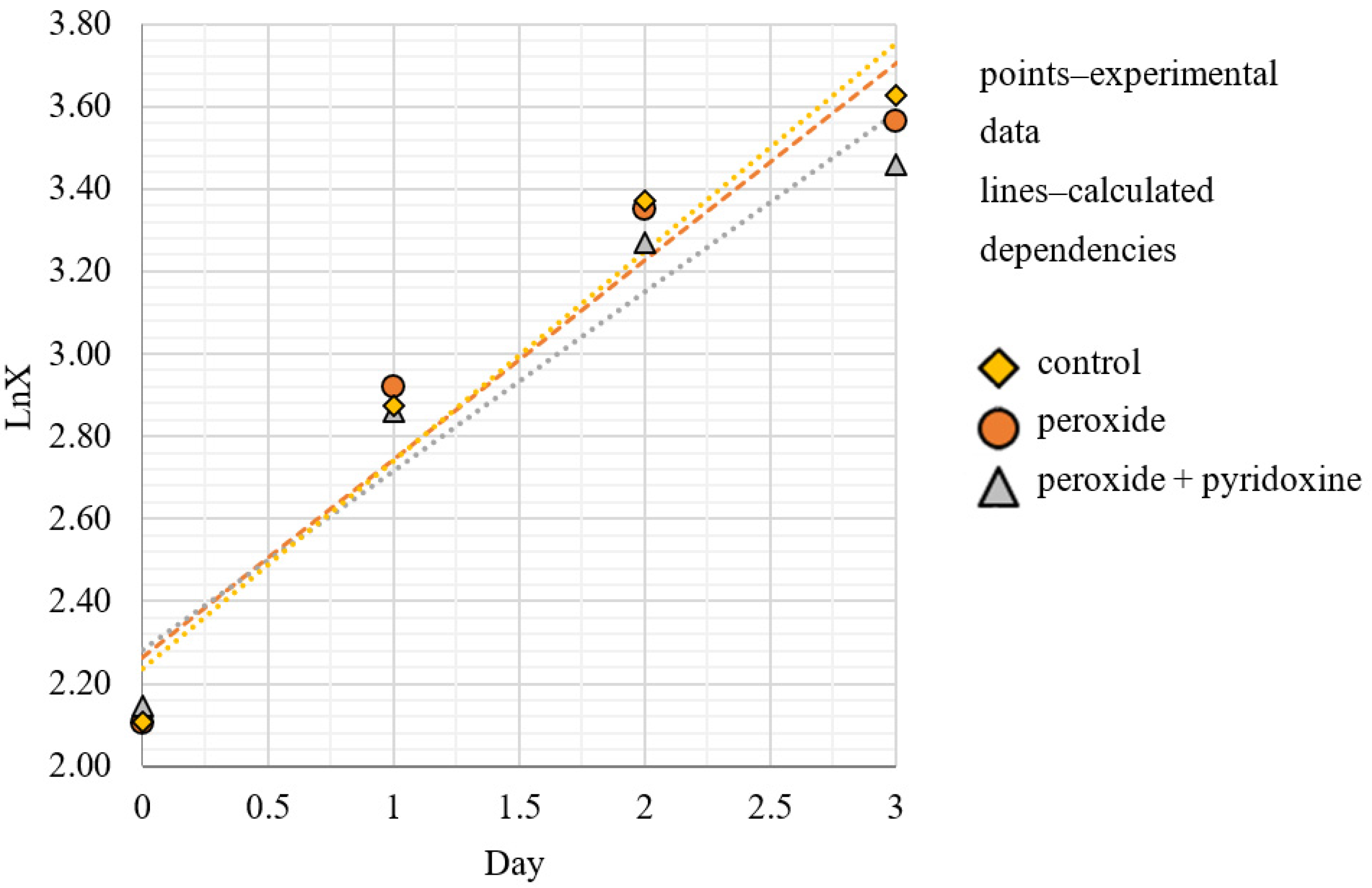
3.1. Dynamics and Specific Rate of C. vulgaris BIN Population Growth
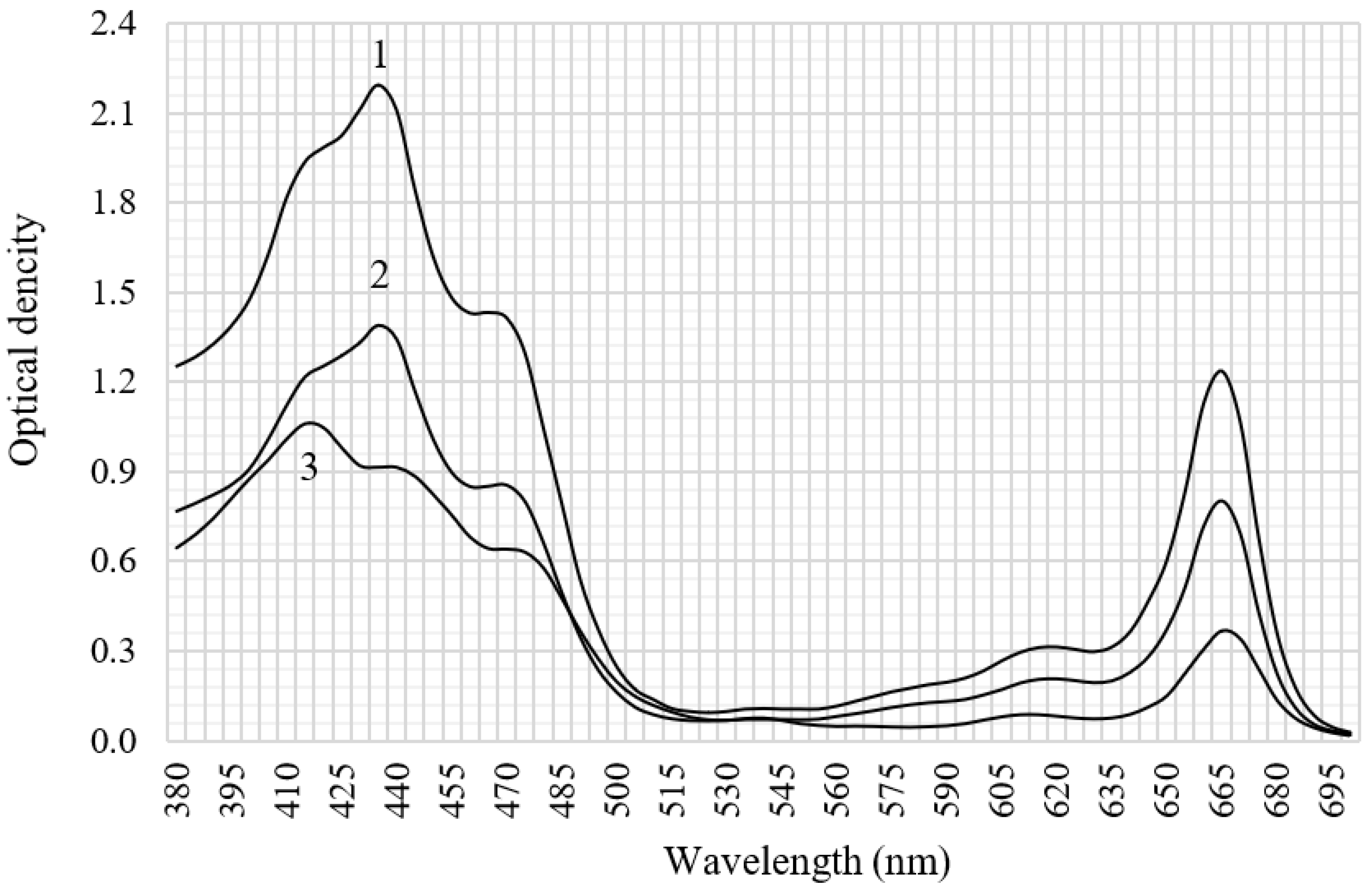
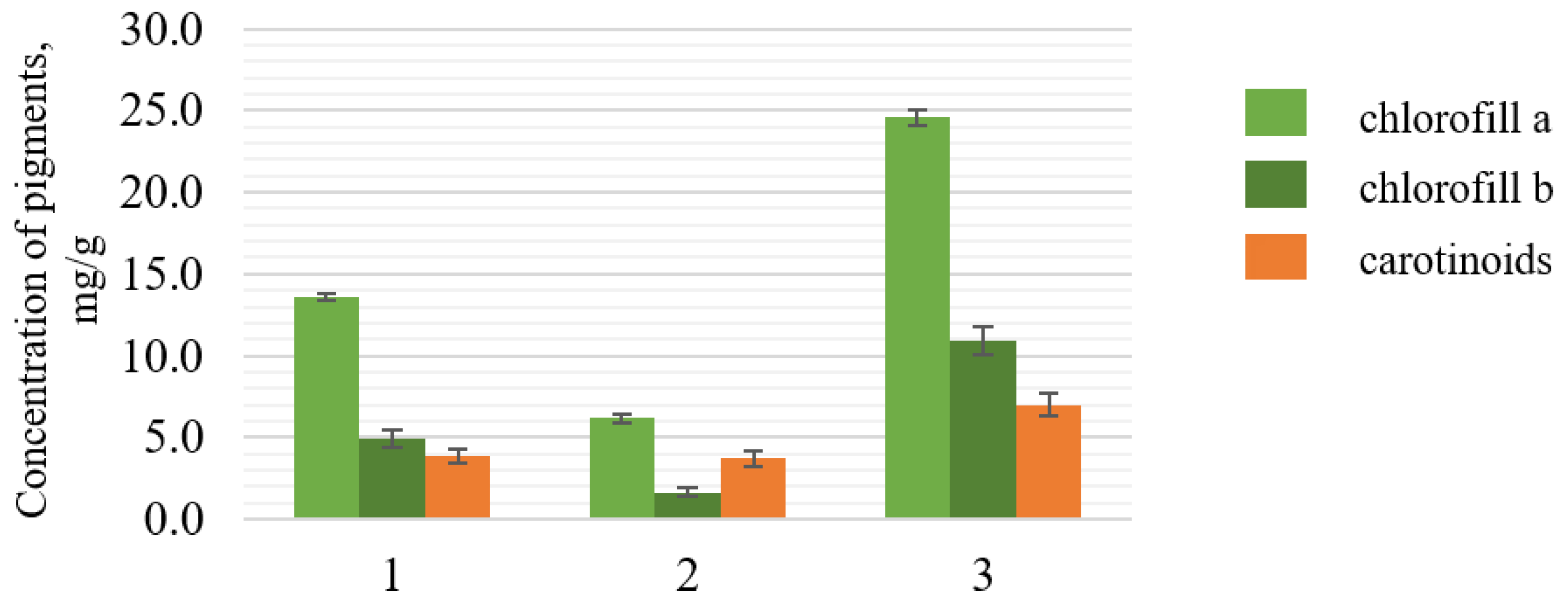
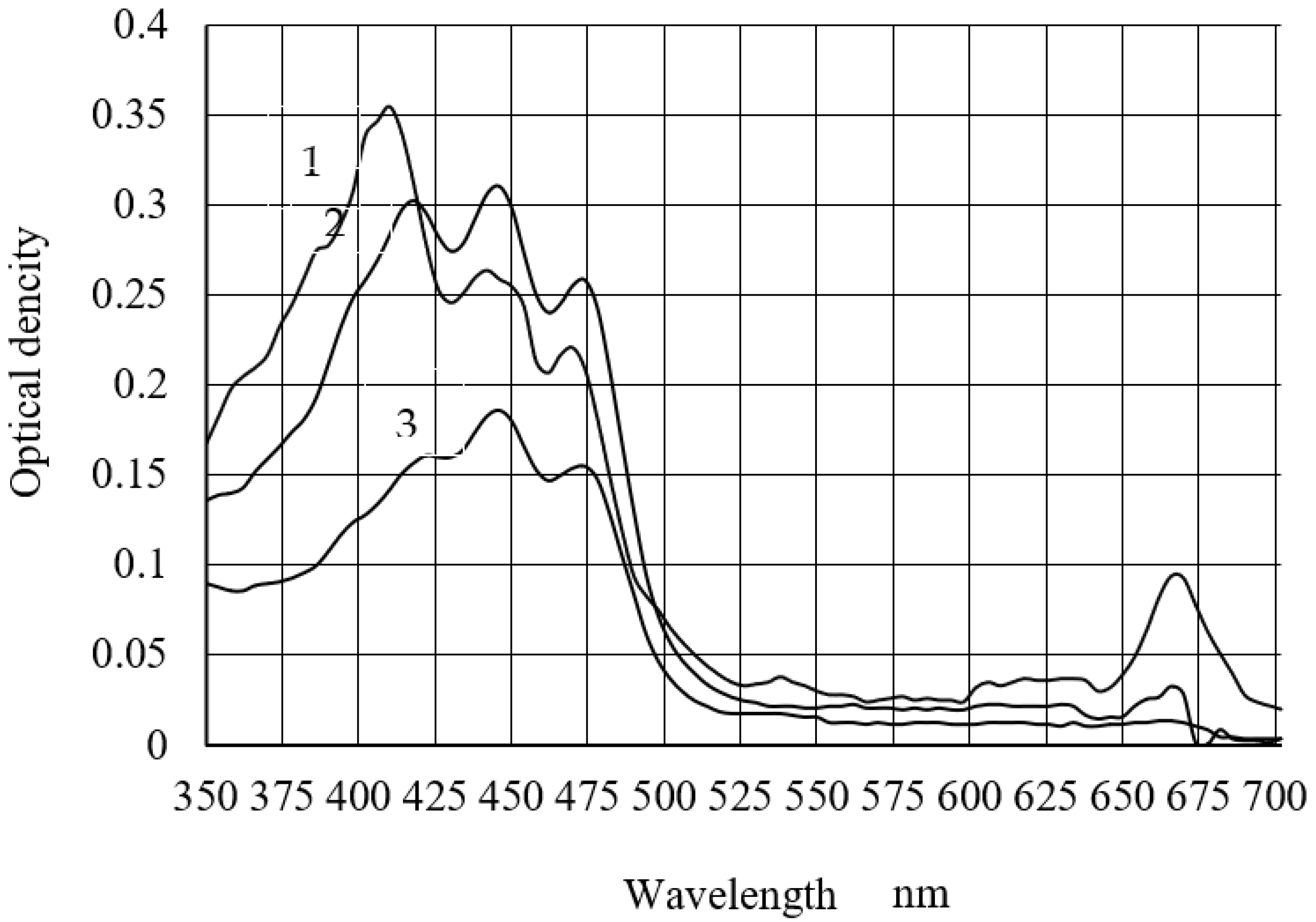
3.2. Spectral Characteristics and Carotenoid Content of C. vulgaris BIN Biomass Extracts
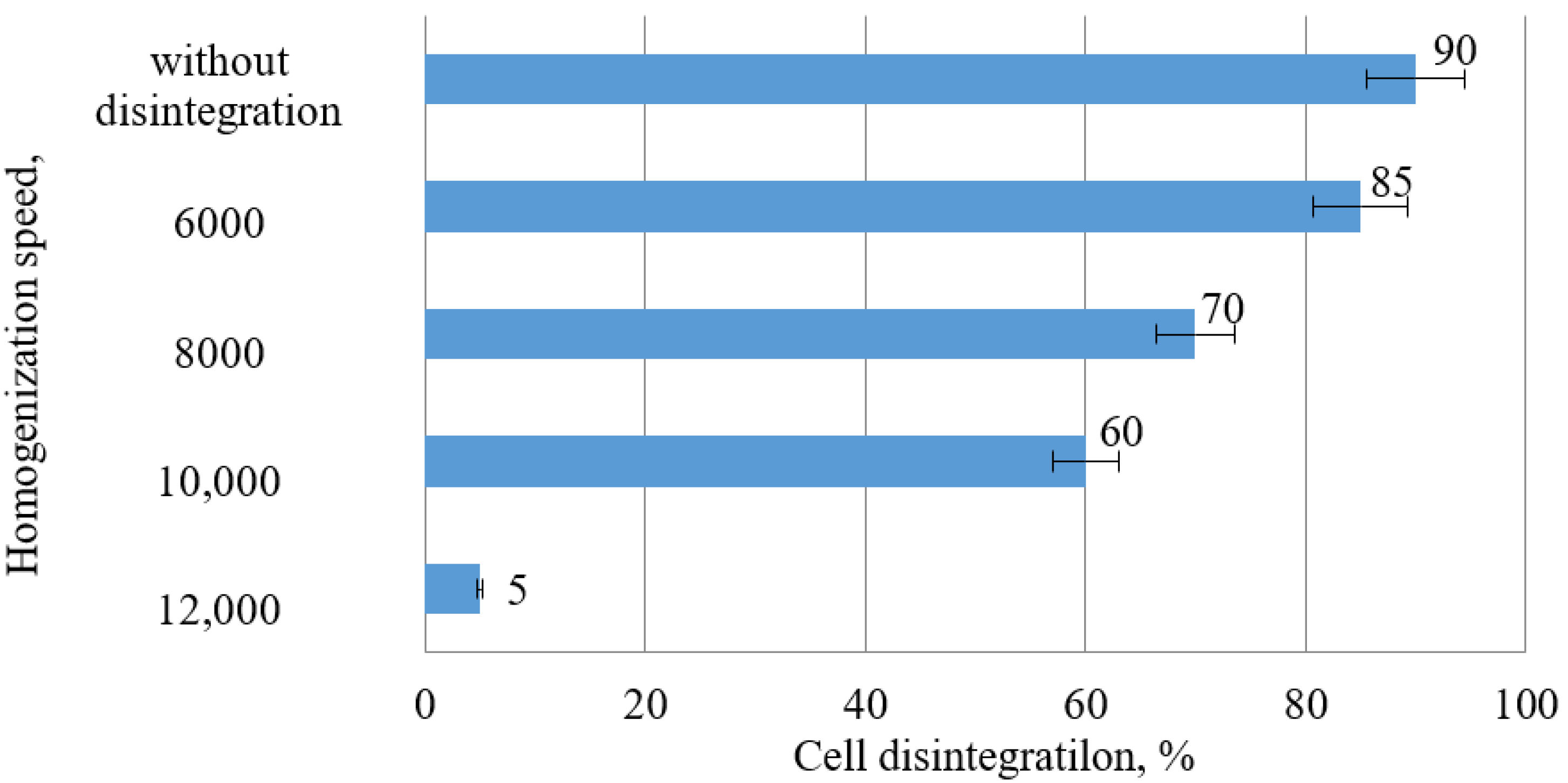
3.3. Influence of Activated C. vulgaris BIN Biomass on the Yield of Carotenoids in the Extracts
3.4. Extraction and Fractionation of the Pigments
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Eggersdorfer, M.; Wyss, A. Carotenoids in human nutrition and health. Arch. Biochem. Biophys. Res. Commun. 2018, 652, 18–26. [Google Scholar] [CrossRef] [PubMed]
- Machado Sierra, E.; Serrano, M.C.; Manares, A.; Guerra, A.; Aranguren Díaz, Y. Microalgae: Potential for Bioeconomy in Food Systems. Appl. Sci. 2021, 11, 11316. [Google Scholar] [CrossRef]
- Hyrslova, I.; Krausova, G.; Smolova, J.; Stankova, B.; Branyik, T.; Malinska, H.; Huttl, M.; Kana, A.; Curda, L.; Doskocil, I. Functional Properties of Chlorella vulgaris, Colostrum, and Bifidobacteria, and Their Potential for Application in Functional Foods. Appl. Sci. 2021, 11, 5264. [Google Scholar] [CrossRef]
- Singh, D.P.; Khattar, J.S.; Rajput, A.; Chaudhary, R.; Singh, R. High production of carotenoids by the green microalga Asterarcys quadricellulare PUMCC 5.1.1 under optimized culture conditions. PLoS ONE 2019, 14, e0221930. [Google Scholar] [CrossRef] [Green Version]
- Kleinegris, D.M.M.; Janssen, M.; Brandenburg, W.A. The Selectivity of Milking of Dunaliella salina. Mar. Biotechnol. 2010, 12, 14–23. [Google Scholar] [CrossRef] [Green Version]
- Harvey, P.J.; Ben-Amotz, A. Towards a sustainable Dunaliella salina microalgal biorefinery for 9-cis β-carotene production. Algal Res. 2020, 50, 102002. [Google Scholar] [CrossRef]
- Diprat, A.B.; Thys, R.C.S.; Rodrigues, E.; Rech, R. Chlorella sorokiniana: A new alternative source of carotenoids and proteins for gluten-free bread. LWT 2020, 134, 109974. [Google Scholar] [CrossRef]
- Guedes, A.C.; Amaro, H.M.; Malcata, F.X. Microalgae as sources of carotenoids. Mar. Drugs 2011, 9, 625–644. [Google Scholar] [CrossRef]
- Matsukawa, R.; Hotta, M.; Masuda, Y.; Chihara, M.; Karube, I. Antioxidants from carbon dioxide fixing Chlorella sorokiniana. J. Appl. Phycol. 2000, 12, 263–267. [Google Scholar] [CrossRef]
- Marino, T.; Casella, P.; Sangiorgio, P.; Verardi, A.; Ferraro, A.; Hristoforou, E.; Molino, A.; Musmarra, D. Natural be-ta-carotene: A microalgae derivate for nutraceutical applications. Chem. Eng. Trans. 2020, 79, 103–108. [Google Scholar] [CrossRef]
- Menezes, R.; Soares, A.; Marques, J.; Lopes, R.; Arantes, R.; Derner, R.; Antoniosi, F.N. Culture medium influence on growth, fatty acid, and pigment composition of Choricystis minor var. minor: A suitable microalga for biodiesel production. J. Appl. Phycol. 2016, 28, 2679–2686. [Google Scholar] [CrossRef]
- Ambati, R.R.; Phang, S.-M.; Ravi, S.; Aswathanarayana, R.G. Astaxanthin: Sources, Extraction, Stability, Biological Activities and Its Commercial Applications—A Review. Mar. Drugs 2014, 12, 128–152. [Google Scholar] [CrossRef] [PubMed]
- Shlykova, A.N.; Balabaev, A.A.; Trukhina, E.V.; Bazarnova, Y.G. Obtaining carotenoid pigments from Chlorella microalgae. Vestn. PNRPU. Chem. Technol. Biotechnol. 2020, 3, 20–37. Available online: https://cyberleninka.ru/article/n/poluchenie-karotinoidnyh-pigmentov-iz-mikrovodorosley-chlorella (accessed on 10 February 2022).
- Damergi, E.; Schwitzguébel, J.-P.; Refardt, D.; Shivom, S.; Holliger, C.; Ludwig, C. Extraction of carotenoids from Chlorella vulgaris using green solvents and syngas production from residual biomass. Algal Res. 2017, 25, 488–495. [Google Scholar] [CrossRef]
- Novoveská, L.; Ross, M.E.; Stanley, M.S.; Pradelles, R.; Wasiolek, V.; Sassi, J.F. Microalgal Carotenoids: A Review of Production, Current Markets, Regulations, and Future Direction. Mar. Drugs 2019, 17, 640. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kul’nev, V.; Stupin, V.; Borzenkov, A. Biological Waste Water Rehabilitation of Sugar Factories by Algocoenosis Correction. Ecol. Ind. Russ. 2017, 21, 16–20. (In Russian) [Google Scholar] [CrossRef]
- Kulnev, V.; Nasonov, A.; Zhogin, I.; Tsvetkov, I.; Grabarnik, V.; Karelin, N. The Experiences of the Managed Algoritmization Recreational Pond. Ecol. Ind. Russ. 2020, 24, 58–64. (In Russian) [Google Scholar] [CrossRef]
- Becker, E.W. Microalgae in human and animal nutrition. In Handbook of Microalgal Culture; Richmond, A., Ed.; Blackwell: Oxford, UK, 2004; pp. 312–351. [Google Scholar]
- Kuznetsova, T.A.; Nikitina, M.S.; Sevastyanova, A.D. Directed cultivation of Chlorella sorokiniana to increase carotenoid synthesis. Vestn. VGUIT 2019, 81, 34–39. [Google Scholar] [CrossRef]
- Machmudah, S.; Goto, M. Methods for extraction and analysis of carotenoids. Nat. Prod. 2013, 3367–3411. [Google Scholar] [CrossRef]
- Bazarnova, J.; Kuznetsova, T.; Aronova, E.; Popova, L.; Pochkaeva, E. A method for obtaining plastid pigments from the biomass of Chlorella microalgae. Agron. Res. 2020, 18, 2317–2327. [Google Scholar] [CrossRef]
- Bazarnova, J.; Nilova, L.; Trukhina, E.; Bernavskaya, M.; Smyatskaya, Y.; Aktar, T. Use of microalgae biomass for fortification of food products from grain. Foods 2021, 10, 3018. [Google Scholar] [CrossRef] [PubMed]
- Saseendran, S. A revised method for pigment extraction from marine nannoplanktonic algal cultures. J. Algal Biomass Util. 2017, 4, 47–52. [Google Scholar]
- Martin-Gordillo, L.; Cuaresma, M.; Fernández-Recamales, M.Á.; Sayago, A.; Vílchez, C.; Garbayo, I. Cultivation of Microalgae Chlorella Using Wine Industry by-products. Proceedings 2020, 66, 30. [Google Scholar] [CrossRef]
- Parra, M.; Stahl, S.; Hellmann, H. Vitamin B₆ and Its Role in Cell Metabolism and Physiology. Cells 2018, 22, 84. [Google Scholar] [CrossRef] [Green Version]
- Farghl, A.A. Thiamine and Pyridoxine Alleviate Oxidative Damage by Copper Stress in Green Alga Chlorella vulgaris. Egypt. J. Microbiol. 2012, 47, 97–110. [Google Scholar] [CrossRef] [Green Version]
- Shavirina, O.B. Toxicity of copper for green algae culture Scenedesmus quadricauda during pH fluctuations. Int. J. Appl. Basic Res. 2016, 4, 741–743. [Google Scholar]
- Bazarnova, Y.G.; Kuznetsova, T.; Trukhina, E. Aquabiotechnology of directed cultivation of microalgae Chlorella sorokiniana biomass. IOP Conf. Ser. Earth Environ. Sci. 2019, 288, 012037. [Google Scholar] [CrossRef]
- Britton, G.B. Biochemistry of natural pigments. Mosc. Mir 1986, 422. [Google Scholar]
- Shivalkar, Y.K.; Prabha, R. Effect of pH and temperature on carotenoid pigments produced from Rhodotorula minuta International. J. Fermented Foods 2014, 3, 105–113. [Google Scholar] [CrossRef]
- Kuregyan, A.G. Spectrophotometry in carotenoid analysis. Basic Res. 2015, 2, 5166–5172. [Google Scholar]








{kind=link}
{kind=link}
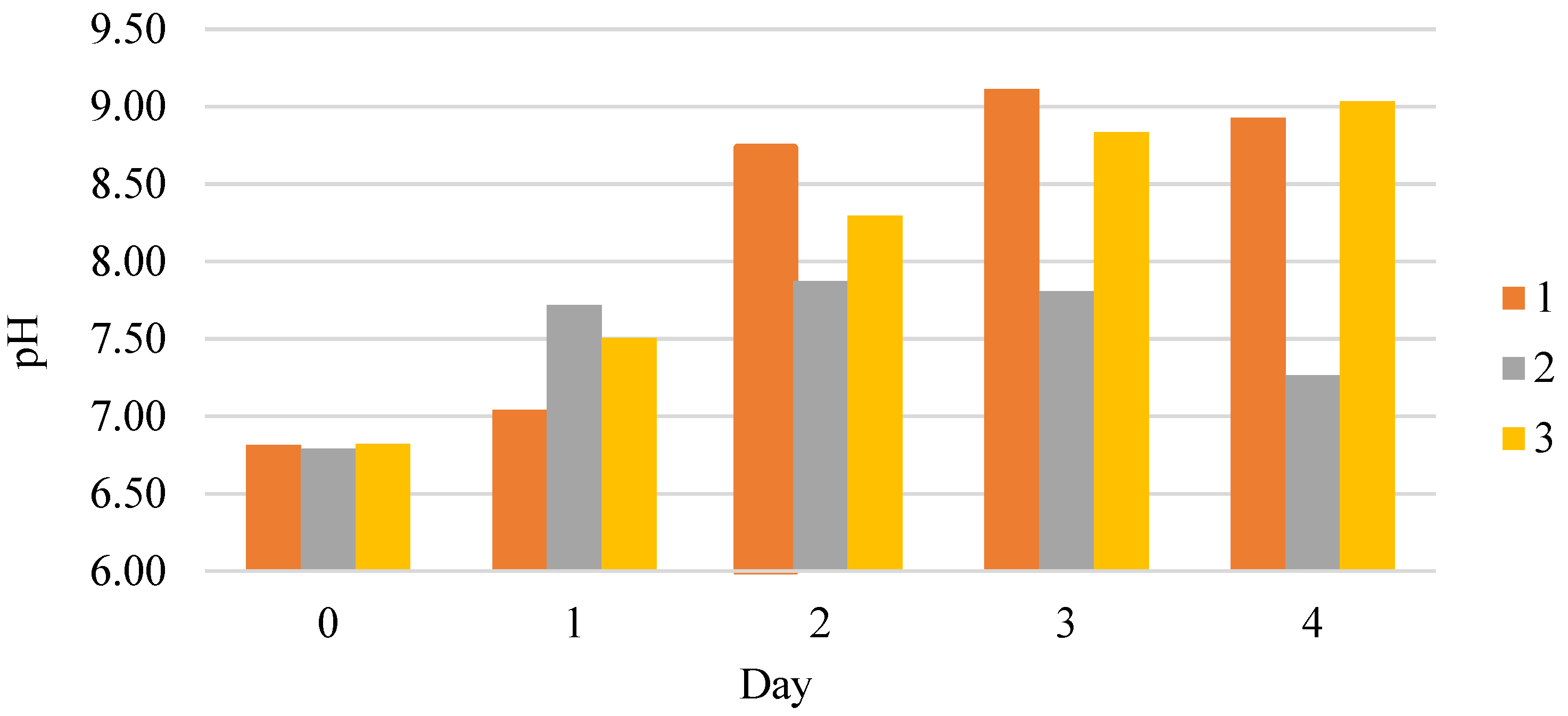{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Culture Medium A | ||||
| Substance | Molecular Weight,(g/mol) | Concentration in Stock Solution, (mg/L) | Concentration in Culture Medium, (µg/L) | Consumption,(mL/L) |
| ZnSO4·7H2O | 287.53 | 0.1 | 100 | 10.0 |
| CuSO4·5H2O | 249.66 | 0.1 | 10 | 0.1 |
| CoSO4·7H2O | 281.06 | 0.1 | 100 | 5.0 |
| MnCl2·4H2O | 197.91 | 0.1 | 500 | 5.0 |
| H3BO3 | 61.83 | 0.1 | 50 | 0.5 |
| Na2MoO4·2H2O | 241.96 | 0.1 | 100 | 0.1 |
| Culture Medium B (Microelements) | ||||
| Substance | Molecular Weight,(g/mol) | Concentration in Stock Solution, (mg/L) | Concentration in Culture Medium, (µg/L) | Consumption,(mL/L) |
| FeCl3·6H2O | 270.21 | 1.0 | 4.000 | 4.0 |
| Na2EDTA·2H2O | 372.24 | 1.0 | 6.000 | 6.0 |
| Macroelements | ||||
| Substance | Molecular Weight,(g/mol) | Concentration in Mother Medium, (mg/L) | Consumption,(g/L) | |
| KNO3 | 101.1 | 1.000 | 3.26 | |
| KH2PO4 | 136.07 | 100 | 0.32 | |
| MgSO4·7H2O | 246.48 | 240 | 2.46 | |
| Homogenization Speed, rpm | CCar, µg/mL |
|---|---|
| 0 (control) | 11.46 |
| 6000 | 23.16 |
| 8000 | 14.85 |
| 10,000 | 17.52 |
| 12,000 | 14.85 |
| Identified Pigments | Reverse Phase HPLC | Spectrophotometry |
|---|---|---|
| Total amount of carotenoids | 12.1 ± 1.2 | 6.55 ± 0.67 |
| Lutein | 5.4 ± 0.5 | − |
| Fucoxanthin | 1.4 ± 0.1 | − |
| β-carotene | 0.52 ± 0.05 | − |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bazarnova, J.; Smyatskaya, Y.; Shlykova, A.; Balabaev, A.; Đurović, S. Obtaining Fat-Soluble Pigments—Carotenoids from the Biomass of Chlorella Microalgae. Appl. Sci. 2022, 12, 3246. https://doi.org/10.3390/app12073246
Bazarnova J, Smyatskaya Y, Shlykova A, Balabaev A, Đurović S. Obtaining Fat-Soluble Pigments—Carotenoids from the Biomass of Chlorella Microalgae. Applied Sciences. 2022; 12(7):3246. https://doi.org/10.3390/app12073246
Chicago/Turabian StyleBazarnova, Julia, Yulia Smyatskaya, Antonina Shlykova, Aleksei Balabaev, and Saša Đurović. 2022. "Obtaining Fat-Soluble Pigments—Carotenoids from the Biomass of Chlorella Microalgae" Applied Sciences 12, no. 7: 3246. https://doi.org/10.3390/app12073246
APA StyleBazarnova, J., Smyatskaya, Y., Shlykova, A., Balabaev, A., & Đurović, S. (2022). Obtaining Fat-Soluble Pigments—Carotenoids from the Biomass of Chlorella Microalgae. Applied Sciences, 12(7), 3246. https://doi.org/10.3390/app12073246