


Effects of Fermented Onion on Gut Health in Dextran Sodium Sulfate (DSS)-Induced Inflammatory Bowel Disease (IBD) Rats

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Featured Application
Abstract
1. Introduction
2. Materials and Methods
2.1. Materials and Chemicals
2.2. Selection of a Lactic Acid Bacteria for the Fermentation of Onion
2.3. Preparation of Fermented Onion for Animal Experiment
2.4. Animal Studies
2.5. Experimental Design
2.6. Colonic Histomorphology
2.6.1. Hematoxylin and Eosin Staining
2.6.2. Alcian Blue and Nuclear Fast Red Staining
2.6.3. Mucus Layer Thickness Using Periodic Acid-Schiff (PAS) Staining
2.7. Oxidative Stress Parameters
2.7.1. Determination of Protein Carbonyl Content (PCOs)
2.7.2. Determination of Thiobarbituric Acid Reactive Substances (TBARS)
2.8. Immunoglobulin Enzyme-Linked Immunosorbent Assay (ELISA)
2.9. Protein Quantitative Assay
2.10. Western Blotting
2.11. Analysis of SCFAs in Colon Content Using GC
2.12. Statistics
3. Results
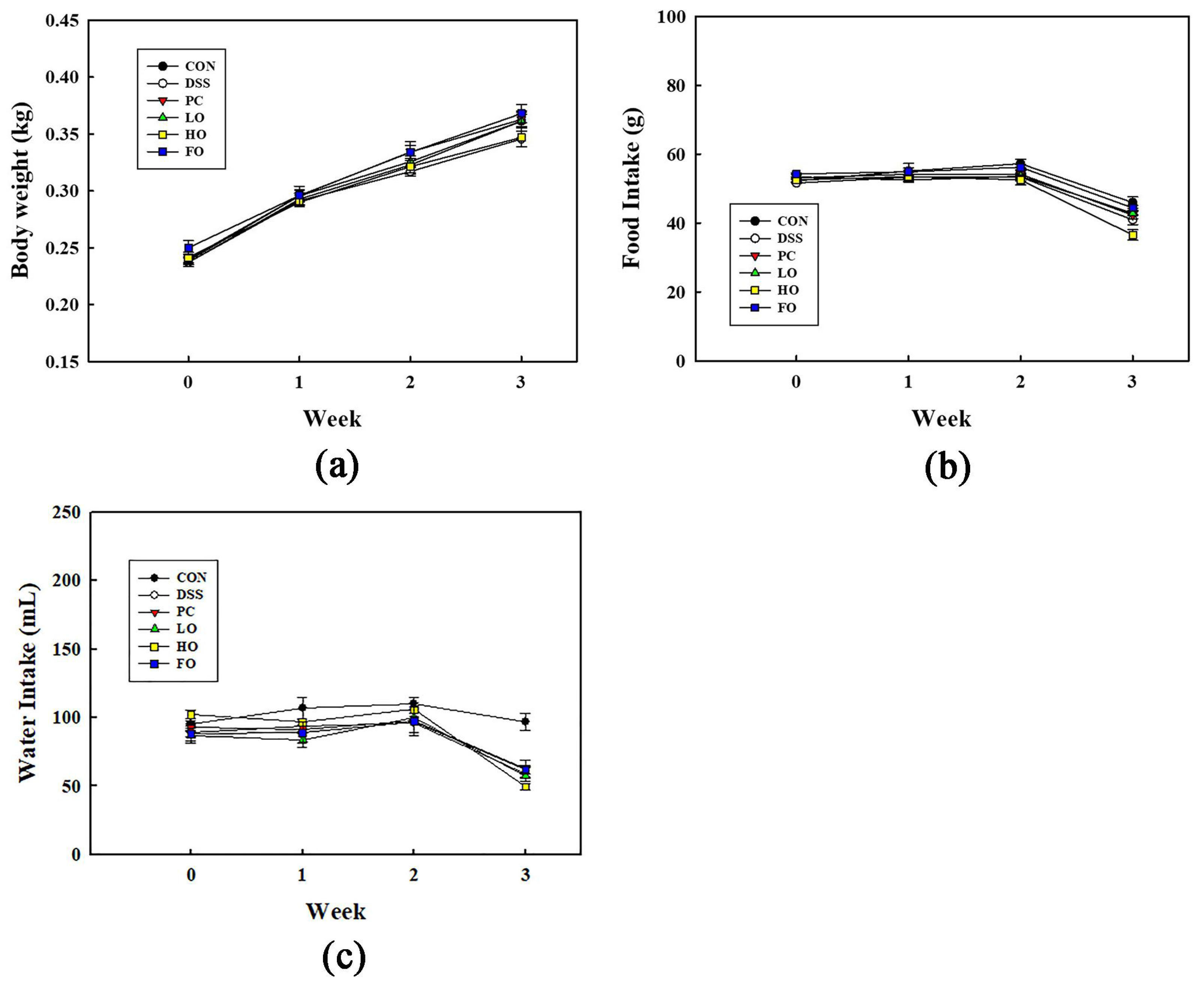
3.1. General Characteristics
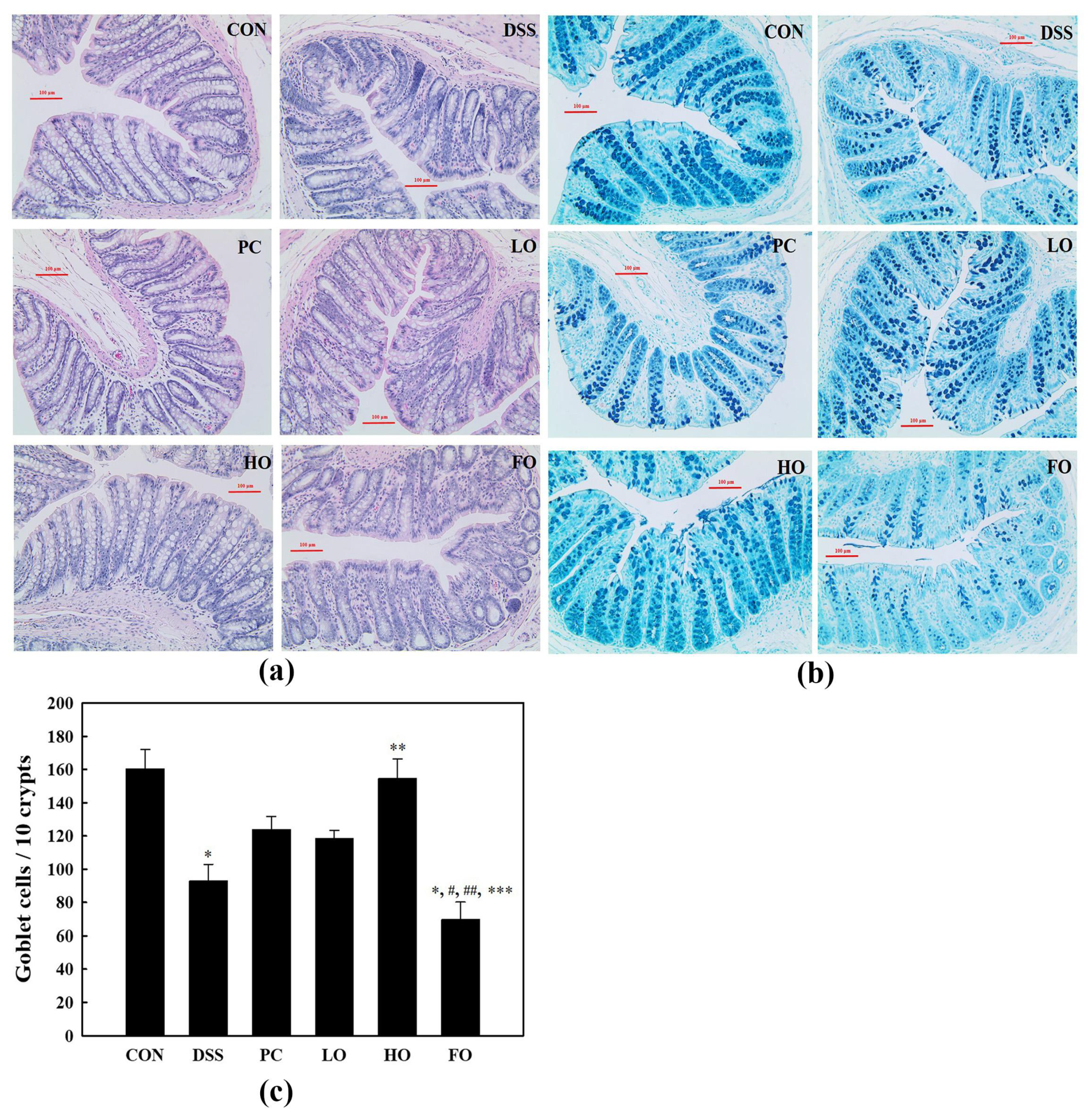
3.2. Colonic Histo-Morphology
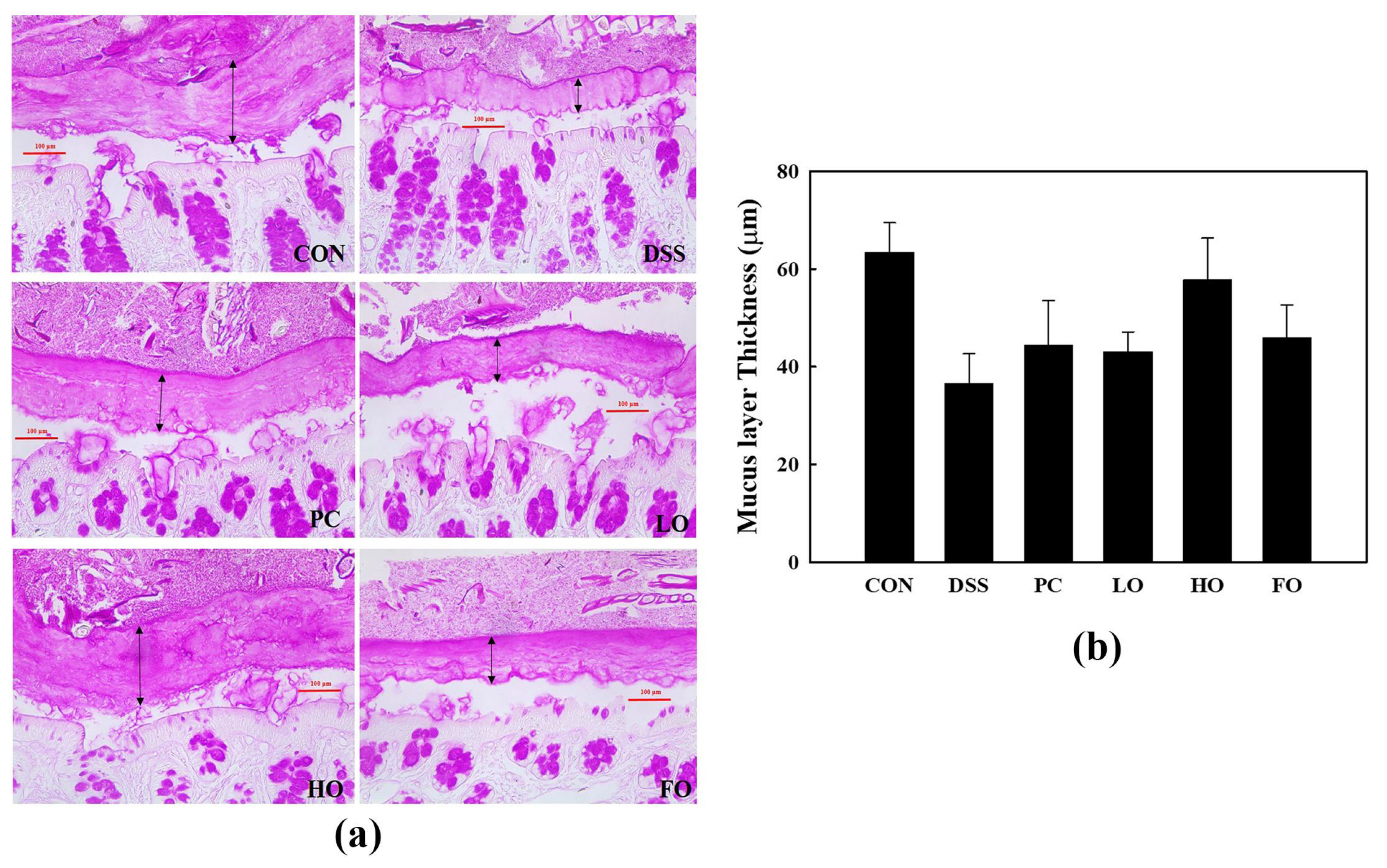
3.3. Inner Mucus Layer Thickness
3.4. Oxidative Stress Profiling
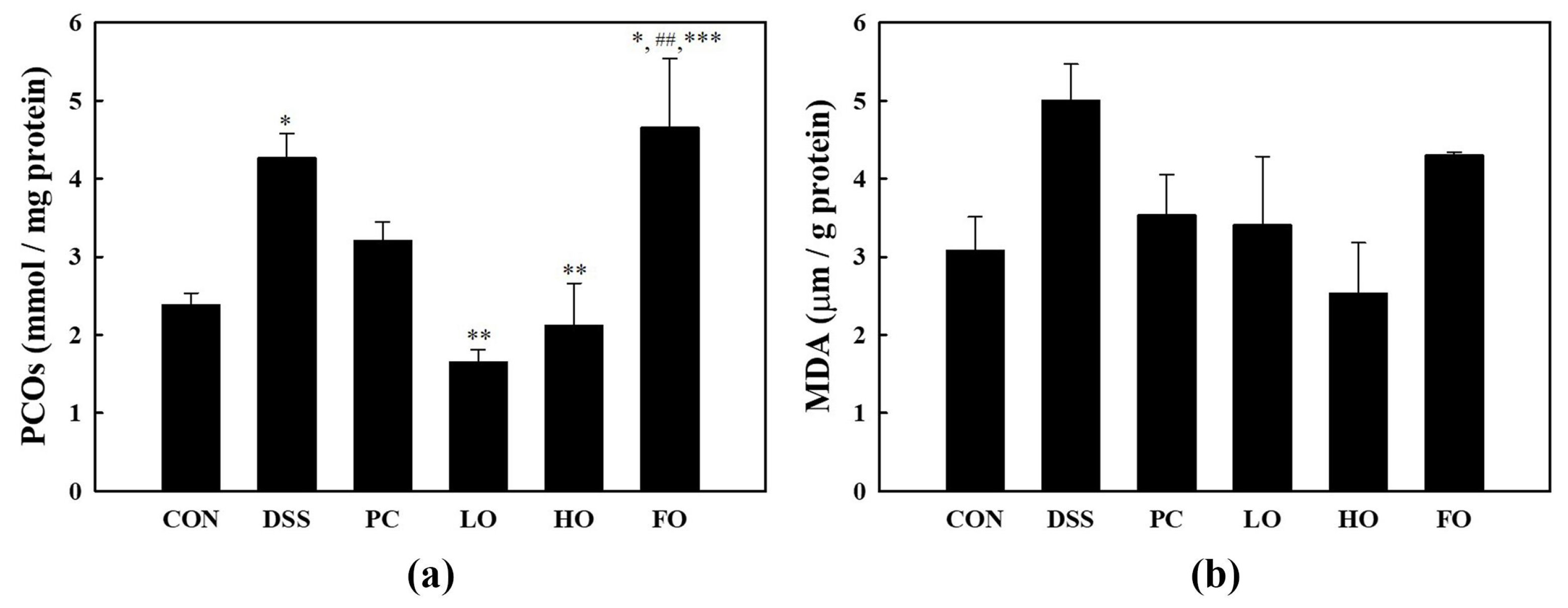
3.4.1. Protein Carbonyl (PCO) Content
3.4.2. Thiobarbituric Acid Reactive Substances (TBARS) Content

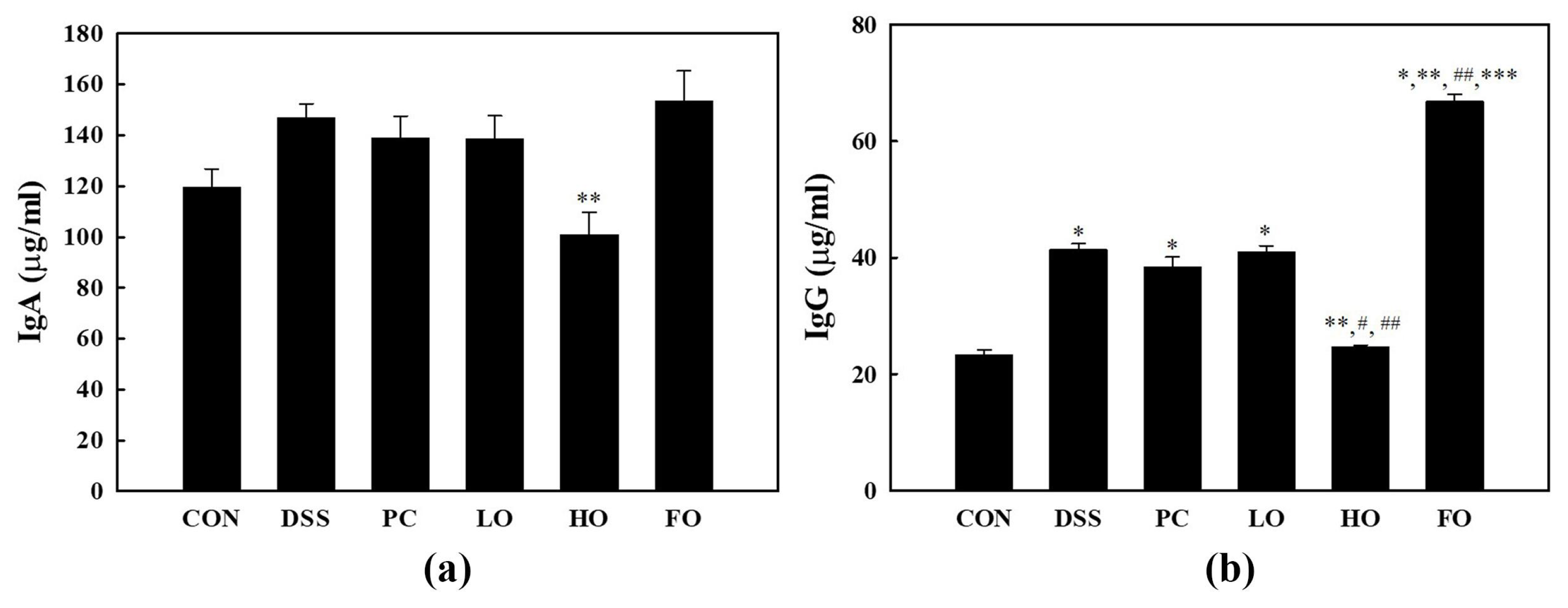
3.5. Effect on Host Immunity
3.6. Assessment of Inflammation
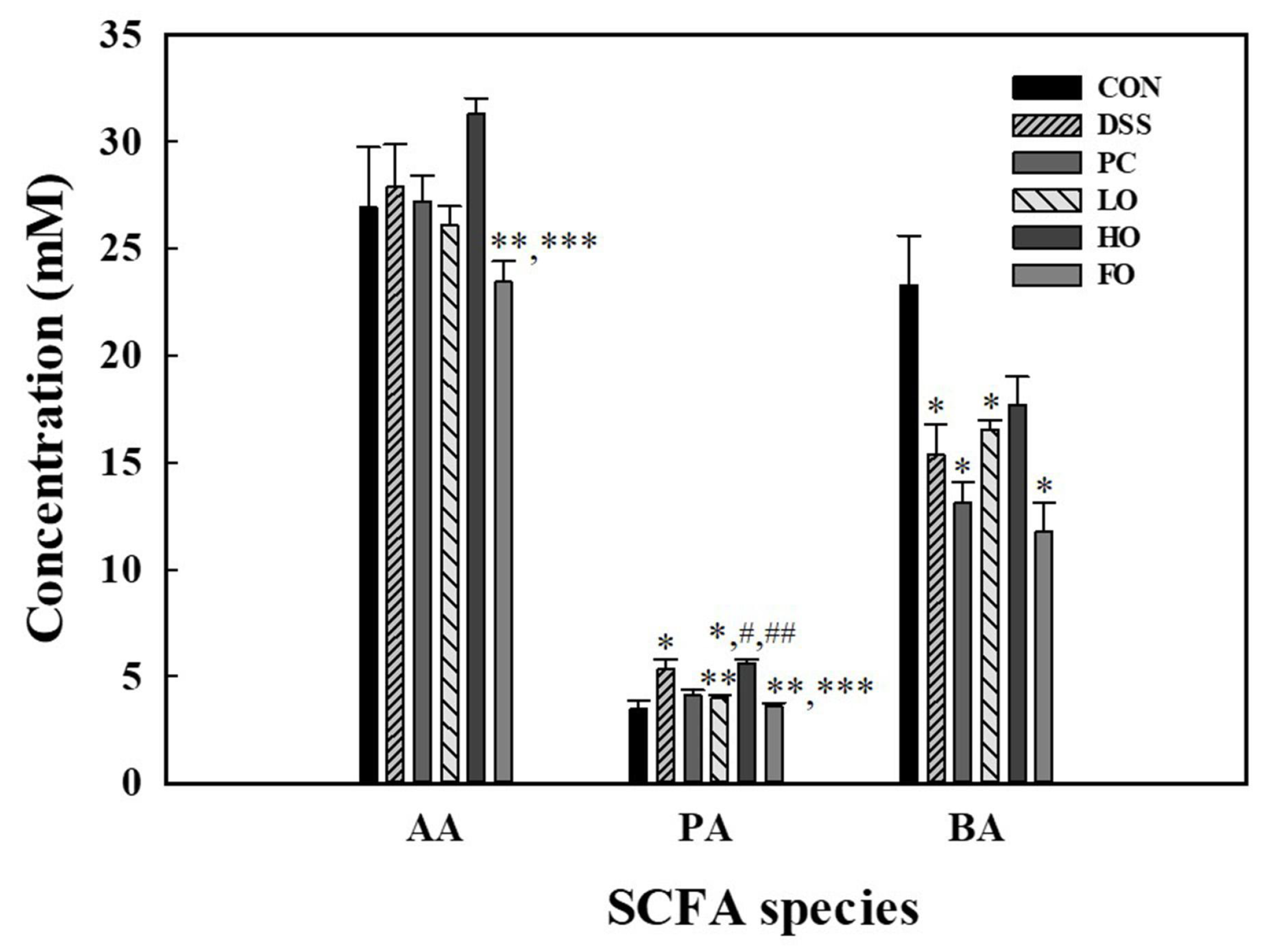
3.7. Short-Chain Fatty Acids (SCFAs) or Gut Metabolites
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Limdi, J.K. Dietary practices and inflammatory bowel disease. Indian J. Gastroenterol. 2018, 37, 284–292. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bhattacharyya, A.; Chattopadhyay, R.; Mitra, S.; Crowe, S.E. Oxidative stress: An essential factor in the pathogenesis of gastrointestinal mucosal diseases. Physiol. Rev. 2014, 94, 329–354. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Antoni, L.; Nuding, S.; Wehkamp, J.; Stange, E.F. Intestinal barrier in inflammatory bowel disease. World J. Gastroenterol. 2014, 20, 1165–1179. [Google Scholar] [CrossRef] [PubMed]
- McLean, L.P.; Cross, R.K. Adverse events in IBD: To stop or continue immune suppressant and biologic treatment. Expert Rev. Gastroenterol. Hepatol. 2014, 8, 223–240. [Google Scholar] [CrossRef] [Green Version]
- Andersen, V.; Olsen, A.; Carbonnel, F.; Tjønneland, A.; Vogel, U. Diet and risk of inflammatory bowel disease. Dig. Liver Dis. 2012, 44, 185–194. [Google Scholar] [CrossRef]
- Hong, Z.; Piao, M. Effect of Quercetin Monoglycosides on Oxidative Stress and Gut Microbiota Diversity in Mice with Dextran Sodium Sulphate-Induced Colitis. Biomed Res. Int. 2018, 2018, 8343052. [Google Scholar] [CrossRef]
- Yang, D.; Wang, T.; Long, M.; Li, P. Quercetin: Its Main Pharmacological Activity and Potential Application in Clinical Medicine. Oxid. Med. Cell. Longev. 2020, 2020, 8825387. [Google Scholar] [CrossRef]
- Panche, A.N.; Diwan, A.D.; Chandra, S.R. Flavonoids: An overview. J. Nutr. Sci. 2016, 5, 9–10. [Google Scholar] [CrossRef] [Green Version]
- Yang, E.J.; Kim, S.I.; Park, S.Y.; Bang, H.Y.; Jeong, J.H.; So, J.H.; Rhee, I.K.; Song, K.S. Fermentation enhances the in vitro antioxidative effect of onion (Allium cepa) via an increase in quercetin content. Food Chem. Toxicol. 2012, 50, 2042–2048. [Google Scholar] [CrossRef]
- Annunziata, G.; Arnone, A.; Ciampaglia, R.; Tenore, G.C.; Novellino, E. Fermentation of foods and beverages as a tool for increasing availability of bioactive compounds. focus on short-chain fatty acids. Foods 2020, 9, 999. [Google Scholar] [CrossRef]
- Wastyk, H.C.; Fragiadakis, G.K.; Perelman, D.; Dahan, D.; Merrill, B.D.; Yu, F.B.; Topf, M.; Gonzalez, C.G.; Van Treuren, W.; Han, S.; et al. Gut-microbiota-targeted diets modulate human immune status. Cell 2021, 184, 4137–4153.e14. [Google Scholar] [CrossRef]
- Hill, D.; Sugrue, I.; Tobin, C.; Hill, C.; Stanton, C.; Ross, R.P. The Lactobacillus casei group: History and health related applications. Front. Microbiol. 2018, 9, 5–6. [Google Scholar] [CrossRef] [Green Version]
- Horvat, I.B.; Gobin, I.; Kresović, A.; Hauser, G. How can probiotic improve irritable bowel syndrome symptoms. World J. Gastrointest. Surg. 2021, 13, 923. [Google Scholar] [CrossRef] [PubMed]
- Reznick, A.Z.; Packer, L. Oxidative damage to proteins: Spectrophotometric method for carbonyl assay. Methods Enzymol. 1994, 233, 357–363. [Google Scholar] [CrossRef] [PubMed]
- Zhao, G.; Nyman, M.; Jönsson, J.Å. Rapid determination of short-chain fatty acids in colonic contents and faeces of humans and rats by acidified water-extraction and direct-injection gas chromatography. Biomed. Chromatogr. 2006, 20, 674–682. [Google Scholar] [CrossRef]
- Chassaing, B.; Aitken, J.D.; Malleshappa, M.; Vijay-Kumar, M. DSS Protocol. Curr. Protoc. Immunol. 2015, 27, 1–19. [Google Scholar] [CrossRef]
- Tarakci, N.G.; Erdem, N.Z.; Dumen, E. Probiotic Foods Are Effective on Weight Loss, Biochemical Parameters, and Intestinal Microbiota in Wistar Albino Rats with Obese Microbiota. Int. J. Clin. Pract. 2022, 2022, 4569100. [Google Scholar] [CrossRef] [PubMed]
- Omotoso, G.O.; Muonagolu, J.N.; Enaibe, B.U. Histological Evaluation of the Jejunum and Ileum of Rats after Administration of High Dose Garlic Aqueous Extract. Int. J. Health Sci. 2012, 6, 135–140. [Google Scholar] [CrossRef] [PubMed]
- Celi, P.; Gabai, G. Oxidant/antioxidant balance in animal nutrition and health: The role of protein oxidation. Front. Vet. Sci. 2015, 2, 48. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abeyrathne, E.D.N.S.; Nam, K.; Ahn, D.U. Analytical methods for lipid oxidation and antioxidant capacity in food systems. Antioxidants 2021, 10, 1587. [Google Scholar] [CrossRef]
- Kumar, S.; Krishna Chaitanya, R.; Preedy, V.R. Assessment of Antioxidant Potential of Dietary Components; Elsevier Inc.: Amsterdam, The Netherlands, 2017; ISBN 9780128098547. [Google Scholar]
- Song, D.J.; Shen, J.; Chen, M.H.; Liu, Z.J.; Cao, Q.; Hu, P.J.; Gao, X.; Qian, J.M.; Wu, K.C.; Lai, L.J.; et al. Association of Serum Immunoglobulins Levels With Specific Disease Phenotypes of Crohn’s Disease: A Multicenter Analysis in China. Front. Med. 2021, 8, 621337. [Google Scholar] [CrossRef] [PubMed]
- Marrero, J.A.; Matkowskyj, K.A.; Yung, K.; Hecht, G.; Benya, R.V. Dextran sulfate sodium-induced murine colitis activates NF-κB and increases galanin-1 receptor expression. Am. J. Physiol. Gastrointest. Liver Physiol. 2000, 278, 797–804. [Google Scholar] [CrossRef] [PubMed]
- Silva, Y.P.; Bernardi, A.; Frozza, R.L. The Role of Short-Chain Fatty Acids From Gut Microbiota in Gut-Brain Communication. Front. Endocrinol. 2020, 11, 25. [Google Scholar] [CrossRef] [Green Version]
- Shepherd, S.J.; Lomer, M.C.E.; Gibson, P.R. Short-chain carbohydrates and functional gastrointestinal disorders. Am. J. Gastroenterol. 2013, 108, 707–717. [Google Scholar] [CrossRef] [PubMed]
- Septembre-Malaterre, A.; Remize, F.; Poucheret, P. Fruits and vegetables, as a source of nutritional compounds and phytochemicals: Changes in bioactive compounds during lactic fermentation. Food Res. Int. 2018, 104, 86–99. [Google Scholar] [CrossRef] [PubMed]
- Saeed, F.; Pasha, I.; Arshad, M.U.; Muhammad Anjum, F.; Hussain, S.; Rasheed, R.; Nasir, M.A.; Shafique, B. Physiological and Nutraceutical Perspectives of Fructan. Int. J. Food Prop. 2015, 18, 1895–1904. [Google Scholar] [CrossRef]
- Prince, A.C.; Myers, C.E.; Joyce, T.; Irving, P.; Lomer, M.; Whelan, K. Fermentable carbohydrate restriction (Low FODMAP Diet) in clinical practice improves functional gastrointestinal symptoms in patients with inflammatory bowel disease. Inflamm. Bowel Dis. 2016, 22, 1129–1136. [Google Scholar] [CrossRef] [Green Version]
- Kalantar-Zadeh, K.; Berean, K.J.; Burgell, R.E.; Muir, J.G.; Gibson, P.R. Intestinal gases: Influence on gut disorders and the role of dietary manipulations. Nat. Rev. Gastroenterol. Hepatol. 2019, 16, 733–747. [Google Scholar] [CrossRef]
- Barrett, J.S.; Gearry, R.B.; Muir, J.G.; Irving, P.M.; Rose, R.; Rosella, O.; Haines, M.L.; Shepherd, S.J.; Gibson, P.R. Dietary poorly absorbed, short-chain carbohydrates increase delivery of water and fermentable substrates to the proximal colon. Aliment. Pharmacol. Ther. 2010, 31, 874–882. [Google Scholar] [CrossRef]
- Ong, D.K.; Mitchell, S.B.; Barrett, J.S.; Shepherd, S.J.; Irving, P.M.; Biesiekierski, J.R.; Smith, S.; Gibson, P.R.; Muir, J.G. Manipulation of dietary short chain carbohydrates alters the pattern of gas production and genesis of symptoms in irritable bowel syndrome. J. Gastroenterol. Hepatol. 2010, 25, 1366–1373. [Google Scholar] [CrossRef]
- Murray, K.; Wilkinson-Smith, V.; Hoad, C.; Costigan, C.; Cox, E.; Lam, C.; Marciani, L.; Gowland, P.; Spiller, R.C. Differential effects of FODMAPs (Fermentable Oligo-, Di-, Mono-Saccharides and Polyols) on small and large intestinal contents in healthy subjects shown by MRI. Am. J. Gastroenterol. 2014, 109, 110–119. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paramithiotis, S.; Das, G.; Shin, H.S.; Patra, J.K. Fate of Bioactive Compounds during Lactic Acid Fermentation of Fruits and Vegetables. Foods 2022, 11, 733. [Google Scholar] [CrossRef] [PubMed]
- Huynh, N.T.; Van Camp, J.; Smagghe, G.; Raes, K. Improved release and metabolism of flavonoids by steered fermentation processes: A review. Int. J. Mol. Sci. 2014, 15, 19369–19388. [Google Scholar] [CrossRef] [PubMed]
- Car, B.D.; Eng, V.M.; Everds, N.E.; Bounous, D.I. Clinical Pathology of the Rat. In The Laboratory Rat; Academic Press: Cambridge, MA, USA, 2006; pp. 127–146. [Google Scholar] [CrossRef]
- Borton, M.A.; Sabag-Daigle, A.; Wu, J.; Solden, L.M.; O’Banion, B.S.; Daly, R.A.; Wolfe, R.A.; Gonzalez, J.F.; Wysocki, V.H.; Ahmer, B.M.M.; et al. Chemical and pathogen-induced inflammation disrupt the murine intestinal microbiome. Microbiome 2017, 5, 47. [Google Scholar] [CrossRef] [Green Version]
- Lin, R.; Chen, H.; Shu, W.; Sun, M.; Fang, L.; Shi, Y.; Pang, Z.; Wu, W.; Liu, Z. Clinical significance of soluble immunoglobulins A and G and their coated bacteria in feces of patients with inflammatory bowel disease. J. Transl. Med. 2018, 16, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.S.; Lee, K.J. Immunoglobulin G4-related immune responses to common food antigens in patients with ulcerative colitis and Crohn’s disease. Turkish J. Gastroenterol. 2019, 30, 408–414. [Google Scholar] [CrossRef]
- Wang, Z.; Zhu, M.; Luo, C.; Zhen, Y.; Mu, J.; Zhang, W.; Ouyang, Q.; Zhang, H. High level of IgG4 as a biomarker for a new subset of inflammatory bowel disease. Sci. Rep. 2018, 8, 10018. [Google Scholar] [CrossRef] [Green Version]
- Castro-Dopico, T.; Dennison, T.W.; Ferdinand, J.R.; Mathews, R.J.; Fleming, A.; Clift, D.; Stewart, B.J.; Jing, C.; Strongili, K.; Labzin, L.I.; et al. Anti-commensal IgG Drives Intestinal Inflammation and Type 17 Immunity in Ulcerative Colitis. Immunity 2019, 50, 1099–1114.e10. [Google Scholar] [CrossRef] [Green Version]
- Kazemi-Shirazi, L.; Gasche, C.H.; Natter, S.; Gangl, A.; Smolen, J.; Spitzauer, S.; Valent, P.; Kraft, D.; Valenta, R. IgA autoreactivity: A feature common to inflammatory bowel and connective tissue diseases. Clin. Exp. Immunol. 2002, 128, 102–109. [Google Scholar] [CrossRef]
- Hamer, H.M.; Jonkers, D.M.A.E.; Bast, A.; Vanhoutvin, S.A.L.W.; Fischer, M.A.J.G.; Kodde, A.; Troost, F.J.; Venema, K.; Brummer, R.J.M. Butyrate modulates oxidative stress in the colonic mucosa of healthy humans. Clin. Nutr. 2009, 28, 88–93. [Google Scholar] [CrossRef]
- Farag, M.A.; Tawfike, A.F.; Donia, M.S.; Ehrlich, A.; Wessjohann, L.A. Influence of pickling process on Allium cepa and citrus limon metabolome as determined via mass spectrometry-based metabolomics. Molecules 2019, 24, 928. [Google Scholar] [CrossRef] [PubMed]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sangpreecha, N.; Chanmuang, S.; Park, K.-H.; Sangar, M.; Sharma, D.; Song, D.; Park, Y.-J.; Sung, H.-M.; Promyo, K.; Ham, K.-S. Effects of Fermented Onion on Gut Health in Dextran Sodium Sulfate (DSS)-Induced Inflammatory Bowel Disease (IBD) Rats. Appl. Sci. 2023, 13, 1590. https://doi.org/10.3390/app13031590
Sangpreecha N, Chanmuang S, Park K-H, Sangar M, Sharma D, Song D, Park Y-J, Sung H-M, Promyo K, Ham K-S. Effects of Fermented Onion on Gut Health in Dextran Sodium Sulfate (DSS)-Induced Inflammatory Bowel Disease (IBD) Rats. Applied Sciences. 2023; 13(3):1590. https://doi.org/10.3390/app13031590
Chicago/Turabian StyleSangpreecha, Neeracha, Saoraya Chanmuang, Kyung-Hee Park, Madhuri Sangar, Divya Sharma, Doyoung Song, Yun-Ja Park, Hea-Mi Sung, Kitipong Promyo, and Kyung-Sik Ham. 2023. "Effects of Fermented Onion on Gut Health in Dextran Sodium Sulfate (DSS)-Induced Inflammatory Bowel Disease (IBD) Rats" Applied Sciences 13, no. 3: 1590. https://doi.org/10.3390/app13031590
APA StyleSangpreecha, N., Chanmuang, S., Park, K. -H., Sangar, M., Sharma, D., Song, D., Park, Y. -J., Sung, H. -M., Promyo, K., & Ham, K. -S. (2023). Effects of Fermented Onion on Gut Health in Dextran Sodium Sulfate (DSS)-Induced Inflammatory Bowel Disease (IBD) Rats. Applied Sciences, 13(3), 1590. https://doi.org/10.3390/app13031590