


Virulence Bioassay of Entomopathogenic Fungi against Adults of Atta mexicana under Controlled Conditions


 and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Biological Material
2.2. Virulence Bioassay
2.3. Preparation of the Synthetic Diet
2.4. Evaluation of Insecticidal Activity
2.5. Statistical Analysis
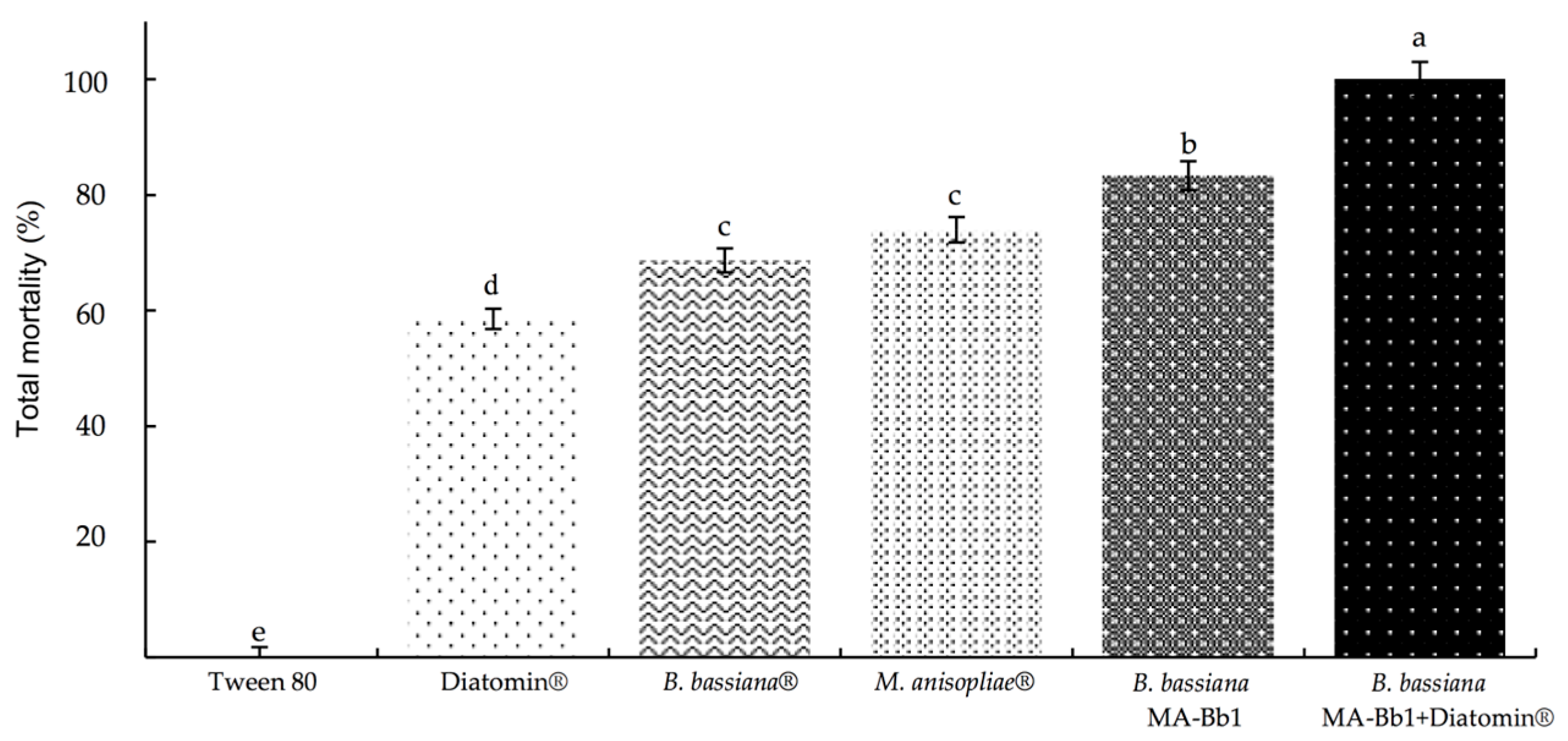
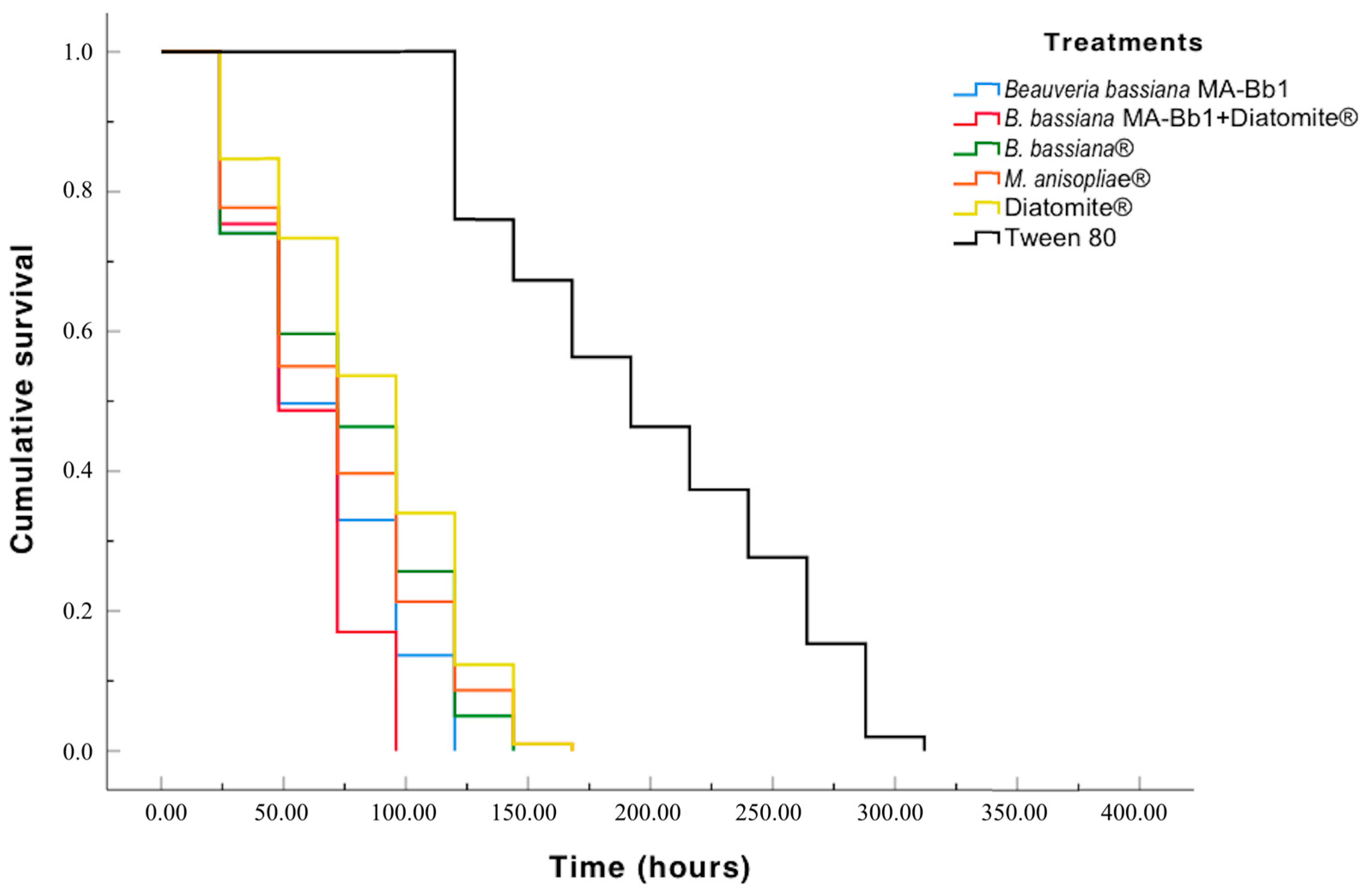
3. Results
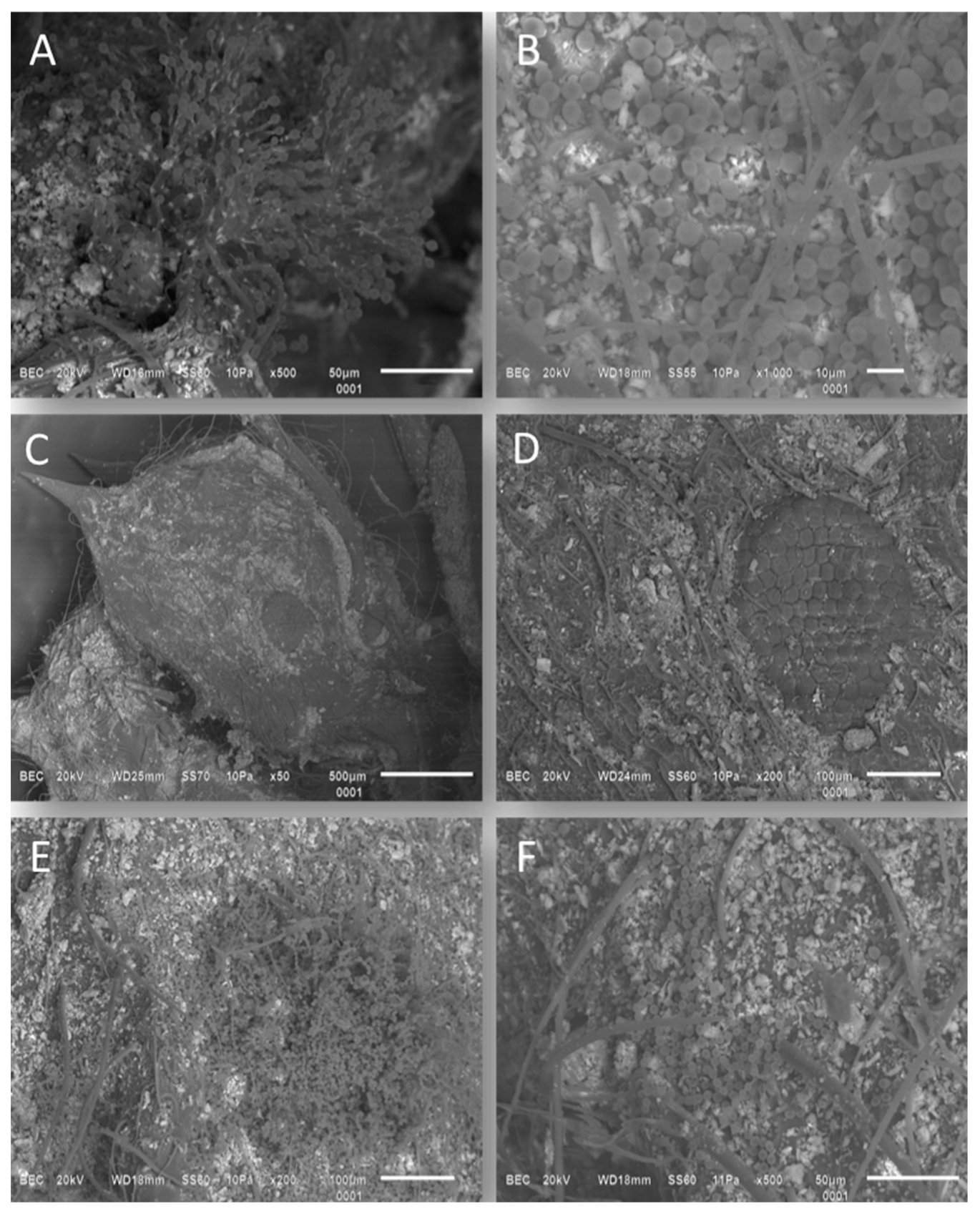
3.1. Viability of Different Biopreparation Formulations
3.2. Mortality of A. mexicana under Controlled Conditions
3.3. Lethal Time (LT)
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Hölldobler, B.; Wilson, E.O. The Leafcutter Ants. Civilization by Instinc; WW Norton & Company: London, UK, 2011; pp. 1–160. [Google Scholar]
- Lemus, Y.; Rodríguez, G.; Cuervo, R.; Vanegas, J.A.D.; Zuluaga, C.L. Feasibility of the Metarhizium anisopliae fungus to be used as a biological control of the “arriera” ant (Atta cephalotes). Rev. Guillermo Ockham 2008, 6, 91–98. [Google Scholar]
- Robledo- y Monterrubio, M.S.; Sánchez-Montes de Oca, G.; Arias-Velázquez, H.F.; Arias-Robledo, G.; Arias-Robledo, M.J. Control of leafhopper ant (Atta mexicana F. Smith) populations with Beauveria bassiana (Bals.) Vuill in rose laurel (Nerium oleander L.) crops in Yecapixtla, Morelos. Agro-Divulgación 2022, 2, 41–44. [Google Scholar] [CrossRef]
- Luna, J.M. “Arriera” ants, Atta spp. (Hymenoptera: Formicidae) from México. Dugesiana 1996, 3, 33–45. [Google Scholar]
- Vigueras, G.; Paredes-Hernández, D.; Revah, S.; Valenzuela, J.; Olivares-Hernández, R.; Le Borgne, S. Growth and enzymatic activity of Leucoagaricus gongylophorus, a mutualistic fungus isolated from the leaf-cutting ant Atta mexicana, on cellulose and lignocellulosic biomass. Lett Appl Microbiol. 2017, 65, 173–181. [Google Scholar] [CrossRef] [PubMed]
- Espinoza, C.; Zavala Izquierdo, I.; Couttolenc, A.; Landa-Cadena, G.; Valenzuela, J.; Trigos, Á. In vitro isolation and identification of Leucoagaricus gongylophorus from Atta mexicana (Hymenoptera: Formicidae) fungal garden. Rev. Mex. Mic. 2017, 46, 3–8. [Google Scholar]
- Aragón-García, A.; Pérez-Torres, B.C.; Calderón-García, G.; Castillo-Hernández, D.; Aragón-Sánchez, M.; Ramón, D.J. The tortilla of maize fungus complex for control leaf-cutter ant Atta mexicana (Smith 1858) (Hymenoptera: Formicidae). Entomol. Mex. 2016, 3, 153–158. [Google Scholar]
- Leal, I.R.; Wirth, R.; Tabarelli, M. The multiple impacts of leaf-cutting ants and their novel ecological role in humand-modified neotropical forests. Biotropica 2014, 46, 516–528. [Google Scholar] [CrossRef]
- Kooij, P.W.; Dentinger, B.M.; Donoso, D.A.; Shik, J.Z.; Gaya, E. Cryptic diversity in Colombian edible leaf-cutting ants (Hymenoptera: Formicidae). Insects 2018, 9, 191. [Google Scholar] [CrossRef] [PubMed]
- Pereira, C.C.; Sperandei, V.D.F.; Henriques, N.R.; Silva, Á.A.N.; Fernandes, G.W.; Cornelissen, T. Gallers as leaf rollers: Ecosystem engineering in a tropical system and its effects on arthropod biodiversity. Ecol. Entomol. 2021, 46, 470–481. [Google Scholar] [CrossRef]
- Vázquez-Jorge, M.D.L.Á.; Aragón-García, A.; Bibbins-Martínez, M.D.; Castillo-Hernández, D.; Nava-Galicia, S.B.; Pérez-Torres, B.C. Sphenarium purpurascens control with Beauveria bassiana and extracts amaranth (Amaranthus hypocondriacus L.). Rev. Mexicana Cienc. Agric. 2016, 7, 235–247. [Google Scholar]
- Forti, L.C.; Pretto, D.R.; Nagamoto, N.S.; Padovani, C.R.; Camargo, R.S.; Andrade, A.P.P. Dispersal of the delayed action insecticide sulfluramid in colonies of the leaf-cutting ant Atta sexdens rubropilosa (Hymenoptera: Formicidae). Sociobiology 2007, 50, 1149–1164. [Google Scholar]
- Teixeira, F.V.; de Melo Souza, T.R.; Castellani, M.A.; da Silva Camargo, R.; Forti, L.C.; de Paula, V.F.; Sabattini, J.A. Bioactivity of plant extracts against Atta sexdens (Hymenoptera: Formicidae) workers by ingestion. Int. J. Trop. Insect. Sci. 2023, 43, 1227–1236. [Google Scholar] [CrossRef]
- Qbal, M.; Gogi, M.D.; Atta, B.; Nisar, M.J.; Arif, M.J.; Javed, N. Assessment of pathogenicity of Beauveria bassiana, Metarhizium anisopliae, Verticillium lecanii and Bacillus thuringiensis var. kurstaki against Bactrocera cucurbitae Coquillett (Diptera: Tephritidae) via diet-bioassay technique under controlled conditions. Int. J. Trop. Insect. Sci. 2021, 41, 1129–1145. [Google Scholar] [CrossRef]
- Atta, B.; Rizwan, M.; Sabir, A.M.; Gogi, M.D.; Farooq, M.A.; Batta, Y. Efficacy of entomopathogenic fungi against brown planthopper Nilaparvata lugens (Stål) (Homoptera: Delphacidae) under controlled conditions. Gesunde Pflanz. 2020, 72, 101–112. [Google Scholar] [CrossRef]
- Farooq, M.A.; Atta, B.; Gogi, M.D.; Arif, M.J.; Arain, Q.A. Compatibility of entomopathogenic fungi and Azadirachta indica extract against the cotton pink bollworm, Pectinophora gossypiella (Saunders) (Lepidoptera: Gelechiidae) under controlled conditions. Egypt J Biol Pest Co. 2020, 30, 63. [Google Scholar] [CrossRef]
- Ali, S.; Huang, Z.; Ren, S. Production of cuticle degrading enzymes by Isaria fumosorosea and their evaluation as a biocontrol agent against diamondback moth. J Pest Sci. 2010, 83, 361–370. [Google Scholar] [CrossRef]
- Cerenius, L.; Trörnqvist, P.O.; Vey, A.; Johansson, M.W.; Söderhäll, K. The effect of the fungal toxin destruxin E on isolated crayfish haemocytes. J. Insect Physiol. 1990, 36, 785–789. [Google Scholar] [CrossRef]
- Polar, P.; Moore, D.; Kairo, M.; Ramsubhag, A. Topically applied myco acaricides for the control of cattle ticks: Overcoming the challenges. Exp. Appl. Acarol. 2008, 46, 119–148. [Google Scholar] [CrossRef]
- Castillo, L.D.; Sword, G.A. The endophytic fungal entomopathogens Beauveria bassiana and Purpureocillium lilacinum enhance the growth of cultivated cotton (Gossypium hirsutum) and negatively affect survival of the cotton bollworm (Helicoverpa zea). Biol. Control 2015, 89, 53–60. [Google Scholar]
- Fernandez-Daza, F.F.; Rodriguez-Roman, G.; Valencia-Rodriguez, M.; Gonzalez-Vargas, I.A.; Cardenas-Heano, H.; Pascol-Cereda, M.; Cuervo-Mulet, R.A. Spores of Beauveria bassiana and Trichoderma lignorum as a bioinsecticide for the control of Atta cephalotes. Biol. Res. 2019, 52, 51. [Google Scholar] [CrossRef]
- Romero-Arenas, O.; Amaro-Leal, L.J.; Rivera, A.; Parraguirre-Lezama, C.; Sánchez-Morales, P.; Villa-Ruano, N. Formulations of Beauveria bassiana MA-Bb1 and mesoporous materials for the biological control of Sphenarium purpurascens in maize crops from Puebla, Mexico. J. Asia Pac. Entomol. 2020, 23, 653–659. [Google Scholar] [CrossRef]
- Hu, X.; Xiao, G.; Zheng, P.; Shang, Y.; Su, Y.; Zhang, X.; Liu, X.; Zhan, S.; Leger, R.J.S.; Wang, C. Trajectory and genomic determinants of fungal-pathogen speciation and host adaptation. Proc. Natl. Acad. Sci. USA 2014, 111, 16796–16801. [Google Scholar] [CrossRef] [PubMed]
- Jiang, W.; Peng, Y.; Ye, J.; Wen, Y.; Liu, G.; Xie, J. Effects of the entomopathogenic fungus Metarhizium anisopliae on the mortality and immune response of Locusta migratoria. Insects 2020, 11, 36. [Google Scholar] [CrossRef] [PubMed]
- Amaro-Leal, L.J.; Romero-Arenas, O.; Lopez-Olguin, J.F.; Parraguirre-Lezama, C.; Sanchez-Morales, P.; Rivera Tapia, J.A. Mortality of Sphenarium purpurascens with Bio-preparations of Beauveria bassiana and Diatomin® in vitro. Southwest. Entomol. 2018, 43, 1029–1039. [Google Scholar] [CrossRef]
- Loureiro, E.D.S.; Monteiro, A.C. Pathogenicity of isolates of three entomopathogenic fungi against soldiers of Atta sexdens sexdens (Linnaeus, 1758) (Hymenoptera: Formicidae). Rev. Árvore. 2005, 29, 553–561. [Google Scholar] [CrossRef]
- Zanetti, R.; Cola Zanuncio, J.; Santos, J.C.; Da Silva, W.L.P.; Ribeiro, G.T.; Lemes, P.G. An overview of integrated management of leaf-cutting ants (Hymenoptera: Formicidae) in Brazilian forest plantations. Forests 2014, 5, 439–454. [Google Scholar] [CrossRef]
- Benítez, N. Microbiology Laboratory Manual. Dpto. de Biología; Universidad del Valle: Cali, Colombia, 2002. [Google Scholar]
- Zhang, X.; Lei, Z.; Reitz, S.R.; Wu, S.; Gao, Y. Laboratory and greenhouse evaluation of a granular formulation of Beauveria bassiana for control of western flower thrips, Frankliniella occidentalis. Insects 2019, 10, 58. [Google Scholar] [CrossRef] [PubMed]
- Cook, D.A.; Armitage, D.M. Efficacy of a diatomaceous earth against mite and insect populations in small beans of wheat under conditions of low temperatura and high humidity. Pest Manag. Sci. 2000, 56, 591–596. [Google Scholar] [CrossRef]
- Zeni, V.; Baliota, G.V.; Benelli, G.; Canale, A.; Athanassiou, C.G. Diatomaceous earth for arthropod pest control: Back to the future. Molecules 2021, 26, 7487. [Google Scholar] [CrossRef] [PubMed]
- Burgerjon, A. Pulvérisation et poudrage au laboratoire par des préparations pathogènes insecticides. Ann. Epiphyt. 1956, 7, 675–683. [Google Scholar]
- Bueno, F.C.; Godoy, M.P.; Leite, A.C.; Bueno, O.C.; Pagnocca, F.C.; Fernandez, J.B.; Hebling, M.J.A.; Bacci, M.; Vieira, P.C.; Silva, M.F.G.F. Toxicity of Cedrela fissilis to Atta sexdens rubropilosa (Hymenoptera: Formicidae) and its symbiotic fungus. Sociobiology 2005, 45, 389–399. [Google Scholar]
- Bueno, O.C.; Morini, M.S.; Pagnocca, F.C.; Hebling, M.J.; Silva, O.A. Survival of workers of Atta sexdens rubropilosa Forel (Hymenoptera: Formicidae) isolated from the ant-nest and fed artificial diets. An. Soc. Entomol. Brasil. 1997, 26, 107–113. [Google Scholar] [CrossRef]
- Castaño-Quintana, K.; Montoya-Lerma, J.; Giraldo-Echeverri, C. Toxicity of foliage extracts of Tithonia diversifolia (Asteraceae) on Atta cephalotes (Hymenoptera: Myrmicinae) workers. Ind. Crops Prod. 2013, 44, 391–395. [Google Scholar] [CrossRef]
- Abbott, W.S. A method of computing the effectiveness of an insecticide. J. Am. Mosquito Contr. 1987, 3, 301–302. [Google Scholar] [CrossRef]
- Püntener, W. Manual for Field Trials in Plant Protection, 2nd ed.; Documenta Ciba-Geigy: Basel, Switzerland, 1981; p. 205. [Google Scholar]
- Finney, D.J. Probit Analysis: A Statistical Treatment of the Sigmoid Response Curve, 7th ed.; Cambridge University Press: Cambridge, UK, 1972; p. 33. [Google Scholar]
- Rodríguez-Gómez, D.; Loera, O.; Saucedo-Castañeda, G.; Viniegra-Gonzalez, G. Substrate influence on physiology and virulence of Beauveria bassiana acting on larvae and adults of Tenebrio molitor. World J. Microbiol. Biotechnol. 2009, 25, 513–518. [Google Scholar] [CrossRef]
- Feng, M.G.; Poprawski, T.J.; Khachatourians, G.G. Production, formulation and application of the entomopathogenic fungus Beauveria bassiana for insect control: Current status. Biocontrol Sci. Technol. 1994, 4, 3–34. [Google Scholar] [CrossRef]
- De Faria, M.R.; Wraight, S.P. Mycoinsecticides and Mycoacaricides: A comprehensive list with worldwide coverage and international classification of formulation types. Biol. Control 2007, 43, 237–256. [Google Scholar] [CrossRef]
- Valero-Jiménez, C.A.; Faino, L.; Smit, S.; Zwaan, B.J.; van Kan, J.A. Comparative genomics of Beauveria bassiana: Uncovering signatures of virulence against mosquitoes. BMC Genom. 2016, 17, 986. [Google Scholar] [CrossRef] [PubMed]
- Gao, Y.P.; Luo, M.; Wang, X.Y.; He, X.Z.; Lu, W.; Zheng, X.L. Pathogenicity of Beauveria bassiana PfBb and Immune Responses of a Non-Target Host, Spodoptera frugiperda (Lepidoptera: Noctuidae). Insects 2022, 13, 914. [Google Scholar] [CrossRef] [PubMed]
- Peng, Y.J.; Wang, J.J.; Lin, H.Y.; Ding, J.L.; Feng, M.G.; Ying, S.H. HapX, an indispensable bZIP transcription factor for iron acquisition, regulates infection initiation by orchestrating conidial oleic acid homeostasis and cytomembrane functionality in mycopathogen Beauveria bassiana. Msystems 2020, 5, e00695-20. [Google Scholar] [CrossRef]
- López, E.; Orduz, S. Metarhizium anisopliae and Trichoderma viride for control of nests of the fungus-growing ant, Atta cephalotes. Biol. Control 2003, 27, 194–200. [Google Scholar] [CrossRef]
- Hughes, W.O.H.; Thomsen, L.; Eilenberg, J.; Boomsma, J.J. Diversity of entomopathogenic fungi near leaf-cutting ant nests in a Neotropical forest, with particular reference to Metarhizium anisopliae var. anisopliae. J. Invertebr. Pathol. 2004, 85, 46–53. [Google Scholar] [CrossRef] [PubMed]
- Rodrigues, A.; Bacci, M., Jr.; Mueller, U.G.; Ortiz, A.; Pagnocca, F.C. Microfungal “weeds” in the leafcutter ant symbiosis. Microb. Ecol. 2008, 56, 604–614. [Google Scholar] [CrossRef]
- Stefanelli, L.E.P.; Mota Filho, T.M.M.; Camargo, R.D.S.; Matos, C.A.O.D.; Forti, L.C. Effects of entomopathogenic fungi on individuals as well as groups of workers and immatures of Atta sexdens rubropilosa leaf-cutting ants. Insects 2020, 12, 10. [Google Scholar] [CrossRef] [PubMed]
- Diehl-Fleig, E.; Da Silva, M.E.; Pacheco, M.R.M. Tests of pathogenicity of entomopathogenic fungi Beauveria bassiana and Metarhizium anisopliae in Atta sexdens piriventris (Santschi, 1919) under different temperatures. Ciênc. Cult. 1988, 40, 1103–1105. [Google Scholar]
- Ribeiro, M.M.; Amaral, K.D.; Seide, V.E.; Souza, B.M.; Della Lucia, T.M.C.; Kasuya, M.C.M.; Souza, D.J. Diversity of fungi associated with Atta bisphaerica (Hymenoptera: Formicidae): The activity of Aspergillus ochraceus and Beauveria bassiana. Psyche J. Entomol. 2012, 2012, 389806. [Google Scholar] [CrossRef]
- Choe, A.; Chuman, T.; von Reuss, S.H.; Dossey, A.T.; Yim, J.J.; Ajredini, R.; Edison, A.S. Sex-specific mating pheromones in the nematode Panagrellus redivivus. Proc. Natl. Acad. Sci. USA 2012, 109, 20949–20954. [Google Scholar] [CrossRef] [PubMed]
- Pedrini, N.; Crespo, R.; Juarez, M.P. Biochemistry of insect epicuticle degradation by entomopathogenic fungi. Pharmacology 2007, 146, 124–137. [Google Scholar] [CrossRef]
- Song, M.H.; Yu, J.S.; Kim, S.; Lee, S.J.; Kim, J.C.; Nai, Y.S.; Kim, J.S. Downstream processing of Beauveria bassiana and Metarhizium anisopliae based fungal biopesticides against Riptortus pedestris: Solid culture and delivery of conidia. Biocontrol. Sci. Technol. 2019, 29, 514–532. [Google Scholar] [CrossRef]
- Akbar, W.; Lord, J.C.; Nechols, J.R.; Howard, R.W. Diatomaceous earth increases the efficacy of Beauveria bassiana against Tribolium castaneum larvae and increases conidia attachment. J. Econ. Entomol. 2004, 97, 273–280. [Google Scholar] [CrossRef]
- Vassilakos, T.N.; Athanassiou, C.G.; Kavallieratos, N.G.; Vayias, B.J. Influence of temperature on the insecticidal effect of Beauveria bassiana in combination with diatomaceous earth against Rhyzopertha dominica and Sitophilus oryzae on stored wheat. Biol. Control 2006, 38, 270–281. [Google Scholar] [CrossRef]
- Xu, J.; Zhang, K.; Cuthbertson, A.G.; Du, C.; Ali, S. Toxicity and biological effects of Beauveria brongniartii Fe0 nanoparticles against Spodoptera litura (Fabricius). Insects 2020, 11, 895. [Google Scholar] [CrossRef] [PubMed]
- Quesada-Moraga, E.; Ruiz-García, A.; Santiago-Álvarez, C. Laboratory evaluation of entomopathogenic fungi Beauveria bassiana and Metarhizium anisopliae against puparia and adults of Ceratitis capitata (Diptera: Tephritidae). J. Econ. Entomol. 2006, 99, 1955–1966. [Google Scholar] [CrossRef] [PubMed]
- Léger, R.J.S.; May, B.; Alle, L.L.; Frank, D.C.; Roberts, D.W. Genetic differences in allozymes and in formation of infection structures among isolates of the entomopathogenic fungus Metarhizium anisopliae. J. Invertebr. Pathol. 1992, 60, 89–101. [Google Scholar] [CrossRef]
- Leal, S.C.M.; Bertioli, D.J.; Butt, T.M.; Carder, J.H.; Burrows, P.R.; Peberdy, J.F. Amplification and restriction endonuclease digestion of the Pr1 gene for the detection and characterization of Metarhizium strains. Mycol. Res. 1997, 101, 257–265. [Google Scholar] [CrossRef]
- Léger, R.J.S.; Joshi, L.; Roberts, D. Ambient pH is a major determinant in the expression of cuticle-degrading enzymes and hydrophobin by Metarhizium anisopliae. Appl. Environ. Microbiol. 1998, 64, 709–713. [Google Scholar] [CrossRef] [PubMed]
- Currie, C.R.; Wong, B.; Stuart, A.E.; Schultz, T.R.; Rehner, S.A.; Mueller, U.G.; Straus, N.A. Ancient tripartite coevolution in the attine ant-microbe symbiosis. Science 2003, 299, 386–388. [Google Scholar] [CrossRef] [PubMed]
- Léger, R.J.S. Metarhizium anisopliae as a model for studying bioinsecticidal host pathogen interactions. In Novel Biotechnologies for Biocontrol Agent Enhancement and Management; Springer: New York, NY, USA; Heidelberg, Germany, 2007; pp. 179–204. [Google Scholar]
- Ekesi, S.; Egwurube, E.A.; Akpa, A.D.; Onu, I. Laboratory evaluation of the entomopathogenic fungus, Metarhizium anisopliae for the control of the groundnut bruchid, Caryedon serratus on groundnut. J. Stored Prod. Res. 2001, 37, 313–321. [Google Scholar] [CrossRef]
- Amala, U.; Jiji, T.; Naseema, A. Laboratory evaluation of local isolate of entomopathogenic fungus, Paecilomyces lilacinus Thom Samson (ITCC 6064) against adults of melon fruit fly, Bactrocera cucurbitae Coquillett. J. Trop. Agric. 2013, 51, 132–134. [Google Scholar]
- Akmal, M.; Freed, S.; Malik, M.N.; Gul, H.T. Efficacy of Beauveria bassiana (Deuteromycotina: Hypomycetes) against different aphid species under laboratory conditions. Pak. J. Zool. 2013, 45, 71–78. [Google Scholar]
- Ortiz-Urquiza, A.; Keyhani, N.O. Action on the Surface: Entomopathogenic Fungi versus the Insect Cuticle. Insects 2013, 4, 357–374. [Google Scholar] [CrossRef]
- Xiao, G.; Ying, S.H.; Zheng, P.; Wang, Z.L.; Zhang, S.; Xie, X.Q.; Shang, Y.; Leger, R.J.S.; Zhao, G.P.; Wang, C.; et al. Genomic perspectives on the evolution of fungal entomopathogenicity in Beauveria bassiana. Sci. Rep. 2012, 2, 483. [Google Scholar] [CrossRef] [PubMed]
- Vestergaard, S.; Gillespie, A.T.; Butt, T.M.; Schreiter, G.; Eilenberg, J. Pathogenicity of the hyphomycete fungi Verticillium lecanii and Metarhizium anisopliae to the western flower thrips, Frankliniella occidentalis. Biocontrol Sci. Technol. 1995, 5, 185–192. [Google Scholar] [CrossRef]
- Wang, Y.H.; Zheng, C.Y.; Wang, J.P. Virulence of Beauveria bassiana to Frankliniella occidentalis Adults and Scanning Electron Microscopic Observation on Its Infection Process. Chin. J. Biol. Control 2011, 3, 324–330. [Google Scholar]
- Thomas, M.B.; Wood, S.N.; Langewald, J.; Lomer, C.J. Persistence of Metarhizium flavoviride and consequences for biological control of grasshoppers and locusts. Pestic. Sci. 1997, 49, 47–55. [Google Scholar] [CrossRef]
- Butt, T.M.; Coates, C.J.; Dubovskiy, I.M.; Ratcliffe, N.A. Entomopathogenic fungi: New insights into host–pathogen interactions. Adv. Genet. 2016, 94, 307–364. [Google Scholar]
- Wang, H.H.; Liu, S.; Wang, S.Y.; Lei, Z.R. Research and development of wettable powder of Beauveria bassiana and its control and application to Frankliniella occidentalis. Chin. J. Biol. Control 2020, 36, 858–861. [Google Scholar]
- Pu, X.Y.; Feng, M.G.; Shi, C.H. Impact of three application methods on the field efficacy of a Beauveria bassiana-based mycoinsecticide against the false-eye leafhopper, Empoasca vitis (Homoptera: Cicadellidae) in the tea canopy. Crops Prot. 2005, 24, 167–175. [Google Scholar] [CrossRef]
- Aw, K.M.S.; Hue, S.M. Mode of Infection of Metarhizium spp. Fungus and Their Potential as Biological Control Agents. J. Fungi. 2017, 3, 30. [Google Scholar] [CrossRef] [PubMed]
- Schabel, H.G. Percutaneous infection of Hylobius pales by Metarrhizium anisopliae. J. Invertebr. Pathol. 1978, 31, 180–187. [Google Scholar] [CrossRef]
- Dornelas, A.S.P.; de Almeida Sarmento, R.; dos Santos, G.R.; Nascimento, M.O.; de Souza, D.J. Filamentous Fungi Associated With Atta sexdens (Linnaeus) and Atta laevigata (F. Smith) (Hymenoptera: Formicidae). EntomoBrasilis 2016, 9, 26–30. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatments | Conidia/mL | g | mL |
|---|---|---|---|
| Beauveria bassiana MA-Bb1 | 1 × 108 | - | 250 |
| B. bassiana MA-Bb1+ Diatomin® | 1 × 108 | - | 250 |
| a B. bassiana® | 1 × 108 | - | 250 |
| b M. anisopliae® | 1 × 108 | - | 250 |
| c Diatomin® (Diatomaceous earth) | - | 10 | 100 |
| Tween 80 (0.01%) | - | - | 1000 |
| Treatments | Conidia/mL * | Viability (% *) |
|---|---|---|
| (μ ± σ) | ||
| Beauveria bassiana MA-Bb1 | 9.38 × 107 b | 93.83 ± 0.14 b |
| B. bassiana MA-Bb1+ Diatomin® | 9.86 × 107 a | 98.63 ± 0.04 a |
| B. bassiana® | 7.31 × 107 c | 73.13 ± 0.31 c |
| M. anisopliae® | 6.86 × 107 d | 68.63 ± 0.02 d |
| Treatments | LT50 (Hours *) | CI of 95% | p Value | LT80 (Hours *) | CI of 95% | p Value |
|---|---|---|---|---|---|---|
| Beauveria bassiana MA-Bb1 | 48 a | 41.37–54.62 | 0.001 | 96 b | 94.7–97.2 | 0.001 |
| B. bassiana MA-Bb1+Diatomin® | 48 a | 43.34–52.65 | 0.001 | 72 a | 70.7–73.2 | 0.001 |
| * B. bassiana® | 72 c | 64.03–79.96 | 0.001 | 120 c | 118.7–121.2 | 0.001 |
| + M. anisopliae® | 72 c | 63.33–80.66 | 0.001 | 120 c | 118.7–121.2 | 0.001 |
| Diatomin® | 96 d | 89.45–102.54 | 0.001 | 120 c | 119.7–122.2 | 0.001 |
| Tween 80 (0.01%) | 192 e | 177.74–206.25 | 0.001 | 264 d | 262.7–265.2 | 0.001 |
| Variables | (1) | (2) | (3) | (4) | (5) | (6) | ||
|---|---|---|---|---|---|---|---|---|
| (1) | LT50 | Pearson correlation | 1 | 0.950 ** | 0.908 ** | −0.908 ** | 0.805 ** | 0.825 ** |
| Covariance | 2898 | 1197 | −1197 | 6801 | 69 | |||
| (2) | LT80 | Pearson correlation | 1 | 0.953 ** | −0.953 ** | 0.828 ** | 0.782 ** | |
| Covariance | 1650 | −1650 | 9201 | 87 | ||||
| (3) | Mortality | Pearson correlation | 1 | −1.000 ** | 0.865 ** | 0.807 ** | ||
| Covariance | −747 | 4149 | 38 | |||||
| (4) | Survival | Pearson correlation | 1 | −0.865 ** | −0.807 ** | |||
| Covariance | −4149 | −38 | ||||||
| (5) | Time of mycosis manifestation | Pearson correlation | 1 | 0.956 ** | ||||
| Covariance | 294 | |||||||
| (6) | Treatment | Pearson correlation | 1 | |||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Leal, L.J.A.; Huerta de la Peña, A.; Ocampo Fletes, I.; Antonio López, P.; Villa-Ruano, N.; Romero-Arenas, O. Virulence Bioassay of Entomopathogenic Fungi against Adults of Atta mexicana under Controlled Conditions. Appl. Sci. 2024, 14, 3039. https://doi.org/10.3390/app14073039
Leal LJA, Huerta de la Peña A, Ocampo Fletes I, Antonio López P, Villa-Ruano N, Romero-Arenas O. Virulence Bioassay of Entomopathogenic Fungi against Adults of Atta mexicana under Controlled Conditions. Applied Sciences. 2024; 14(7):3039. https://doi.org/10.3390/app14073039
Chicago/Turabian StyleLeal, Luis J. Amaro, Arturo Huerta de la Peña, Ignacio Ocampo Fletes, Pedro Antonio López, Nemesio Villa-Ruano, and Omar Romero-Arenas. 2024. "Virulence Bioassay of Entomopathogenic Fungi against Adults of Atta mexicana under Controlled Conditions" Applied Sciences 14, no. 7: 3039. https://doi.org/10.3390/app14073039
APA StyleLeal, L. J. A., Huerta de la Peña, A., Ocampo Fletes, I., Antonio López, P., Villa-Ruano, N., & Romero-Arenas, O. (2024). Virulence Bioassay of Entomopathogenic Fungi against Adults of Atta mexicana under Controlled Conditions. Applied Sciences, 14(7), 3039. https://doi.org/10.3390/app14073039