
Heterotrophic and Photoautotrophic Media Optimization Using Response Surface Methodology for the Novel Microalga Chlorococcum amblystomatis

 ,
, 
 ,
,  ,
,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Microalgal Strain and Growth Conditions
2.2. Growth Assessment
2.3. Experimental Design
2.4. Biochemical Composition
2.5. Statistical Analysis
3. Results and Discussion
3.1. Preliminary Screening Using Plackett–Burman Design
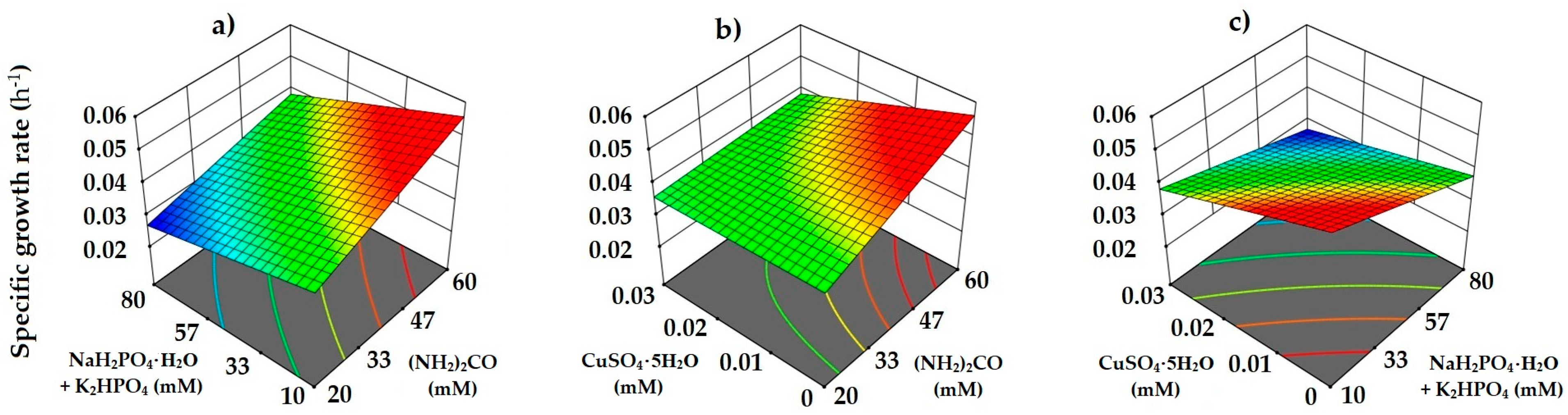
3.2. Heterotrophic Medium Optimization Using Box–Behnken Design
3.3. Validation of Heterotrophic Medium Optimization
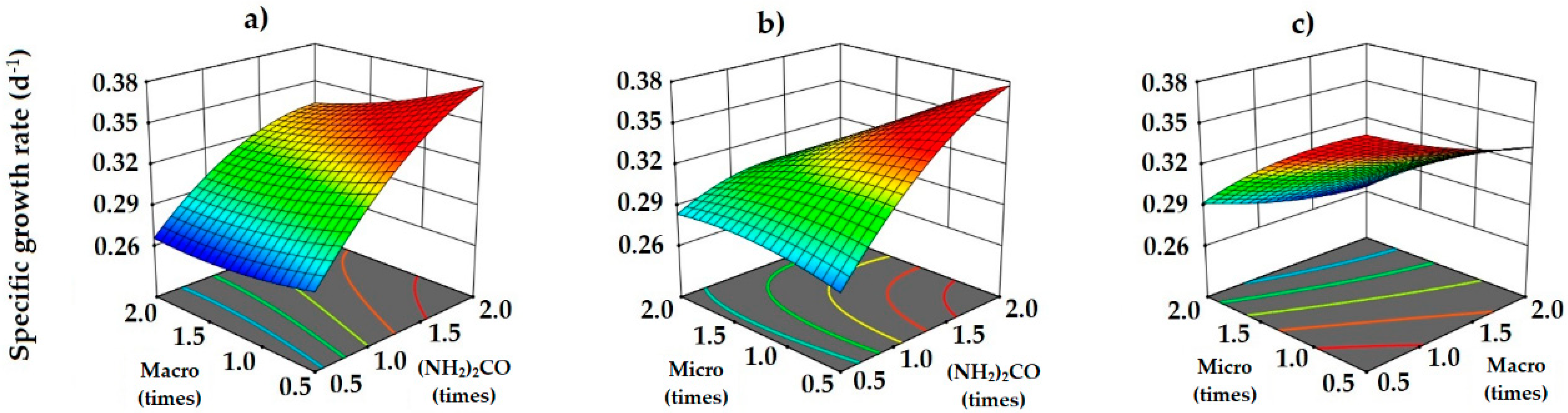
3.4. Photoautotrophic Medium Optimization Using Box–Behnken Design
3.5. Validation of Photoautotrophic Medium Optimization
3.6. Biochemical Composition
3.6.1. Proximate Composition
3.6.2. Fatty Acids Profile
3.6.3. Chlorophylls and Carotenoids Contents
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Pulz, O.; Gross, W. Valuable products from biotechnology of microalgae. Appl. Microbiol. Biotechnol. 2004, 65, 635–648. [Google Scholar] [CrossRef]
- Hlaing, S.A.A.; Sadiq, M.B.; Anal, A.K. Enhanced yield of Scenedesmus obliquus biomacromolecules through medium optimization and development of microalgae based functional chocolate. J. Food Sci. Technol. 2020, 57, 1090–1099. [Google Scholar] [CrossRef]
- Priyadarshani, I.; Rath, B. Commercial and industrial applications of micro algae—A review. J. Algal Biomass Util. 2012, 3, 89–100. [Google Scholar]
- Ejike, C.E.; Collins, S.A.; Balasuriya, N.; Swanson, A.K.; Mason, B.; Udenigwe, C.C. Prospects of microalgae proteins in producing peptide-based functional foods for promoting cardiovascular health. Trends Food Sci. Technol. 2017, 59, 30–36. [Google Scholar] [CrossRef]
- Caporgno, M.P.; Mathys, A. Trends in Microalgae Incorporation Into Innovative Food Products With Potential Health Benefits. Front. Nutr. 2018, 5, 58. [Google Scholar] [CrossRef] [PubMed]
- Rumin, J.; Junior, R.G.D.O.; Bérard, J.-B.; Picot, L. Improving Microalgae Research and Marketing in the European Atlantic Area: Analysis of Major Gaps and Barriers Limiting Sector Development. Mar. Drugs 2021, 19, 319. [Google Scholar] [CrossRef] [PubMed]
- de Medeiros, V.P.B.; da Costa, W.K.A.; da Silva, R.T.; Pimentel, T.C.; Magnani, M. Microalgae as source of functional ingredients in new-generation foods: Challenges, technological effects, biological activity, and regulatory issues. Crit. Rev. Food Sci. Nutr. 2021, 62, 4929–4950. [Google Scholar] [CrossRef] [PubMed]
- Spolaore, P.; Joannis-Cassan, C.; Duran, E.; Isambert, A. Commercial applications of microalgae. J. Biosci. Bioeng. 2006, 101, 87–96. [Google Scholar] [CrossRef]
- Barros, A.; Pereira, H.; Campos, J.; Marques, A.; Varela, J.; Silva, J. Heterotrophy as a tool to overcome the long and costly autotrophic scale-up process for large scale production of microalgae. Sci. Rep. 2019, 9, 13935. [Google Scholar] [CrossRef]
- Daneshvar, E.; Ok, Y.S.; Tavakoli, S.; Sarkar, B.; Shaheen, S.M.; Hong, H.; Luo, Y.; Rinklebe, J.; Song, H.; Bhatnagar, A. Insights into upstream processing of microalgae: A review. Bioresour. Technol. 2021, 329, 124870. [Google Scholar] [CrossRef]
- Hu, J.; Nagarajan, D.; Zhang, Q.; Chang, J.-S.; Lee, D.-J. Heterotrophic cultivation of microalgae for pigment production: A review. Biotechnol. Adv. 2018, 36, 54–67. [Google Scholar] [CrossRef]
- Bumbak, F.; Cook, S.; Zachleder, V.; Hauser, S.; Kovar, K. Best practices in heterotrophic high-cell-density microalgal processes: Achievements, potential and possible limitations. Appl. Microbiol. Biotechnol. 2011, 91, 31–46. [Google Scholar] [CrossRef]
- Perez-Garcia, O.; Escalante, F.M.E.; De-Bashan, L.E.; Bashan, Y. Heterotrophic cultures of microalgae: Metabolism and potential products. Water Res. 2011, 45, 11–36. [Google Scholar] [CrossRef] [PubMed]
- Chen, G.-Q.; Chen, F. Growing Phototrophic Cells without Light. Biotechnol. Lett. 2006, 28, 607–616. [Google Scholar] [CrossRef] [PubMed]
- Ogbonna, J.C.; Masui, H.; Tanaka, H. Sequential heterotrophic/autotrophic cultivation-An efficient method of producing Chlorella biomass for health food and animal feed. J. Appl. Phycol. 1997, 9, 359–366. [Google Scholar] [CrossRef]
- Dionisi, H.M.; Lozada, M.; Olivera, N.L. Bioprospection of marine microorganisms: Biotechnological applications and methods. Rev. Argent Microbiol. 2012, 44, 49–60. [Google Scholar] [CrossRef]
- Pandey, M.K.; Dasgupta, C.N.; Mishra, S.; Srivastava, M.; Gupta, V.K.; Suseela, M.R.; Ramteke, P.W. Bioprospecting microalgae from natural algal bloom for sustainable biomass and biodiesel production. Appl. Microbiol. Biotechnol. 2019, 103, 5447–5458. [Google Scholar] [CrossRef]
- Fábregas, J.; Domínguez, A.; Regueiro, M.; Maseda, A.; Otero, A. Optimization of culture medium for the continuous cultivation of the microalga Haematococcus pluvialis. Appl. Microbiol. Biotechnol. 2000, 53, 530–535. [Google Scholar] [CrossRef]
- Mandenius, C.-F.; Brundin, A. Review: Biocatalysts and bioreactor design Bioprocess Optimization Using Design-of-Experiments Methodology. Biotechnol. Prog. 2008, 24, 1191–1203. [Google Scholar] [CrossRef] [PubMed]
- Cheng, K.-C.; Demirci, A.; Catchmark, J.M. Enhanced pullulan production in a biofilm reactor by using response surface methodology. J. Ind. Microbiol. Biotechnol. 2010, 37, 587–594. [Google Scholar] [CrossRef]
- Sarabia, L.A.; Ortiz, M.C. Response Surface Methodology. Compr. Chemom. 2009, 1, 345–390. [Google Scholar] [CrossRef]
- Steinberg, D.M.; Bursztyn, D. Response Surface Methodology in Biotechnology. Qual. Eng. 2010, 22, 78–87. [Google Scholar] [CrossRef]
- Chen, J.; Lan, X.; Jia, R.; Hu, L.; Wang, Y. Response Surface Methodology (RSM) Mediated Optimization of Medium Components for Mycelial Growth and Metabolites Production of Streptomyces alfalfae XN-04. Microorganisms 2022, 10, 1854. [Google Scholar] [CrossRef] [PubMed]
- Chlorococcum Amblystomatis (F.D.Lambert ex N.Wille) N.Correia, J.Varela & Leonel Pereira: AlgaeBase. Available online: https://www.algaebase.org/search/species/detail/?species_id=175845 (accessed on 16 September 2022).
- Bligh, E.G.; Dyer, W.J. A rapid method of total lipid extraction and purification. Can. J. Biochem. Physiol. 1959, 37, 911–917. [Google Scholar] [CrossRef]
- Pereira, H.; Barreira, L.; Mozes, A.; Florindo, C.; Polo, C.; Duarte, C.V.; Custódio, L.; Varela, J. Microplate-based high throughput screening procedure for the isolation of lipid-rich marine microalgae. Biotechnol. Biofuels 2011, 4, 61. [Google Scholar] [CrossRef]
- Nunez, M.; Quigg, A. Changes in growth and composition of the marine microalgae phaeodactylum tricornutum and nannochloropsis salina in response to changing sodium bicarbonate concentrations. J. Appl. Phycol. 2016, 28, 2123–2138. [Google Scholar] [CrossRef]
- Lepage, G.; Roy, C.C. Improved recovery of fatty acid through direct transesterification without prior extraction or purification. J. Lipid Res. 1984, 25, 1391–1396. [Google Scholar] [CrossRef] [PubMed]
- Couso, I.; Vila, M.; Vigara, J.; Cordero, B.F.; Vargas, M.; Rodríguez, H.; León, R. Synthesis of carotenoids and regulation of the carotenoid biosynthesis pathway in response to high light stress in the unicellular microalga Chlamydomonas reinhardtii. Eur. J. Phycol. 2012, 47, 223–232. [Google Scholar] [CrossRef]
- Chew, K.W.; Yap, J.Y.; Show, P.L.; Suan, N.H.; Juan, J.C.; Ling, T.C.; Lee, D.-J.; Chang, J.-S. Microalgae biorefinery: High value products perspectives. Bioresour. Technol. 2017, 229, 53–62. [Google Scholar] [CrossRef]
- Guedes, A.C.; Malcata, F.X. Nutritional Value and Uses of Microalgae in Aquaculture. Aquaculture 2012, 10, 59–78. [Google Scholar] [CrossRef]
- Becker, E.W. Micro-algae as a source of protein. Biotechnol. Adv. 2007, 25, 207–210. [Google Scholar] [CrossRef]
- Khanra, S.; Mondal, M.; Halder, G.; Tiwari, O.; Gayen, K.; Bhowmick, T.K. Downstream processing of microalgae for pigments, protein and carbohydrate in industrial application: A review. Food Bioprod. Process. 2018, 110, 60–84. [Google Scholar] [CrossRef]
- Del Río, E.; García-Gómez, E.; Moreno, J.; Guerrero, M.G.; García-González, M. Microalgae for oil. Assessment of fatty acid productivity in continuous culture by two high-yield strains, Chlorococcum oleofaciens and Pseudokirchneriella subcapitata. Algal Res. 2017, 23, 37–42. [Google Scholar] [CrossRef]
- Correia, N.; Pereira, H.; Silva, J.T.; Santos, T.; Soares, M.; Sousa, C.B.; Schüler, L.M.; Costa, M.; Varela, J.; Pereira, L.; et al. Isolation, Identification and Biotechnological Applications of a Novel, Robust, Free-living Chlorococcum (Oophila) amblystomatis Strain Isolated from a Local Pond. Appl. Sci 2020, 10, 3040. [Google Scholar] [CrossRef]
- Pereira, H.; Barreira, L.; Figueiredo, F.; Custódio, L.; Vizetto-Duarte, C.; Polo, C.; Rešek, E.; Engelen, A.; Varela, J. Polyunsaturated fatty acids of marine macroalgae: Potential for nutritional and pharmaceutical applications. Mar. Drugs 2012, 10, 1920–1935. [Google Scholar] [CrossRef] [PubMed]
- Schmitz, G.; Ecker, J. The opposing effects of n-3 and n-6 fatty acids. Prog. Lipid Res. 2008, 47, 147–155. [Google Scholar] [CrossRef] [PubMed]
- Morales-Sánchez, D.; A Martinez-Rodriguez, O.; Martinez, A. Heterotrophic cultivation of microalgae: Production of metabolites of commercial interest. J. Chem. Technol. Biotechnol. 2017, 92, 925–936. [Google Scholar] [CrossRef]
- Balder, H.F.; Vogel, J.; Jansen, M.C.; Weijenberg, M.P.; van den Brandt, P.A.; Westenbrink, S.; Van Der Meer, R.; Goldbohm, R.A. Heme and chlorophyll intake and risk of colorectal cancer in the Netherlands cohort study. Cancer Epidemiol. Biomark. Prev. 2006, 15, 717–725. [Google Scholar] [CrossRef]
- Paiva, S.A.R.; Russell, R.M. Beta-carotene and other carotenoids as antioxidants. J. Am. Coll. Nutr. 1999, 18, 426–433. [Google Scholar] [CrossRef]
- Tang, G.; Russell, R.M. Carotenoids as Provitamin, A. Carotenoids 2009, 5, 149–172. [Google Scholar] [CrossRef]
- Chen, B.H.; Chuang, J.R.; Lin, J.H.; Chiu, C.P. Quantification of Provitamin A Compounds in ChInese Vegetables by High-Performance Liquid Chromatography. J. Food Prot. 1993, 56, 51–54. [Google Scholar] [CrossRef] [PubMed]
- Mostafa, S.S.M. Microalgal Biotechnology: Prospects and Applications. Plant Sci. 2012, 12, 276–314. [Google Scholar] [CrossRef]
- Fernández-Sevilla, J.M.; Fernández, F.A.; Grima, E.M. Biotechnological production of lutein and its applications. Appl. Microbiol. Biotechnol. 2010, 86, 27–40. [Google Scholar] [CrossRef] [PubMed]
- Del Campo, J.A.; García-González, M.; Guerrero, M.G. Outdoor cultivation of microalgae for carotenoid production: Current state and perspectives. Appl. Microbiol. Biotechnol. 2007, 74, 1163–1174. [Google Scholar] [CrossRef] [PubMed]
- Kotake-Nara, E.; Asai, A.; Nagao, A. Neoxanthin and fucoxanthin induce apoptosis in PC-3 human prostate cancer cells. Cancer Lett. 2005, 220, 75–84. [Google Scholar] [CrossRef] [PubMed]
- Market Research Report. Available online: https://www.fortunebusinessinsights.com/industry-reports/carotenoids-market-100180 (accessed on 19 December 2022).



{kind=link}
{kind=link}
| Factor | Low Level (−) | High Level (+) | Effect | t-Value | p-Value |
|---|---|---|---|---|---|
| Nitrogen source | (NH2)2CO | (NH4)2SO4 | −0.006 | −7.33 | 0.000 |
| (NH2)2CO (NH4)2SO4 | 20.00 mM | 60.00 mM | −0.002 | −2.24 | 0.067 |
| NaH2PO4·H2O + K2HPO4 * | 10.00 mM | 100.00 mM | −0.005 | −6.05 | 0.001 |
| CaCl2·2H2O | 1.00 mM | 5.00 mM | 0.000 | 0.45 | 0.667 |
| MgSO4·7H2O | 1.00 mM | 10.00 mM | −0.002 | −2.43 | 0.051 |
| FeSO4·7H2O | 0.05 mM | 0.50 mM | 0.001 | 1.69 | 0.142 |
| ZnSO4·7H2O | 0.50 mM | 2.50 mM | 0.000 | −0.08 | 0.935 |
| Cl2Co·6H2O | 0.00 mM | 0.04 mM | 0.000 | −0.52 | 0.621 |
| Na2MoO4·2H2O | 0.00 mM | 0.10 mM | 0.000 | −0.12 | 0.905 |
| MnSO4·H2O | 0.20 mM | 1.50 mM | 0.000 | 0.29 | 0.778 |
| NiCl2·6H2O | 0.00 mM | 0.02 mM | 0.001 | 1.37 | 0.221 |
| CuSO4·5H2O | 0.00 mM | 0.02 mM | −0.002 | −2.88 | 0.028 |
| H3BO3 | 0.50 mM | 1.50 mM | 0.000 | −0.44 | 0.673 |
| Vitamins Mix ** | 0.5 dose | 3 doses | 0.000 | 0.22 | 0.835 |
| Temperature | 26 °C | 30 °C | 0.000 | 0.55 | 0.601 |
| Run Order | (NH2)2CO mM | NaH2PO4·H2O + K2HPO4 mM | CuSO4·5H2O mM | µ Experimental h−1 | µ Predicted h−1 |
|---|---|---|---|---|---|
| 1 | 60 | 80 | 0.015 | 0.032 | 0.033 |
| 2 | 60 | 45 | 0.005 | 0.046 | 0.048 |
| 3 | 20 | 10 | 0.015 | 0.036 | 0.037 |
| 4 | 40 | 45 | 0.015 | 0.037 | 0.036 |
| 5 | 40 | 10 | 0.025 | 0.037 | 0.037 |
| 6 | 60 | 45 | 0.025 | 0.032 | 0.033 |
| 7 | 20 | 80 | 0.015 | 0.028 | 0.029 |
| 8 | 40 | 45 | 0.015 | 0.039 | 0.036 |
| 9 | 40 | 80 | 0.025 | 0.029 | 0.029 |
| 10 | 20 | 45 | 0.005 | 0.032 | 0.032 |
| 11 | 40 | 45 | 0.015 | 0.037 | 0.036 |
| 12 | 60 | 10 | 0.015 | 0.048 | 0.048 |
| 13 | 20 | 45 | 0.025 | 0.034 | 0.033 |
| 14 | 40 | 10 | 0.005 | 0.047 | 0.047 |
| 15 | 40 | 80 | 0.005 | 0.034 | 0.033 |
| Source | Sum of Squares | Degree of Freedom | Mean Square | Coefficient Estimate | F-Value | p-Value |
|---|---|---|---|---|---|---|
| Model | 0.0005 | 6 | 0.0001 | 65.25 | <0.0001 | |
| A-(NH2)2CO | 0.0001 | 1 | 0.0001 | 0.0034 | 76.66 | <0.0001 |
| B-NaH2PO4·H2O + K2HPO4 | 0.0003 | 1 | 0.0003 | −0.0064 | 186.76 | <0.0001 |
| C-CuSO4·5H2O | 0.0001 | 1 | 0.0001 | −0.0035 | 67.35 | <0.0001 |
| AB | 0.0000 | 1 | 0.0000 | −0.0010 | 8.40 | 0.0200 |
| AC | 0.0001 | 1 | 0.0001 | −0.0042 | 47.02 | 0.0001 |
| BC | 0.0000 | 1 | 0.0000 | 0.0013 | 5.33 | 0.0498 |
| Residual | 0.0000 | 8 | 0.0000 | |||
| Lack of Fit | 0.0000 | 6 | 0.0000 | 1.24 | 0.5107 | |
| Pure Error | 0.0000 | 2 | 0.0000 | |||
| Corr. Total | 0.0006 | 14 |
| Volumetric Productivity g·L−1·h−1 | Specific Growth Rate h−1 | Biomass Production g·L−1 | ||
|---|---|---|---|---|
| Global | Maximum | |||
| Basal medium | 0.12 ± 0.01 b | 0.29 ± 0.02 b | 0.03 ± 0.00 b | 7.71 ± 0.62 b |
| Optimized medium | 0.19 ± 0.01 a | 0.60 ± 0.06 a | 0.05 ± 0.00 a | 10.53 ± 0.59 a |
| % of increment | 67.4 | 109.7 | 44.9 | 36.6 |
| Run Order | [Urea] Times | [Other Macro] Times | [Micro] Times | µ Experimental d−1 | µ Predicted d−1 |
|---|---|---|---|---|---|
| 1 | 1.25 | 0.50 | 2.00 | 0.30 | 0.30 |
| 2 | 2.00 | 0.50 | 1.25 | 0.34 | 0.34 |
| 3 | 0.50 | 1.25 | 2.00 | 0.29 | 0.28 |
| 4 | 1.25 | 1.25 | 1.25 | 0.31 | 0.31 |
| 5 | 1.25 | 0.50 | 0.50 | 0.35 | 0.34 |
| 6 | 1.25 | 1.25 | 1.25 | 0.31 | 0.31 |
| 7 | 1.25 | 2.00 | 2.00 | 0.29 | 0.29 |
| 8 | 0.50 | 1.25 | 0.50 | 0.27 | 0.27 |
| 9 | 0.50 | 0.50 | 1.25 | 0.28 | 0.29 |
| 10 | 1.25 | 1.25 | 1.25 | 0.32 | 0.31 |
| 11 | 1.25 | 2.00 | 0.50 | 0.31 | 0.32 |
| 12 | 2.00 | 1.25 | 2.00 | 0.27 | 0.27 |
| 13 | 0.50 | 2.00 | 1.25 | 0.28 | 0.28 |
| 14 | 2.00 | 2.00 | 1.25 | 0.31 | 0.30 |
| 15 | 2.00 | 1.25 | 0.50 | 0.35 | 0.35 |
| Source | Sum of Squares | Degrees of Freedom | Mean Square | Coefficient Estimate | F-Value | p-Value |
|---|---|---|---|---|---|---|
| Model | 0.0093 | 9 | 0.0010 | 17.07 | 0.0030 | |
| A-Urea | 0.0029 | 1 | 0.0029 | 0.0190 | 47.53 | 0.0010 |
| B-Other Macronutrients | 0.0008 | 1 | 0.0008 | −0.0097 | 12.44 | 0.0168 |
| C-Micronutrients | 0.0019 | 1 | 0.0019 | −0.0155 | 31.46 | 0.0025 |
| AB | 0.0003 | 1 | 0.0003 | −0.0085 | 4.80 | 0.0799 |
| AC | 0.0022 | 1 | 0.0022 | −0.0237 | 36.82 | 0.0018 |
| BC | 0.0001 | 1 | 0.0001 | 0.0040 | 1.04 | 0.3552 |
| A2 | 0.0009 | 1 | 0.0009 | −0.0158 | 15.08 | 0.0116 |
| B2 | 0.0001 | 1 | 0.0001 | 0.0050 | 1.51 | 0.2743 |
| C2 | 0.0002 | 1 | 0.0002 | −0.0066 | 2.65 | 0.1645 |
| Residual | 0.0003 | 5 | 0.0001 | |||
| Lack of Fit | 0.0001 | 3 | 0.0000 | 0.39 | 0.7775 | |
| Pure Error | 0.0002 | 2 | 0.0001 | |||
| Corr. Total | 0.0096 | 14 |
| Volumetric Productivity g·L−1·d−1 | Specific Growth Rate d−1 | Biomass Production g·L−1 | ||
|---|---|---|---|---|
| Global | Maximum | |||
| Basal medium | 0.12 ± 0.00 b | 0.17 ± 0.01b | 0.23 ± 0.01 b | 1.36 ± 0.02 b |
| Optimized medium | 0.19 ± 0.01a | 0.27 ± 0.02 a | 0.35 ± 0.01 a | 1.91 ± 0.06 a |
| % of increment | 55.8 | 65.8 | 51.2 | 40.8 |
| Medium | Proteins (%) | Lipids (%) | Carbohydrates (%) | Ashes (%) | |
|---|---|---|---|---|---|
| Hetero. | Basal | 33.49 ± 0.38 d | 7.41 ± 1.89 c | 47.68 ± 2.19 a | 11.42 ± 0.07 c |
| Optimized | 61.49 ± 0.13 b | 8.88 ± 0.70 c | 14.11 ± 0.58 b | 15.52 ± 0.24 b | |
| Photoauto. | Basal | 56.67 ± 1.06 c | 19.74 ± 1.07 a | 4.26 ± 0.47 c | 19.39 ± 1.53 a |
| Optimized | 73.45 ± 1.91 a | 13.28 ± 0.45 b | 8.23 ± 0.02 c | 6.07 ± 0.07 d |
| FAME % | Heterotrophy | Photoautotrophy | ||
|---|---|---|---|---|
| Basal | Optimized | Basal | Optimized | |
| C14:0 | 0.88 ± 0.05 a | 0.66 ± 0.03 b | 0.66 ± 0.00 b | nd |
| C16:4n-3 | 14.09 ± 0.14 b | 18.63 ± 1.509 a | 15.07 ± 0.40 b | 14.88 ± 0.12 b |
| C16:3n-3 | nd | 1.10 ± 0.38 a | 1.31 ± 0.07 a | 1.14 ± 0.03 a |
| C16:2n-6 | 2.38 ± 0.08 b | 1.68 ± 0.03 c | 1.23 ± 0.10 d | 3.12 ± 0.23 a |
| C16:1 | 9.68 ± 0.33 a | 5.50 ± 0.33 b | 1.21 ± 0.19 c | 1.77 ± 0.94 c |
| C16:0 | 30.95 ± 0.38 a | 32.33 ± 1.51 a | 23.25 ± 0.16 b | 22.35 ± 0.76 b |
| C17:3 | 5.07 ± 0.22 a | 4.32 ± 0.20 b | 2.71 ± 0.10 c | 3.06 ± 0.18 c |
| C18:4n-3 | 2.56 ± 0.03 d | 5.03 ± 0.15 b | 6.03 ± 0.35 a | 3.49 ± 0.29 c |
| C18:3n-3 | nd | nd | 34.26 ± 0.20 a | 28.46 ± 0.38 b |
| C18:3n-6 | 8.10 ± 0.11 b | 10.30 ± 0.09 a | 4.47 ± 0.04 d | 5.27 ± 0.06 c |
| C18:2n-6 | 12.26 ± 0.62 a,b | 10.59 ± 0.56 b | 5.38 ± 0.19 c | 13.38 ± 0.79 a |
| C18:1 | 11.46 ± 0.06 a | 7.58 ± 3.71 a,b | 2.05 ± 0.02 b,c | 1.54 ± 0.05 c |
| C18:0 | 2.55 ± 0.53 a | 2.29 ± 0.14 a,b | 2.03 ± 0.11 a,b | 1.52 ± 0.19 b |
| Σ SFA | 34.39 ± 0.29 a | 35.28 ± 1.67 a | 26.27 ± 0.48 b | 23.87 ± 0.94 b |
| Σ MUFA | 21.14 ± 0.37 a | 13.08 ± 4.01 a | 3.27 ± 0.20 b | 3.31 ± 0.89 b |
| Σ PUFA | 44.47 ± 0.57 c | 51.64 ± 2.37 b | 70.46 ± 0.63 a | 72.82 ± 0.07 a |
| Σ n-3 | 16.66 ± 0.15 d | 24.76 ± 2.02 c | 56.68 ± 0.38 a | 47.26 ± 0.62 b |
| Σ n-6 | 22.74 ± 0.75 a | 22.57 ± 0.55 a | 11.07 ± 0.29 b | 21.78 ± 0.56 a |
| Σn-6/Σn-3 | 1.37 ± 0.06 a | 0.92 ± 0.05 b | 0.20 ± 0.00 d | 0.45 ± 0.02 c |
| PUFA/SFA | 1.29 ± 0.03 c | 1.46 ± 0.02 c | 2.68 ± 0.07 b | 3.06 ± 0.12 a |
| Pigments (mg·g−1) | Heterotrophy | Photoautotrophy | ||
|---|---|---|---|---|
| Basal | Optimized | Basal | Optimized | |
| Neoxanthin | 0.52 ± 0.05 c | 0.79 ± 0.06 c | 1.46 ± 0.12 b | 3.66 ± 0.33 a |
| Violaxanthin | 0.02 ± 0.00 c | 0.06 ± 0.01 c | 0.51 ± 0.03 b | 0.75 ± 0.06 a |
| Lutein | 1.23 ± 0.10 c | 1.60 ± 0.11 c | 4.32 ± 0.15 b | 5.27 ± 0.37 a |
| β-carotene | 0.81 ± 0.16 b | 4.15 ± 0.22 a | 5.37 ± 0.44 a | 5.84 ± 0.98 a |
| Chlorophyll a and b | 8.53 ± 0.35 c | 14.59 ± 0.74 b | 12.88 ± 0.61 b | 29.32 ± 0.39 a |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Correia, N.; Pereira, H.; Schulze, P.S.C.; Costa, M.M.; Santo, G.E.; Guerra, I.; Trovão, M.; Barros, A.; Cardoso, H.; Silva, J.L.; et al. Heterotrophic and Photoautotrophic Media Optimization Using Response Surface Methodology for the Novel Microalga Chlorococcum amblystomatis. Appl. Sci. 2023, 13, 2089. https://doi.org/10.3390/app13042089
Correia N, Pereira H, Schulze PSC, Costa MM, Santo GE, Guerra I, Trovão M, Barros A, Cardoso H, Silva JL, et al. Heterotrophic and Photoautotrophic Media Optimization Using Response Surface Methodology for the Novel Microalga Chlorococcum amblystomatis. Applied Sciences. 2023; 13(4):2089. https://doi.org/10.3390/app13042089
Chicago/Turabian StyleCorreia, Nádia, Hugo Pereira, Peter S. C. Schulze, Monya M. Costa, Gonçalo E. Santo, Inês Guerra, Mafalda Trovão, Ana Barros, Helena Cardoso, Joana L. Silva, and et al. 2023. "Heterotrophic and Photoautotrophic Media Optimization Using Response Surface Methodology for the Novel Microalga Chlorococcum amblystomatis" Applied Sciences 13, no. 4: 2089. https://doi.org/10.3390/app13042089
APA StyleCorreia, N., Pereira, H., Schulze, P. S. C., Costa, M. M., Santo, G. E., Guerra, I., Trovão, M., Barros, A., Cardoso, H., Silva, J. L., Gouveia, L., & Varela, J. (2023). Heterotrophic and Photoautotrophic Media Optimization Using Response Surface Methodology for the Novel Microalga Chlorococcum amblystomatis. Applied Sciences, 13(4), 2089. https://doi.org/10.3390/app13042089