
Nitrogen Metabolism and Antioxidant Capacity of Selected Vegetables from Organic and Conventional Crops


Abstract
:1. Introduction
2. Materials and Methods
2.1. The Origin of the Plant Material
- celery family Apiaceae:
- -
- dill [Anethum graveolens L.]—dill (marking on the charts),
- -
- common carrot [Daucus carota L.]—carrot,
- -
- celery [Apium graveolens L.]—celery,
- -
- parsley [Petroselinum crispum (Mill.)]—parsley,
- amaranth family Amaranthaceae:
- -
- beet [Beta vulgaris L.]—beet,
- amaryllis family Amaryllidaceae:
- -
- spring onions [Allium cepa L.]—onion,
- cabbage family Brassicaceae:
- -
- Chinese cabbage [Brassica rapa L. var. pekinensis]—ch. cabbage,
- -
- white head cabbage [Brassica oleracea var. capitata L.]—w. cabbage,
- -
- rocket [Eruca vesicaria subsp. Sativa]—rocket.
2.2. Nitrate Content
2.3. Nitrate Reductase Activity
2.4. Glutamine Synthetase Activity
2.5. Total Chlorophyll Content
2.6. Phenolic Compound Content
2.7. Ascorbate Content
2.8. Total Antioxidant Capacity
2.9. Statistical Analyses
3. Results and Discussion
3.1. Nitrate Content
3.2. Nitrate Reductase Activity
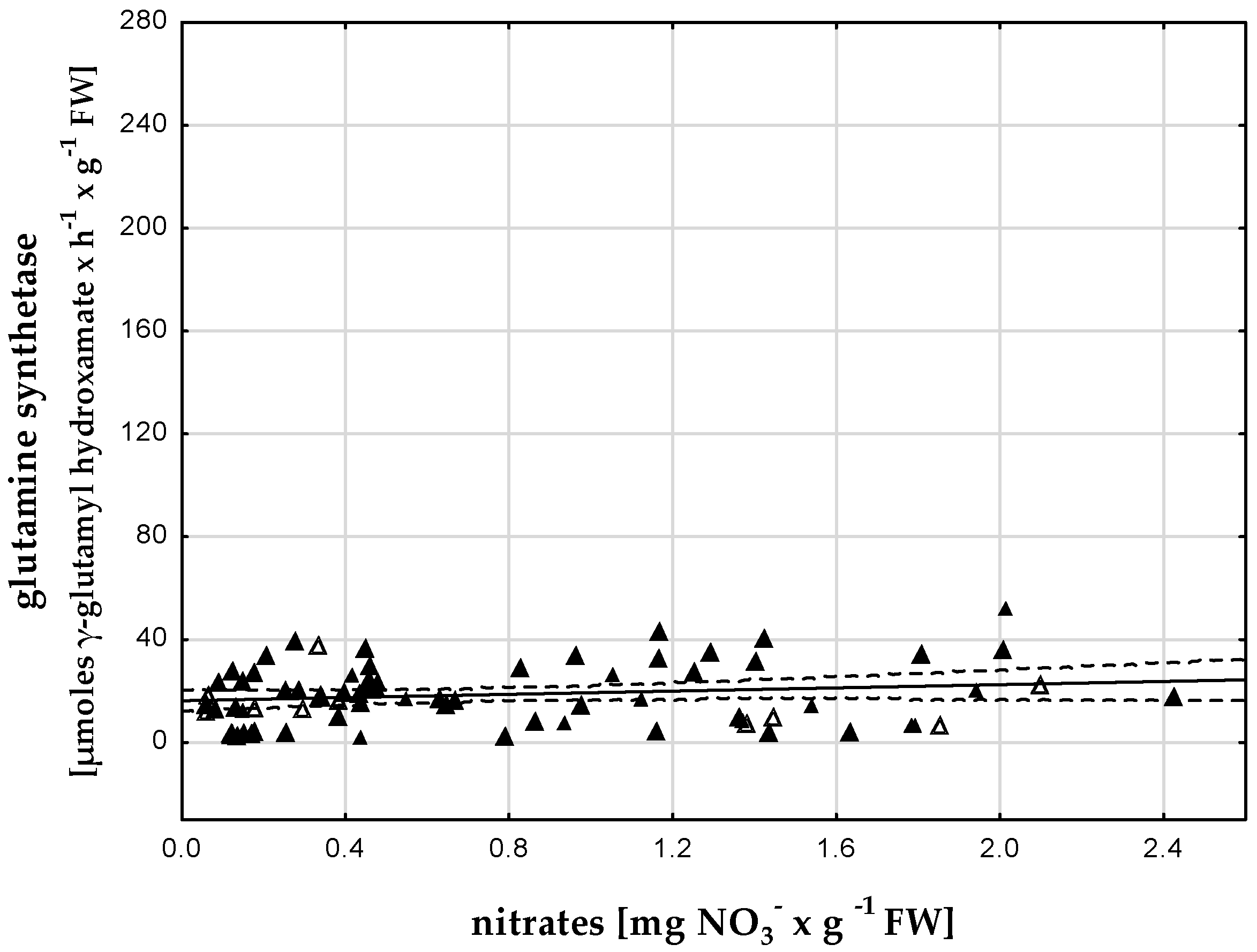
3.3. Glutamine Synthetase Activity
3.4. Total Chlorophyll Content
3.5. Phenolic Compound Content
3.6. Ascorbate Content
3.7. Total Antioxidant Capacity
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- IFOAM. Consolidated Annual Report of IFOAM-Organics International; IFOAM: Bonn, Germany, 2018. [Google Scholar]
- Willer, H.; Lernoud, J. The World of Organic Agriculture. Statistics and Emerging Trends; Research Institute of Organic Agriculture (FiBL) & IFOAM-Organics International: Frick, Switzerland, 2017. [Google Scholar]
- Hurtado-Barroso, S.; Tresserra-Rimbau, A.; Vallverdu-Queralt, A.; Lamuela-Raventos, R.M. Organic food and the impact on human health. Crit. Rev. Food Sci. Nutr. 2017, 59, 704–714. [Google Scholar] [CrossRef]
- Apaolaza, V.; Hartmann, P.; D’Souza, C.; López, C.M. Eat organic–Feel good? The relationship between organic food consumption, health concern and subjective wellbeing. Food Qual. Prefer. 2018, 63, 51–62. [Google Scholar] [CrossRef]
- Vigar, V.; Myers, S.; Oliver, C.; Arellano, J.; Robinson, S.; Leifert, C. A Systematic Review of Organic Versus Conventional Food Consumption: Is There a Measurable Benefit on Human Health? Nutrients 2020, 12, 7. [Google Scholar] [CrossRef] [PubMed]
- Györéné, K.G.; Varga, A.; Lugasi, A. A comparison of chemical composition and nutritional value of organically and conventionally grown plant derived foods. Orv. Hetil. 2006, 147, 2081–2090. [Google Scholar]
- Palupi, E.; Jayanegara, A.; Ploeger, A.; Kahl, J. Comparison of nutritional quality between conventional and organic dairy products: A meta-analysis. J. Sci. Food Agric. 2012, 92, 2774–2781. [Google Scholar] [CrossRef] [PubMed]
- Smith-Spangler, C.; Brandeau, M.L.; Hunter, G.E.; Clay Bavinger, J.; Pearson, M.; Eschbach, P.J.; Sundaram, V.; Liu, H.; Schirmer, P.; Stave, C.; et al. Are organic foods safer or healthier than conventional alternatives? A systematic review. Ann. Intern. Med. 2012, 157, 348–366. [Google Scholar] [CrossRef]
- Barański, M.; Średnicka-Tober, D.; Volakakis, N.; Seal, C.; Sanderson, R.; Stewart, G.B.; Benbrook, C.; Biavati, B.; Markellou, E.; Giotis, C.; et al. Higher antioxidant and lower cadmium concentrations and lower incidence of pesticide residues in organically grown crops: A systematic literature review and meta-analyses. Br. J. Nutr. 2014, 112, 794–811. [Google Scholar] [CrossRef] [PubMed]
- Brantsæter, A.L.; Ydersbond, T.A.; Hoppin, J.A.; Haugen, M.; Meltzer, H.M. Organic food in the diet: Exposure and health implications. Annu. Rev. Public Health 2017, 38, 295–313. [Google Scholar] [CrossRef] [PubMed]
- Mie, A.; Andersen, H.R.; Gunnarsson, S.; Kahl, J.; Kesse-Guyot, E.; Rembiałkowska, E.; Quaglio, G.; Grandjean, P. Human health implications of organic food and organic agriculture: A comprehensive review. Environ. Health 2017, 16, 111. [Google Scholar] [CrossRef]
- González, N.; Marquès, M.; Nadal, M.; Domingo, J.L. Occurrence of environmental pollutants in foodstuffs: A review of organic vs. conventional food. Food Chem. Toxicol. 2019, 125, 370–375. [Google Scholar] [CrossRef]
- Rathke, G.W.; Behrens, T.; Diepenbrock, W. Integrated nitrogen management strategies to improve seed yield, oil content and nitrogen efficiency of winter oilseed rape (Brassica napus L.): A review. Agric. Ecosyst. Environ. 2006, 117, 80–108. [Google Scholar] [CrossRef]
- Brandt, K.; Mølgaard, J.P. Organic agriculture: Does it enhance or reduce the nutritional value of plant foods? J. Sci. Food Agric. 2001, 81, 924–931. [Google Scholar] [CrossRef]
- Winter, C.K.; Davis, S.F. Organic food. J. Food Sci. 2006, 71, 117–124. [Google Scholar] [CrossRef]
- Hallmann, E.; Rembiałkowska, E. Characterisation of antioxidant compounds in sweet bell pepper (Capsicum annuum L.) under organic and conventional growing systems. J. Sci. Food Agric. 2012, 92, 2409–2415. [Google Scholar] [CrossRef]
- Dixon, R.A. Natural products and plant disease resistance. Nature 2001, 411, 843–847. [Google Scholar] [CrossRef]
- Wallis, C.M.; Galarneau, E.R.A. Phenolic compound induction in plant-microbe and plant-insect interactions: A meta-analysis. Front. Plant Sci. 2020, 11, 580753. [Google Scholar] [CrossRef]
- Chowdhary, V.; Alooparampil, S.; Pandya, R.V.; Tank, J.G. Physiological function of phenolic compounds in plant defense system. In Phenolic Compounds—Chemistry, Synthesis, Diversity, Non-Conventional Industrial, Pharmaceutical and Therapeutic Applications; IntechOpen: Rijeka, Croatia, 2021. [Google Scholar]
- Vibhakar, C.; Sheena, A.; Rohan, V.P.; Jigna, G.T. Physiological Function of Phenolic Compounds in Plant Defense System; IntechOpen: Rijeka, Croatia, 2021; pp. 185–205. [Google Scholar]
- Fine, P.V.A.; Miller, Z.J.; Mesones, I.; Irazuzta, S.; Appel, H.M.; Stevens, M.H.H. The growth defense tradeoff and habitat specialisation by plants in Amazonian forest. Ecology 2006, 87, 150–162. [Google Scholar] [CrossRef]
- Yamamoto, Y.; Gaynor, R.B. Therapeutic potential of inhibition of the NF-JB pathway in the treatment of inflammation and cancer. J. Clin. Investig. 2001, 107, 135–142. [Google Scholar] [CrossRef] [PubMed]
- Cushnie, T.P.T.; Lamb, A.J. Antimicrobial activity of flavonoids. Int. J. Antimicrob. Agents. 2005, 26, 343–356. [Google Scholar] [CrossRef] [PubMed]
- Shahidi, F.; Wanasundara, P.K. Phenolic antioxidants. Crit. Rev. Food Sci. Nutr. 1992, 32, 67–103. [Google Scholar] [CrossRef] [PubMed]
- Migliaccio, S.; Anderson, J.B. Isoflavones and skeletal health: Are these molecules ready for clinical application. Osteoporos. Int. 2003, 14, 361–368. [Google Scholar]
- Abad-Garcia, B.; Berrueta, L.A.; Garmon-Lobato, S.; Gallo, B.; Vicente, F. A general analytical strategy for the characterisation of phenolic compounds in fruit juices by high-performnace liquid chromatography with diode array detection coupled to electrospray ionisation and triple quadrupole mass spectrometry. J. Chromatol. 2009, 1216, 5398–5415. [Google Scholar] [CrossRef]
- Faller, A.L.K.; Fialho, E.F.N.U. Polyphenol content and antioxidant capacity in organic and conventional plant foods. J. Food Compos. Anal. 2010, 23, 561–568. [Google Scholar] [CrossRef]
- de Oliveira Pereira, F.; dos Santos Pereira, R.; de Souza Rosa, L.; Teodoro, A.J. Organic and conventional vegetables: Comparison of the physical and chemical characteristics and antioxidant activity. Afr. J. Biotechnol. 2016, 15, 1746–1754. [Google Scholar]
- Ren, F.; Reilly, K.; Kerry, J.P.; Gaffney, M.; Hossain, M.; Rai, D.K. Higher antioxidant activity, total flavonols, and specific quercetin glucosides in two different onion (Allium cepa L.) varieties grown under organic production: Results from a 6-year field study. J. Agric. Food Chem. 2017, 65, 5122–5132. [Google Scholar] [CrossRef] [PubMed]
- Basay, S.; Cimen, A.; Baba, Y.; Yildirim, A.B.; Turker, A.U. Organic and conventional solanaceous vegetables: Comparison of phenolic constituents, antioxidant and antibacterial potentials. Acta Aliment. 2021, 50, 442–452. [Google Scholar] [CrossRef]
- Czech, A.; Szmigielski, M.; Sembratowicz, I. Nutritional value and antioxidant capacity of organic and conventional vegetables of the genus Allium. Sci. Rep. 2022, 12, 18713. [Google Scholar] [CrossRef] [PubMed]
- Cataldo, D.A.; Haroon, M.; Schrader, L.E.; Youngs, V.L. Rapid colorimetric determination of nitrate in plant tissue by nitration of salicylic acid. Commun. Soil Sci. Plant Anal. 1975, 6, 71–80. [Google Scholar] [CrossRef]
- Jaworski, E.G. Nitrate reductase assay in intact plant tissues. Biochem. Biophys. Res. Commun. 1971, 43, 1274–1279. [Google Scholar] [CrossRef]
- Hipkin, C.; Syrret, P.J. Some effects of nitrogen-starvation on nitrogen and carbohydrate metabolism in Ankistrodesmus brauni. Planta 1977, 133, 209–214. [Google Scholar] [CrossRef]
- Hiscox, J.D.; Israelstram, G.F. A method for the extraction of chlorophyll from leaf tissue without maceration. Can. J. Bot. 1979, 57, 1332–1334. [Google Scholar] [CrossRef]
- Wellburn, A.R. The spectral determination of chlorophylls a and b, as well as total carotenoids, using various solvents with spectrophotometers of different resolution. J. Plant Physiol. 1994, 144, 307–313. [Google Scholar] [CrossRef]
- Singleton, V.L.; Rossi, J.A. Colorimetry of total phenolics with phosphomolybdic-phosphotungstic acid reagents. Am. J. Enol. Vitic. 1965, 16, 144–158. [Google Scholar] [CrossRef]
- Law, M.Y.; Charles, S.A.; Halliwell, B. Glutathione and ascorbic acid in spinach (Spinacia oleracea) chloroplasts. The effect of hydrogen peroxide and of paraquat. Biochem. J. 1983, 210, 899–903. [Google Scholar] [PubMed]
- Costa, H.; Gallego, S.M.; Tomaro, M.L. Effect of UV-B radiation on antioxidant defense system in sunflower cotyledons. Plant Sci. 2002, 162, 939–945. [Google Scholar] [CrossRef]
- Re, R.; Pellegrini, N.; Proteggente, A.; Pannala, A.; Yang, M.; Rice-Evans, C. Antioxidant activity applying an improved ABTS radical cation decolorisation assay. Free Radic. Biol. Med. 1999, 26, 1231–1237. [Google Scholar] [CrossRef] [PubMed]
- Bartosz, G. Druga Twarz Tlenu; Naukowe PWN: Wydaw, Poland, 1995. [Google Scholar]
- Gajewska, M.; Czajkowska, A.; Bartodziejska, B. Zawartość azotanów (III) i (V) w wybranych warzywach dostępnych w handlu detalicznym regionu łódzkiego. Ochr. Sr. I Zasobów Nat. 2009, 40, 388–395. [Google Scholar]
- Niewczas, J.; Kamionowska, M.; Mitek, M. Zawartość azotanów (III) i (V) w owocach nowych odmian dyni olbrzymiej (Cucurbita maxima). Żywność Nauka Technol. Jakość 2006, 2, 238–245. [Google Scholar]
- Wojciechowska, R. Akumulacja azotanów a jakość produktów rolniczych. In W: Ochrona Środowiska Naturalnego w XXI Wieku—Nowe Wyzwania i Zagrożenia; Wiech, K., Kłokoczka, H., Kaszycki, P., Eds.; Politechnika Koszalińska: Kraków, Poland, 2005; pp. 21–27. [Google Scholar]
- Grudzińska, M.; Zgórska, K. Wpływ obróbki wstępnej oraz metod gotowania na zawartość azotanów w warzywach. Rocz. Ochr. Sr. 2005, 7, 233–241. [Google Scholar]
- Tietze, M.; Burghardt, A.; Brągiel, P.; Mac, J. Zawartość związków azotowych w produktach spożywczych. Ann. Univ. Mariae Curie-Skłodowska. Sect. EE Zootech. 2007, 25, 71–77. [Google Scholar]
- Murawa, D.; Banaszkiewicz, T.; Majewska, E.; Błaszczuk, B.; Sulima, J. Zawartość azotanów (III) i (V) w wybranych gatunkach warzyw i ziemniakach dostępnych w handlu w Olsztynie w latach 2003–2004. Bromat. Chem. Toksykol.—XLI 2008, 1, 67–71. [Google Scholar]
- Regulation of the Minister of Health of 13 January 2003. Journal of Laws No. 37, Items 325 and 326, on the Maximum Levels of Chemical and Biological Contaminants that May Be Present in Food, Food Ingredients, Permitted Additives, Processing Aids or on the Surface of Food. Available online: https://eli.gov.pl/eli/DU/2003/326/ogl (accessed on 3 May 2017).
- Bano, M.J.; Lorente, J.; Castillo, J.; Benavente-Garcia, O.; Rio, J.A.; Ortuno, A.; Quirin, K.W.; Gerard, D. Phenolic diterpens, flavones, and rosmarinic acid distribution during the development of leaves, flowers, stems, and roots of Rosmarinum officinalis. Antioxidant activity. J. Agric. Food Chem. 2003, 51, 4247–4253. [Google Scholar] [CrossRef]
- Raczuk, J.; Wadas, W.; Głozak, K. Nitrates and nitrites in selected vegetables purchased at supermarkets in Siedlce, Poland. Rocz. Panstw. Zakl. Hig. 2014, 65, 15–20. [Google Scholar] [PubMed]
- Krężel, J. Przydatność wybranych odmian buraka ćwikłowego do uprawy na zbiór pęczkowy, do przechowania i przetwórstwa. Zeszyty Naukowe AR Wrocław, Roln. 2005, 87, 325–332. [Google Scholar]
- Krężel, J.; Kołota, E. Wpływ nawożenia azotowego na plonowanie i wartość iologiczną buraka ćwikłowego, uprawianego na zbiór pęczkowy. Zeszyty Naukowe AR Wrocław Ogrodn. 2007, 41, 547–552. [Google Scholar]
- Zhou, Z.Y.; Wang, M.J.; Wang, J.S. Nitrate and nitrite contamination in vegetables in China. Food Rev Int. 2000, 16, 61–76. [Google Scholar] [CrossRef]
- Majkowska−Gadomska, J.; Arcichowska, K.; Wierzbicka, B. Nitrate content of the edible parts of vegetables and spice plants. Acta Sci. Pol. Hortorum. Cultus 2009, 8, 25–35. [Google Scholar]
- Kozik, E. Wpływ terminu zbioru oraz nawożenia azotem i potasem na zawartość azotanów w salacie uprawianej w szklarni. Acta Agrophys. 2006, 7, 633–642. [Google Scholar]
- Weightman, R.M.; Dyer, C.; Buxton, J.; Farrington, D.S. Effects of light level, time of harvest and position within field on the variability of tissue nitrate concentration in commercial crops of lettuce (Lactuca sativa) and endive (Cichorium endiva). Food Addit. Contam. 2006, 23, 462–469. [Google Scholar] [CrossRef]
- Parks, S.E.; Irving, D.E.; Milham, P.J. A critical evaluation of on-farm rapid tests for measuring nitrate in leafy vegetables. Sci. Hortic. 2012, 134, 1–6. [Google Scholar] [CrossRef]
- Obidoska, G.; Sadowska, A. Wpływ pH podłożą i jego zasobność w azot na plonowanie Withania somnifera (L.) Dun. Biuletyn IHAR 2003, 228, 373–381. [Google Scholar]
- Santamaria, P.; Elia, A.; Serio, F.; Todaro, E. A survey of nitrate and oxalate content in fresh vegetables. J. Sci. Food Agric. 1999, 79, 1882–1888. [Google Scholar] [CrossRef]
- Anjana; Umar, S.; Iqbal, M.; Abrol, Y.P. Are nitrate concentrations in leafy vegetables within safe limits? Curr. Sci. 2007, 92, 355–360. [Google Scholar]
- Wadas, W.; Jabłońska-Ceglarek, R.; Kosterna, E. The nitrates content in early potato tubers depending on growing conditions. EJPAU. Horiculture 2005, 8, 18. Available online: http://www.ejpau.media.pl/ (accessed on 27 May 2013).
- Tamme, T.; Reinik, M.; Roasto, M.; Jukham, K.; Tenno, T.; Kiis, A. Nitrates and nitrites in vegetables and vegetable-based products and their intakes by the Estonian population. Addit. Contam. 2006, 23, 355–361. [Google Scholar] [CrossRef] [PubMed]
- Radziejewska-Kubzdela, E.; Walkowiak-Tomczak, D.; Bieganska-Marecik, R. Wpływ pakowania i przechowywania w atmosferze modyfikowanej na zawartość azotanów [V] i [III] w salatce warzywnej typu coleslaw oraz na jej cechy sensoryczne i fizykochemiczne. Żywność Nauka Technol. Jakość 2008, 15, 261–268. [Google Scholar]
- Gajewski, M. Czynniki wpływające na jakość marchwi przeznaczonej do przetwórstwa. Kated. Roślin Warzywnych I Lecz. SGGW 2010, 1–5. [Google Scholar]
- Rożek, S. Czynniki wpływające na akumulację azotanów w plonie warzyw. Zesz. Nauk. Akad. Rol. W Krakowie 2000, 71, 19–31. [Google Scholar]
- Wawrzyniak, A.; Kwiatkowski, S.; Gronowska-Senger, A. Ocena zawartości azotanów i azotynów oraz białka ogółem w wybranych warzywach uprawianych konwencjonalnie i ekologicznie. Rocz. PZH 1997, 48, 181–186. [Google Scholar]
- Rutkowska, B. Azotany i azotyny w ziemniakach z gospodarstw ekologicznych i konwencjonalnych. Rocz. PZH 2001, 52, 231–236. [Google Scholar]
- Guadagnin, S.G.; Rath, S.; Reyes, F.G.R. Evaluation of the nitrate content in leaf vegetables produced through different agricultural systems. Food Addit Contam. 2005, 22, 1203–1208. [Google Scholar] [CrossRef] [PubMed]
- Murawska, B.; Piekut, A.; Jachymska, J.; Mitura, K.; Lipińska, K.J. Ekologiczny i konwencjonalny system gospodarowania a wielkość i jakość plonu wybranych roślin uprawnych. In Infrastruktura i Ekologia Terenów Wiejskich; Nr III, Polska Akademia Nauk; Oddział w Krakowie: Krakow, Poland, 2015; pp. 663–675. [Google Scholar]
- Aslam, M.; Travis, R.L.; Rains, D.W.; Huffaker, R.C. Differential effect of ammonium on the induction of nitrate and nitrite reductase activities in roots of barley (Hordeum vulgare) seedlings. Physiol. Plant. 1997, 101, 612–619. [Google Scholar] [CrossRef]
- Tischner, R.; Waldeck, B.; Goyal, S.S.; Rains, W.D. Effect of nitrate pulses on the nitrate-uptake rate, synthesis of mRNA coding for nitrate reductase, and nitrate-reductase activity in the roots of barley seedlings. Planta 1993, 189, 533–537. [Google Scholar] [CrossRef]
- Samuelson, M.E.; Öhlén, E.; Lind, M.; Larsson, C.M. Nitrate regulation of nitrate uptake and nitrate reductase expression in barley grown at different nitrate: Ammonium ratios at constant relative nitrogen addition rate. Physiol. Plant. 1995, 94, 254–260. [Google Scholar] [CrossRef]
- Smoleń, S.; Wojciechowska, R.; Sady, W.; Szura, A. Wpływ formy azotu nawozowego i dokarmiania dolistnego na plon i gospodarkę azotową korzeni spichrzowych marchwi (Daucus carota L.). Acta Agrophys. 2006, 7, 721–732. [Google Scholar]
- Lillo, C.; Meyer, C.; Lea, U.S.; Provan, F.; Oltedal, S. Mechanism and importance of posttranslational regulation of nitrate reductase. J. Exp. Bot. 2004, 55, 1275–1282. [Google Scholar] [CrossRef] [PubMed]
- Man, H.M.; Abd-El Baki, G.K.; Stegmann, P.; Weiner, H.; Kaiser, W.M. The activation state of nitrate reductase is not always correlated with total nitrate reductase activity in leaves. Planta 1999, 209, 462–468. [Google Scholar] [CrossRef]
- Reda, M.; Kłobus, G.; Buczek, J. Structure and regulation of nitrate reductase. Postepy Biochem. 2000, 46, 99–106. [Google Scholar]
- Starck, Z. Różnorodne funkcje węgla i azotu w rośliny. Kosmos 2006, 55, 243–257. [Google Scholar]
- Małolepsza, U. Biosynteza tlenku azotu w roślinach. Postępy Biochem. 2007, 53, 263–271. [Google Scholar]
- Miflin, B.J.; Habash, D.Z. The role of glutamine synthetase and glutamate dehydrogenase in nitrogen assimilation and possibilities for improvement in the nitrogen utilisation of crops. J. Exp. Bot. 2002, 53, 979–987. [Google Scholar] [CrossRef]
- Kozłowski, S.; Goliński, P.; Golińska, B. Barwniki chlorofilowe, jako wskaźniki wartości użytkowej gatunków i odmian traw. Zesz. Probl. Post. Nauk Rol. 2001, 474, 215–223. [Google Scholar]
- Balder, F.H.; de Vogel, J.; Jansen, C.J.F.; Weijenberg, M.P.; Piet Van den Brandt, A.; Westenbrink, S.; Van der Meer, R.R.; Goldbohm, A. Heme and chlorophyll intakeand risk of colorectal cancer in the Netherlands Cohort Study. Cancer Epidemiol. Biomark. Prev. 2006, 15, 717–725. [Google Scholar] [CrossRef]
- McQuistan, T.J.; Simonich, M.T.; Pratt, M.M.; Pereira, C.B.; Hendricks, J.D.; Dashwood, R.H.; Williams, D.E.; Bailey, G.S. Cancer chemoprevention by dietary chlorophylls: A 12,000-animal dose-dose matrix biomarker and tumour study. Food Chem. Toxicol. 2012, 50, 341–352. [Google Scholar] [CrossRef]
- Pietrzak, M.; Halicka, H.D.; Wieczorek, Z.; Wieczorek, J.; Darżynkiewicz, Z. Attenuation of acridine mutagen ICR-191-DNA interactions and DNA damage by the mutagen interceptor chlorophyllin. Biophys. Chem. 2008, 135, 69–75. [Google Scholar] [CrossRef]
- Jubert, C.; Mata, J.; Bench, G.; Dashwood, R.; Pereira, C.; Tracewell, W.; Turteltaub, K.; Williams, D.; Bailey, G. Effects of chlorophyll and chlorophyllin on low-dose afl atoxin B(1) pharmacokinetics in human volunteers. Cancer Prev. Res. 2009, 2, 1015–1022. [Google Scholar] [CrossRef] [PubMed]
- Niewiadomska, A.; Sawicka, A. Wpływ zróżnicowanych poziomów nawożenia azotowego i sodowego na aktywność nitrogenazy i koncentrację chlorofilu w mieszance koniczyny łąkowej z festulolium. Woda-Sr.-Obsz. Wiej. 2010, 30, 121–131. [Google Scholar]
- Benbrook, C.; Zhao, X.; Yáñez, J.; Davies, N.; Andrews, P. New evidence confirms the nutritional superiority of plant-based organic foods. In State of Science Review: Nutritional Superiority of Organic Foods; The Organic Center: Washington, DC, USA, 2008; pp. 421–431. [Google Scholar]
- Hallmann, E. Wartość odżywcza i zawartość związków bioaktywnych w warzywach i owocach z produkcji ekologicznej. In Materiały Szkoleniowe XII Forum Rolników Ekologicznych Baszkowice; Szkoła Główna Gospodarstwa Wiejskiego w Warszawie: Borzkowice, Poland, 2014; pp. 1–21. [Google Scholar]
- Wieczorek, J.; Wieczorek, Z. Części nadziemne popularnych warzyw jako źródło karotenoidów i chlorofilu w żywności. Bromat. Chem. Toksykol. XLIX 2016, 3, 422–426. [Google Scholar]
- Chassy, A.W.; Bui, L.; Renaud, E.N.C.; Van Horn, M.; Mitchell, A.E. Three-year comparison of the content of antioxidant micro constituents and several quality characteristics in organic and conventionally managed tomatoes and bell peppers. J. Agric. Food Chem. 2006, 54, 8244–8252. [Google Scholar] [CrossRef]
- Crinnion, W.J. Organic foods contain higher levels of certain nutrients, lower levels of pesticides, and may provide health benefits for the consumer. Altern. Med. Rev. J. Clin. Ther. 2010, 15, 4–12. [Google Scholar]
- Yu, X.; Guo, L.; Jiang, G.; Song, Y.; Muminov, M.A. Advances of organic products over conventional productions with respect to nutritional quality and food security. Acta Ecol. Sinica. 2018, 38, 53–60. [Google Scholar] [CrossRef]
- Heimler, D.; Romani, A.; Ieri, F. Plant Polyphenol Content, Soil Fertilization and Agricultural Management: A Review. Eur. Food Res. Technol. 2017, 243, 1107–1115. [Google Scholar] [CrossRef]
- Rembiałkowska, E. Quality of plant products from organic agriculture. J. Sci. Food Agric. 2007, 87, 2757–2762. [Google Scholar] [CrossRef]
- Mazzoncini, M.; Antichi, D.; Silvestri, N.; Ciantelli, G.; Sgherri, C. Organically vs conventionally grown winter wheat: Effects on grain yield, technological quality, and on phenolic composition and antioxidant properties of bran and refined flour. Food Chem. 2015, 175, 445–451. [Google Scholar] [CrossRef] [PubMed]
- Silva, L.F.L.E.; Souza, D.C.; Resende, L.V.; Nassur, R.D.C.M.; Samartini, C.Q.; Gonçalves, W.M. Nutritional evaluation of non-conventional vegetables in Brazil. An. Da Acad. Bras. De Ciências 2018, 90, 1775–1787. [Google Scholar] [CrossRef]
- Giannakourou, M.C.; Taoukis, P.S. Effect of alternative preservation steps and storage on vitamin C stability in fruit and vegetable products: Critical review and kinetic modelling approaches. Foods 2021, 10, 2630. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.K.; Kader, A.A. Preharvest and postharvest factors influencing vitamin C content of horticultural crops. Postharvest Biol. Technol. 2000, 20, 207–220. [Google Scholar] [CrossRef]
- Zhang, L.; Zhang, F.; Wang, Y.; Ma, X.; Shen, Y.; Wang, X.; Deng, Y. Physiological and metabolomic analysis reveals maturity stage-dependent nitrogen regulation of vitamin C content in pepper fruit. Front. Plant Sci. 2023, 13, 1049785. [Google Scholar] [CrossRef]
- Hesari, N.; Szegő, A.; Mirmazloum, I.; Pónya, Z.; Kiss-Bába, E.; Kolozs, H.; Papp, I. High-Nitrate-Supply-Induced Transcriptional Upregulation of Ascorbic Acid Biosynthetic and Recycling Pathways in Cucumber. Plants 2023, 12, 1292. [Google Scholar] [CrossRef]
- Gomiero, T. Food quality assessment in organic vs. conventional agricultural produce: Findings and issues. Appl. Soil Ecol. 2018, 123, 714–728. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| (A) Source of Variation | Nitrate | Nitrate Reductase | ||||
| d.f. | F | p | d.f. | F | p | |
| Species (S) | 8 | 91.557 | <0.0001 | 8 | 246.116 | <0.0001 |
| Cultivation (C) | 2 | 1.451 | NS | 2 | 9.816 | <0.0001 |
| S × C | 16 | 10.013 | <0.0001 | 16 | 75.976 | <0.0001 |
| Error | 54 | 111 | ||||
| (B) Source of Variation | Glutamine Synthetase | Total Chlorophyll | ||||
| d.f. | F | p | d.f. | F | p | |
| Species (S) | 8 | 38.898 | <0.0001 | 8 | 70.755 | <0.0001 |
| Cultivation (C) | 2 | 4.407 | 0.0153 | 2 | 73.39 | <0.0001 |
| S × C | 16 | 53.052 | <0.0001 | 16 | 25.106 | <0.0001 |
| Error | 81 | 54 | ||||
| Variable | Nitrate | Nitrate Reductase | Glutamine Synthetase | Total Chlorophyll | Phenolic Compounds | Ascorbate | Total Antioxidant Capacity |
|---|---|---|---|---|---|---|---|
| N = 81 (exception correlation between enzyme activities N = 106) | |||||||
| nitrate | --- | r = −0.0182 | 0.1669 | 0.1415 | −0.1155 | 0.0045 | 0.036 |
| p = 0.872 | 0.137 | 0.208 | 0.305 | 0.968 | 0.749 | ||
| nitrate reductase | --- | --- | r = 0.4669 | −0.0308 | 0.0251 | −0.116 | 0.0342 |
| p = 0.000 | 0.785 | 0.824 | 0.918 | 0.762 | |||
| glutamine synthetase | --- | --- | --- | r = 0.0473 | 0.0084 | −0.1358 | 0.0173 |
| p = 0.675 | 0.941 | 0.227 | 0.878 | ||||
| total chlorophyll | --- | --- | --- | --- | r = −0.3617 | −0.3065 | −0.3182 |
| p = 0.001 | 0.005 | 0.004 | |||||
| phenolic compounds | --- | --- | --- | --- | --- | r = 0.8317 | 0.8164 |
| p = 0.00 | 0.00 | ||||||
| ascorbate | --- | --- | --- | --- | --- | --- | r = 0.8981 |
| p = 0.00 | |||||||
| t. antioxidant capacity | --- | --- | --- | --- | --- | --- | --- |
| (C) Source of Variation | Phenolic Compounds | Ascorbate | Total Antioxidant Capacity | ||||||
|---|---|---|---|---|---|---|---|---|---|
| d.f. | F | p | d.f. | F | p | d.f. | F | p | |
| Species (S) | 8 | 170.24 | <0.0001 | 8 | 176.275 | < 0.0001 | 8 | 90.062 | <0.0001 |
| Cultivation (C) | 2 | 12.499 | <0.0001 | 2 | 35.823 | < 0.0001 | 2 | 15.414 | <0.0001 |
| S × C | 16 | 4.258 | <0.0001 | 16 | 6.456 | < 0.0001 | 16 | 4.993 | <0.0001 |
| Error | 54 | 54 | 54 | ||||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chadzinikolau, T.; Formela-Luboińska, M. Nitrogen Metabolism and Antioxidant Capacity of Selected Vegetables from Organic and Conventional Crops. Appl. Sci. 2023, 13, 11170. https://doi.org/10.3390/app132011170
Chadzinikolau T, Formela-Luboińska M. Nitrogen Metabolism and Antioxidant Capacity of Selected Vegetables from Organic and Conventional Crops. Applied Sciences. 2023; 13(20):11170. https://doi.org/10.3390/app132011170
Chicago/Turabian StyleChadzinikolau, Tamara, and Magda Formela-Luboińska. 2023. "Nitrogen Metabolism and Antioxidant Capacity of Selected Vegetables from Organic and Conventional Crops" Applied Sciences 13, no. 20: 11170. https://doi.org/10.3390/app132011170
APA StyleChadzinikolau, T., & Formela-Luboińska, M. (2023). Nitrogen Metabolism and Antioxidant Capacity of Selected Vegetables from Organic and Conventional Crops. Applied Sciences, 13(20), 11170. https://doi.org/10.3390/app132011170